
Abstract
Dr. Arne Holmgren (Ph.D., 1968) is recognized here as a redox pioneer, because he has published at least one article on redox biology that has been cited over 1000 times and has published at least 10 articles, each cited over 100 times. He is widely known for his seminal discoveries and in-depth studies of thioredoxins, thioredoxin reductases, and glutaredoxins. Dr. Holmgren, active throughout his career at Karolinska Institutet, Sweden, has led the field of research about these classes of proteins for more than 45 years, continuously building upon his sequence determination of Escherichia coli thioredoxin in the late 1960s and discovery of the thioredoxin fold in the 1970s. He discovered and named glutaredoxin and he determined the structure and function of several members of these glutathione-dependent disulfide oxidoreductases. He still continues to broaden the frontiers of knowledge of thioredoxin and glutaredoxin systems. The thioredoxin fold is today recognized as one of the most common protein folds and the intriguing complexity of redox systems, redox signaling, and redox control of cellular function is constantly increasing. The legacy of Dr. Holmgren's research is therefore highly relevant and important also in the context of present science. In a tribute to his work, questions need to be addressed toward the physiological importance of redox signaling and the impact of glutaredoxin and thioredoxin systems on health and disease. Dr. Holmgren helped lay the foundation for the redox biology field and opened new vistas in the process. He is truly a redox pioneer. Antioxid. Redox Signal. 15, 845–851. Experiments carried out with the highest precision should be analyzed with an unbiased mind. Redox biology has a great future. It has always been a great inspiration to know that redox proteins have been around during evolution from the beginning of life on earth and there is yet so much to discover.
Educational and Professional Training of Dr. Holmgren
Area of Interest in Redox Biology—The Thioredoxin and Glutaredoxin Systems
In this short article recognizing the work of Dr. Holmgren, we will reflect upon his key findings on the thioredoxin and glutaredoxin systems in particular, but we should also learn from his strategy of careful, patient, and thorough studies of the molecular mechanisms of these proteins. The type of meticulous biochemical work performed by Dr. Holmgren is as important as ever in this present era of popularity in high-throughput screening projects and beliefs in systems biology. Although modern approaches of high-throughput studies can certainly yield important novel insights of crucial importance for the understanding of organisms, it is probably only from work of lone, focused, curious scientists that genuine discoveries of novel molecular mechanisms can arise. The work of Dr. Holmgren should thus serve as an inspiration for any aspiring researcher who may be interested in the very details of the chemistry of life.

The very first study arising from the work of Holmgren, then working with his fellow student Lars Thelander in the group of Dr. Reichard in Uppsala, related to the effects of ATP on CDP reduction and thus the allosteric regulation of ribonucleotide reductase (36). The regulation of fidelity and activity of ribonucleotide reductase through two separate allosteric effector sites is of utmost intricacy (56) and could have caught anyone's full interest. However, Holmgren choose to focus on the thioredoxin system and this led him into his tremendously successful career in the redox biology field. He published his second paper in 1967 together with Lubert Stryer and Peter Reichard on the conformational change of thioredoxin upon its reduction (61)—an important property of the protein, considering that the redox status-related conformational changes of thioredoxin are today known to affect its binding to master control factors such as apoptosis regulating kinase-1 (49). In his third to seventh papers, thus completing his Ph.D. thesis work, Holmgren worked out a reproducible scheme to prepare thioredoxin from E. coli cells, and by having spent time in Dr. Richard Perham's laboratory in Cambridge, England, to learn sequencing techniques, he determined the amino acid sequence of E. coli thioredoxin (17 –19, 34, 35). The knowledge of the sequence of E. coli thioredoxin and the mapping of its active site -Trp-Cys-Gly-Pro-Cys- laid the very foundation for the whole thioredoxin field. Holmgren subsequently spent more than 2 years crystallizing thioredoxin and, together with Dr. Carl-Ivar Brändén, published in 1975 the 2.8 Å structure of oxidized E. coli thioredoxin (37). It should be noted that the thioredoxin fold is today known as one of the most utilized folds among proteins, catalyzing a wide range of important functions (1b). With the many thioredoxin fold proteins exerting highly diverse reactions within different subcellular compartments and under different conditions, clearly there must be individual protein-specific features of each particular member of this superfamily of proteins. Still, many aspects of the thioredoxin fold proteins are rather universal, and by learning from the key findings of Dr. Holmgren, we can learn much of these specific properties of the thioredoxin fold proteins and the many systems within which they act. Three of his key findings shall be summarized here in slightly more detail.
