
Abstract
Maintenance of healthy arteries requires a balance between injuries to the arterial wall and processes of intrinsic arterial repair. Such repair requires the availability of progenitor cells that are local to the wall itself. Progenitor cells from distant reservoirs like the bone marrow may also contribute to repair. Arterial repair seems to degrade over a lifetime, particularly with risk factors such as smoking and diabetes. Hence, a potential preventive/therapeutic strategy for atherosclerosis could be transfusion of competent bone marrow cells (BMCs) to restore effective repair in the face of arterial injury and depleted endogenous repair reservoirs. The challenge with this strategy has been the reliable collection and/or generation of BMCs that support arterial repair. In this study, we describe a set of experiments to elucidate a method of culturing BMCs that robustly retards atherosclerosis development in apolipoprotein E knockout mice. Identifying such a method would represent an important step in developing cell-based treatments for patients with proclivity for developing atherosclerosis. Antioxid. Redox Signal. 16, 85–91.
Introduction
While there is a substantial body of work regarding arterial repair mediated by bone marrow-derived progenitor cells (1 –3, 5), there remains considerable controversy (6 –9). For example, work by Hagensen et al. demonstrated that progenitor cells originating from the bone marrow do not contribute to endothelial repair (3). Instead, Ergun et al. showed that progenitor cells located within the arterial wall, rather than circulating progenitor cells, were responsible for arterial repair (1). Similarly, there have been discrepancies regarding the effects of endogenous (6) and exogenously administered (7) progenitor cells on atherosclerosis. As reviewed by Zampetaki et al. (9), injections of exogenous progenitor cells have led to reductions as well as increases in atherosclerosis in the apolipoprotein E knockout (ApoE−/− ) mouse model of atherosclerosis (7). Zampetaki et al. noted that each report used different experimental conditions and methods used different cell fractions, injection frequency, number of total injections, and cell processing methodologies (9). In considering the entire body of work to date, clearly progenitor cells modulate atherosclerosis development; however, the method for obtaining reliable and robust reductions in atherosclerosis with injected progenitor cells has yet to be defined.
Innovation
Even with the development of advanced treatments modalities, atherosclerosis remains the leading cause of mortality in the industrialized world. As with other chronic diseases, cellular therapy is an intriguing new avenue of therapeutic intervention. Initial enthusiasm for using bone marrow-derived stem cells to treat atherosclerosis has been tempered over the years by conflicting results from different research groups. The findings from each group have been difficult to compare given differences in their methodologies.
A series of studies is presented here to determine the optimal experimental conditions necessary to generate BMCs that can reduce atherosclerosis in a mouse model of atherosclerosis in a robust and reliable way. From this work, a method was discovered for generating such BMCs by first culturing them in a specific combination of cytokines and growth factors. Regular injections of these cultured BMCs reduces atherosclerosis in a mouse atherosclerosis model of over 50% compared to control animals. This represents a significant next step in the application of cellular therapy for treating atherosclerosis.
Our group pioneered the concept that cultured bone marrow cells (BMCs) could reduce atherosclerosis when repeatedly injected into ApoE−/− recipients [1]. Over the past several years, we tested different experimental protocols to define the optimal conditions under which exogenously administered BMCs will reduce atherosclerosis in a robust and reproducible manner. In our original experiments, we cultured BMCs in minimum essential medium (MEM) with fetal bovine serum (FBS) and hydrocortisone to isolate an enriched pool of progenitor cells before injection (5). Injections of these cultured cells slowed disease progression in the ApoE−/− mouse model. In our current report, we show that culturing BMCs is necessary but not sufficient to produce cells capable of robust and reproducible arterial repair. Through a series of experiments, we show that the repair capacity of cultured BMCs is substantially improved in two ways: (i) utilizing a lineage negative (Lin−) enriched cell fraction and (ii) enhancing the culture media by adding selected cytokines. We examine the efficacy of cells cultured according to our prior work (5), versus Lin− BMCs that undergo the enhanced culturing process. The implication of our results is that human Lin− BMCs that are pretreated under similar culture conditions may produce cells capable of restoring arterial repair in patients with a propensity for atherosclerosis and its thromboembolic complications.
