
Abstract
Introduction
We and others have demonstrated that proinflammatory cytokine (PIC) production and several renin–angiotensin system (RAS) components are increased in the hypertensive kidney (18, 23, 40). Reactive oxygen species (ROS) production is also increased in the renal tissue of the SHR. Further, PICs and RAS components have been found to increase ROS production (9, 36, 38), which in turn can activate various intracellular signaling pathways, including that of the transcription factor nuclear factor-kappa B (NF-κB) (18). Activation of NF-κB induces transcription of PIC genes, leading to further increases in ROS production and fostering a cyclic positive feedback mechanism, thereby accelerating the progression of hypertension and its associated renal changes.
Innovation
Exercise is currently recommended as an adjunct to many pharmacological therapies for several chronic diseases, and has been shown to benefit end organs, including the kidney, in hypertension. However, the mechanistic basis for the positive effects of exercise on the kidney has not yet been elucidated. This study is the first to provide mechanistic evidence that exercise training can delay the development of hypertension and resulted in structural and functional improvements in the kidneys of hypertensive rats by decreasing oxidative stress, inflammation, and renin–angiotensin system activation in the kidney itself. Here, it was found that the early initiation of ExT in a rat model of hypertension delayed the development of hypertension by preventing oxidative stress, preserving antioxidant status, and decreasing inflammation. These benefits of ExT are likely responsible for the preservation of renal hemodynamics and structure in this model. Importantly, these findings on hydralazine-treated SHR animals suggest that exercise-induced benefits in hypertensive kidneys, at least, in part, are pressure independent. Although, a direct cause-effect relationship between various parameters studied could not be established, this study is clinically significant in that it demonstrates that exercise may be a viable nonpharmacological approach to delay hypertension and preserve renal function as well as renal structure in patients that are predisposed to hypertension.
Several previous studies have investigated the effects of exercise on hypertension and kidney diseases; however, most of the studies were performed on patients or animals with established disease or the exercise was combined with other interventions (7, 8, 14, 21, 47). However, the effects of ExT on delaying or preventing the progression of hypertension-induced renal injury have not yet been elucidated. The mechanisms by which chronic ExT may affect renal function are unknown, although various mechanisms have been proposed, which include a direct lowering of blood pressure, leading to reduced peripheral vascular resistance (39) and reductions in oxidative stress (21). Here, we hypothesized that chronic ExT would preserve renal structure and function by modulating oxidative stress and inflammation in the SHR model of hypertension.
Results
Body weights
There were no significant differences in body weights between WKY and SHR groups at baseline (Table 1). However, body weights were significantly lower in both WKY and SHR exercise groups at study end when compared to their sedentary counterparts (Table 2).
Rats were obtained at 7 weeks of age.
Data are presented as mean±SEM.
BW, body weight; g, grams; GFR, glomerular filtration rate; KW, kidney weight; MAP, mean arterial pressure; mmHg, millimeters of mercury; RBF, renal blood flow; RVR, renal vascular resistance; SBP, systolic blood pressure.
Data were obtained at conclusion of study. Data are presented as mean±SEM.
p<0.05 vs. WKY-S; * p<0.05 vs. SHR-S; † p<0.05 vs. SHR-E.
BW, body weight; g, grams; GFR, glomerular filtration rate; KW, kidney weight; MAP, mean arterial pressure; mmHg, millimeters of mercury; RBF, renal blood flow; RVR, renal vascular resistance; SBP, systolic blood pressure.
Assessment of training efficacy
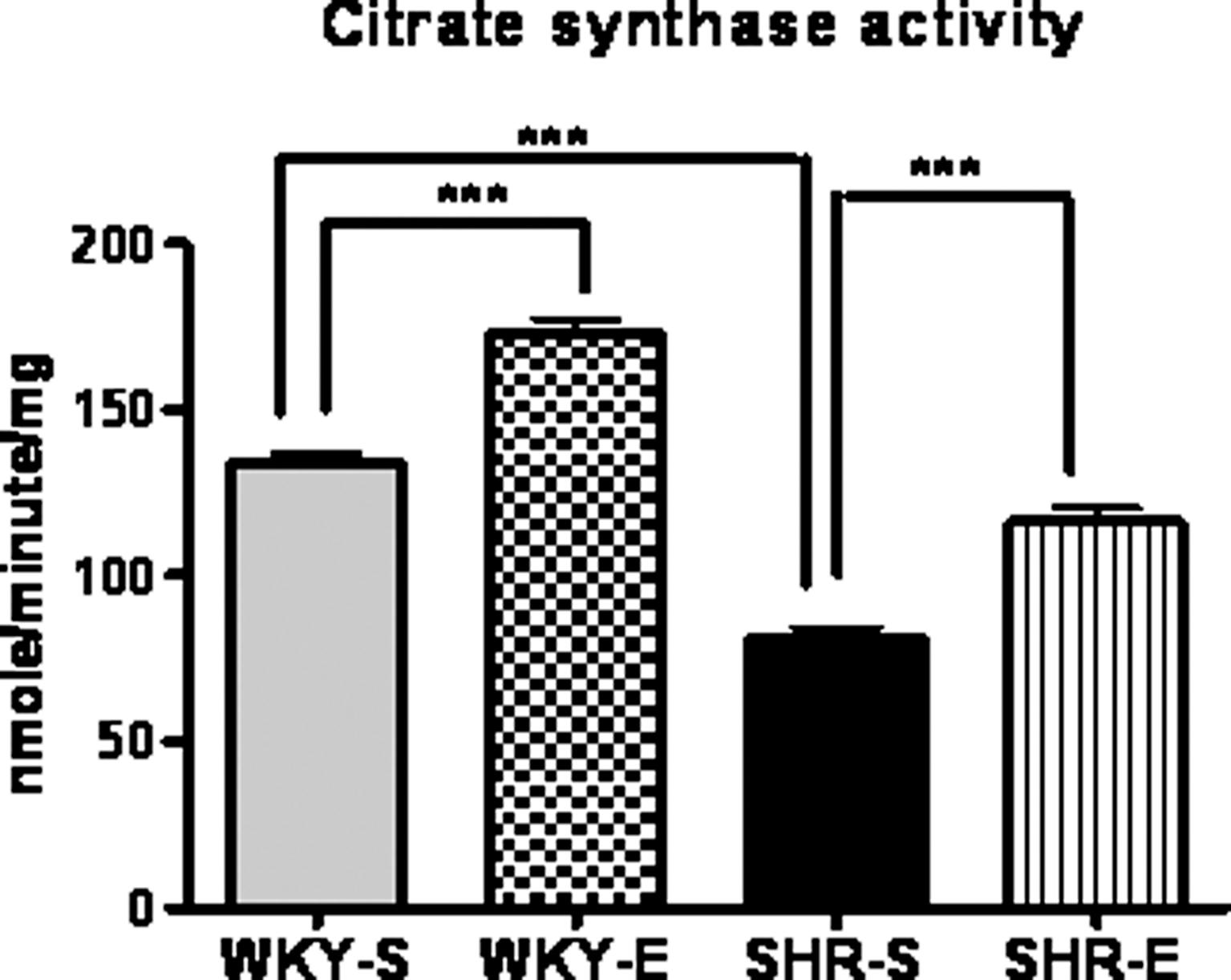
Citrate synthase (CS) activity in soleus muscle was used as a marker of training efficacy. After the period of 16 weeks of exercise, the CS activity was significantly higher in SHR and WKY rats compared with their sedentary control groups, indicating the efficacy of the exercise protocol (Fig. 1). CS activity was higher in WKY rats compared with SHR both in the exercise and the sedentary group.