Description of Key Finding 1
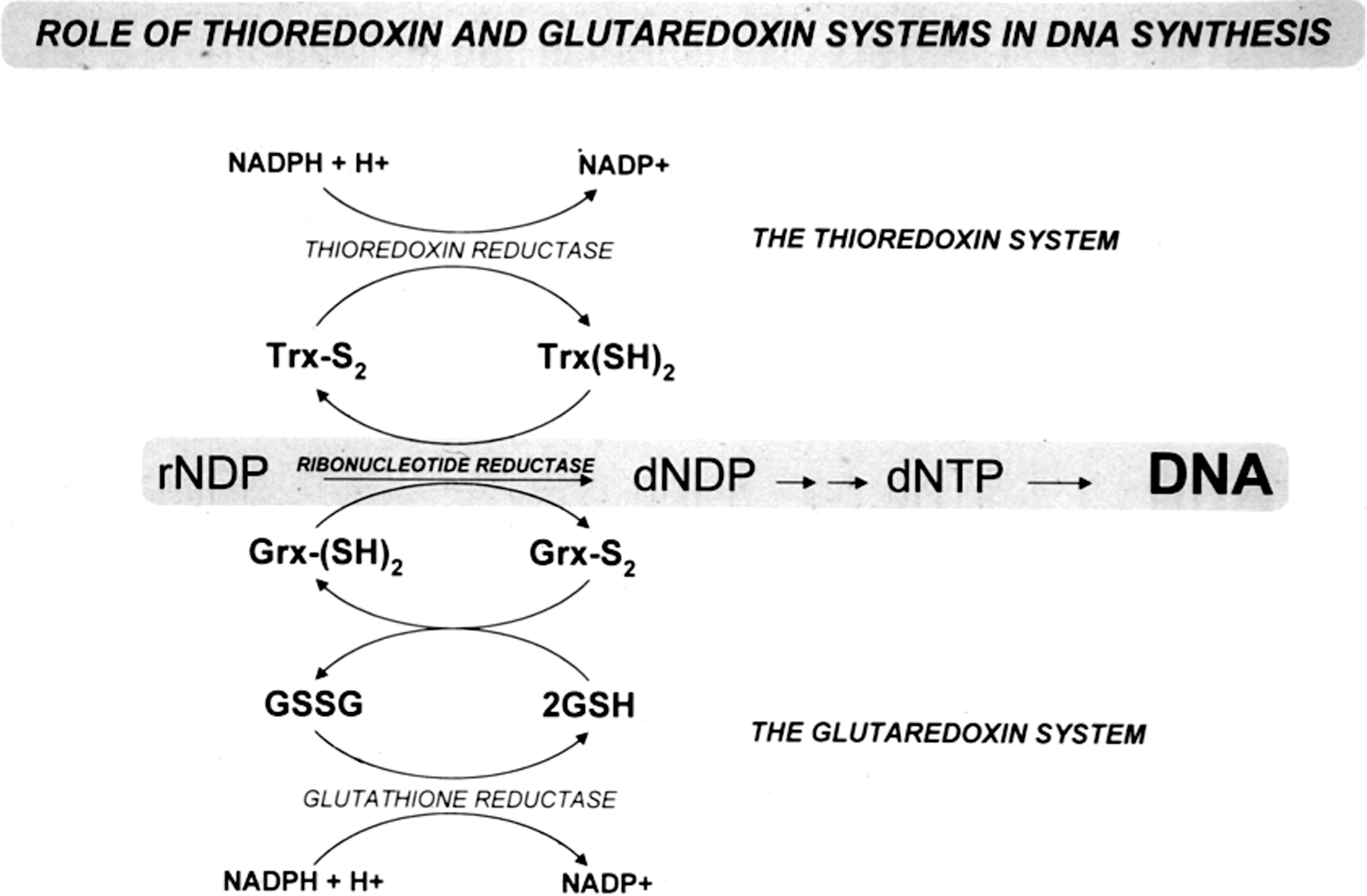
The discovery of glutaredoxin and characterization of glutaredoxin systems
A major contribution of Dr. Holmgren was achieved already during his graduate studies, as was described earlier, that is, the determination of the E. coli thioredoxin primary sequence. However, the two first major awards that he received, the Svedberg Prize (1979) and the Eric K. Fernström Prize (1980), recognized his discovery of E. coli glutaredoxin. His discovery rose from a thorough analysis of the mechanisms by which a strain of E. coli (tsnC7004) shown to lack thioredoxin could actually grow, thus suggesting that a second reducing system for ribonucleotide reductase existed. Through this work, a heat-stable protein that could support ribonucleotide reductase, required glutathione, and clearly was not thioredoxin was discovered by Dr. Holmgren. He named the protein “glutaredoxin” in the first description of this study, published by Holmgren as single author in 1976 (20). Early follow-up studies revealed the presence of similar glutaredoxin-like proteins also in T4 phage-infected E. coli (22) and calf thymus extracts (47). Purification of the E. coli glutaredoxin to homogeneity revealed the specific features of its glutathione-dependent activity with ribonucleotide reductase and additional enzymatic properties, such as being a general glutathione-disulfide oxidoreductase. Interestingly, already at that time there was also evidence for large amounts of additional glutaredoxins being present in E. coli (23, 24). Today, it is well known from the work of Holmgren and many others that most if not all organisms express several isoforms of both thioredoxins and glutaredoxins, carrying out functions that may be either specific or complementary to each other. The major differences between glutaredoxins and thioredoxins are found in their use of glutathione. Even if both classes of proteins belong to the thioredoxin fold superfamily, glutaredoxins utilize glutathione as reductant and thereby also glutathione reductase, whereas thioredoxins are directly dependent upon thioredoxin reductases (Fig. 1). This key difference in function was