
Abstract
Introduction
A complex interplay of reactive oxygen species (ROS) generation and genes that protect against ROS damage has been implicated in the onset and progression of numerous aging-associated disease states, including type-2 diabetes, solid cancers, Alzheimer's dementia, osteoporosis, myocardial infarction, and cerebrovascular accidents (45). Blockade of insulin/IGF-1 signaling has beneficial effects: it increases lifespan, confers resistance to oxidative stress, and reduces or postpones many age-associated conditions and diseases (2, 8, 17, 38). How could different stress states, including those mediated by ROS, oxidants and other electrophiles, contribute to aging and explain the age-dependent incidence of diverse diseases? A possible shared mechanism, increasingly invoked in both geriatric and gerontologic contexts, is inflammation (13).
Innovation
Aspirin is a versatile drug that confers protection against multiple age-associated diseases, including atherosclerosis, diabetes, and a variety of cancers. Evidence in mice and humans suggests that aspirin modestly reduces all-cause mortality, but it is unknown whether that reduction is merely the sum of the disease-specific benefits or a more direct amelioration of aging itself. Our observations demonstrate for the first time that aspirin slows many measures of aging per se, and provides evidence that insulin-like signaling and antioxidant defenses are involved. The nematode model is a particularly suitable one in which to establish the mechanisms underlying aspirin's diverse benefits.
Aspirin (acetylsalicylic acid), a prototypic cyclooxygenase inhibitor, is a widely used analgesic agent which opposes inflammation and platelet aggregation. It regulates the activity of a number of pro-inflammatory signaling molecules, such as TGF-β (37) and PDGF (48). In addition to its anti-inflammatory effect mediated through cyclooxygenase inhibition, the salicylate moiety in aspirin also reduces the generation of pro-oxidant species (28). In keeping with these pharmacologic effects, aspirin reduces the severity of aging-related endothelial dysfunction in mice (7), modestly lowers blood pressure in hypertensive mice (26), and reduces cardiovascular events in both primary and secondary prevention trials in humans (11). High-dose aspirin was reported to improve glucose metabolism and reduce fatty acid levels in patients with type-2 diabetes (20). Aspirin also extends the lifespan of male mice (43), and significantly reduces all-cause mortality in patients with type-2 diabetes (35). The mechanisms by which aspirin increases longevity, however, remain unclear.
The present study was designed to assess the effects of aspirin on endogenous ROS levels and regulation of antioxidant genes in a simple model system, Caenorhabditis elegans. Superoxide dismutases (SODs) convert superoxide into oxygen and peroxide, which in turn is broken down by catalases to molecular oxygen and water. Peroxide can trigger quite harmful free-radical chain reactions in cell lipids, and thus is likely to be the main source of oxidative damage associated with aging (41), opposed chiefly by certain glutathione S-transferases (GSTs) (50). We now demonstrate protective effects of aspirin, accompanied by induction of antioxidant enzymes (SODs, catalases, and lipoperoxidation-specific GSTs). Moreover, we document its ability to attenuate aging-associated functional declines and to extend lifespan in the nematode, largely dependent on the FOXO transcription factor that mediates most downstream effects of insulin-like signaling.
Results
Aspirin and salicylate reduce endogenous oxidative stress, and upregulate antioxidant enzymes and phase-2 detoxification enzymes
In vivo steady-state ROS levels can be inferred from fluorogenic activation of dichlorodihydrofluorescein diacetate (H2DCF-DA, Invitrogen/Life Technologies), provided that the intensity and duration of excitation (incident light) are held rigorously constant (10). Total H2DCF-DA fluorescence was reduced ∼40% by treating worms with either 1 mM aspirin or 1 mM salicylate (each p<0.001 vs. untreated controls; Fig. 1A and 1B). H2DCF-DA responds most sensitively to hydroxyl radical (

Because neither aspirin nor salicylate is itself an antioxidant, we sought mechanisms by which they might reduce ROS abundance. Transcript levels were measured for three distinct classes of antioxidant enzymes: SODs, catalases, and GSTs. With one exception, all were induced by aspirin and to a lesser extent by salicylate, each at 1 mM concentration. Of the genes encoding SODs (Fig. 1C), the most strongly induced (1.6-fold by salicylate; 2.7-fold by aspirin, p<0.0003) was sod-3, encoding an Fe/Mn-SOD believed to be localized to mitochondria. Sod-4, encoding an extracellular Cu/Zn-SOD, was downregulated 3.5-fold by 1 mM aspirin (p<10−4), while sod-5 transcripts were increased ∼20% (Supplementary Fig. S1; supplementary data are available online at
Consistent with transcriptional regulation via DAF-16, the most affected antioxidant genes are believed to be negatively regulated by insulin/IGF-1 signaling, and show increased expression in long-lived daf-2 mutants in which the insulin-like receptor is disrupted (18). The most affected gst genes, gst-5 and gst-10, were the only two of 44 genes tested that extend lifespan when disrupted (3). We examined the expression of GFP-reporter constructs driven by promoters for gst-4 and gst-10, encoding GSTs that protect against lipoperoxidation; their expression is also upregulated in daf-2 mutants (27), although chiefly via the SKN-1 transcription factor that is closely intertwined with insulin-like signaling (34). Expression of Pgst-4::GFP and Pgst-10::GFP increased at least 3.5-fold in aspirin-treated worms relative to untreated controls (Fig. 2; each p<0.0001), corroborating aspirin induction of antioxidant defenses. Expression of the Psod-3:: GFP reporter in transgenic worms was increased at least 1.6-fold by aspirin (Fig. 2, bottom panels), somewhat less than the upregulation observed at the transcript level by RT-PCR (Fig. 1C).
