
Abstract
Introduction
In keratinocytes, EGFR can be activated by disparate mechanisms under physiological or pathological conditions by specific pre-existing ligands, such as epidermal growth factor (EGF), transforming growth factor alpha (TGF-α), etc., and via matrix metalloproteinase (MMP)-mediated cleavage of mature ligands from their membrane-bound precursors (25, 30). The autocrine formation of EGFR ligands may be stimulated by the proinflammatory cytokines tumor necrosis factor alpha (TNF-α) and interferon gamma (IFN-γ) (24, 26, 27, 29). Many of the proinflammatory cytokines that are produced upon stimulation with TNF-α/IFN-γ depend on the activation of transcription factors nuclear factor kappa B (NFκB) and activator protein-1 (AP-1) by redox mechanisms (4, 11, 29). Upon activation, the cytoplasmic domain of EGFR dimer undergoes auto- or trans-phosphorylation of distinct tyrosine residues that serve as docking sites for cytoplasmic signal transduction proteins (1, 24, 28, 30, 43). Several lines of evidence suggest the existence of two modes of EGFR signaling. Traditional cytoplasmic EGFR route involves transduction of mitogenic signals through activation of numerous signaling cascades, such as phospholipase C-gamma-protein kinase C, Ras-Raf-mitogen-activated protein kinases (MAPKs), phosphatidylinositol-3-kinase (PI3K)-protein kinase B (Akt), and signal transducer and activator of transcription (STATs) (1, 5, 22, 24, 25). In the nuclear pathway, activated EGFR undergoes fast nuclear translocation, where it physically or functionally interacts with other transcription factors possessing DNA-binding activity and STAT3, that leads to upregulation of distinct genes controlling cell proliferation and DNA repair (20, 24, 44). Both EGFR pathways can be activated in a ligand-free manner, for example, by UV and ionizing radiation, by heat, cisplatin, and H2O2 (8, 22, 24, 25, 30, 43, 44). It seems that cytoplasmic EGFR mechanism mainly controls adaptive inflammatory/stress responses in keratinocytes, whereas the nuclear pathway is involved in keratinocyte proliferation and motility and in their resistance to oxidative stress and heat shock (8, 24, 25). A number of publications from our group (26, 27, 29) has shown that EGFR controls expression of inflammatory chemokines (CXCL8/interleukin 8 [IL-8], CCL2/monocyte chemotactic protein-1 [MCP-1], CXCL10/interferon gamma-induced protein of 10 kDa [IP-10], CCL5/regulated upon activation, normal T-cell expressed [RANTES]) and granulocyte-macrophage growth factor in normal human keratinocytes, and EGFR impairment accounts for overexpression of distinct chemokines in the skin affected by chronic inflammation. It has also been suggested that adverse skin reactions (inflammation and hyperkeratosis) to anti-EGFR antitumor therapies may be a consequence of the impaired control of suppressed EGFR over keratinocyte proliferation and their inflammatory responses (10, 25, 29, 30, 46). Although EGFR involvement in cancer cell proliferation and their chemo-resistance attracts major attention (24), there is substantial evidence that EGFR system is essential to guide keratinocyte proliferation, migration, and survival to maintain normal skin homeostasis or restore epidermal integrity after wounding. Indeed, skin wounding releases EGFR ligands (21) and they potentiate keratinocyte migration (2). The application of EGFR ligands accelerates wound healing (39), whereas pharmacological (28) or genetic (35) EGFR impairment leads to retardation of wound closure/epithelization.
Innovation
Traditionally, anti-inflammatory activity of plant polyphenols (PPs) is ascribed to their direct antioxidant and free radical scavenging activity. Their wound healing effect is often seen as a consequence of suppression of acute inflammation. The in vitro experiments are usually carried out on immortalized or tumor cell lines, which bear significant molecular peculiarities as compared with normal cells. Here, we used primary human keratinocytes for mechanistic studies. We showed for the first time that PPs, depending on the chemical structure of polyphenolic core and the presence of sugar moieties, time-dependently up- and downregulated proinflammatory and repair-regulating chemokines and accelerated in vitro and in vivo wound healing through their interaction with different cytoplasmic and nuclear components of epidermal growth factor receptor (EGFR) system. No correlation with PPs antioxidant/superoxide scavenging properties was found. Very first experimental evidence was obtained on the involvement of EGFR nuclear translocation and retention as possible molecular targets for the skin inflammation/repair modulating PPs.
There is a continuously growing interest in beneficial effects of plant polyphenols (PPs) toward human skin: anti-inflammatory, wound healing, cancer preventing, antiage, etc. The skin benefits of PPs have been traditionally attributed to their chain-breaking antioxidant or free radical scavenging activities (6, 14). However, evidence from numerous in vitro skin cell studies suggests that PPs can influence cellular functions by multiple other mechanisms, such as interaction with several receptors, modulation of signal transduction and transcription of a number of genes, post-translational modulation of enzymatic activities (9, 12, 16, 18, 19, 31, 33, 36, 40), and epigenetic regulation of gene expression (42). Although most of these functions are redox-dependent, their modulation does not compulsory depend on direct antioxidant/free radical scavenging/metal chelating properties of PPs (15, 18).
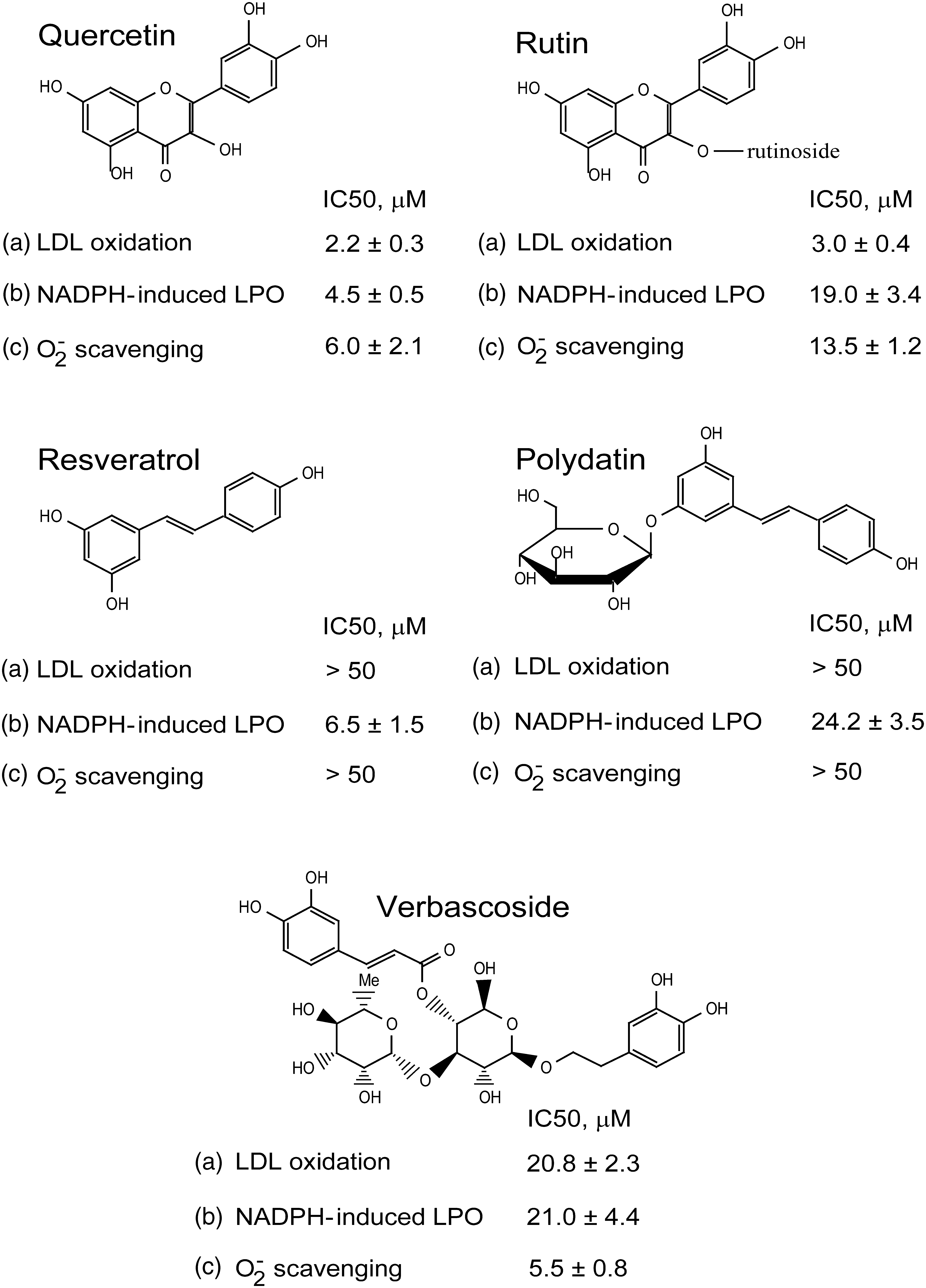
Here, we focused on the elucidation of EGFR-related mechanisms underlying modulation of inflammatory responses and skin repair processes by selected PPs-antioxidants belonging to three major groups: flavonoids, stilbenes, and phenylpropanoids. The pairs of glycosylated and nonglycosylated flavonoids (rutin [Rt] and quercetin [Qr], respectively), glycosylated and nonglycosylated forms of stilbenoids (polydatin [Pd] and resveratrol [Rv], respectively), and glycosylated phenylpropanoid verbascoside (Vb) (Fig. 1) were studied. Primary cultures of human keratinocytes were used for mechanistic studies and the in vitro data were confirmed in the animal model of excision wound. We found that PPs could differentially affect cytoplasmic and nuclear parts of EGFR system. These effects did not correspond to antioxidant or superoxide scavenging capacities of PPs.