ExT preserves renal hemodynamics in SHR
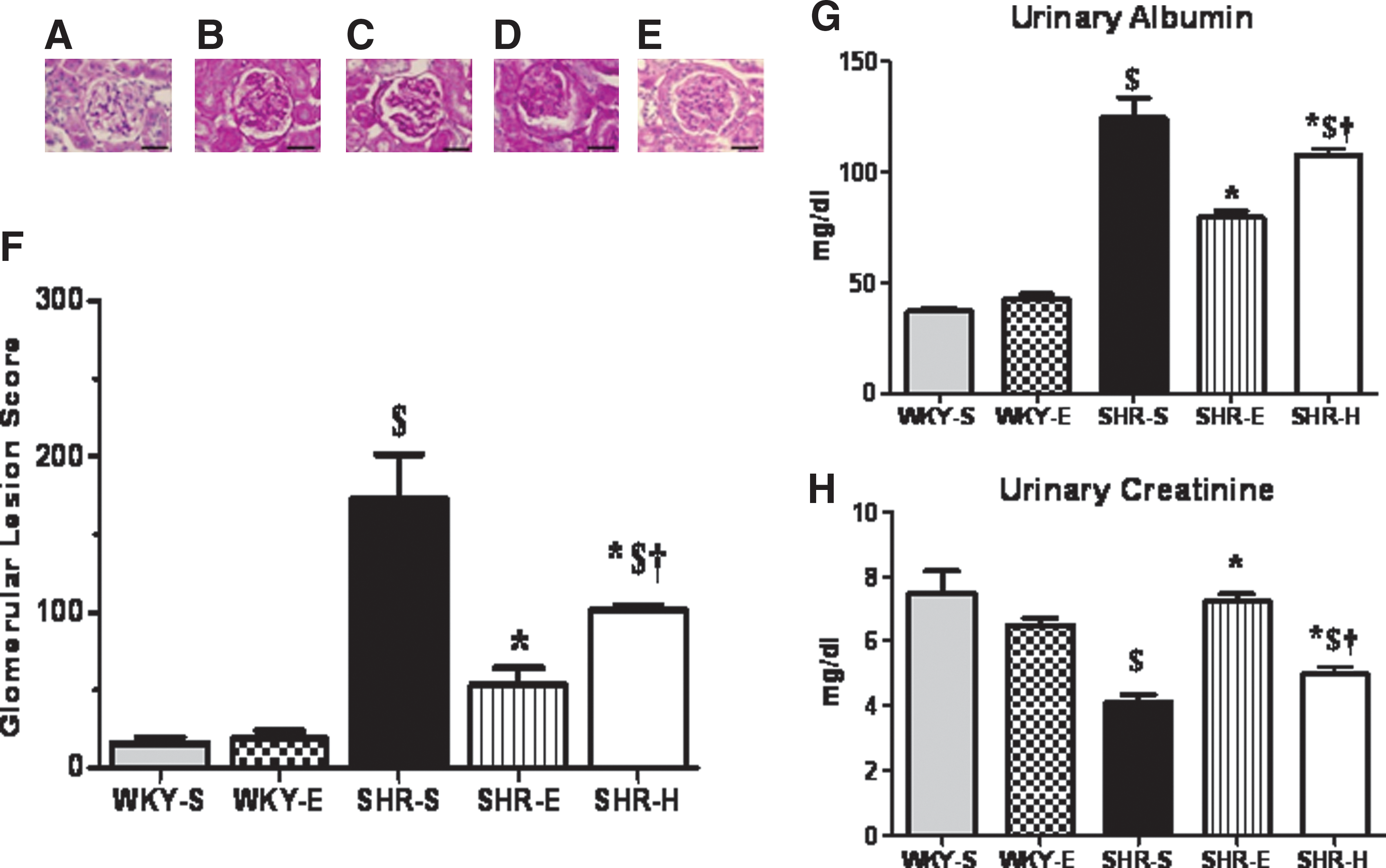
No differences in systolic blood pressure (SBP) or mean arterial pressure (MAP) were noted between WKY and SHR at baseline (Table 1); however, baseline SBP in SHR (124.7±1.99 mmHg) was slightly higher than WKY (120.6±3.32), suggesting that the BP began to increase. At study end, the pressures for SHR-E animals were significantly lower than those for SHR-S animals (Table 2). We observed significantly lowered GFR and RBF and elevated RVR values in SHR-S rats when compared to SHR-E at the study end. However, there was no significant difference in GFR, RBF, or RVR between WKY-S and WKY-E rats. Additionally, significantly higher urine albumin and lower creatinine levels were found in SHR-S than in WKY-S (Figs. 2G and 2H) and ExT resulted in reveral of urine albumin and creatinine levels in SHR.
ExT preserves glomerular morphology and structure in the SHR kidney
PAS-stained kidney sections from rats (n=5 from each group) were examined by a veterinary pathologist who was blinded to the experimental conditions. One hundred glomeruli from each section were scored. Representative photomicrographs of glomeruli for each lesion score appear in Figures 2A–2E. There was no significant difference in glomerular lesion scores for WKY or SHR at baseline (WKY score 27±1.44 and SHR score 34±2.61). SHR-S rats had significantly higher glomerular lesion scores than WKY-S and SHR-E rats at the completion of the study (Fig. 2F).
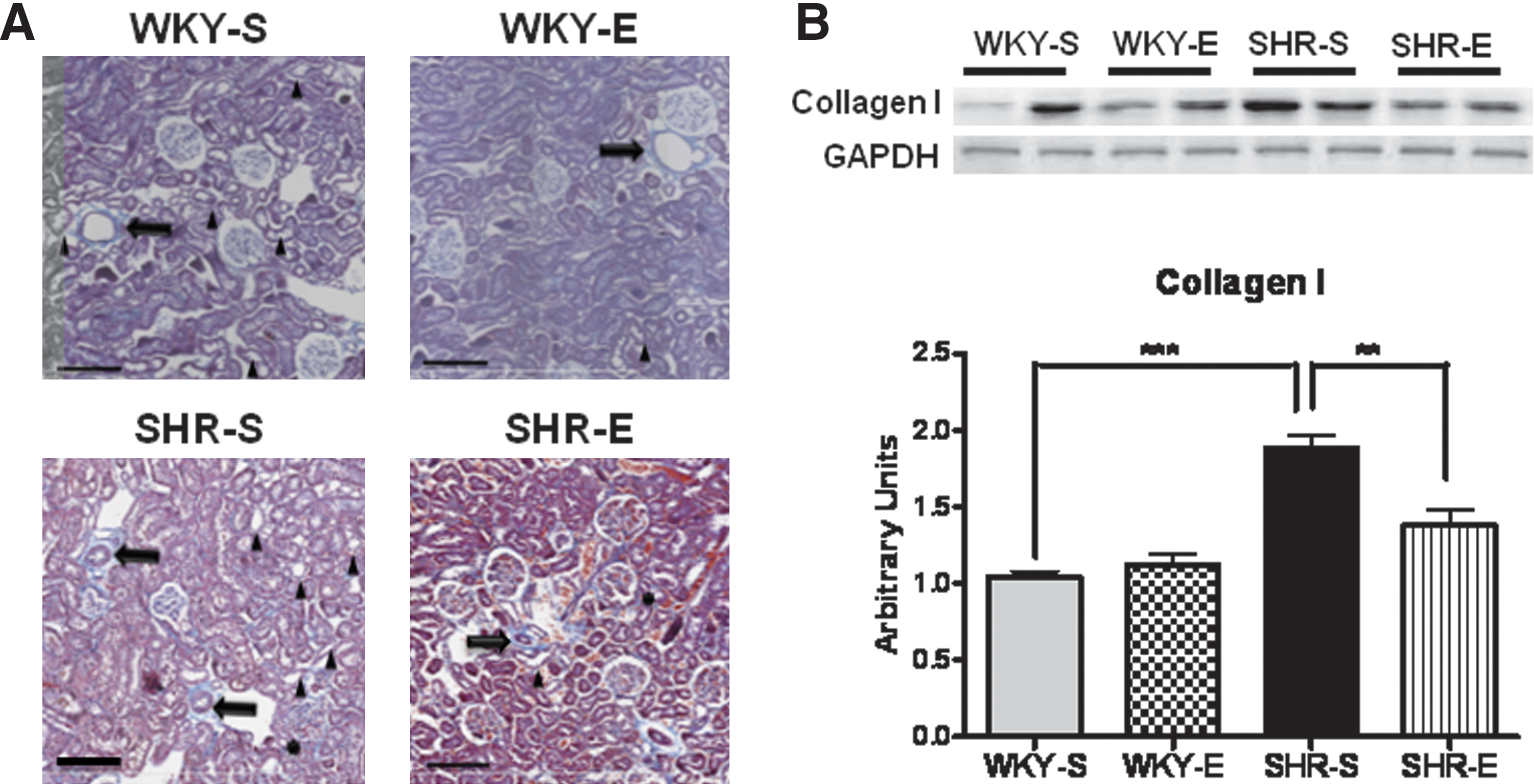
Additonally, kidney sections stained with Masson's trichrome (Fig. 3A) revealed more arterial myointimal hyperplasia, tubular degeneration, ectasia, interstitial, periglomerular and periarterial fibrosis, and glomerular parietal metaplasia in SHR-S when compared to WKY-S. Interestingly, when compared to SHR-S, SHR-E exhibited less arterial myointimal hyperplasia, tubular degeneration, ectasia, interstitial, periglomerular and periarterial fibrosis, and glomerular parietal metaplasia. There were no appreciable differences between WKY-S and WKY-E in aforementioned renal structure except presence for less tubular degeneration and ectasia in WKY-E. Exercise-induced reduction in renal fibrosis was further confirmed, at the molecular level, by assessment of collagen I protein expression. Collagen I levels in SHR-S were nearly 2-fold higher than WKY-S and exercise was found to significantly reduce Collagen I in SHR-S (Fig. 3B).