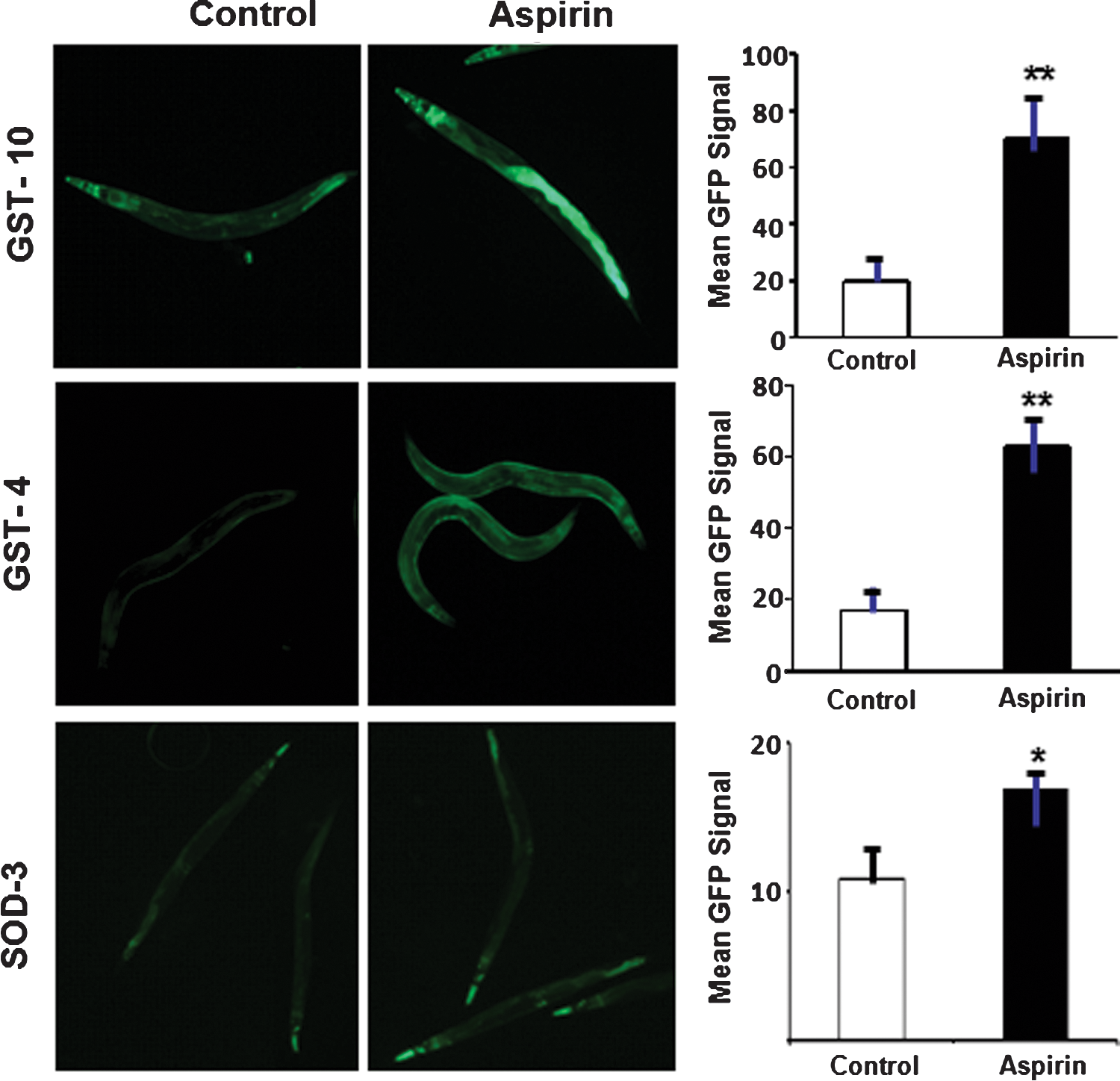
Aspirin improves resistance to an oxidative stress and extends C. elegans lifespan
Wild-type C. elegans were exposed to aspirin or salicylate from the last larval stage (L4, 2 days after hatching from eggs), and transferred on day 5 post-hatch to liquid medium containing a toxic level (5 mM) of H2O2. As shown in Figure 3A, aspirin and to a lesser extent salicylate (each at 1 mM) significantly extended survival of wild-type (N2) nematodes in the presence of a toxic level of H2O2 (p<0.001 and p<0.01, respectively).