Results
Antioxidant and superoxide scavenging properties of selected PPs
Antioxidant properties of PPs were closely tested in two in vitro systems resembling free radical-driven oxidation under physiological conditions, namely, low-density lipoprotein (LDL) oxidation by myeloperoxidase (MPO)+H2O2 in the presence of nitrite and NADPH-induced lipid peroxidation (LPO) in microsomes. Two flavonoids, Rt and its aglycon Qr were the most effective inhibitors of LDL oxidation, whereas Vb was 10 times less effective, and both stilbenes Rv and Pd did not inhibit the reaction up to 50 μM concentration (Fig. 1). On contrast, two aglycons, Qr and Rv inhibited LPO at low micromolar concentrations, and three glycosylated PPs, Rt, Pd, and Vb were less effective. To avoid the interference of PPs with xanthine oxidase routinely applied for the in vitro superoxide generation, we used the reaction of photo-oxidation of riboflavin as a source of O2 −, and the scavenging action changed in the range: Qr=Vb > Rt > Rv=Pd.
PPs modulate constitutive EGFR-extracellular regulation kinase signal transduction and chemokine expression
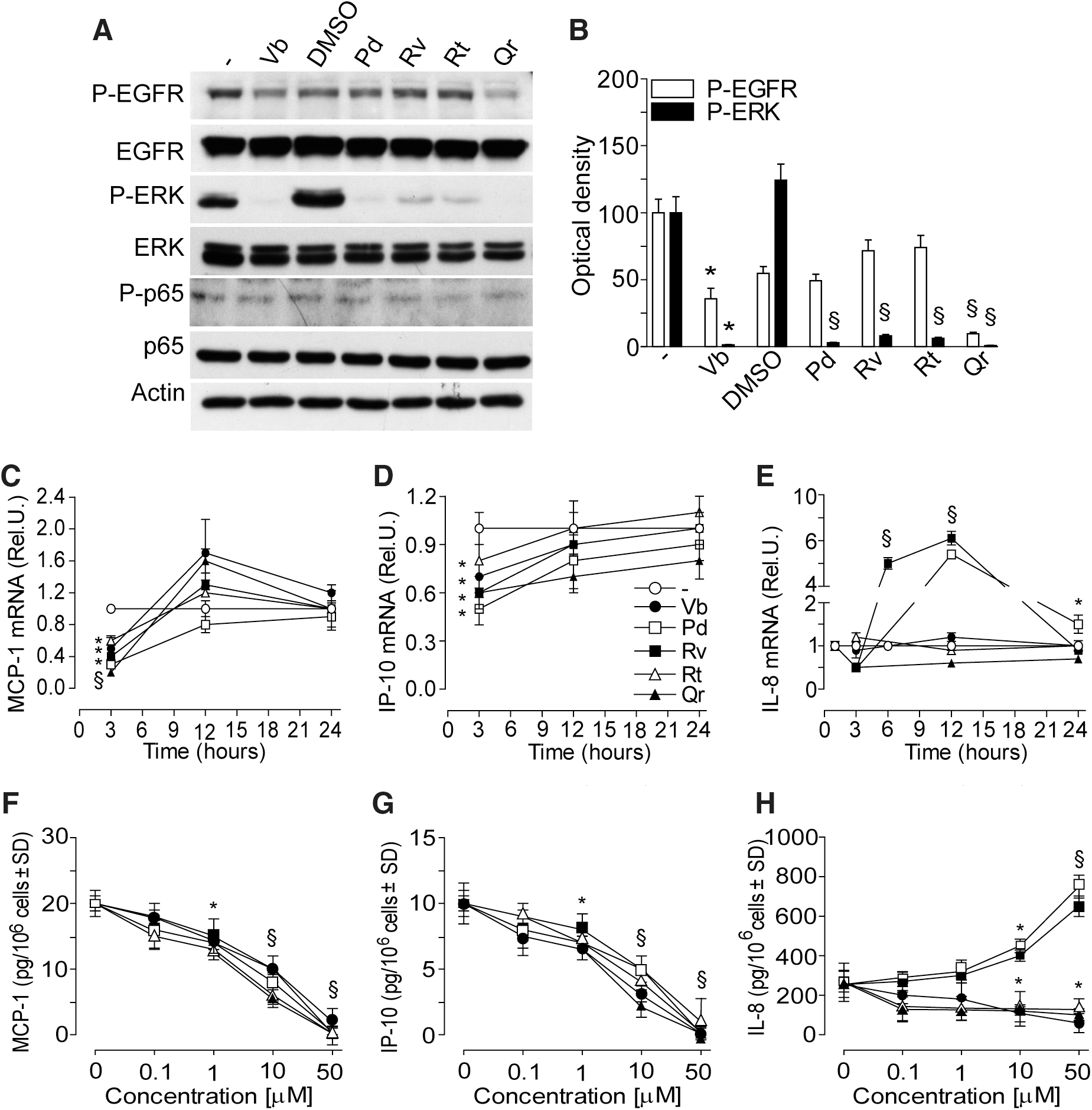
At 15 min, only 50 μM Vb and Qr significantly reduced EGFR phosphorylation, whereas all PPs abrogated extracellular regulation kinase (ERK) phosphorylation (Fig. 2A, B), and did not affect low background level of p65 phosphorylation (Fig. 2A). At 3 h incubation, the transcripts of both MCP-1 and IP-10 were significantly and transiently downregulated by all the PPs (50 μM), then, at 12 and 24 h time-points the constitutive levels were restored (Fig. 2C, D). The expression of IL-8 was dramatically upregulated by Rv at 6 and 12 h, whereas Pd significantly increased IL-8 transcripts at 12 and 24 h (Fig. 2E). All the PPs significantly and dose-dependently reduced the content of MCP-1 and IP-10 in the 24 h supernatant (Fig. 2F, G). Vb, Rt, and Qr dose-dependently reduced, whereas both Pd and Rv (10 or 50 μM) significantly enhanced IL-8 release by normal human epidermal keratinocytes (NHEK) (Fig. 2H).