evident already in the earliest work of Holmgren and has generally held true until today, even if newer findings such as mammalian Grx2 being reduced by thioredoxin reductase (40) show that there may be also a cross-talk between the glutaredoxin and thioredoxin systems. With the glutaredoxins gaining more and more interest, based upon the many functions they carry out that are not necessarily overlapping with those of thioredoxins (14), the initial papers of Holmgren discovering and describing these proteins are truly pioneering works.
Description of Key Finding 2
Catalytic mechanisms of thioredoxins and glutaredoxins
To fully understand the functions and roles of thioredoxin and glutaredoxin (14, 28, 30), Dr. Holmgren has painstakingly studied many crucial details of their structures (6), conformational dynamics (6), substrate specificities (23, 25, 32), and several additional features of these proteins. These studies have laid the foundation for our current understanding of the catalytic mechanisms of both thioredoxins and glutaredoxins. This includes the low pK a of the nucleophilic thiolate of the N-terminally located Cys residue in the -CXXC- active site motif typical for these proteins (10 –12) and the slight but important conformational differences between reduced and oxidized species of thioredoxin that may guide the binding to other protein partners, as mentioned earlier. Importantly, Dr. Holmgren has also characterized the differences between the “dithiol” reduction mechanism for disulfide reduction supported by both thioredoxins and dithiol glutaredoxins, as well as the “monothiol” reduction mechanism seen with glutaredoxins, involving the recognition and reduction of a mixed protein-glutathione disulfide and thus supporting deglutathionylation reactions (5, 6, 51). The functional impact of these qualitatively different mechanisms of glutaredoxin and thioredoxin may be significant, also considering that many “monothiol” glutaredoxins, that is, thioredoxin fold proteins with glutaredoxin homology having only one Cys residue in the active site, have been indeed identified (14). The principal differences between the dithiol and monothiol mechanisms are shown in Figure 2.