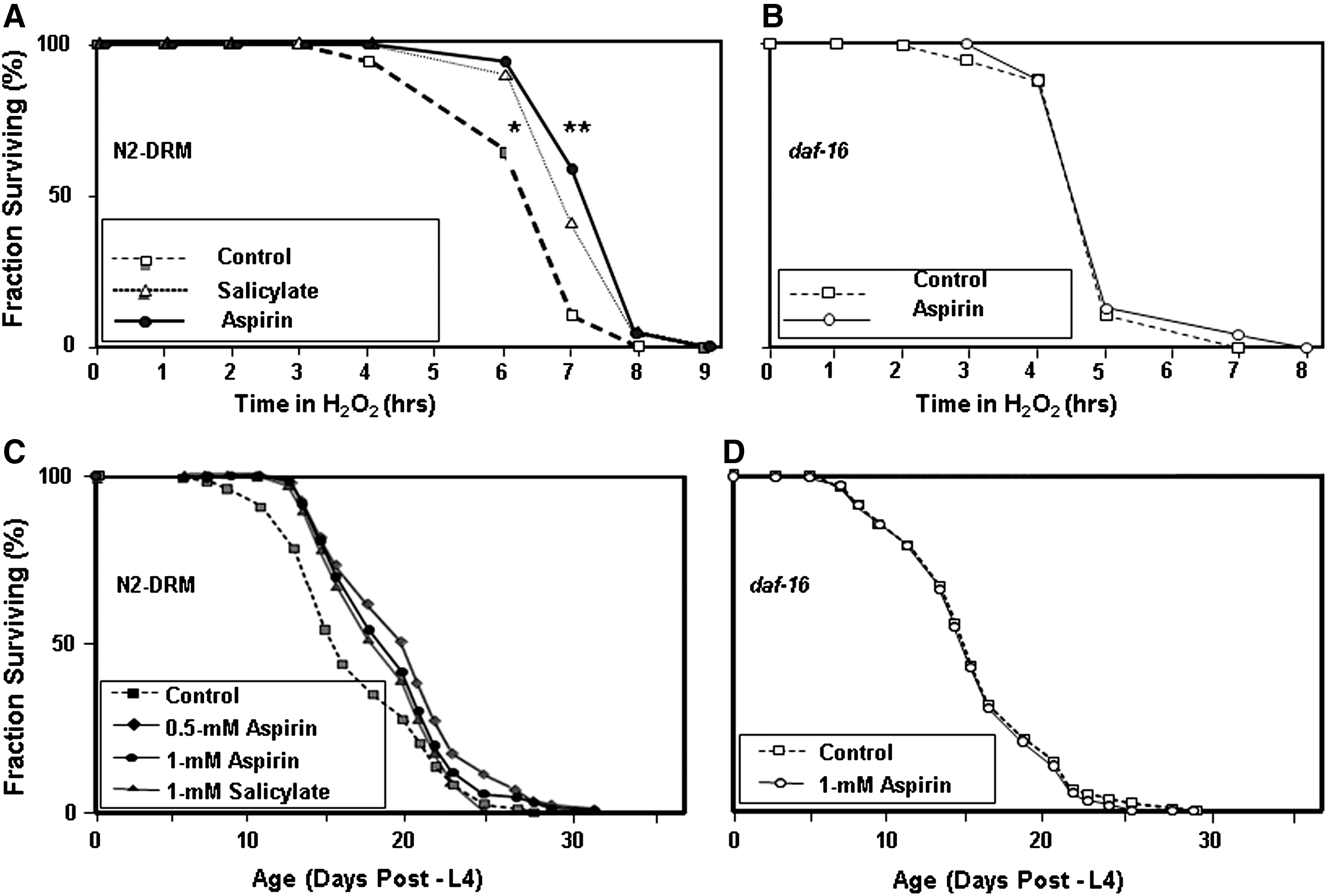
Because stress resistance can provide a surrogate biomarker for longevity (22, 29, 40 –42), we asked whether aspirin might also extend nematode lifespan. Aspirin (0.5 or 1.0 mM) increased the lifespan of wild-type worms by 12%–30% in three experiments (each p<0.001, data summarized in Table 1). A typical set of survivals is shown in Figure 3C; overall, the weighted average life extensions by 0.5 and 1.0 mM aspirin were 23% and 21%, respectively, with a combined p value of <10−9 at 1 mM (Table 1). The salicylate moiety of aspirin, at 1 mM, also enhanced C. elegans lifespan in three experiments (e.g., Fig. 3C), producing an average extension of 14% (combined p<10−7; Table 1).