PPs differently affect TGF-α-driven EGFR-dependent intracellular signaling and TGF-α-associated chemokine expression
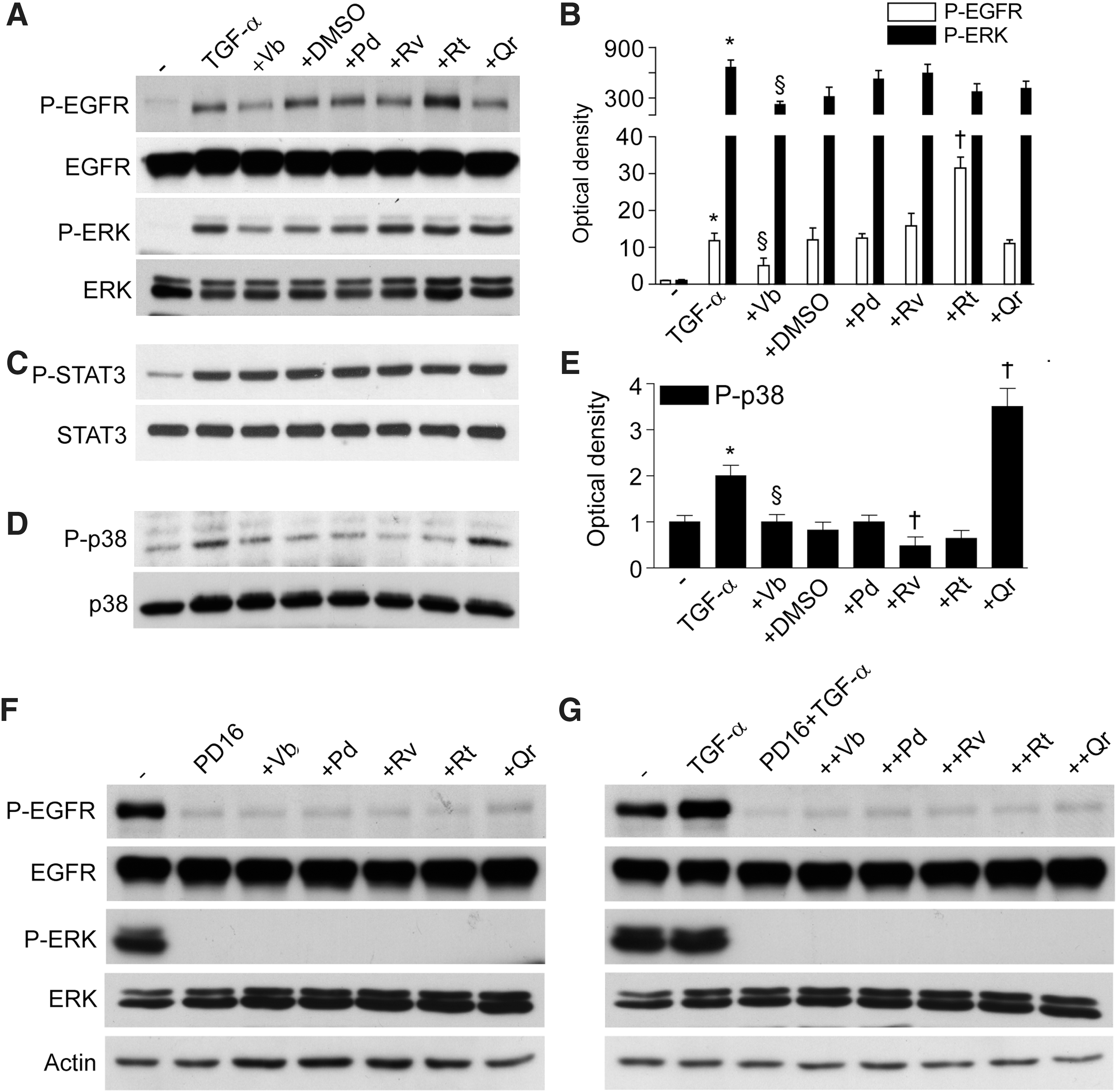
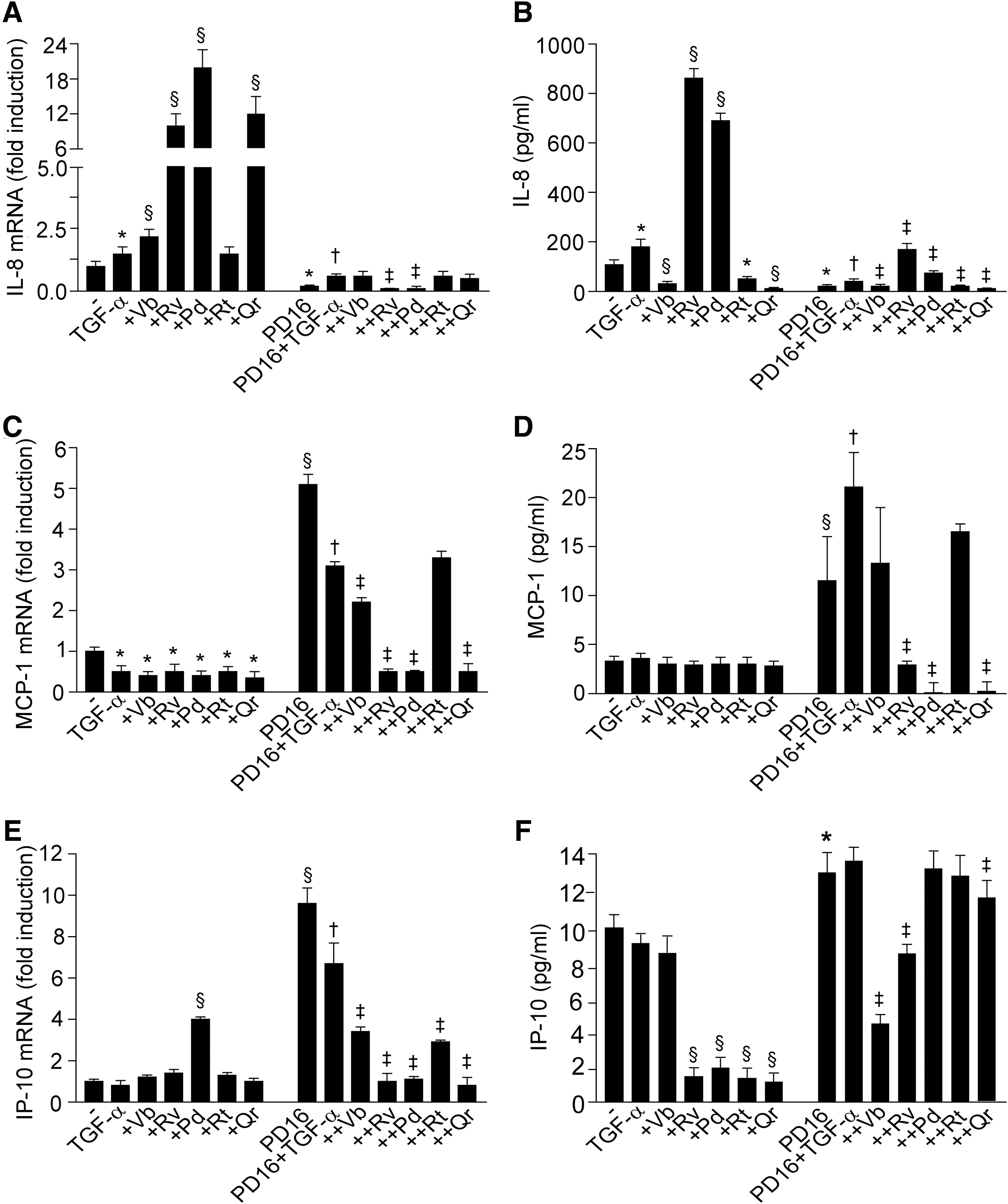
Being a prototypic EGFR ligand, TGF-α (50 ng/ml, 15 min) stimulated phosphorylation of EGFR and ERK (Fig. 3A, B). This TGF-α-induced phosphorylation was not influenced by PPs, except for Vb inhibiting, and Rt further enhancing EGFR activation. As expected, TGF-α also induced STAT3 and p38 phosphorylation. While none of the PPs studied (50 μM, 1 h preincubation) affected STAT3 phosphorylation (Fig. 3C), Vb and Rv inhibited, and Qr further enhanced p38 phosphorylation (Fig. 3D, E). NHEK incubation with PD168393 (2 μM, 30 min preincubation), a specific inhibitor of EGFR tyrosine kinase, resulted in abrogation of spontaneous and TGF-α-induced EGFR and ERK phosphorylation, and PPs did not reverse this dramatic effect (Fig. 3F, G). Of note, PD168393 effectively opposed both IL-8 gene overexpression associated to TGF-α, TGF-α+Vb, TGF-α+Rv, TGF-α+Pd, and TGF-α+Qr (Fig. 4A) and also the enhanced IL-8 protein release induced by TGF-α, TGF-α+Rv, and TGF-α+Pd (Fig. 4B). We also observed that PD168393 strongly upregulated both constitutive and TGF-α-associated MCP-1 (Fig. 4C) and IP-10 (Fig. 4E) gene expression. A significant PD168393-induced increase of these two chemokines was also detected at the protein level (Fig. 4D, F). Notably, Rv, Pd, and Qr strongly inhibited the effect of PD168393 on MCP-1 expression (Fig. 4C, D), and also on IP-10 transcript (Fig. 4E). By contrast, Vb and Rv were the most effective inhibitors of IP-10 protein release (Fig. 4F).