Results
Injection of fresh (noncultured) BMCs
There were six treatment groups: (i) freshly harvested Lin− BMCs, (ii) lineage positive (Lin+) BMCs, (iii) a cell subpopulation isolated on the basis of small size and paucity of organelles (simple little cells [SLC]) (Supplementary Reference S1; Supplementary Data are available online at
Injections of noncultured subpopulations of BMCs
In our prior work (5), we observed that regular injections of BMCs led to a reduction in atherosclerosis in the ApoE
Injection of whole bone marrow cultured in standard conditions, without added cytokines or growth factor
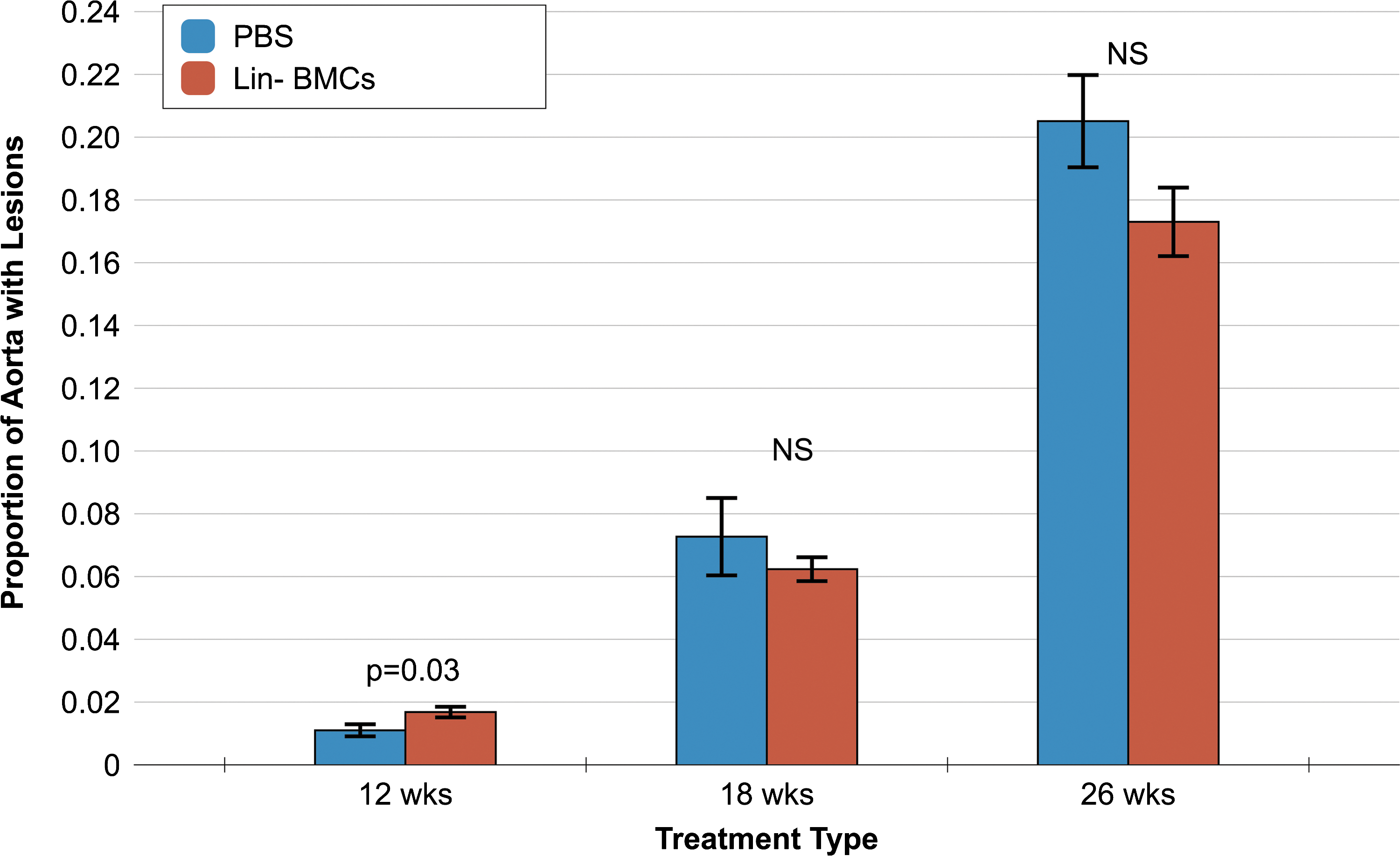
This set of studies was a close replication of experiments from our prior report (5). We injected BMCs that were cultured in MEM alpha medium with hydrocortisone and FBS for 2 days to generate an enriched population of progenitor cells. Our intention was to determine whether the culturing step was necessary and sufficient in generating a population of BMCs that could lead to a reduction in atherosclerosis. We observed that, at the early time point (after 3 injections), there was actually an increase in atherosclerosis in mice receiving the standard culture BMCs compared to PBS (Fig. 1). With increasing numbers of injections (6 and 10 injections), we observed a trend toward a significant reduction in the proportion of atherosclerotic lesions. However, the reduction in aortic atherosclerosis did not reach statistical significance.