ExT decreases total ROS and superoxide production in the SHR kidney
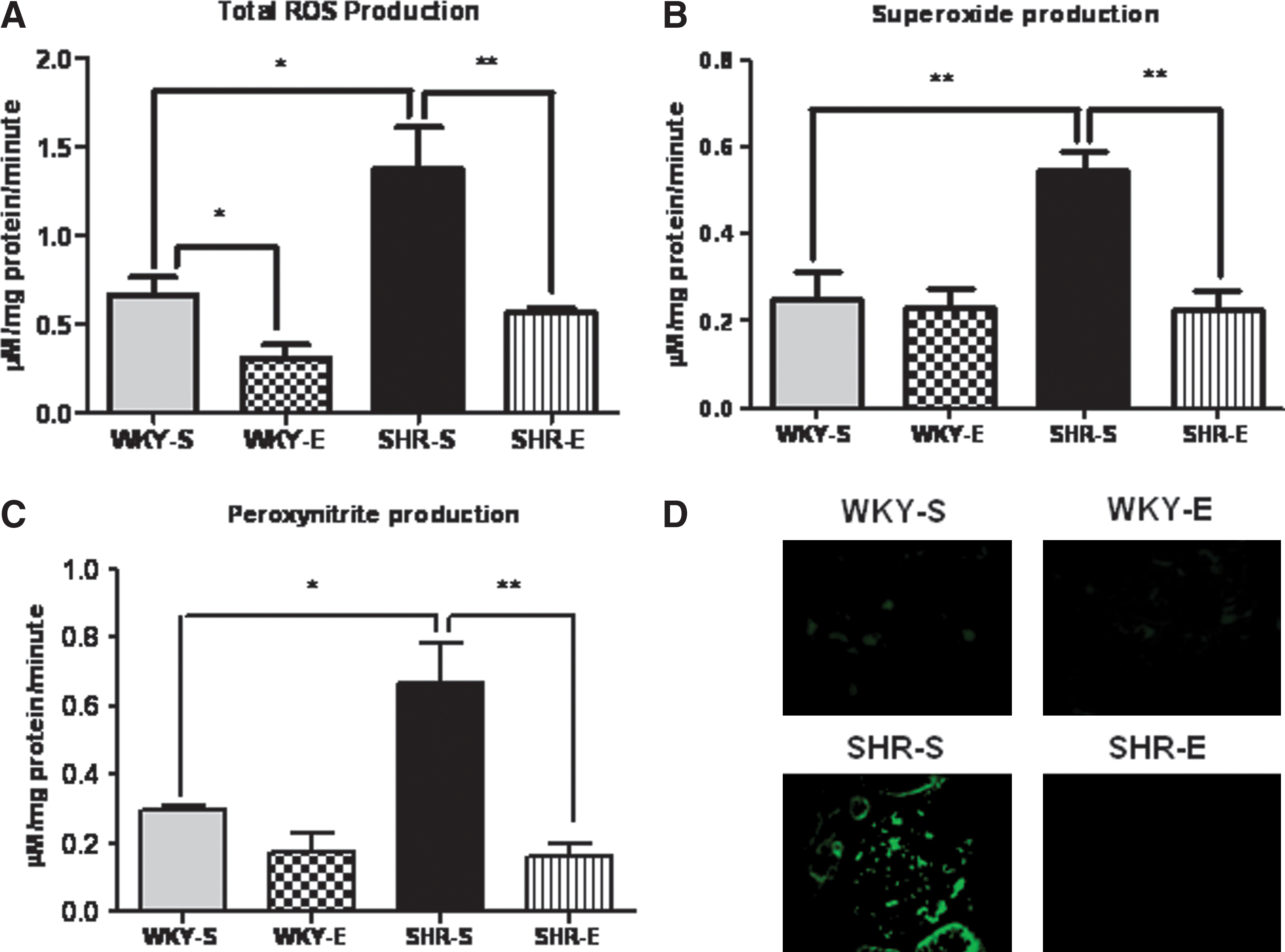
We measured total ROS and superoxide production rates using EPR spectroscopy in cortical tissues from rats (n=6 per group) in all experimental groups. Production rates of both species were significantly decreased in cortical tissues of SHR-E rats when compared to SHR-S rats (Figs. 4A and 4B). No significant differences in total ROS or superoxide measurements were found between WKY-S and WKY-E rats or between WKY-E and SHR-E rats.
ExT decreases peroxynitrite production and NT formation in the SHR kidney
We measured cortical peroxynitrite levels in all rat groups (n=6 per group) using EPR spectroscopy. We also examined expression of 3-nitrotyrosine (NT), a footprint of peroxynitrite formation, via immunofluoresence. In the SHR-E rats, peroxynitrite production rates were significantly decreased and NT expression levels were lower when compared to those of SHR-S rats (Figs. 4C and 4D, respectively).