PPs differently affect TNF-α+IFN-γ-induced chemokine expression and intracellular signaling
Analysis of dose-dependent effects of PPs on IL-8, and IP-10 release from NHEK exposed to TNF-α+IFN-γ (T/I) showed that Pd and Rv (50 μM) maintained their ability to enhance, whereas Vb > Qr=Rt inhibited IL-8 release (Fig. 5A). In contrast, all the PPs dose-dependently inhibited IP-10 (Fig. 5B) and MCP-1 (data not shown) release. Cell penetrable compounds, SOD-mimicking MnTMPyP, and iron chelator L1, did not exert any effect (Fig. 5A, B). As expected, NHEK treatment with T/I induced the phosphorylation of EGFR, ERK, and p65. All the PPs significantly inhibited ERK and p65 phosphorylation, whereas only Vb and Qr effectively opposed EGFR phosphorylation (Fig. 5C, D). Vb, Rv, and Qr were all effective inhibitors of both T/I-induced NFκB and AP-1 binding activity, with Vb strongly acting against both transcription factors (Fig. 5E, F).

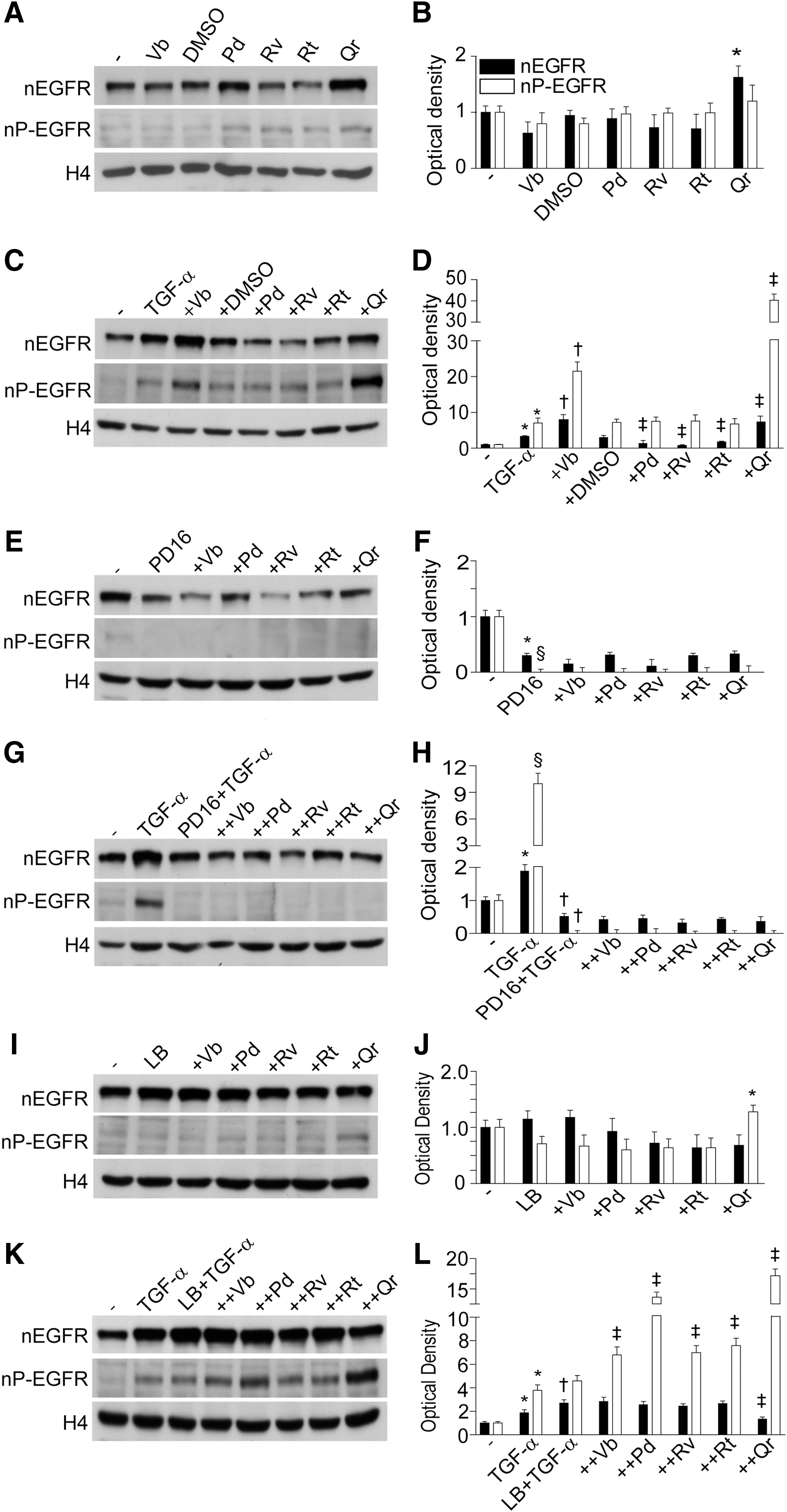
PPs differently affect EGFR translocation or/and retention in the nucleus
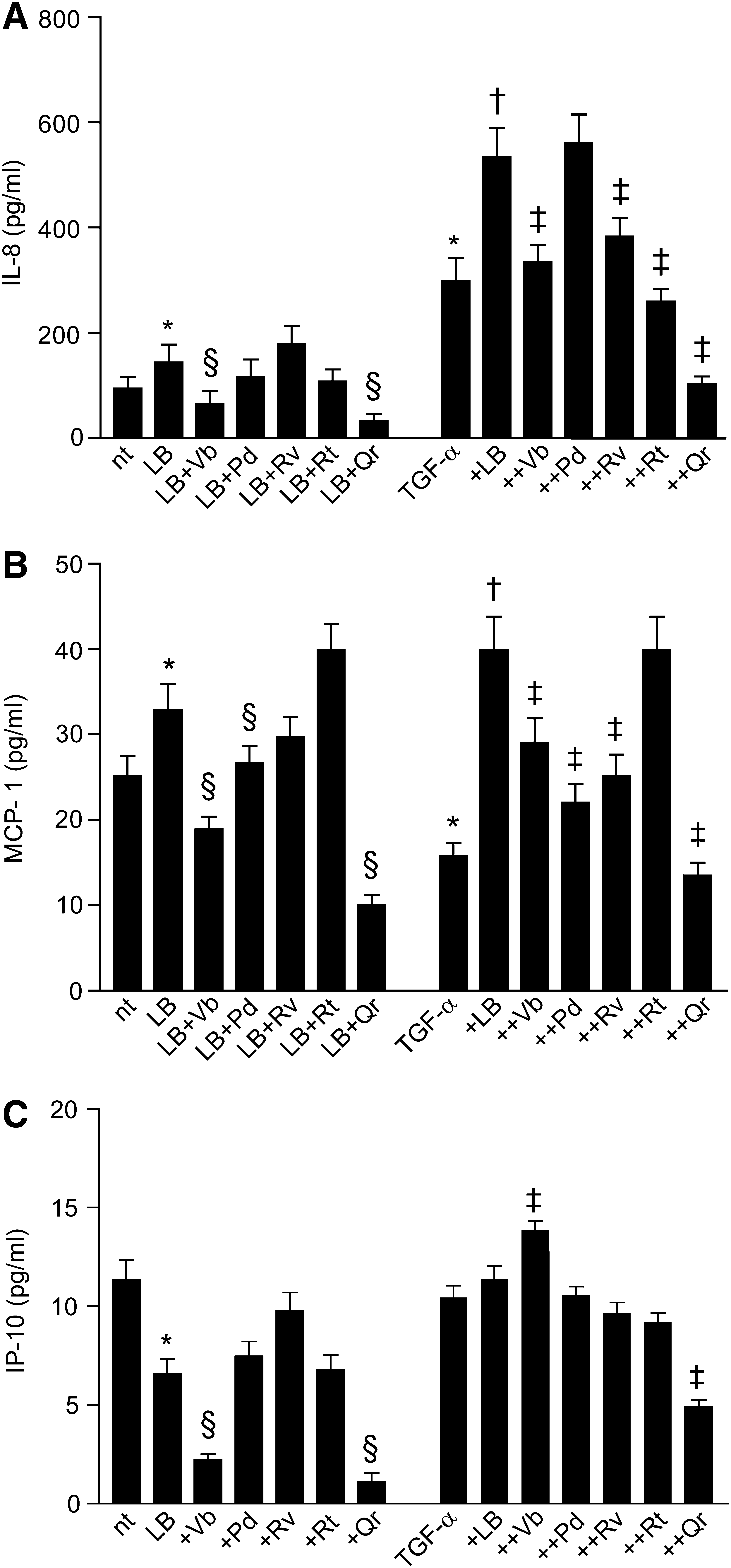
We observed that Qr exclusively enhanced background nuclear levels of EGFR (nEGFR) with no perturbation of the nuclear levels of P-EGFR (nP-EGFR) (Fig. 6A, B). NHEK treatment with TGF-α led to an early (15 min) increase in both nEGFR and nP-EGFR. In this condition, both Vb and Qr further increased nEGFR and nP-EGFR, whereas Pd, Rv, and Rt inhibited nEGFR only (Fig. 6C, D). The presence of