Mean adult lifespans are expressed in days after the L4/adult molt. Total lifespans are approximately 2.5 days longer (the developmental interval from egg-lay to adult) at 20°C. The mean increase (last 4 rows) has been weighted by the number of worms per experiment.
Significance of survival differences was ascertained by Gehans-Wilcoxon log-rank test, comparing treatment groups to untreated controls. The composite p value combining multiple independent experiments is the product of p values observed in the individual experiments.
Insulin/IGF-1 signaling has been implicated in aging of C. elegans, results replicated in Drosophila and mice (14), and is almost entirely dependent on the DAF-16/FOXO transcription factor (25). We assessed whether lifespan extension and stress resistance conferred by aspirin also require DAF-16. As shown in Figure 3B and 3D (data summarized in Table 1), aspirin had no detectable effect on either survival of H2O2 stress, or lifespan, for a C. elegans mutant in which the daf-16 gene was severely disrupted, implying that these salutary effects of aspirin depend, largely or entirely, on the DAF-16/FOXO transcription factor.
Aspirin delays age-associated functional declines in C. elegans
Aging is associated with a progressive decline in many physiological functions, such as spontaneous movement and pharyngeal pumping in the nematode (16, 19). As shown in Figure 4A, the age-dependent decline in pharyngeal pumping was reduced or delayed by 1 mM aspirin or salicylate (each p<0.001 on day 6, and p<0.05 on day 10 of adulthood). In this respect, aspirin- and salicylate-treated worms appeared physiologically “younger” than the untreated controls. Since food intake was initially unchanged (at the first assay, on adult day 2), and was subsequently higher in the treated worms than in controls, it can be surmised that aspirin and salicylate do not exert their salutary effects via dietary restriction (i.e., by suppressing food intake).
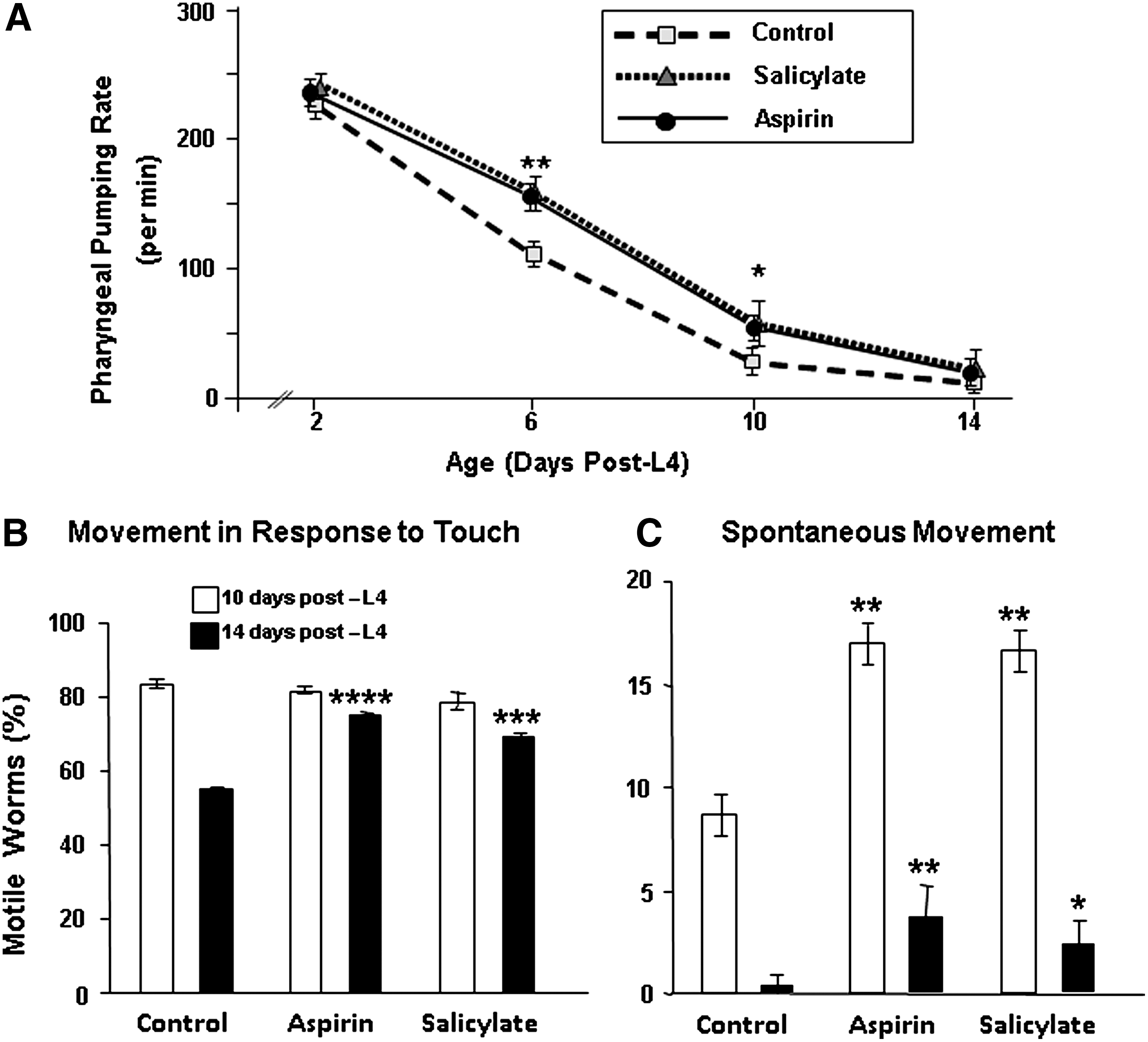