ExT improves antioxidant status in the SHR kidney
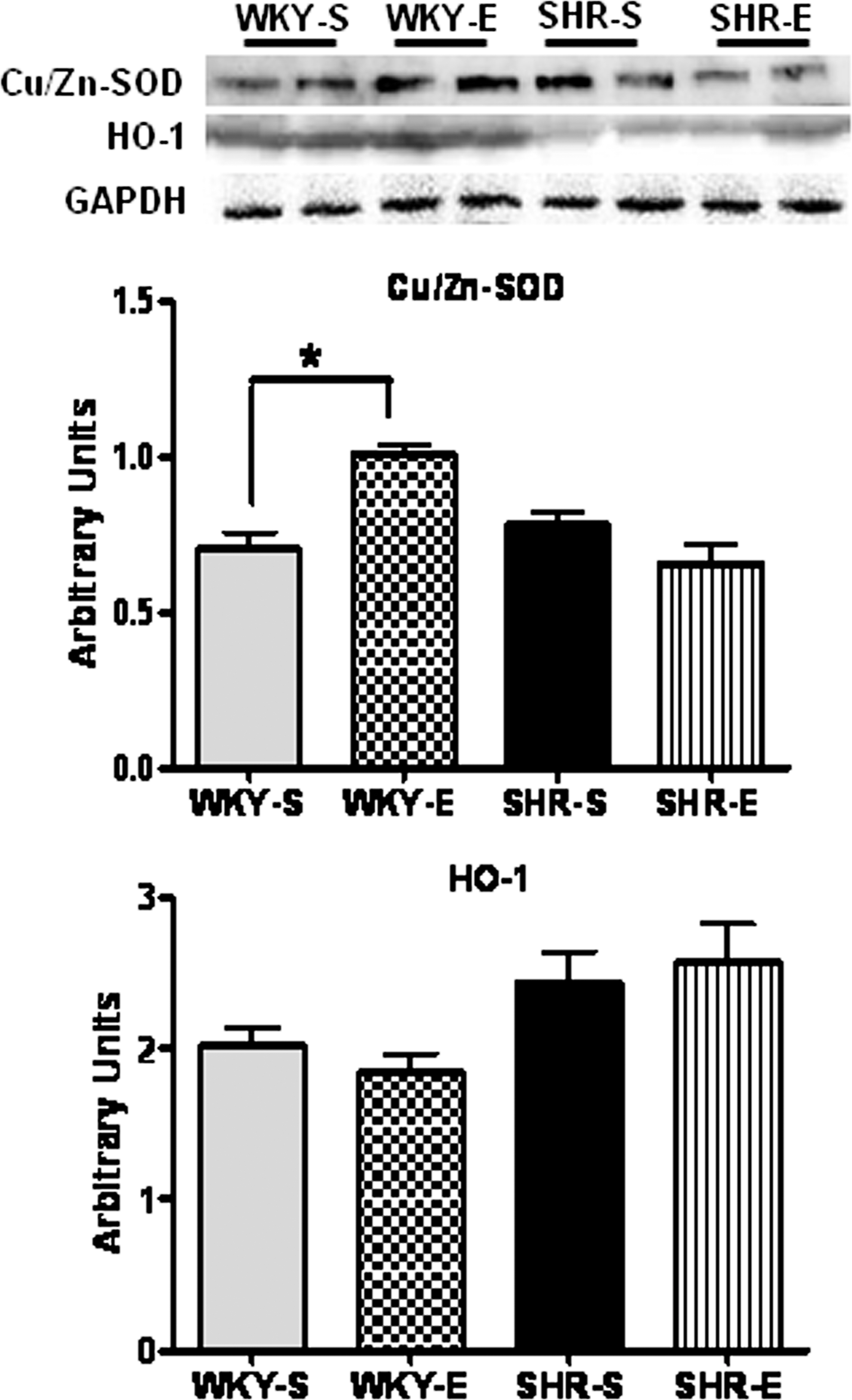
We measured glutathione peroxidase (GPx) and reduced and oxidized glutathione (GSH and GSSG, respectively) in cortical tissues from each experimental group (n=8 per group), using commercially available kits (Figs. 5A–5D). Levels of GPx and GSH were significantly lower in SHR-S when compared to WKY-S. In addition, SHR-S exhibited significantly increased GSSG levels and reduced GSH/GSSG ratio (an important marker of cellular redox balance (43)) in comparison with WKY-S. ExT increased GPx, GSH, and decreased GSSG levels and GSH/GSSG ratio in SHRs. Furthermore, our Western blot analyses did not show any significant difference in the basal levels of renal Cu/Zn-superoxise dismutase (Cu/Zn-SOD) expression between WKY and SHR (Fig. 6). ExT significantly increased SOD and GPx levels in WKY but not in SHRs. Additionally, HO-1 levels were not significantly different among all four groups; however, SHR had slightly higher levels compared to WKY rats (Fig. 6).

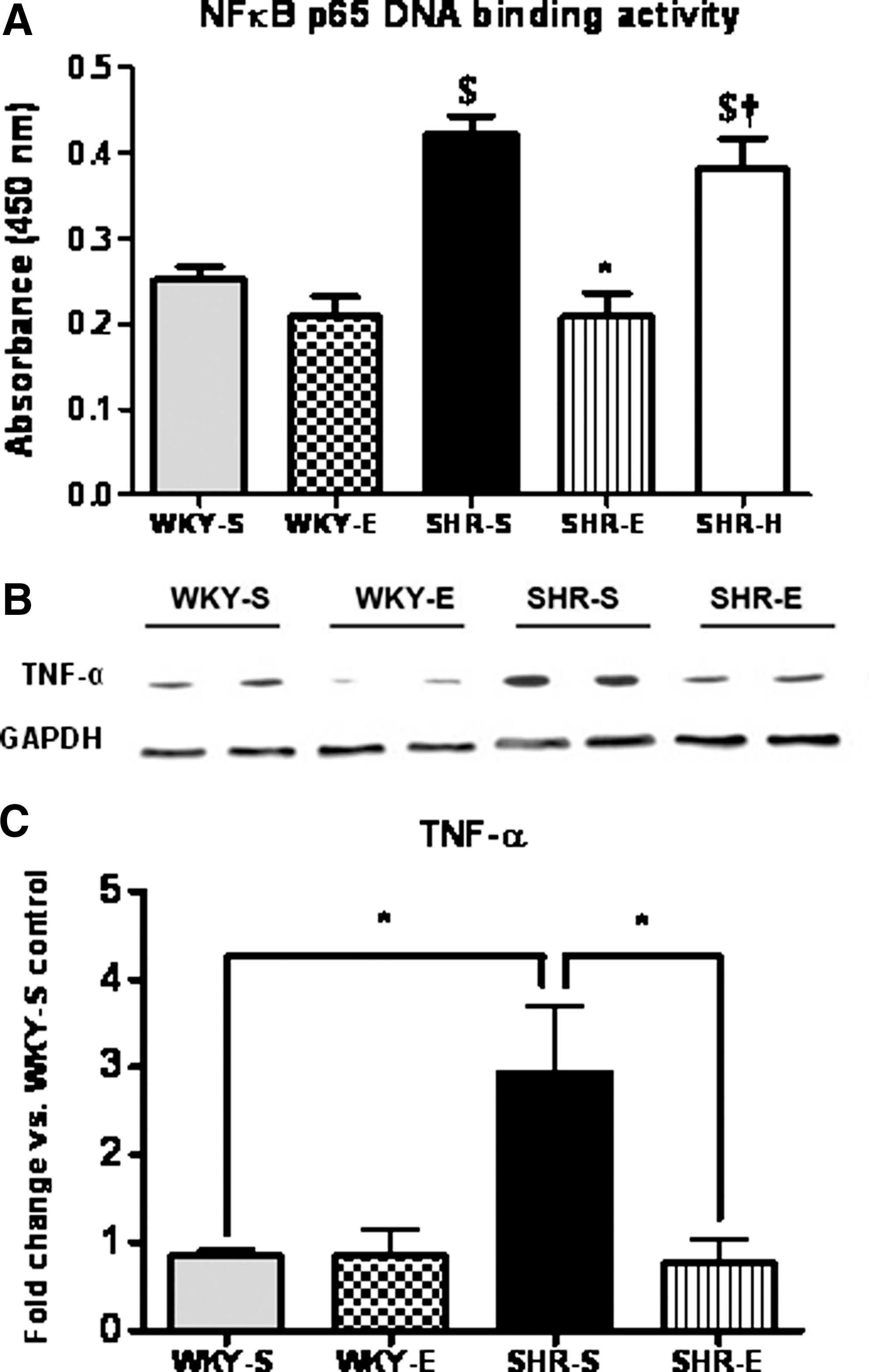
ExT decreases NF-κB activity and TNF-α expression in the SHR kidney
Renal cortical NF-κB p65 DNA binding activity was measured in tissues from all groups (n=8 per group). Rats from the SHR-S group had significantly higher cortical NF-κB activity than WKY-S rats (Fig. 7A). The SHR-E rats had NF-κB activity levels comparable to the WKY-S and WKY-E rats, suggesting that chronic ExT prevents the increase in cortical NF-κB activity seen in SHR. Since the proinflammatory cytokine tumor necrosis factor-alpha (TNF-α) acts through a NF-κB-dependent pathway, we measured protein and gene expression of this cytokine in the cortical tissues of animals from all experimental groups. Expression of TNF-α was significantly increased in SHR-S rats, but was completely normalized in SHR-E rats (Figs. 7B, 7C, and Supplementary Fig. S1).
ExT alters NO production and NOS isoform expression in the SHR kidney
Protein and gene expression levels of eNOS and iNOS were measured in all experimental groups at study completion (Figs. 8A, 8B, 8D, and Supplementary Fig. S1). Cortical nitrate/nitrite levels (indirect indicator of NO production) were also measured (Fig. 8C). Both protein and gene expression levels of eNOS, along with NO production, were decreased in SHR-S rats, while iNOS levels were significantly increased. These effects were normalized in the SHR-E animals.