Injection of Lin− BMCs cultured with cytokines and growth factor
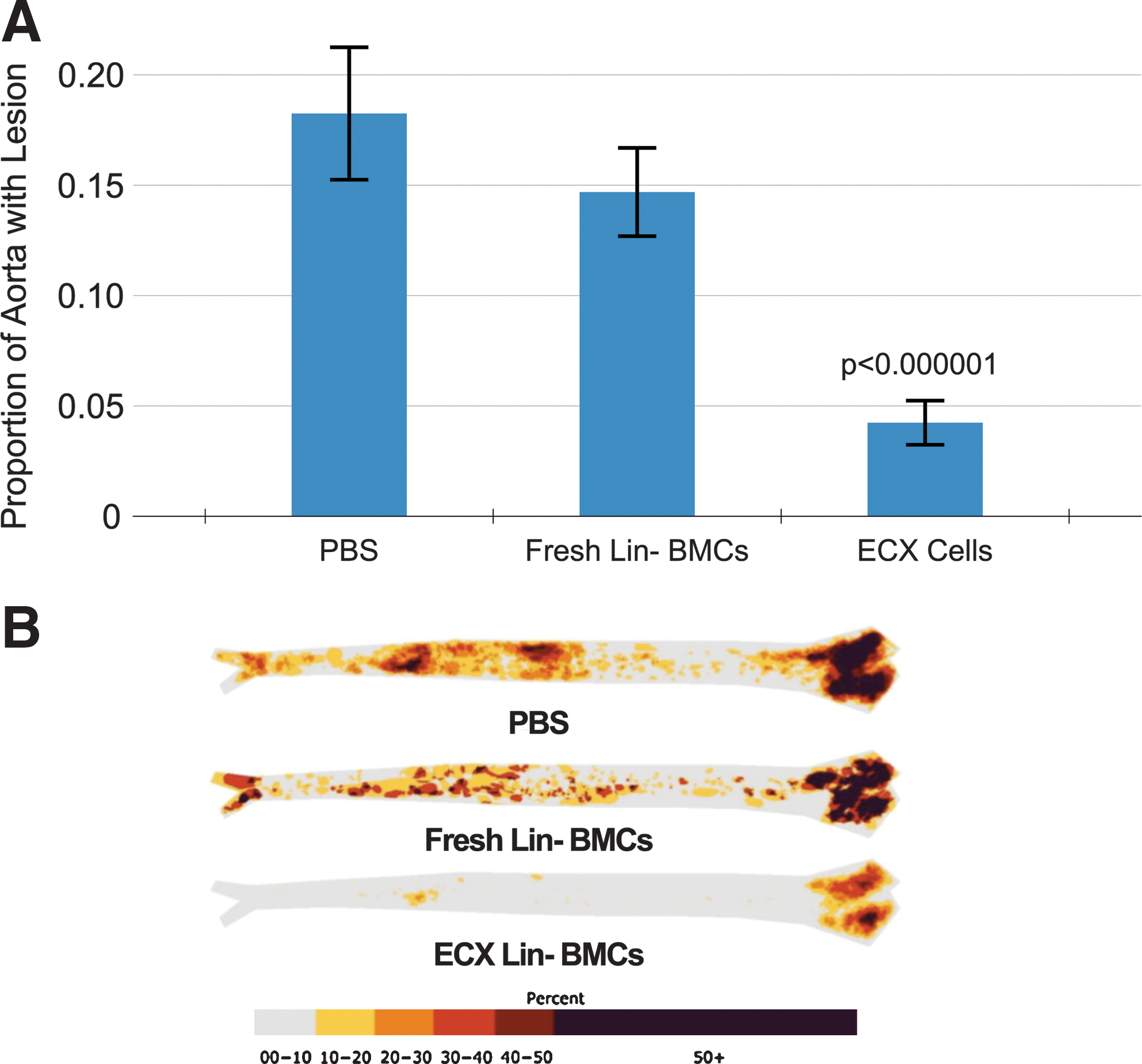
In reviewing the two sets of experiments described above, it appeared that the freshly harvested Lin− fraction had a greater effect on atherosclerosis than other cell fractions, possibly because of the enrichment for progenitor cells. Further, cultured BMCs appeared to reduce atherosclerosis more than noncultured BMCs likely through further progenitor cell enrichment as well as an unspecified transformation of the cells. Hence, in our next experiments, we combined the use of the Lin− bone marrow fraction with an enhancement of the culturing process through the addition of selected cytokines and/or growth factors. When Lin− BMCs were cultured using an enriched culture media (designated as enhanced culture [ECX] cells) with interleukins (ILs) 3 and 6 and stem cell factor (SCF), we observed a substantial reduction in atherosclerosis development (Fig. 2). The mice receiving PBS or freshly harvested Lin− BMCs injection developed atherosclerotic lesions covering 18% and 15% of their aortas, respectively. Mice receiving Lin− BMCs incubated in the enhanced culture medium only had atherosclerosis on 4% of their aortic surface (p<0.000001).
As was observed in our previously published studies, injection of any cell, including the ECX cells did not significantly alter total cholesterol, low-density lipoprotein (LDL) and high-density lipoprotein (HDL) levels (Supplementary Fig. S2).