PD168393 dramatically reduced both nEGFR and nP-EGFR in nonstimulated (Fig. 6E, F) and TGF-α-stimulated (Fig. 6G, H) NHEK, no effects of PPs were observed. By contrast, the combination Qr+LB increased nP-EGFR levels (Fig. 6I, J). In TGF-α-stimulated NHEK, leptomycin B (LB) further selectively enhanced nEGFR (Fig. 6K, L). The TGF-α+LB+PP combination induced a significant increase of nP-EGFR (Qr>Pd>Rv=Vb=Rt) and TGF-α+LB+Qr led also to significant nEGFR reduction (Fig. 6K, L). LB substantially increased background release of IL-8 and MCP-1 from NHEK, and effect toward IL-8 was significantly impaired by Vb and Qr (Fig. 7A), and toward MCP-1 by Pd (Fig. 7B). By contrast, IP-10 release was downregulated by LB and Vb and Qr further reduced the IP-10 levels (Fig. 7C). In TGF-α-stimulated NHEK, LB worked by further increasing IL-8 release, and, apart from Pd, all the PPs opposed LB-associated increase of IL-8 (Fig. 7A). On the other hand, LB contrasted TGF-α-associated suppression of MCP-1 and strongly enhanced the release of this chemokine, whereas Vb, Rv, Rt, and Qr significantly reduced LB activity (Fig. 7B).
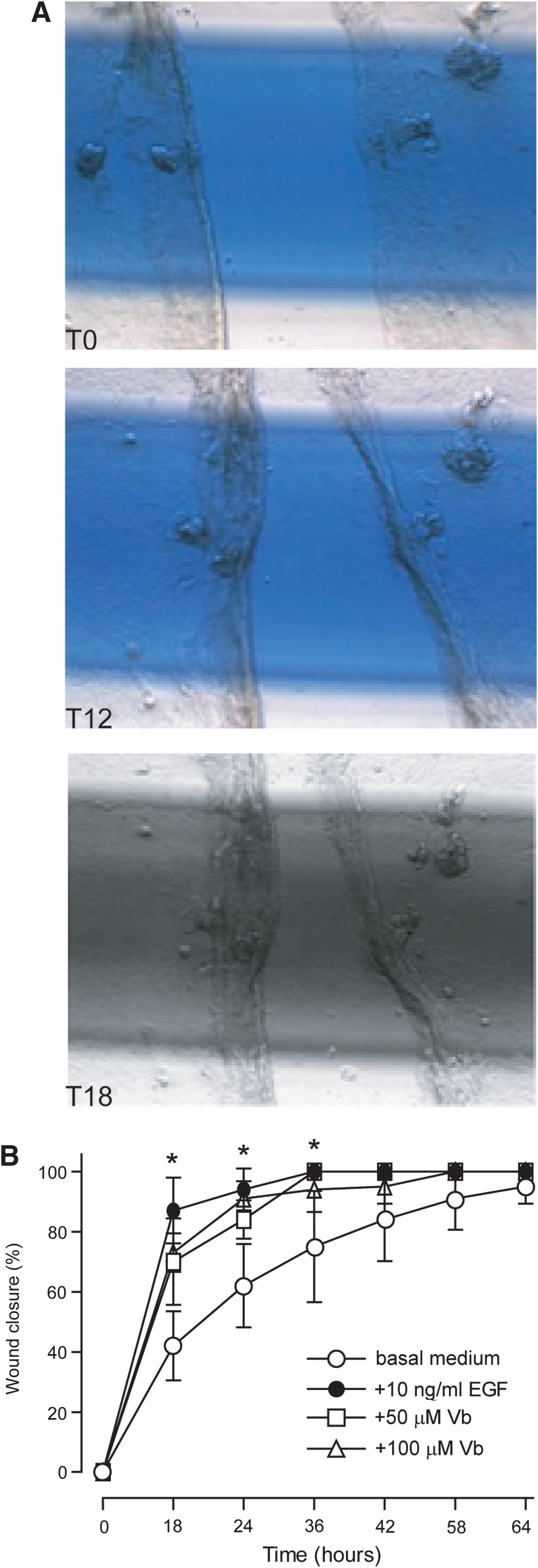
Vb accelerates in vitro scratch wound healing and exerts anti-inflammatory and wound healing effects in vivo
In the in vitro model of scratch wound healing (Fig. 8A), Vb significantly accelerated restoration of damaged NHEK layer at 18 and 24 h time points, with an effect comparable to the healing effect of EGF (Fig. 8B).