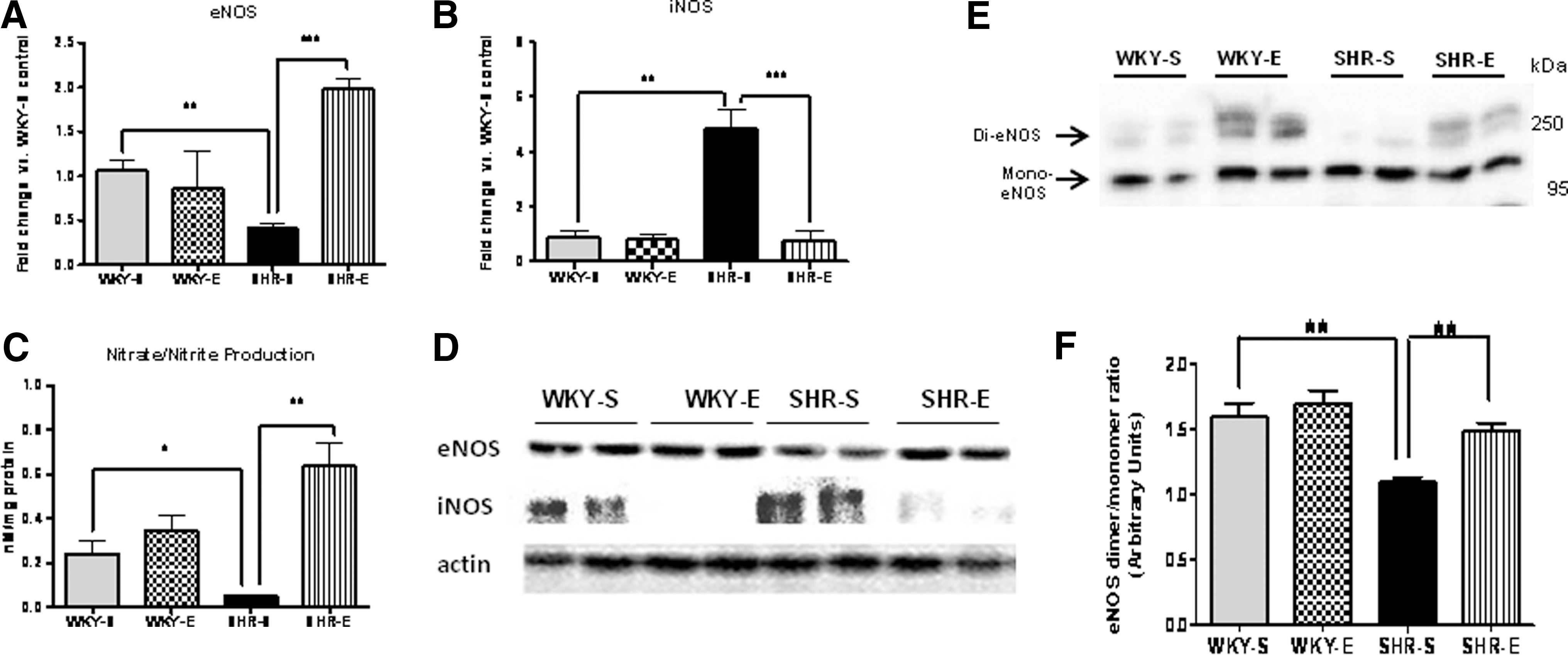
ExT attenuates eNOS uncoupling in SHR
Previous studies have shown that the formation of eNOS homodimers is necessary for eNOS activity (48); therefore, we examined the eNOS dimer and monomer levels by low-temperature SDS-PAGE (LT-PAGE). In sedentary SHRs, eNOS was found to be present predominantly as the monomer; whereas, in WKY-S it was predominantly in the active homodimeric form, suggesting low eNOS activity in SHR-S (Fig. 8E). Densitometric analyses showed that total quantity of homodimeric eNOS and the ratio of dimer:monomer band intensities were more in WKY-S and WKY-E when compared to SHR-S (Fig. 8F). Interestingly, the dimer:monomer ratio was high in SHR-E in comparison with SHR-S. These results suggest that ExT attenuates eNOS uncoupling in SHR, suggesting increased eNOS activity.
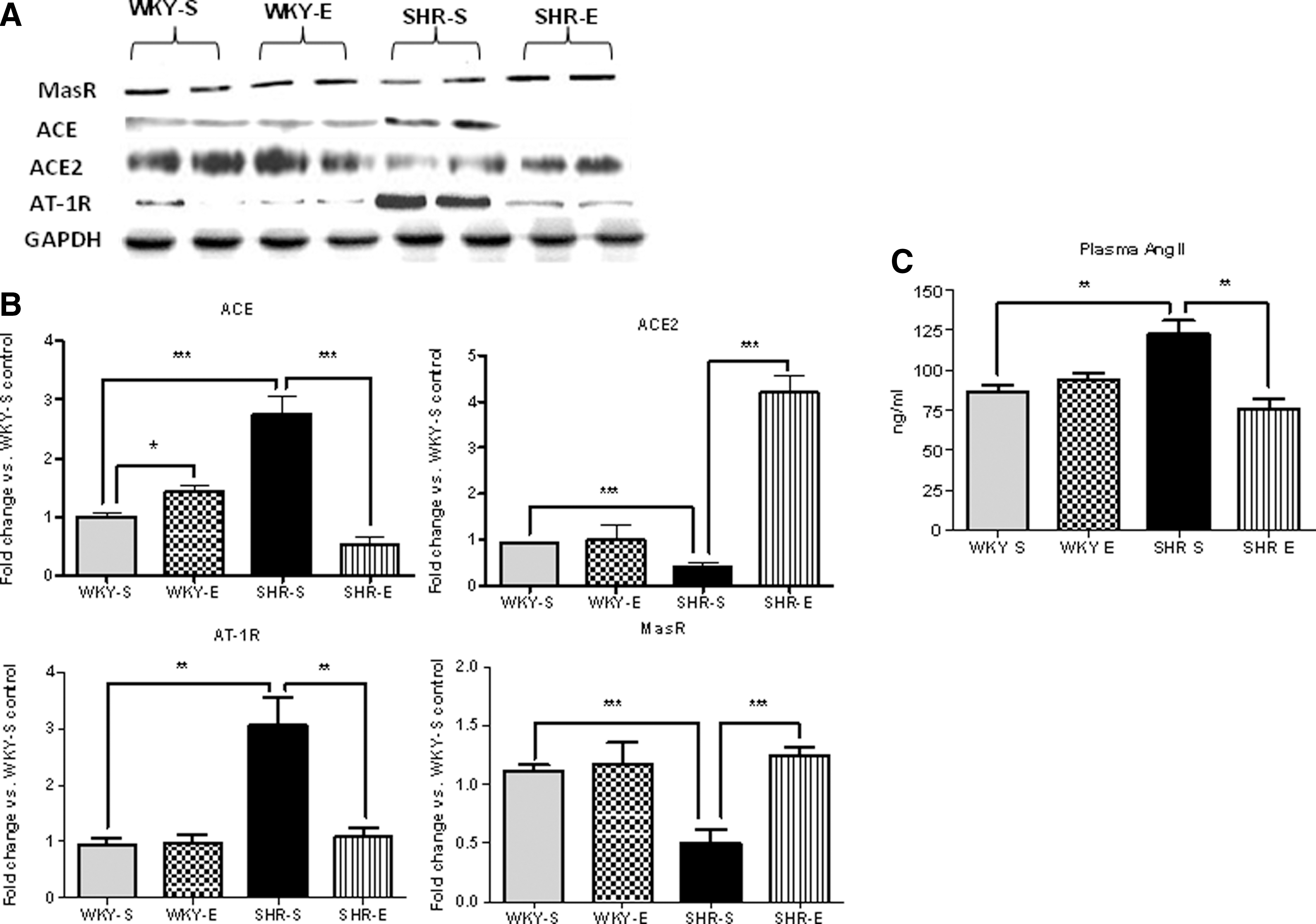
ExT alters expression of RAS components in the SHR kidney
Protein and gene expression (Figs. 9A, 9B, and Supplementary Fig. S1) levels of ACE, ACE2, AT-1 receptor (AT-1R), and Mas receptor (MasR) were measured in all experimental groups at study completion. Circulating AngII levels were also measured in plasma samples from animals from all experimental groups at study end (Fig. 9C). Both protein and gene expression levels of ACE and AT-1R were higher in SHR-S rats, while ACE2 and MasR levels were significantly lower. Similarly, AngII levels were also elevated in SHR-S animals. In SHR-E animals, ACE2 and MasR expression levels were elevated, while ACE and AT-1R expression levels were decreased. Circulating AngII levels were also lower in SHR-E animals.