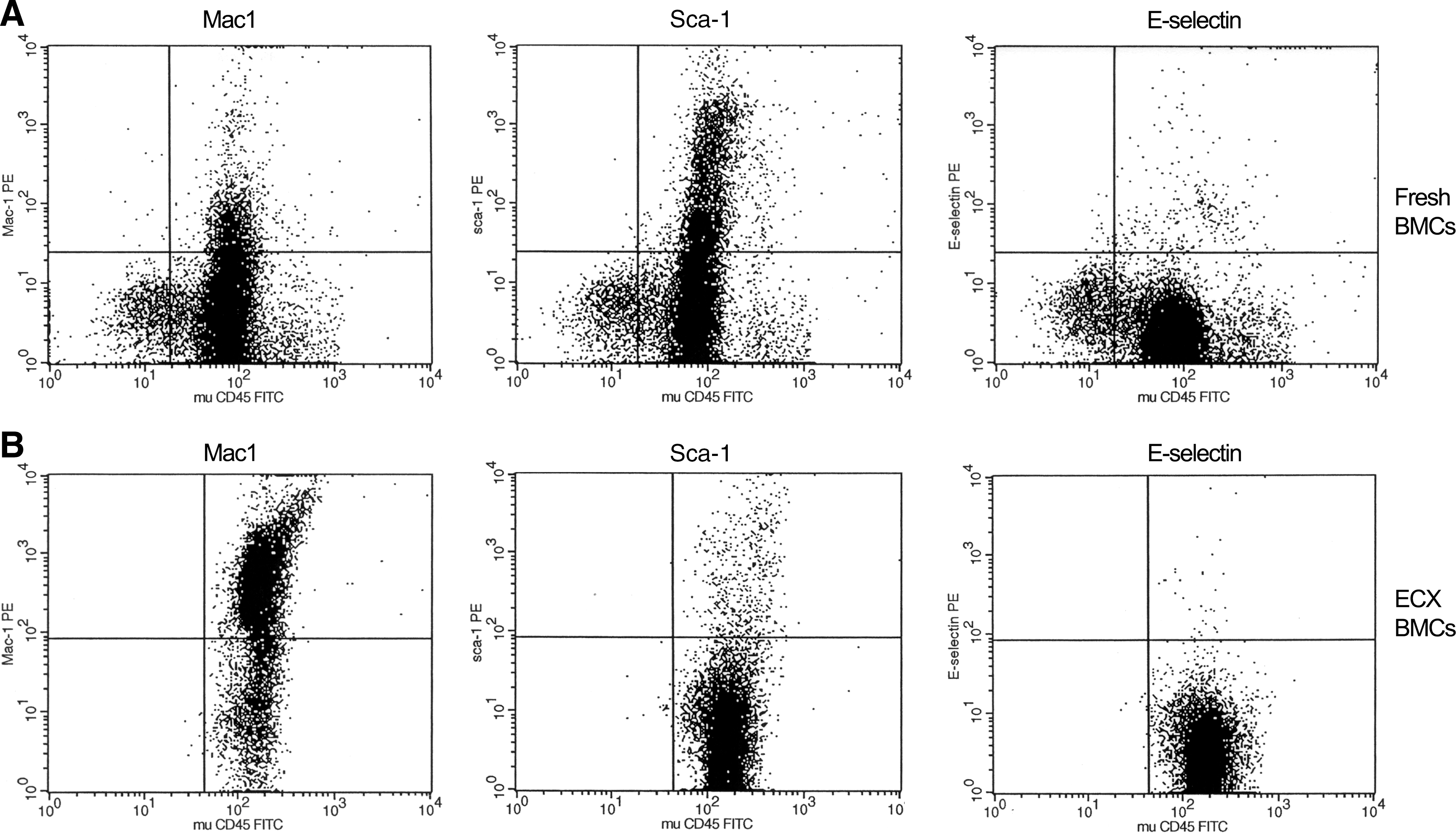
Standard flow cytometry analysis comparing ECX versus non-ECX BMCs showed that markers for all the different cell lineages were decreased in the ECX BMCs with the exception of Mac1 (see below). In particular, we observed a relative decrease in hematopoietic and endothelial progenitor cells (Sca1+ and E-selectin+ cells) in the Lin− ECX BMCs relative to fresh Lin− BMCs (Fig. 3). There was increased expression of the Mac1 marker among the ECX BMCs detected. However, in subsequent experiments injecting mice with either Lin− Mac1+ ECX BMCs versus Lin− Mac1− ECX BMCs, we found an equivalent reduction in the total area of the aorta containing atherosclerosis from 18% in the PBS control to 8.2% and 8.0%, respectively (Supplementary Fig. S3).
We explored whether the improved reduction in atherosclerosis in animals receiving Lin− ECX BMCs cells was due to factors secreted by the cells rather than the cells themselves. We compared mice that received injections of supernatant produced by ECX cells to mice that received PBS injections. Again, we found no significant reduction in atherosclerosis; in fact, we saw an increase in atherosclerosis upon injection of supernatant (Supplementary Fig. S4).