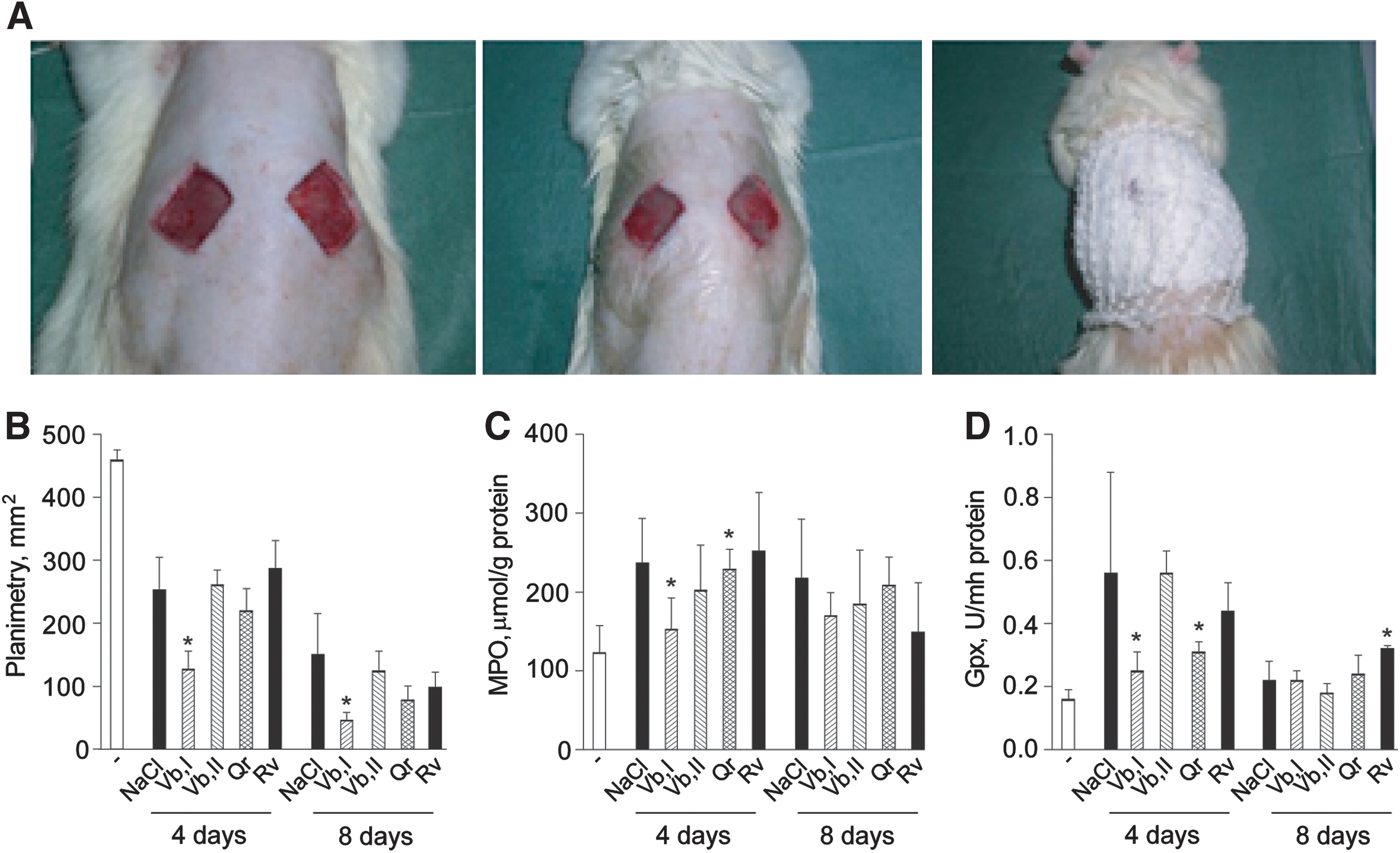
In the animal model of full-thickness excision wound (Fig. 9A), 50 μM Vb applied topically substantially accelerated wound closure (Fig. 9B) and inhibited both MPO and glutathione peroxidase (Gpx) activities in the skin biopsies closely taken to wound area (Fig. 9C, D) at the 4th postwounding day. On contrast, 100 μM Vb and 50 μM Rv or Qr did not have any of the above effects.
Discussion
Being a ubiquitously expressed, cell surface transmembrane receptor with intrinsic protein-tyrosine kinase activity (1, 24, 25, 30, 43), EGFR is highly expressed in human keratinocytes in vivo and in vitro (26, 27, 29, 43, 44). Growing evidence suggests that EGFR is a key functional regulator of keratinocyte responses to a vast array of extracellular stimuli including specific EGFR ligands, ligands for other cell surface receptors such as G-protein-coupled and cytokine receptors, and a number of nonligand agents such as H2O2, UV, heat, radiation, and some chemotherapeutics (8, 24, 25, 30). Disparate mechanisms of EGFR activation in skin cells under physiological and pathological conditions suggest that it could mediate numerous cellular functions, for example, changes of cell shape, proliferation, migration, adhesion, and inflammatory responses during wound healing (28, 30, 34, 45). The experimental data demonstrate that the EGFR system plays a relevant role in the epidermal cell reaction to wounding, first by mounting a robust, but transient inflammation (7, 26, 27, 30). Taking into account that EGFR is a redox-regulated receptor (1, 4, 18, 43), here, we attempted to elucidate the modulation of EGFR-dependent chemokine production and repair-related processes in NHEK by redox-active PPs (14). In nature, many PPs exist in glycosylated forms with sugar moieties substituting hydrogen in one of the polyphenolic hydroxyl groups. Since both number and position of OH-groups in the phenolic core determine free radical scavenging and metal chelating properties of PPs, glycosylated forms are often less effective antioxidants than corresponding aglycons (6). The antioxidant potency of PPs depends also on the oxidation model used (Fig. 1). Antioxidant properties of the selected PPs were confirmed in two physiologically relevant models of LDL oxidation by MPO+H2O2+nitrite and NADPH-induced LPO in liver microsomes. Our data on superoxide scavenging action of Qr, Rt, Rv, and Vb in xanthine oxidase-free O2- -producing system well correspond to literature data (6, 9, 15). Exclusively Vb and Qr, which were the strongest superoxide scavengers (Fig. 1), effectively inhibited constitutive (Fig. 2A) TGF-α-induced (Fig. 3A, B), and T/I-induced (Fig. 5C, D) EGFR phosphorylation. Auto-phosphorylation of EGFR tyrosine residues 1068 and 1173 important for the functional activity of the receptor is counter-balanced by receptor-type protein tyrosine phosphatase (RT-PTP) that directly de-phosphorylates it to maintain low levels of active P-EGFR (43, 44). Several publications indicate that reversible oxidative inactivation of RT-PTP takes place as a consequence of reactive oxygen species generated in response to EGF and cytokine receptor activation (1, 12, 43). We suggest that Vb and Qr could invariably inhibit EGFR phosphorylation via redox regulation of RT-PTP. Another possibility of direct inhibition of EGFR tyrosine kinase activity by Vb and Qr is less probable due to serious differences between the action of classical inhibitor of the activity P168393 and the PPs: (i) P168393 completely abrogated both EGFR and downstream ERK phosphorylation induced by TGF-α whereas PPs did not affect ERK (Fig. 3A) and (ii) P168393 and Vb/Qr effects on TGF-α-connected chemokine expression were opposite (Fig. 4). Although P168393 inhibited EGFR-dependent IL-8 gene expression, the PPs synergized with TGF-α in its overexpression. At the same time, P168393 induced overexpression of MCP-1 and IP-10, whereas Vb/Qr did not affect the chemokine gene expression and MCP-1 protein. Only Qr inhibited IP-10 synthesis.