The effects of ExT on the kidney are, in part, pressure independent
To investigate whether effects of exercise on hypertensive kidneys are pressure-dependent or pressure-independent, a group (n=8) of SHR rats (SHR-H) were treated with hydralazine for the duration of 16 weeks to clamp the BP. Although hydralazine treatment normalized MAP and SBP in SHR (Table 2), it did not appreciably alter the activity of NF-κB (Fig. 7A), the most important redox-sensitive transcription factor. Additionally, similar to SHR-E, SHR-H exhibited significantly improved GFR, RBF, and RVR when compared to SHR-S, and these values were not statistically different between SHR-H and SHR-E (Table 2). Surprisingly, however, urinary albumin levels in SHR-H were slightly increased and creatinine was reduced when compared to SHR-S, however, these values were still significantly different from SHR-E (Figs. 2G and 2H). Importantly, glomerular lesion scores in SHR-H were significantly lower in comparison with SHR-S, although these values were still higher than SHR-E (Fig. 2F). Taken together, these findings suggest that hydralazine treatment in SHR normalizes BP and moderately improves renal function but does not alters glomerular structural changes and NF-κB activity to the level of SHR-E.
Discussion
Primary (essential) hypertension remains a major cause of morbidity and mortality in Western society, and is the second leading cause of end-stage renal disease in the United States (46). The most effective way to avoid the development of hypertension-induced renal injury is to prevent hypertension or to delay its progression. Current pharmacological therapies have proven beneficial in the treatment of hypertension, but the focus has recently shifted to include nonpharmacological approaches, such as exercise, as adjunct therapies to prevent or mitigate hypertension and its end-organ effects. Here, we examined the effects of ExT on renal function and injury and assessed exercise-induced changes in oxidative, nitrosative, and inflammatory parameters in SHR. We initiated chronic ExT at 7 weeks of age, an age when SHR began to develop hypertension and alterations in renal hemodynamic indices that is shown to be preceded by proteinuria (19, 30). We also performed baseline measurements of arterial pressures and renal hemodynamic parameters in 7-week-old WKY and age-matched SHR, with no statistically significant differences found between strains for any of the parameters measured (Table 1). In addition, a group of SHR rats was given hydralazine for 16 weeks, the same length of time that the exercise group was subjected to exercise, to investigate whether exercise-mediated effects are pressure-dependent/independent. The first and foremost finding of this study was that besides significantly reducing SBP and MAP in SHRs, chronic regular ExT preserved renal structure and renal hemodynamics in these animals. Second, moderate-intensity ExT prevents renal oxidative stress and inflammation, improves renal antioxidant defense, and modulates both intrarenal and extrarenal renin–angiotensin system (RAS) components in SHR. Third, ExT attenuates elevated NFκB–DNA binding activity in hypertensive rats. Finally, this study revealed that hydralazine treatment, although able to normalize MAP and SBP in SHR, did not appreciably alter NF-κB activity (Fig. 7A and Table 2). Taken together, these findings clearly suggest that chronic regular exercise reduces BP, causes structural and functional improvements in hypertensive kidneys, and improves RAS components, possibly via reduction in oxidative stress and NF-κB activity in SHRs. In addition, exercise-induced effects in this study, at least in part, were found to be pressure independent.
Our findings revealed that hydralazine treatment in SHR normalizes BP and moderately improves renal function but does not alters glomerular structural changes and NF-κB activity to the level of SHR-E, indicating that factors other than reduction in BP may have contributed to exercise-induced renal changes in SHRs. It is apparent from hydralazine-induced improvements in renal functional parameters that effects of exercise on kidneys of SHR could, partially, be pressure dependent. However, in this study, hydralazine treatment was started from the beginning of the study (at 7 weeks of age) when SHR animals began to develop hypertension and hence, this chronic treatment may have resulted in renal function improvement. In addition, being an antihypertensive drug, hydralazine not only clamps the pressure but also has several other effects on heart, vasculature, and kidney (4, 11). Therefore, it is plausible to suggest that using a compound other than hydralazine that does not have significant effects on renal function would be more appropriate to prove or rule out the pressure-independent effects of exercise in hypertensive kidneys. Nevertheless, lower glomerular scores and albuminuria, higher urinary creatinine levels, and attenuated NF-κB activity in SHR-E in comparison with SHR-H provide evidence that chronic exercise exerts its effects on hypertensive kidneys, at least, in part, might be pressure independent.
Oxidative stress is characteristic of the adult SHR, and has been shown to precede the development of hypertension in this rat strain by several weeks (31, 37). Conversely, hypertension has been shown to cause oxidative stress in the kidney (51). This self-perpetuating cycle, if left unchecked, can lead to progressive renal disease. Several key mediators of renal oxidative stress have been identified, including PIC and the effector peptide of the renin-angiotensin system, AngII; both can cause activation of the key redox-sensitive transcription factor, NF-κB, and increase production of ROS and reactive nitrogen species (RNS), such as superoxide and peroxynitrite, respectively. These ROS/RNS themselves can increase NF-κB activity, leading to further oxidative/nitrosative insult and RAS activation, which perpetuates this vicious positive feedback cycle and accelerates hypertension-induced renal damage (18). A schematic representation of this cycle appears in Figure 10.
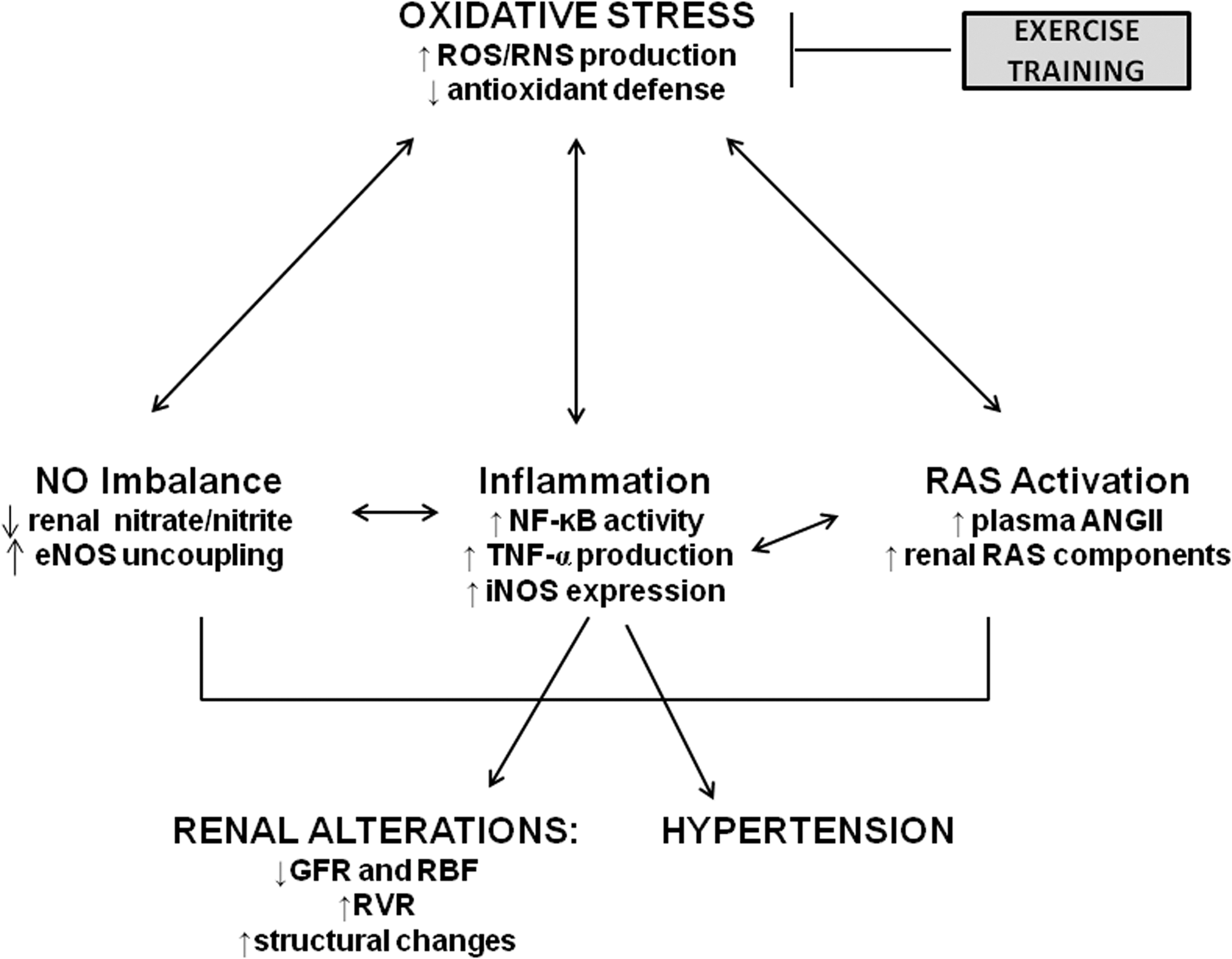
NF-κB can induce and respond to oxidative stress; when activated, NF-κB can activate the promoters of two proinflammatory molecules: TNF-α (12) and iNOS (56). A key finding in the present study is that NF-κB p65 DNA binding activity, along with cortical protein and gene expression levels of iNOS and TNF-α, did not increase in the SHR kidney when ExT was initiated prior to development of hypertension, while all three parameters increased significantly in sedentary SHR. Conversely, gene and protein expression levels of eNOS were significantly higher in SHR-E animals than in SHR-S animals. Further, chronic ExT not only prevented the increases normally seen in cortical total ROS, superoxide, and peroxynitrite production rates in SHR but also significantly upregulated the eNOS protein levels. Our current finding that decreased oxidative stress is associated with decreased TNF-α expression and decreased NF-κB activity in SHR-E rats raises the possibility that decreased PICs might be responsible for the exercise-induced decrease in oxidative stress in SHR.