Description of Key Finding 3
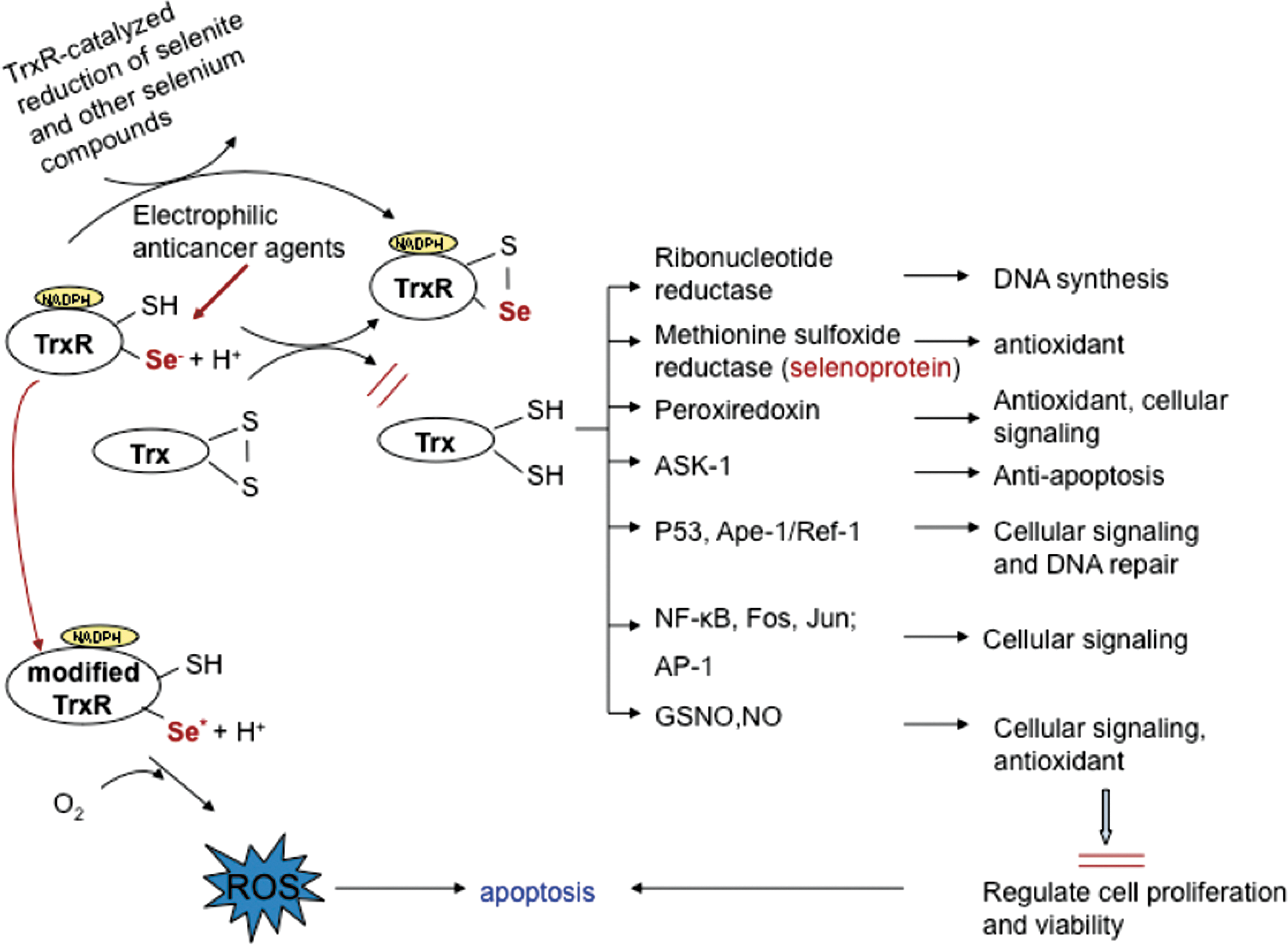
The links between mammalian thioredoxin reductase and selenium
Dr. Holmgren was the first to purify mammalian thioredoxin reductase to homogeneity, from bovine (21), rat (48) and human tissues, thereby discovering that the enzyme was larger and very different from its bacterial counterpart. Dr. Holmgren was also the first to show that a number of selenium compounds could be directly reduced by mammalian thioredoxin reductase, including selenite (42), selenodiglutathione (4), and selenocystine (3). These findings were all fascinatingly integrated into a selenium-centered system when it was discovered by others that human thioredoxin reductase itself is a selenoprotein (16, 62). Dr. Holmgren rapidly showed that the selenocysteine residue of the enzyme was indeed needed for its catalytic activity and he revealed molecular mechanisms by which it functioned (67 –69). He also discovered additional selenium-containing substrates for the enzyme, such as ebselen (64 –66), as well as completely selenocysteine-dependent activities of thioredoxin reductase including reduction of nitrosoglutathione (50) and diverse peroxides (69). He also published the first crystal structure of mammalian thioredoxin reductase (58). Together, these findings intimately link the diverse functions of the complete mammalian thioredoxin system to that of selenium status and thus to the many aspects of selenium in health and disease. This may be also of special importance in relation to cancer growth and anticancer therapy, as summarized in Figure 3.