Since analysis of cell surface markers by flow was of limited benefit, we performed microarray analyses to try to identify activated or suppressed biological pathways that could explain the significant improvement of atherosclerosis prevention produced by the enhanced culture of the Lin− BMCs cells. Microarray analyses had been used previously by our group to define molecular signatures of arterial repair (4). We performed microarray analyses on (i) freshly harvested Lin− BMCs, (ii) whole bone marrow grown in standard culture, and (iii) Lin− BMCs grown in the enhanced culture. Hierarchical clustering analysis showed distinct molecular signatures resulting in the clear delineation of the three different cell groups (Fig. 4).

Gene expression analysis of freshly harvested Lin− BMCs compared to Lin− ECX cells using the statistical analysis of microarrays (SAM) tool at an false discovery rate (FDR) of <0.000001 identified 524 genes with significantly lower expression in the ECX cells relative to the freshly harvested Lin− BMCs and 877 genes with significantly higher expression in the ECX cells (Supplementary Table S1). To categorize potential biological pathways activated in the ECX cells that might explain their ability to promote arterial repair, we analyzed the candidate genes using the GeneGo Pathways Analysis Metacore Tool (Table 1).
EMT, epithelial to mesenchymal transition; IL, interleukin; PDGFR, platelet-derived growth factor receptor.