Oxidative stress, by definition, involves an excess of free radicals; in the case of renal oxidative stress during hypertension, the predominant free radical is superoxide, which is mostly produced by NADPH oxidases (13). We have previously demonstrated an upregulation in NADPH oxidase expression and activity in the cortical tissues of SHR; this was associated with a decline in renal hemodynamic parameters and with increased arterial pressure (18). In addition to its direct detrimental effects, superoxide can interact with NO to form the highly cytotoxic peroxynitrite radical. Peroxynitrite can then react with tyrosine residues in various proteins to generate NT (6,25). Although NT can be formed from reactions that do not include NO (17), the contribution of these reactions to total tissue NT abundance is minimal; thus, tissue NT abundance is largely considered a function of the interaction between superoxide and NO (6). In this study, the increase in arterial blood pressure and alterations in renal hemodynamics seen in sedentary SHR were accompanied by cortical accumulation of NT and increased tissue production rates of total ROS, superoxide, and peroxynitrite. These findings were also accompanied by a marked reduction in renal cortical NO metabolites, which is suggestive of diminished NO bioavailability, likely resulting from an enhancement of NO inactivation by superoxide. Given the current findings that SHR-S had higher iNOS levels than WKY-S and because iNOS can itself increase superoxide production (54, 55), it is highly likely that superoxide-mediated degradation of NO may have contributed to reduced nitrate/nitrite levels in SHR-S. However, concomitant downregulation of eNOS may have also contributed to reduced nitrate/nitrite in SHRs. Regardless, reduced eNOS accompanied by elevated free radical production and decreased nitrate/nitrite levels in SHR-S suggest uncoupling of eNOS. In support of this statement, our densitometric analyses of LT-PAGE clearly showed that total quantity of homodimeric eNOS and the ratio of dimer:monomer were significantly lowered in SHR-S when compared to WKY-S; whereas,ExT in SHR resulted in higher eNOS dimer:monomer ratio, suggesting that chronic exercise attenuates eNOS uncoupling in SHR. Since tetrahydrobiopterin (BH4) is an essential cofactor for eNOS activity and has been shown to be required for promoting and stabilizing NOS protein monomers into the active homodimeric form (45, 50), the eNOS uncoupling observed in SHR-S could be due to a decrease in BH4 levels. Nonetheless, exercise-induced reduction in free radical production along with upregulation of dimeric eNOS may have contributed to renoprotective effects of exercise in SHR as observed in the present study.
In addition to increased free radical production, SHR-S exhibited decreased GPx levels and GSH/GSSG ratio in renal cortex, which is indicative of deranged antioxidant status. Lee and colleagues recently found similar impairments in the glutathione system in SHR from the age of 8 weeks (32). However, we did not observe any significant difference in the basal levels of renal Cu/Zn-SOD between WKY and SHR. This is not unexpected because not all enzymes upregulate their expression in response to an increase in substrate. Consistent with our findings, previous reports have also shown no difference in renal cortical SOD levels between normotensive and hypertensive rats (1, 20, 57). Similarly, previous studies have shown that SOD levels and activity differed in the liver but not in the myocardium of sedentary and exercising WKY and SHR (53). Therefore, the discrepancies in these results could be due to the variations in tissue types studied. Nevertheless, increased superoxide production in absence of any increases in SOD levels in SHR further highlight the point that oxidative stress caused by overproduction of oxidants and impaired antioxidant defense system exist in SHR and may be responsible for observed renal damage in these rats. Unlike SOD, HO-1 levels were not different among all four groups; however, SHRs had slightly higher levels than WKY. HO-1 is known to be induced by various stressors (eg., hypoxia, poisons, and chemicals); therefore, it is likely that increased HO-1 in SHRs could be a compensatory mechanism in response to increased oxidative stress. Interestingly, however, we observed increases in SOD and GPx levels in exercised WKY when compared to WKY-S. Similar results have also been previously reported (5, 27, 29). These findings explain absence of exercise-induced oxidative stress in trained animals or individuals. Our results suggest that increased free radical production accompanied with impaired antioxidant defense system in SHR continues through age 24 weeks and that ExT prevents these alterations in antioxidant status.
In exercised SHR, there was a delayed, moderate increase in arterial pressure and a normalization of cortical NO metabolites. Further, SHR-E exhibited no evidence of cortical NT accumulation and no appreciable increases in tissue production rates of total ROS, superoxide, or peroxynitrite. These results were associated with a preservation of renal hemodynamics and renal antioxidant status in these animals, and with alterations in cortical gene and protein expression levels of iNOS and eNOS (29). Our results are in line with a previous report from our laboratory, where the same ExT regimen resulted in preserved cardiac function, decreased ROS formation, decreased iNOS expression, and increased NO metabolites in the SHR myocardium (2). Taken together, these observations suggest that chronic ExT can decrease the severity of hypertension and its associated alterations in renal hemodynamics in SHR by ameliorating the renal oxidative stress known to exist in these animals. These beneficial effects seem to involve preservation of redox status and an improvement in NO bioavailability.
Activation of the RAS and the resulting AngII-induced pressor response are key mediators of renal damage (26, 42), and have renal pro-oxidant and proinflammatory effects which negatively alter renal hemodynamic parameters (26, 52). For example, AngII can antagonize the effects of NO (59), potentiate superoxide production (52), and activate NF-κB (33); these actions of AngII represent another arm of the vicious positive feedback cycle involved in hypertension-induced renal injury. We measured protein and gene expression levels of several RAS components (AT-1R, ACE, ACE2, and MasR) and circulating AngII levels to assess the possible involvement of the RAS in the renoprotection afforded by ExT in this study. We found downregulation of AT-1R and ACE and decreased plasma AngII, with concomitant upregulation of ACE2 and MasR, in exercised SHR when compared to SHR-S animals. These findings suggest that exercise-induced improvement in renal function may be attributed, at least in part, to improved cortical RAS components. The exact mechanisms by which improvement in RAS may have contributed to exercise-mediated benefits are still unclear. However, RAS-mediated upregulation of renal COX2 has been shown to play an important role in pathogenesis of hypertensive renal injury (28). Therefore, the possibility that downregulation of renal COX2 mediated by reduced vasoconstrictor RAS may have contributed to exercise-induced improvement in renal function and BP, cannot be ignored. Nevertheless, our results indicate a role for decreased RAS activation in the renoprotective beneficial effects of ExT in SHR.