A progressive decline with age in C. elegans motility, both spontaneous and in response to exogenous stimuli, has been well documented (16, 19). Continuous exposure to either aspirin or salicylate (each at 1 mM) significantly inhibited the age-related decline in motility. The fraction of untreated worms moving in response to touch (Fig. 4B) fell 30% between 10 and 14 days of adult age (Chi-squared p<10−6); in contrast, the decline was <3% in aspirin- or salicylate- treated worms, not significantly changed from day 10 (open bars in Fig. 4B). At 14 days, however, ASA- and salicylate-treated worms showed significantly more motility than untreated controls (solid bars; P<10−6 and 0.0005, respectively). Spontaneous movement was relatively infrequent at adult day 10 (∼8.6%), and declined to 0.3% by day 14 (Fig. 4C, Control bars). Treatment with aspirin or salicylate doubled the spontaneous movement frequency at the earlier time point (Fig. 4C, open bars; each p<0.005), and raised the 14-day value (filled bars) from 0.3% to 3.7% (p<0.005) and 2.3% (p<0.05), respectively. Both the pumping rate and motility data suggest that aspirin and salicylate slow the decline in function that accompanies physiological aging.
Aspirin reduces aging-associated aggregation of proteins in C. elegans
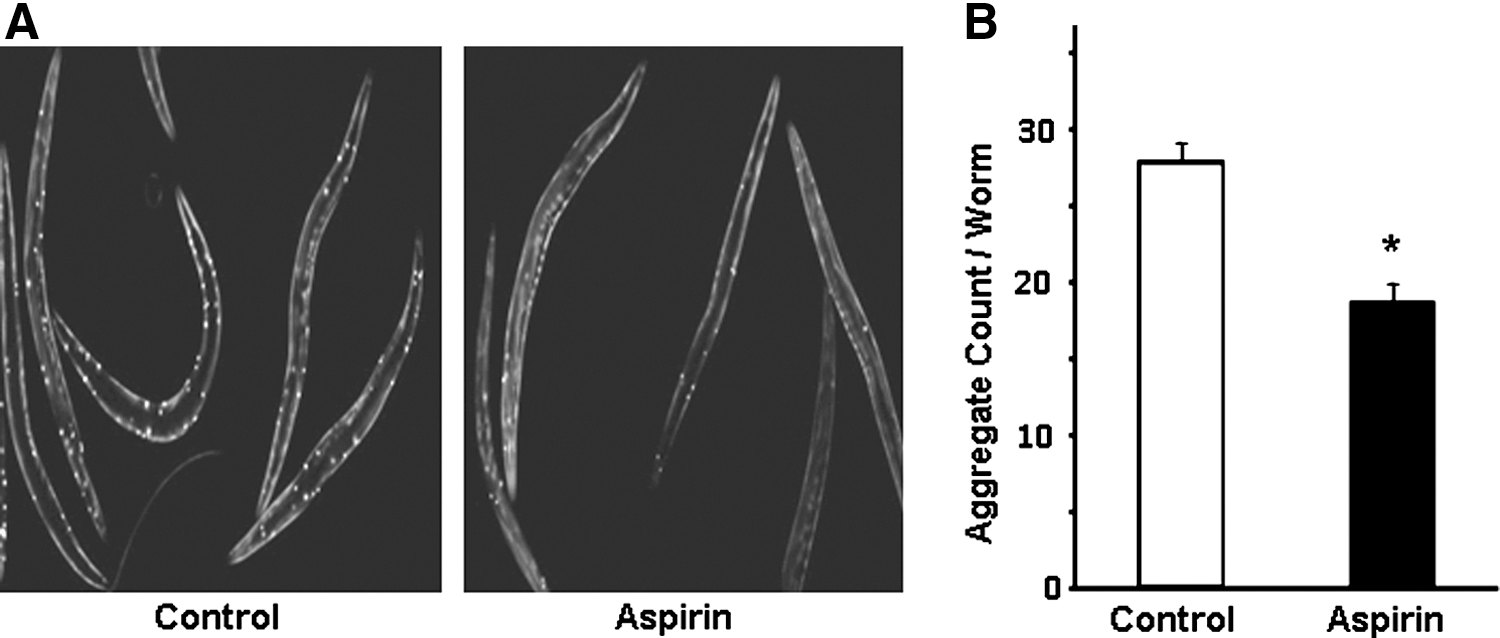
Aggregation of proteins, a characteristic of many progressive neuropathies, increases with age but is slowed by mutations that extend lifespan, and may thus provide a robust molecular biomarker of aging (6, 29). We assessed a transgenic strain expressing a fluorescence-tagged polyglutamine repeat (Q40::YFP) in body wall muscle (6). Such strains undergo age-dependent aggregate formation, as evidenced by a highly punctate pattern of yellow fluorescence in body wall muscle, commencing at day 3 after hatch (adult day 1). Aspirin reduced both the size and number of Q40::YFP aggregates observed at adult day 4, each by >30% (p<0.0001) (Fig. 5).
Discussion