We observed that all PPs independent of their antioxidant capacity inhibited T/I activated p65 phosphorylation (Fig. 5C, D) although the process is regarded as redox-dependent (11). In accord with our previous observation (31), Vb profoundly inhibited inflammatory responses in NHEK significantly downregulating both NFκB and AP-1 DNA binding whereas Qr mainly inhibited AP-1, and Rv less potently affected DNA binding of both transcription factors (Fig. 5E, F). Here, Vb was an effective inhibitor of T/I-induced phosphorylation of the three factors ERK, NFκB, and EGFR; it profoundly suppressed both NFκB and AP-1 DNA binding, and downregulated chemokine production (Fig. 5A, B). The PPs effects toward chemokine expression in NHEK did not depend on their superoxide scavenging and iron chelating action since SOD-mimicking compound and iron chelator exhibited no effect (Fig. 5A, B). Surprisingly, Rv and Pd at a concentration of 50 μM remarkably activated IL-8 production by nonstimulated and T/I-challenged NHEK (Figs. 2E, 2H, and 5) notwithstanding their significant inhibition of ERK at early time points. In addition, although TGF-α alone slightly induced IL-8, its combination with the stilbenes, significantly upregulated IL-8 both at the mRNA and protein levels (Fig. 4). The discrepancy between the chemokine transcript levels measured at an early time point (3 h) and the protein levels at 24 h in the supernatants in the presence of Vb (Fig. 4) suggests that the polyphenol might exert a complex, time-dependent regulation of chemokine gene expression, or that it might differentially affect the two processes of gene transcription and subsequent transcript translation. Notably, none of the PPs inhibited TGF-α-induced ERK or STAT3 phosphorylation, whereas Vb inhibited and Qr further stimulated p38 phosphorylation (Fig. 3). We have recently verified that Rv-induced IL-8 upregulation was not associated to RNA stabilization in T/I-stimulated NHEK, and that Rv-induced aryl hydrocarbon receptor significantly contributes, but not fully explains, IL-8 upregulation (33). On these grounds, we hypothesize that the intracellular mechanisms underlying stilbene-associated IL-8 upregulation cannot be detected in early time points, and that a delayed, strong activation of signal transduction should appear after hours-long contact of NHEK with these PPs.
In keeping with our previous results (26, 27, 29), we observed that the specific EGFR tyrosine kinase inhibitor PD168393 critically perturbed TGF-α-associated expression of chemokines in NHEK, with strong suppression of IL-8 and significant upregulation of MCP-1 and IP-10 (Fig. 4). Importantly, PPs opposed PD168393 inhibitory effects on the release of IL-8 (stilbenes), the mRNA synthesis and protein release of MCP-1, and the transcript of IP-10 (stilbenes and Qr). Altogether, these data indicate that the PPs can be potent modulators of the proinflammatory behavior of keratinocytes in a compound-specific way, despite the quantitative suppression of EGFR and ERK due to PD168393 (Fig. 3F, G). We also investigated the impact of the PPs on the nuclear accumulation of EGFR (44), a process upregulated by TGF-α, suppressed by PD168383, and relevantly perturbed by LB. Although LB alone did not affect the background levels of both nEGFR and nP-EGFR (Fig. 6I, J), Qr increased nP-EGFR in LB-treated cells. In the presence of TGF-α+Qr or TGF-+Vb both EGFR and P-EGFR accumulated in the nucleus (Fig. 6C, D), thus showing a contrasting effect as compared with their own significant downregulation of EGFR phosphorylation in the whole cells (Fig. 5C, D). Also, Vb and Qr were the most effective PPs in downregulating IL-8, MCP-1, and IP-10, either upregulated (IL-8 and MCP-1) or downregulated (IP-10) by LB (Fig. 7), suggesting that these two PPs specifically interfere with LB-driven inhibition of EGFR nuclear-cytoplasm export (44). LB strongly upregulated both TGF-α-associated IL-8 and MCP-1, suggesting that enhanced retention of nEGFR/nP-EGFR in the nucleus might facilitate the expression of these chemokine genes (Fig. 7). PPs exerted strong but disparate effects on (LB+TGF-α)-induced chemokine expression. Collectively, these results suggest that the differential perturbation of EGFR phosphorylation and intracellular trafficking by PPs heavily contribute to their impact on the proinflammatory potential of NHEK.
Nuclear EGFR has been strongly correlated to enhanced proliferation activity (20, 24). During the epidermal response to wounding, epidermal keratinocytes initiate a program of de novo synthesis of growth factors and autocrine production of EGFR ligands (30). Our observation that wound closure was accelerated by Vb both in vitro (Fig. 8) and in vivo (Fig. 9), could be attributed to its potentiation of the nuclear EGFR accumulation in the presence of EGFR ligands. In agreement with its capacity to inhibit EGFR-dependent IL-8, a chemoattractant for granulocytes, Vb suppressed granulocyte recruitment to the wound site at the early inflammatory stage of wound healing (MPO test) thus exhibiting anti-inflammatory properties assessed by Gpx test.
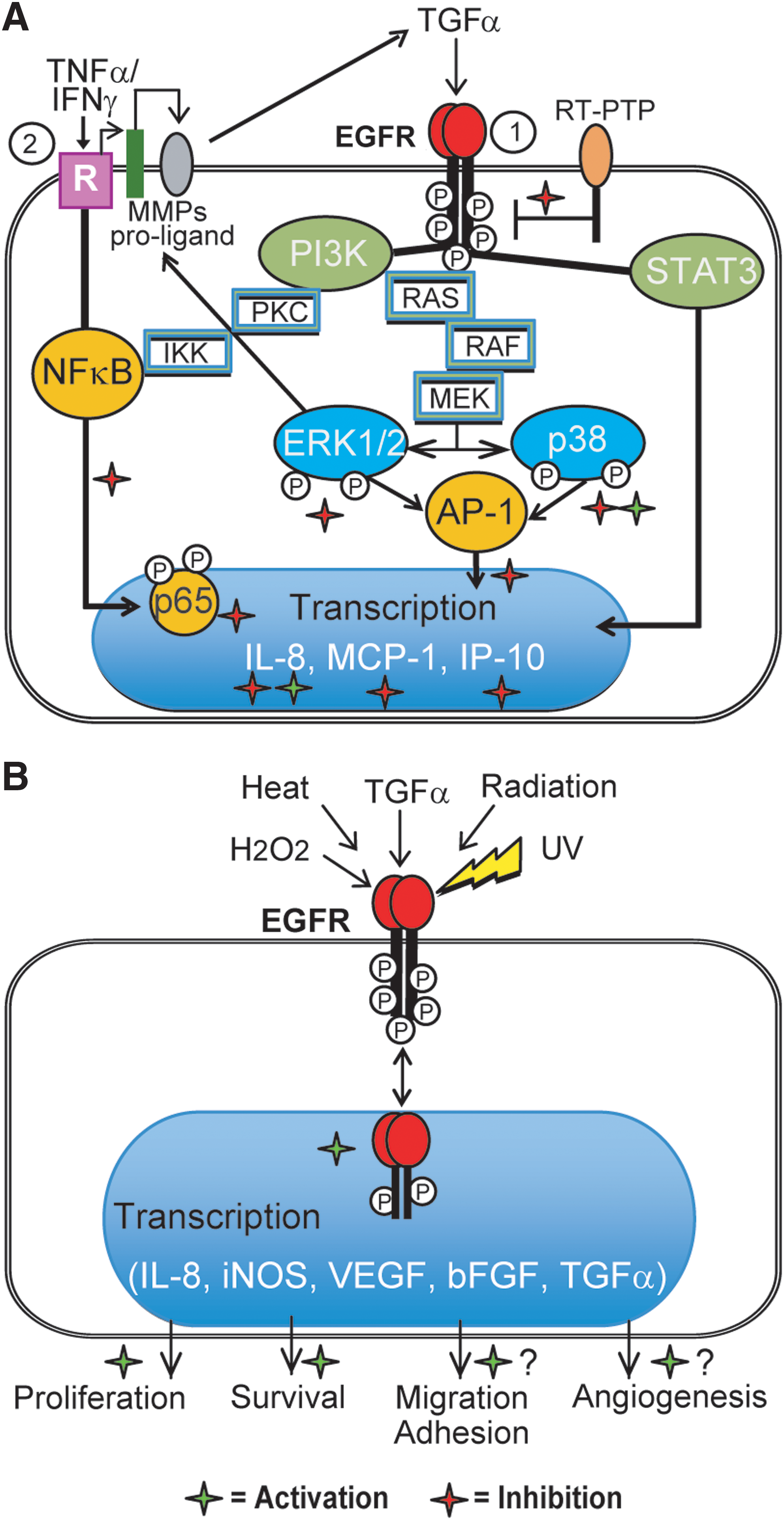
Hypothetical mechanisms of PPs effects on EGFR-driven proinflammatory and repair processes in keratinocytes are proposed in Figure 10.