The normal parietal tissue of Bowman's capsule consists of simple squamous epithelium (24). Tubular metaplasia of Bowman's capsule has been well-characterized in SHR and in aging male Sprague-Dawley rats (10, 22, 24); however, the exact cause of such metaplasia remains uncertain. Metaplasia usually occurs in response to chronic inflammation and allows for substitution of cells that are better able to survive under circumstances in which a more fragile cell type might succumb. Although the normal squamous epithelium that lines Bowman's capsule is robust and more resistant to many insults than is cuboidal epithelium, local changes in cytokine, growth factor, or extracellular matrix components arising from either increased glomerular pressure or proteinuria may favor metaplasia to renal tubular epithelium (49). In this study, a higher incidence of glomerular parietal metaplasia, arterial myointimal hyperplasia, and tubular degeneration were seen in SHR-S than any other group whereas the incidence of aforementioned renal structural changes in SHR-E were similar to that of WKY-S and WKY-E animals. These differences were not present in WKY or SHR that were sacrificed at 7 weeks of age, suggesting that the metaplasia seen in SHR-S glomeruli at study completion may indeed be the result of increased pressure and increased inflammation, as previously suggested (24). Besides the abovementioned changes, ExT resulted in reduced periglomerular and periarterial fibrosis as well as collagen I levels in SHRs. It has previously been shown that ACE2 upregulation inhibits Ang II- induced tubulointerstitial fibrosis; whereas the loss of ACE2 enhances Ang II-mediated increase in collagen type I expression and fibrosis (58). Therefore, in light of these reports and our current findings that exercise reduces AngII levels and upregulates ACE2 in SHRs, it is plausible to suggest that exercise-mediated improvement in RAS may have contributed to observed attenuation of renal fibrosis in SHR-E. The absence of significant structural alterations in SHR-E kidneys was associated with improved renal hemodynamics and decreased inflammation, reduced proteinuria, while the increased incidence of metaplasia in SHR-S kidneys was associated with a decline in renal hemodynamics, increased proteinuria, and an increase in inflammation. These results suggest that ExT has ability to improve renal function and structure at physiological and molecular level, possibly through reduction in oxidative stress and attenuated inflammatory cytokines.
We have previously reported an association of PICs and their transcription factor, NF-κB, with oxidative stress and hemodynamic alterations in the adult SHR kidney (18). Given the results from that report and our current results, it is plausible to suggest that the lower NF-κB activity levels seen in SHR-E animals may be attributable to reductions of PIC and ROS production and of RAS activation, and thus, lead to disruption of the positive feedback cycle involved with hypertension-induced renal dysfunction. This study is the first to provide mechanistic evidence surrounding the effects of chronic exercise on NF-κB activity, oxidative stress, hemodynamics, and structure in the SHR kidney. Our results suggest a major role for exercise training in modulating hypertensive renal injury via decreases in inflammation, oxidative stress, and RAS activation. This study also provides evidence that exercise-induced effects in hypertensive kidneys are, in part, pressure-independent as indicated by hydralazine treatment in SHR. The present study clearly suggests that chronic, moderate-intensity ExT may be a cost-effective nonpharmacological approach to preventing hypertension and preserving renal hemodynamics in susceptible patients.
Materials and Methods
For expanded materials and methods, please see the accompanying data supplement. (Supplementary information is available online at
Animals and experimental design
Seven-week-old male normotensive Wistar-Kyoto (WKY) and spontaneously hypertensive rats (SHR), from colonies maintained at the Louisiana State University School of Veterinary Medicine, were used in this study. The original source of the breeder animals was Harlan (Indianapolis, IN). Prior to the initiation of any experimental protocols, five WKY and five SHR were subjected to acute renal clearance experiments as previously published, to obtain baseline values for glomerular filtration rate (GFR) and renal plasma flow (RPF) (18); immediately following clearance experiments, animals were euthanized by thiobutabarbital overdose.
Four groups of eight animals each were used for the experimental protocol: sedentary WKY (WKY-S), sedentary SHR (SHR-S), exercise WKY (WKY-E), and exercise SHR (SHR-E). Animals in exercise groups were subjected to moderate-intensity exercise on a motor-driven treadmill for a period of 16 weeks (2). At the end of the study, six animals from each experimental group were subjected to acute renal clearance experiments (18). Animals in the exercise groups were sacrificed 24 hours after the last exercise session (at age 24 weeks); sedentary animals were sacrificed at the same age. Animals were euthanized immediately after clearance experiments, and kidneys and soleus muscle were collected for further assessment. One kidney from each animal was sectioned and subjected to immunofluorescence studies and histopathological examination. Cortical tissue was separated from the other kidney and stored for later analyses. We performed the following experimental procedures as previously described: real-time RT-PCR, Western blot, electron paramagnetic resonance (EPR) studies, NF-κB p65 DNA binding assay, immunofluorescence, antioxidant assays, and statistical analysis.
In addition, to investigate whether effects of exercise on hypertensive kidneys are pressure-dependent or pressure-independent, a group (n=8) of SHR rats (SHR-H) was treated with hydralazine (25 mg/kg/day) in drinking water daily for 16 weeks. Subsequently, the SHR-H animals were anesthetized, subjected to acute renal experiments, euthanized, and kidneys were collected for NF-κB p65 DNA binding assay and histopathological studies.
Renal clearance experiments
Acute renal clearance experiments were performed according to previously published methods (18).
Glomerular injury scoring
A semi-quantitative glomerular lesion scoring method was used, which was based upon previously published methods for glomerulosclerosis scoring (44) and expanded to include the following criteria: tubular epithelial metaplasia of Bowman's capsule, glomerulosclerosis, mesangial proliferation, and glomerular capillary basement membrane thickening.
Measurement of cortical nitrate/nitrite production
The reaction of nitric oxide (NO) with oxygen can cause the oxidation of NO, leading to nitrate/nitrite production; therefore, levels of nitrate/nitrite are considered an indirect indicator of NO production. Nitrate/nitrite levels were measured in renal cortical tissues of animals from all experimental groups with a commercially available colorimetric assay kit (Cayman Chemical; Ann Arbor, MI), as previously described (2).
Measurement of cortical NF-κB p65 DNA binding activity
Activated NF-κB translocates to the nucleus, where the p65 subunit binds to DNA to promote transcription. The binding activity of free NF-κB p65 in nuclear extracts was assessed with a NF-κB p65 TransAM ELISA kit (Active Motif, Carlsbad, CA) as per manufacturer's instructions, as previously described (35).
Measurement of cortical oxidized/reduced glutathione and glutathione peroxidase levels
Antioxidant status was assessed in renal cortical tissues of animals from all groups by measurement of reduced and oxidized glutathione and glutathione peroxidase, with commercially available colorimetric assay kits (Cayman Chemical, Ann Arbor, MI), as previously described (36).
Analysis of mRNA expression by real-time PCR
Total RNA isolation from renal cortical tissues, cDNA synthesis and real-time RT-PCR were performed as previously described in detail (2, 18).
Analysis of protein expression by Western blotting
Protein expression in renal cortical tissues was determined by Western blot analysis, as described previously in detail (2, 18).
Low-temperature SDS-PAGE (LT-PAGE) for the measurement of eNOS dimer/monomer
To investigate eNOS homodimer formation in kidney cortex, low-temperature SDS-PAGE was employed, as described previously (41, 60).
Electron paramagnetic resonance spectroscopy
Total ROS, superoxide, and peroxynitrite were measured in renal cortical tissues using electron paramagnetic resonance spectroscopy, as described previously in detail (15, 16, 18, 36).
Immunofluorescence
Immunofluorescence detection of 3-nitrotyrosine (Cayman Chemical; 1:100 dilution) in paraffin-embedded kidney sections was conducted as previously described (18).
Statistical analyses
All data are presented as means±SEM. For baseline analyses between WKY and SHR, unpaired t-tests were used. At study completion, unpaired t-tests were used to execute planned comparisons between WKY-S and SHR-S; WKY-E and SHR-E; and SHR-S and SHR-E groups, using Graph Pad Prism software (version 5.0). In all cases, results were considered significant when P<0.05.
Footnotes
Acknowledgments
The authors gratefully acknowledge Sherry Ring for processing, sectioning, and staining tissue sections. This work was supported by NIH National Heart, Lung, and Blood Institute grant HL-80544-04 (J.F.).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.