Oxidative stress generally elicits an inflammatory response that, under favorable conditions, initiates tissue repair and restoration of a normal physiological state. However, in senescence, excessive oxidative stress is not adequately controlled by endogenous antioxidant reserves and may produce a state of chronic inflammation (13, 47). Aspirin, a very widely used drug, is a potent anti-inflammatory agent. We have shown that this drug, and to a somewhat lesser extent its salicylate moiety, can reduce age-associated oxidative stress and attenuate insulin/IGF-1 signaling (based on known, antioxidant gene targets of this pathway) in C. elegans. Both these agents also delay several measures of physiological aging and extend nematode lifespan.
One shared precept of various “oxidative stress theories of aging” is that endogenous ROS species, including superoxide and peroxide radical, either increase with age or are less effectively offset by antioxidant defenses, resulting in many aging-associated declines in tissue functions (42). In the present studies, we noted that both aspirin and salicylate attenuate, by >40%, total signal reported by the fluorogenic-dye H2DCF-DA as a measure of endogenous ROS levels (10). To evaluate the balance between ROS generation and subsequent elimination by antioxidant enzymes, we also measured the transcript levels of key “longevity-assurance genes” encoding 3 catalases, 5 SODs, and 5 GSTs. We found that aspirin, and to a lesser extent salicylate, increased transcript levels for nearly all of the tested antioxidant genes, with the greatest effect (>2.5-fold for aspirin) on sod-3 and ctl-2. In keeping with these observations, we found that both aspirin and salicylate improved survival in the presence of exogenous H2O2 (Fig. 3A). A curious exception to this pattern is presented by sod-4, which was downregulated more than 3-fold by aspirin (Supplementary Fig. S1). It was previously reported that deletion of the sod-4 gene, which encodes an extracellular SOD and contributes ∼5% of total sod transcripts (46), has no effect on lifespan or resistance to several oxidative stresses in a wild-type background, but further increases lifespan (especially maximal lifespan) in the long-lived daf-2 background (12). Since aspirin appears to reduce insulin-like signaling, and thus resembles a daf-2 hypomorphic mutation, it is likely that aspirin-mediated suppression of sod-4 would serve to promote rather than impair longevity.