Materials and Methods
Animals and excision wound model
Adult Wistar rats (300–400 g, 10 animals per group) were anesthetized with 10 mg/kg ketamine, the dorsal fur shaved, and two 21.5×21.5 mm squares were excised down to muscles (0.2 cm depth) (Fig. 9A). The wounds were daily treated with 100 μl of tested compounds or 0.9% NaCl, then, dressed to maintain moisture. Biopsies were taken in a close vicinity to wounds at days 4 and 8. Tissue samples were homogenized, centrifuged, and enzyme activities were measured in supernatants (16). Animal experiments were approved by the Local Board. The measurements of the wound areas were performed on the 4th and 8th day by planimetry.
Cell cultures and treatments
Primary cultures of NHEK were obtained from skin biopsies of healthy volunteers (n=5) after their informed consent (29). Keratinocytes were grown up to 60%–80% of confluence in serum-free medium (KGM-Gold; Lonza, Walkersville, MD). In the 24 h preceding the experiments, NHEK cultures were switched to EGF-depleted medium.
In the experiments with the in vitro scratch wounds, NHEK were grown in 6×multiwell plates. Bottom of each plate was marked with three horizontal lines. At 80% of confluence, cultures were disrupted by plastic tip drawing three vertical lines (Fig. 9) and the time-dependence of the gap between cell sheet margins at 9 cross points was digitally measured. As positive control, 10 ng/ml EGF (R&D Systems, Abingdon, UK) was used.
Quercetin dihydrate (Qr) and Rt were from Sigma-Aldrich (Milan, Italy), Rv was from Biomol (Research Lab, Plymouth, MA), Pd (95% purity; HPLC grade) was a gift from Prof. Ravagnan (University of Venice, Italy), and Vb (97% purity; HPLC grade) was a gift from Dr. Dal Toso (IRB, AltavillaVicentina, Italy). Chemical structures of tested PPs are given in Figure 1. PPs were added to cell cultures 30 min or 1 h before exposure to proinflammatory triggers. Equal volumes of dimethyl sulfoxide (a vehicle for Rv, Pd, Qr, and Rt) were added to the control cultures.
TGF-α (50 ng/ml; R&D Systems) was added to cell cultures 1 h after preincubation with PPs (50 μM) and protein phosphorylation was measured after 15 min, gene expression after 3, 12, and 24 h, whereas chemokine levels in NHEK supernatant were determined after 24 h.
In the experiments with cytokine-induced chemokine expression, NHEK were preincubated with escalating concentrations of PPs for 1 h, then, the mixture of TNF-α (100 ng/ml) and IFN-γ (100 U/ml) both from R&D Systems was added. Tetrakis(1-methyl-4-pyridyl)porphyrin pentachloride (MnTMPyP), a cell permeable superoxide dismutase mimetic and peroxynitrite scavenger (32) was a kind gift of Prof. Mollace (Pharmacology Department, Catanzaro University, Italy). Cell permeable iron chelator 1,2-dimethyl-3-hydroxypyrid-4-one (L1) (13) was a gift from Lipomed Co. (Zurich, Switzerland).
In the experiments with pharmacological impairment of EGFR, NHEK were preincubated with 2 μM P168393, a specific inhibitor of EGFR tyrosine kinase (Calbiochem, La Jolla, CA) for 30 min before treatment with PPs. Then, PPs alone (50 μM) were added for additional 30 min followed by 50 ng/ml TGF-α. The same experimental design was used with LB (Sigma-Aldrich), an inhibitor of EGFR export from the nucleus (44)
Preparation of cell extracts and immunoblotting
Total nonphosphorylated and phosphorylated EGFR, ERK1/2, and phosphorylated p65 subunit of NFκB were investigated in total cell lysates as previously described (31). Anti-EGFR antibodies were from Santa Cruz, whereas anti-ERK, and anti-p65 NFκB antibodies were from Cell Signaling Technology (Beverly, MA).
NHEK were differentially lysed to obtain nucleic extracts according to (37).
DNA binding activity of transcription factors
NFκB and AP-1 specific DNA binding activity was detected in cell nuclear lysates and quantified using the transcription factor-specific TransAM kits (Active Motif, Florence, Italy).
Assessment of antioxidant and free radical scavenging properties of PPs
Antioxidant effects of PPs were studied in LDL oxidation by MPO plus nitrite as previously described (17). Peroxidation products (LOOH) were quantified using the PeroxiDetect Kit (Sigma-Aldrich).
PPs-related inhibition of NADPH-induced LPO was evaluated in rat liver microsomes (1.2–1.3 mg protein/ml), which were incubated with 0.3 mM NADPH, 20 mM NaCl, and 10 μM FeSO4 dissolved in 0.05 M phosphate buffer, pH 7.4, at 37°C for 5 min. The content of thiobarbituric acid reactive substances was determined (41). Superoxide scavenging activity of PPs was assessed in a photochemical system reducing riboflavin and yielding superoxide as a by-product. Superoxide-dependent reduction of nitroblue tetrazolium was spectrophotometrically monitored (3). Results were expressed as concentrations of 50% inhibition (I 50, μM).
Quantitative real-time-PCR assay
Total RNA was isolated using the GenElute Mammalian Total RNA Kit (Sigma-Aldrich) and was reverse transcribed using the iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA). cDNA was amplified with IQ SYBR green Supermix (Bio-Rad) using the MiniOpticon Real-Time PCR Detection System (Bio-Rad). Two housekeeping genes, ribosomal 18S and beta-actin were chosen as reference and fold changes were calculated (23). The primer sets were synthesized by Eurofins MWG Operon (Ebersberg, Germany): β-actin fwd:5′-AATCTGGCACCACACCTTCTAC-3′; β-actin rev:5′-ATAGCACAGCCTGGATAGCAAC-3′; 18S rRNA fwd:5'- TCCCCCAACTTCTTAGAGG-3'; 18S rRNA rev:5'- GCTTATGACCCGCACTTAC-3'; MCP1 frw:5'-AAGCAGAAGTGGGTTCAGGA−3'; MCP1 rev:5'-TAAAACAGGGTGTCTGGGGA-3'; IL-8 frw:5'-GTCCTTGTTCCACTGTGCCT-3'; IL-8 rev:5'-GCTTCCACATGTCCTCACAA-3'; IP-10 frw:5′-GGGAGCAAAATCGATGCAGTGCT-3′; IP-10 rev:5′-GCAGCCTCTGTGTGGTCCATCC-3′.
Assays for inflammatory chemokine production by NHEK
The proinflammatory chemokines IP-10, MCP-1, and IL-8 were measured in cell supernatants following 24 h stimulation with PPs alone or in combination with TGF-α or (TNF-α+IFN-γ), using BD OptEIA Elisa kits from BD Biosciences (San Diego, CA) (33).
Statistics
All measurements were done in triplicate, and data of at least three independent experiments were statistically evaluated. Statistical evaluation was carried out with the software package for Windows XP. Results were expressed as the mean±SD. To evaluate the difference between experimental groups, two-tailed Student's t-test was applied and p-values<0.05 were considered to be significant.
Footnotes
Acknowledgments
This work was supported by The Italian Ministry for Health (Grant IDI IRCCS-2010). The skillful technical assistance of Dr. Giovanni Primavera and Dr. Virginia Barone in keratinocyte culturing and Dr. Michail Anurov and Dr. Svetlana Titkova in the excision wound model is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.