Two of the biological pathways highlighted by GeneGo analysis implicated the epithelial to mesenchymal transition (EMT) process. Among the candidate genes that are associated with EMT, hepatocyte growth factor, lymphoid enhancer-binding factor 1 (Lef1), and Cdk5r1 were differentially overexpressed in the freshly harvested Lin− BMCs compared to the Lin− BMCs ECX cells.
Quantitative real-time polymerase chain reaction (PCR) was performed to investigate the expression of the major genes of EMT described in the literature, E-cadherin, snail, and vimentin. Expression of E-cadherin was significantly higher in the Lin− ECX BMCs, whereas expression of snail and vimentin was significantly reduced in the Lin− ECX BMCs relative to freshly harvested Lin− BMCs. These findings suggest that in the Lin− ECX BMCs, the opposite process of mesenchymal to epithelial transition (MET) may be induced by the mixture of cytokines and growth factor. This suggests a possible mechanism that may lead to the enhanced repair and may warrant further study.
Discussion
We performed a series of experiments using a wide variety of cell treatments to comprehensively address the question of whether such treatments can reliably and effectively modify BMCs to reduce atherosclerosis. Our results both confirm some prior findings in the literature and also suggest a new method for significantly boosting the ability of cell injections to ameliorate atherosclerosis in the ApoE−/− mice. We found that ApoE−/− mice receiving a small number (3 or fewer) of whole culture bone marrow injections actually developed more atherosclerosis relative to control animals injected with PBS (Fig. 1). This is in line with the previous reports by Silvestre et al. (7). We found that mice receiving a greater number of whole bone marrow injections showed a trend toward atherosclerosis reduction (Fig. 1). While not reaching statistical significance, reduction in atherosclerotic lesions increased with increasing numbers of injections. These findings are in line with previous reports in the literature, including our own (1, 5) in which ApoE−/− mice receiving 6 or more cell injections showed a reduction in atherosclerosis. While the whole bone marrow injections in our investigation showed a trend toward atherosclerosis reduction, the effect was at best modest. Therefore, we sought to identify a method to produce cells that could reproducibly induce a marked reduction in atherosclerosis.
We found that a new method that pretreats Lin− BMCs by culturing them in media containing IL-3, IL-6, and SCF before injection. This pretreatment produces cells that can significantly and reproducibly reduce the development of atherosclerosis in the ApoE−/− mouse model. We recognize that the mechanism responsible for the marked reduction in atherosclerosis remains mostly uncharacterized. Certainly, it is not through a normalization of serum lipid levels (Supplementary Fig. S2). Flow cytometry analysis showed significant differences between freshly harvested Lin− BMCs and Lin− ECX cells, but ultimately these differences were not useful in explaining the augmented reparative capability the Lin− ECX cells. However, gene expression analysis showed substantial differences between fresh Lin− BMCs and Lin− ECX cells. Further, simple pathway analysis highlighted the MET as a key pathway differentiating fresh Lin− BMCs from Lin− ECX cells. The results of the gene expression analysis point toward a potential mechanistic explanation and provide direction for further refinements in using cultured BMCs for preventing and treating atherosclerosis.
Conclusion
Our study contributes to the advancement of the field in several ways, as we have developed a new method for producing cells capable of reproducible and marked reductions in atherosclerosis when infused repeatedly into ApoE−/− mice. The disease reduction occurs in the absence of improvement in serum cholesterol in this hyperlipidemic model. Potential treatments involving the infusion of BMCs pretreated under similar culture conditions could be an important addition to our current clinical armamentarium of risk reduction and revascularization strategies. Thus, in addition to reducing individual risk factors with diet, exercise, smoking cessation, statins, blood pressure medications, and other tools currently available, cell transfusion may become an additional means to suppress atherosclerosis and its thromboembollic complications.
Notes
Materials and methods
Animals
C57BL/6 ApoE−/− mice were purchased from Jackson Laboratories (B6.129P2-ApoEtm1Unc ; Bar Harbor). Pups were weaned at 3 weeks of age. Donor mice were maintained on chow diet and recipient mice were fed a Western diet (TD.88137; Harlan Teklad Madison) containing 21% fat by weight (0.15% cholesterol).
Lin− cells isolation
BMCs were harvested from ApoE−/− mice at 5–8 weeks of age by flushing the long bones (5). Lin− cells were isolated by MACS sorting using Mouse Lineage cell depletion kit (Miltenyi Biotech).