Peroxide is a potent inducer of lipid peroxidation, which poses the greatest oxidative threat to survival because lipoperoxides (unlike other ROS molecules) are sufficiently long-lived to create free-radical chain reactions in vivo (50). The two nematode glutathionyl S-transferases with highest activity against lipoperoxides are GST-4 and GST-10 (3). It is thus particularly significant that protein-level expression of reporters for these GST's was increased >3-fold by aspirin treatment (Fig. 2). Considering that vulnerability to lipid peroxidation is one of the strongest predictors of nematode longevity (41), aspirin's ability to strongly induce these enzymes may contribute to its extension of nematode lifespan.
The GSTs are phase-2 detoxification genes, which have been proposed to play key roles in longevity assurance (21, 27). In addition to GSTs, other phase-1 and phase-2 enzymes mediating resistance to endogenous and exogenous toxicants (which include but extend beyond ROS) comprise the catalases, SODs, cytochromes P450, short-chain dehydrogenases/reductases, and UDP-glucuronosyltransferases (UGTs) (33). These genes are broadly upregulated in long-lived C. elegans mutants including daf-2, age-1 and clk-1, which in particular overexpress sod-3 and ctl-2 (32), the two antioxidant genes most highly induced by aspirin and salicylate treatments (Fig. 1C and 1E), and reported to be essential for daf-16-mediated innate resistance to bacterial infection (9). Taken together, the evidence that aspirin treatment of daf-16 mutant worms enhanced neither peroxide resistance nor lifespan (Fig. 3B and 3D), and failed to induce sod-3 and ctl-2 (Fig. 1D and 1F), implies that signaling through DAF-16/FOXO mediates most or all of aspirin's beneficial effects.
Both aspirin and salicylate slowed the age-dependent declines in motility and pharyngeal pumping, two of the most reliable physiological biomarkers of C. elegans aging (19, 24). Protein aggregation, due to oxidation, misfolding, and unstructured/nonpolar interactions between polyglutamine tracts, underlies aging-related neurodegenerative disorders and possibly many other age-dependent traits (30). Drugs that interfere with protein aggregation protect protein homeostasis, and can extend lifespan (1), and clinical trials indicate a substantial protective effect of long-term aspirin treatment against Alzheimer's dementia, which involves at least two types of protein aggregates (44). Using a nematode model of polyglutamine aggregation, we were able to demonstrate that aspirin indeed reduces aggregate size by >2-fold and the number of aggregates by >30% (Fig. 5). In view of the known association of protein aggregation with both oxidation and aging (29), it is likely that the ability of aspirin to lower ROS levels and induce antioxidant defenses, demonstrated in the present study, is linked to inhibition of protein aggregation and extension of lifespan of the nematode.
The observation of C. elegans life-span extension by aspirin is consistent with the more modest increase in longevity observed for aspirin-treated male mice (43). In human subjects, aspirin ameliorated clinical parameters associated with type-2 diabetes (20), and moderately reduced hypertension (26). At therapeutic concentrations, aspirin is a potent anti-angiogenic agent and inhibits atherosclerotic plaque formation (23). Moreover, aspirin prevents restenosis after vascular injury, reduces the frequency of recurrent myocardial infarctions and strokes, and reduces or delays the development and progression of several cancers in humans (39). One plausible explanation for the remarkable versatility of aspirin in conferring protection against such a wide assortment of age-dependent diseases is that aspirin opposes insulin/IGF signaling, a pathway that contributes to aging and age-associated diseases (22), and/or opposes other pathways that converge on the DAF-16/FOXO transcription factor (49). The 21%–23% mean life extension observed in aspirin-treated nematodes, dependent on functional DAF-16, constitutes the strongest evidence to date for such a general, anti-aging effect. Since salicylate conferred benefits similar to those of aspirin, but by most measures was less effective, it appears that the beneficial effects of aspirin described here are mediated largely, but not entirely, by the salicylate moiety.
“Oxidative damage” theories of aging attribute its ravages to cell damage inflicted by ROS. Such theories have fallen into disfavor of late, due to the inability to forestall aging with genetic or drug interventions that demonstrably reduce markers of oxidative damage (12, 36, 46). Although cogently argued, these critiques challenge a rather narrow and outdated version of such theories. They thus fail to take into account the remarkable redundancy and compensatory ability of antioxidant defenses (4), and the positive role of oxygen free radicals in inflammatory signaling. Thus, an intervention that defeats any one antioxidant effector could induce other overlapping defenses, or might thwart inflammatory reactions that are critical for defense against pathogens (50).
Materials and Methods
Strains
Nematode strains, supplied by the Caenorhabditis Genetics Center (CGC, Minneapolis), or derived in our laboratory from CGC strains, were maintained at 20°C on 0.6% peptone NGM-agar plates seeded with E. coli strain OP50, as described earlier (2). Strains employed in these studies were Bristol-N2 (wild-type) and CF1038 bearing a daf-16(mu86) allele in which most of the exons encoding DAF-16 are deleted.