Other Achievements
Dr. Holmgren has published more than 380 peer-reviewed publications and has clearly had a major impact on research in redox biochemistry. As of today, about 70 of those publications have attracted 100 citations or more (Supplementary Table S1; see • Characterizing thioredoxin as not only a reductant of ribonucleotide reductase but also a powerful general disulfide reductase, and in the process, introducing insulin as an efficient thioredoxin substrate used in assays of thioredoxin activities (25
–27, 33, 59). • Purifying mammalian thioredoxin and thioredoxin reductase to homogeneity, and developing additional specific assays to measure their activities (21, 48). • Beyond discovering glutaredoxin (see earlier text) also determining the glutaredoxin structure using nuclear magnetic resonance (NMR) (5, 7) and catalytic mechanism with ribonucleotide reductase from either E. coli (5) or mammals (63). • Purifying and characterizing Grx1, Grx2, Grx3, and Grx4 from E. coli (1, 1a, 13, 15, 24, 51, 52, 60). • Discovering mammalian glutaredoxin 2 and finding that it is an iron-sulfur cluster protein (38, 44
–46). • Discovering that the N-terminal Cys residue in the thioredoxin active site has a low pK
a and thereby explaining the mechanism of thioredoxin-catalyzed disulfide reduction (41), which was further supported by determination of the structure of reduced thioredoxin (8, 9) (at that time the largest solved 2D NMR protein structure). Also discovering differences between the oxidized and reduced protein (29, 39), which underlies the redox regulation by binding of reduced but not oxidized thioredoxin to selected target proteins. • In a collaborative effort, discovering that thioredoxin regulates photosynthesis (31). • Characterizing extracellular human thioredoxin and the C-terminally truncated thioredoxin-80 and the effects of these proteins as immunomodulatory cytokines (2, 53
–55).
In addition to his many scientific accomplishments, Dr. Holmgren has been also a member of the Nobel Assembly at Karolinska Institutet selecting nobel laureates and of the Swedish Royal Academy of Sciences and recipient of a number of prizes and awards, and he has tutored many prominent scientists in the field of redox biochemistry. Last, but not least, he has vividly entertained many of his colleagues with illustrative recollections of his hunting adventures—his favorite outdoor activity, tracking down moose and wild boars in the Swedish woods aided by his Dachshund dogs. Those dogs have been also loyally joining him in his office. The latest one, Digger, is the current dear and friendly pet in the Holmgren laboratory.
Asking Dr. Holmgren to give some words of wisdom for any future scientists in the field, he gives the following statement:
A citation from the French physiologist Claude Bernard has been a great inspiration: “What we know is a great obstacle to learn more about what is yet unknown to us.” Experiments carried out with the highest precision should be analyzed with an unbiased mind. Redox biology has a great future. It has always been a great inspiration to know that redox proteins have been around during evolution from the beginning of life on earth and there is yet so much to discover. Another great inspiration has been to work on selenium and selenoproteins knowing the pioneering work of Jac Berzelius, who discovered and named selenium in 1818 and was a professor of chemistry and pharmacy here at Karolinska Institutet in Stockholm.
Footnotes
Acknowledgments
The author wishes to thank Dr. Arne Holmgren not only for his accomplishments in redox biochemistry and for the impact that his work has had on the redox biochemistry field, but also for being an excellent scientific mentor and for a long friendship. Dr. Holmgren extends his gratitude to the many extremely talented individuals with whom he has been working over the past 45 years. This includes his mentor, Prof. Peter Reichard, and technicians, secretaries, graduate students, postdocs, and collaborators all over the world. Dr. Holmgren acknowledges that it is their contributions and hard work that has formed the basis for the present knowledge about thioredoxin and glutaredoxin systems.
Abbreviations Used
Reviewing Editors: Ruma Banerjee, James Bardwell, Bob Buchanan, Gary Buettner, William Chillian, Henry Forman, Vadim Gladyshev, Anonymous, Chandan K. Sen, Roberto Sitia, and Christine Winterbourn
Author note: The author of this biography had the privilege and pleasure of joining Dr. Holmgren as a post doc in 1994, subsequently taking over the duties of heading the biochemistry division in 2008. It is with both admiration for his work and delight due to an opportunity to acknowledge the merits of Dr. Holmgren that the author has accepted the invitation by Dr. Sen (Editor, ARS) to summarize the accomplishments of Dr. Homgren—a challenging task indeed.
For a list of frequently cited articles published by Prof. Holmgren, see Supplementary Table S1, available online at
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.