Ex vivo culture of whole BM and Lin− BMCs
For standard culture, whole bone marrow was placed into MEM alpha medium containing 20% FBS and 2 μM hydrocortisone. Cells were incubated at 37°C in 5% CO2 for 2 days. For the enhanced culture, Lin− cells were placed into DMEM containing 20% FBS, 20 ng/ml murine IL-3, 50 ng/ml murine IL-6, and 50 ng/ml murine SCF (R&D Systems). The cells were incubated at 37°C in 5% CO2 for 4 days. For both culturing approaches, the nonadherent, floating cells were collected and used for injections. Single-cell suspensions were prepared by passing the cells through a 45-μm nylon gauze and resuspended in PBS containing 2% bovine serum albumin.
Cell injections
Five sets of cell injection experiments were performed. First, freshly harvested BMCs (noncultured) from donor ApoE
Second, whole BMCs from donor ApoE
Third, Lin− BMCs from donor ApoE
Fourth, Lin− BMCs underwent enhanced culture and were separated into Lin− Mac1+ BMCs or Lin− Mac1− BMCs fractions. Recipient ApoE
Fifth, Lin− BMCs underwent enhanced culture for 4 days, after which the supernatant was collected. Recipient ApoE
Flow cytometry analysis
Phenotypic analysis of freshly harvested Lin− BMCs and Lin− BMCs cultured with cytokines and growth factor was performed using anti-mu-CD 3 PE, anti-Sca-1 PE, anti-mu-Mac-1(CD-11b) PE, anti-mu-CD45 FITC, and anti-E-selectin PE and compared to isotype control staining (Becton Dickson). The stained samples were sorted using a dual laser fluorescence-activated cell sorter (Becton Dickinson, San Jose, CA).
Real-time PCR
TaqMan Gene Expression Assay for Cdh1, Cdh2, Snail, and VIM was purchased from Applied Biosysems (ABI). First-stand cDNAs were synthesized from total RNA using TaqMan Reverse Transcription Reagents (ABI). Gene expression levels were measured by real-time PCR with an ABI/PRISM 7700 sequence detection system (ABI). The expression levels of target mRNAs relative to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was calculated using the comparative threshold cycle (CT) method (2−ΔΔCT).
Microarray analysis
Total RNA was extracted from either freshly harvested Lin− BMCs or Lin− BMCs grown in enhanced culture (IL-3, IL-6, and SCF). Probes were generated and hybridized to the murine GeneST array (Affymetrix) at the University of Miami Microarray Core facility. SAM identified candidate genes with statistically significant differential expression. Raw data underwent RMA normalization. FDR of <0.000001 was used to denote differentially expressed genes. The GeneGo MetaCore Pathways Analysis Package was used for gene annotation and identification of biological pathways that were statistically overrepresented within the candidate genes.
Quantitative assessment of atherosclerosis lesion burden
After sacrificing recipient mice, aortas were removed and formalin-fixed. The Oil Red O staining and measurements of the atherosclerosis were conducted independently by the EEHScience laboratory. The laboratory previously performed the computerized atherosclerosis quantification analysis as previously described (4). EEHScience was blinded to the cell treatment type received by each of the experimental animal groups. En face staining for atherosclerosis was done with Oil Red O. Each aorta image was projected to a reference aorta template, derived from the average size and shape of all aorta surveyed. The extent of disease was quantified by counting pixels staining with Oil Red O compared with the total area of the aorta. EEHScience performed the statistical analysis for each of the animal groups.
Footnotes
Acknowledgments
The work was supported by funding from the National Institutes of Health Grant R01 HL71536 through the National Institute for Aging and the National Institutes of Heath Grant K12RR17630 through Duke University.
Author Contributions
X.S, P.J.G., and D.S. discovered and developed the enhanced culturing strategy. D.S. designed and supervised the overall project. K.W. supervised certain portions of the mouse experiments. X.S., Q.M., and X.L. performed the mouse studies described. E.H. performed the disease mapping. P.S. and E.H. performed the statistical analyses. D.S. performed the microarray analyses. D.S. and P.J.G. were the primary writers and P.S. performed substantial editing.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.