Determination of lifespan
Nematodes, grown on NGM-agar plates containing 0.6% peptone, were harvested, and eggs were isolated by alkaline hypochlorite with 0.5 N NaOH, 1.05% hypochlorite; 5 min at 20°C (2). The recovered eggs were rinsed in S Buffer and placed on fresh agar plates seeded with E. coli strain OP50. Survival cultures were established on 60-mm agar plates; just after the L4/adult molt, 50 adults were transferred to 60-mm dishes containing aspirin, salicylic acid, or ethanol (solvent control, 5% [w/v] final concentration) at the indicated concentrations. Worms were maintained at 20±0.5°C and live worms counted during daily transfer to fresh dishes; worms not moving, either spontaneously or in response to touch, were scored as dead. Worms were maintained until death.
Hydrogen peroxide stress response
Adult worms (N2 or daf-16) were synchronized by alkaline hypochlorite lysis, and then the eggs were rinsed and transferred to fresh NGM-agar plates. On reaching the L4 stage, worms were placed on fresh NGM-agar plates seeded with E. coli strain OP50 and overlaid with 1 mM aspirin in ethanol, or ethanol alone (control). After 48 h, they were transferred to 24-well plates (20–25 worms per well) containing S Medium (S Buffer plus 0.5% cholesterol) and 5 mM hydrogen peroxide (Sigma) at 20°C, as previously described (2, 3). Survival was scored at 1-hour intervals, as above.
DCF assay for endogenous hydrogen peroxide
Endogenous ROS levels were indirectly quantified using dichlorodihydrofluorescein diacetate, H2DCF-DA (Invitrogen), which is deacetylated and retained after uptake into cells. In turn, H2-DCF reacts with endogenous ROS to generate the fluorescent dye, dichlorofluorescein (DCF). Live C. elegans nematodes were treated at day 4 post-hatch (1.5 days after the L4/adult molt) with 10 μM H2DCF-DA for 30 min. Fluorescence intensity was imaged using an epifluorescence microscope (Olympus BX151) with 488-nm incident light, recording light emission at 520±20 nm with a QICAM camera. The intensity and duration of incident-light exposure was kept constant, including the extent of prior light exposure which might introduce photobleaching artefacts. Fluorescence intensity, recorded through a narrow-band-pass emission filter, was quantified from digital images using imageJ software.
Fluorescence measurement of GFP-reporter strains
A confocal epifluorescence microscope (Olympus model BX151) was used to image strains carrying Pgst-10::GFP or Pgst-4::GFP transgenes, with 390-nm excitation light, recording green epifluorescence at 510±20 nm. For each reporter strain, the same image-capture settings were utilized for treated and untreated control worms, using a QICAM. Autofluorescence was measured for non-GFP (background-control) worms of the same age, and subtracted from each image. Total GFP fluorescence per worm was analyzed as above, using imageJ software.
Locomotion, pharyngeal pumping, and protein aggregate measurement
Locomotory movement, either spontaneous or in response to touch, was measured for synchronized wild-type N2 cultures as described previously (16, 19). To assess protein aggregation, AM141 worms expressing a muscle-specific fusion protein, polyglutamine (Q40)::YFP, were synchronized and grown at 20°C as above. From the L4/adult interface onward, treated worms were maintained in the presence of aspirin, salicylate, or the ethanol solvent used to introduce those drugs to plates. Epifluorescence images were captured as above. Each worm was analyzed using imageJ software, and aggregates were counted in at least 30 animals per group.
Statistical analysis
Survival and stress test experiments were repeated at least three times. Median survival (either lifespan or survival of peroxide stress) was calculated from the fitted Gompertz function at 50% survival, using NCSS software (Number Cruncher Statistical Systems, Kaysville, UT). Significance of survival differences was determined by the Gehans-Wilcoxon log-rank test, a nonparametric measure that assesses differences in entire survival curves. For comparisons of proportions (e.g., for motility assays), the chi-squared test was utilized. Other comparisons between groups involved Student's t test for larger samples (N≥10), or the Behrens-Fisher version of the t test appropriate to small samples in which equality of variances could not be established. P values≤0.05 are reported as nominally significant in two-group comparisons, but sufficient information is provided for stringent adjustments in threshold to be made for multiple endpoint comparisons.
Footnotes
Acknowledgments
This study was supported in part by funds from the Department of Veterans Affairs, Veterans Health Administration, Office of Research and Development, Biomedical Laboratory Research and Development, Washington, DC. The contents of this article do not represent the views of the Department of Veterans Affairs or the United States Government.
Author Disclosure Statement
None of the authors declare any conflict of financial interest.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.