
Abstract
Introduction
Butein (3,4,2′,4′-tetrahydroxychalone, Fig. 1), a plant polyphenol, is one of the major biologically active components of the stem bark of cashews (Semecarpus anacardium), the heartwood of Dalbergia odorifera, and the traditional Chinese and Tibetan medicinal herbs Caragana jubata and Rhus verniciflua Stokes. It has been traditionally used for the treatment of pain, thrombotic disease, gastritis, stomach cancer, and parasitic infections (12, 22) in Korea, Japan, and China. It has also long been used as a food additive in Korea (22). Butein reportedly had protective effects in human leukemia (27), hepatoma (25, 26), and breast (21) cancer cells. Butein has been shown to be a specific protein tyrosine kinase inhibitor by causing inhibition of epidermal growth factor (EGF)-stimulated auto-phosphotyrosine level of EGF receptor in HepG2 cells (37). Butein suppressed the growth, induced cell death, inhibit cell proliferation, and induce apoptosis in B16 melanoma cells. (10). It was also found to inhibit the clonogenic growth of small numbers of breast cancer cells co-cultured with fibroblasts in-vitro (31). Butein inhibited migration and invasion through the extracellular signal-regulated kinase and nuclear factor-kappa B (NF-κB) signaling pathways in human bladder cancer cells, and this inhibitory effect was associated with a reversal of epithelial-mesenchymal transition (38). Treatment with butein sensitized tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-resistant human leukemia U937 cells to TRAIL-induced apoptosis (19). Butein was reported to suppress constitutive and inducible signal transducer and activator of transcription (STAT)-3 activation and STAT3-regulated gene products in multiple myeloma cells (29) and hepatocellular carcinoma (HCC) cells. It also inhibited the growth of human HCC xenograft tumors in male athymic nude mice (30). We hypothesized that butein may act as a chemopreventive and/or chemotherapeutic agent against PCa. We report here that butein inhibited phosphatidylinositol 3-kinase (PI3K)/Akt/NF-κB signaling, induced apoptosis in human PCa cells and inhibited tumor growth in athymic nude mice implanted with human PCa cells.

Innovation
The conventional treatments for PCa are associated with significant mortality and side-effects. In the present study, we demonstrated that an antioxidant butein caused decrease in the viability of PCa cells, induced apoptosis, inhibited signaling pathways in human PCa cells and resulted in significant inhibition of tumor growth in athymic nude mice implanted with human PCa cells with decrease in the serum prostate-specific antigen levels. These findings suggest that butein could be developed as an agent against PCa in humans.
Results
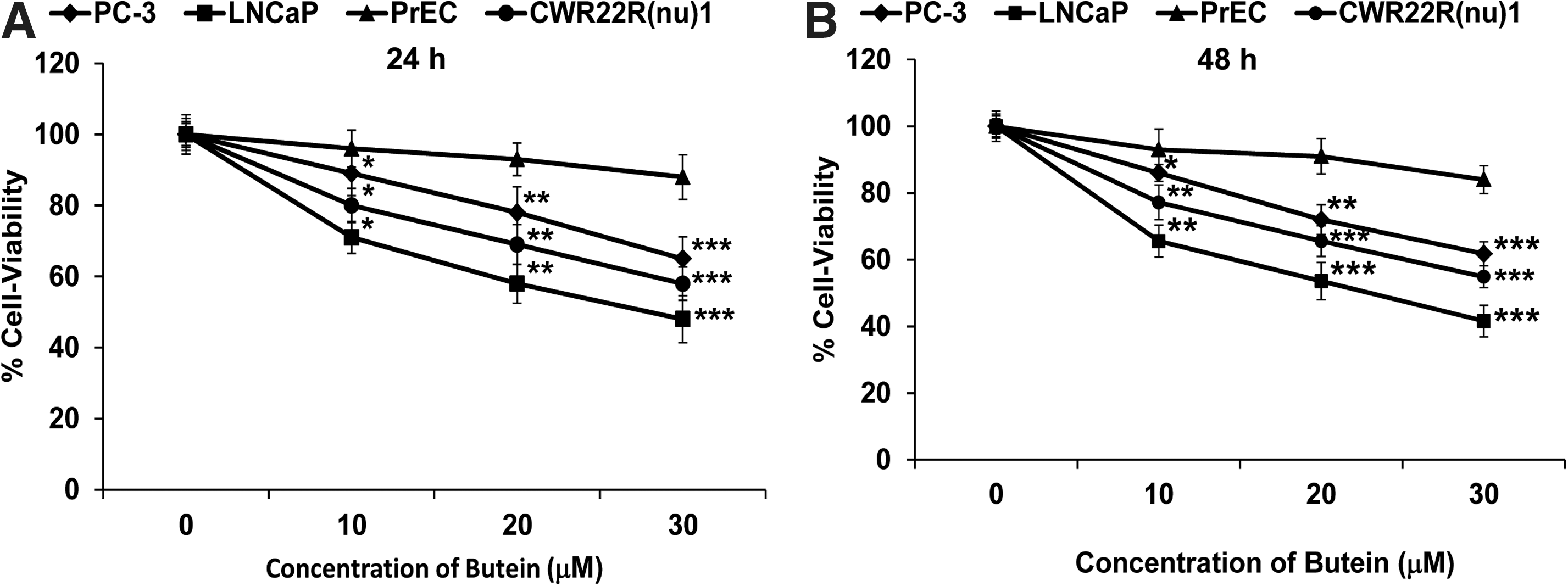
Butein caused inhibition of cell growth in human PCa cells
The dose- and time-dependent effects of treatment with butein (10–30 μM) on the growth of human PCa (LNCaP, CWR22Rν1, and PC-3) cells and normal prostate epithelial cells (PrEC) were investigated. We evaluated the effect of butein on the growth of these cells by 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazoliumbromide (MTT) assay. We compared the antiproliferative effects of butein on LNCaP, CWR22Rν1, PC-3, and PrEC cells. Treatment with butein (10–30 μM) for 24 h decreased cell viability in LNCaP (29%, 42%, and 52%), CWR22Rν1 (20%, 31%, and 42%), and PC-3 (11%, 22% and 35%) cells but had a minimal effect on PrEC cells at these doses (Fig. 2A). At 48 h, there was a much pronounced decrease in cell viability on treatment with butein (10–30 μM) in LNCaP (35%, 46%, and 58%), CWR22Rν1 (23%, 34%, and 45%), and PC-3 (14%, 28% and 39%) cells but had a minimal effect on PrEC cells (Fig. 2B). Since we found a maximum decrease in cell-viability in LNCaP cells as compared with CWR22Rν1 and PC-3 cells, we selected LNCaP cells for further experiments.
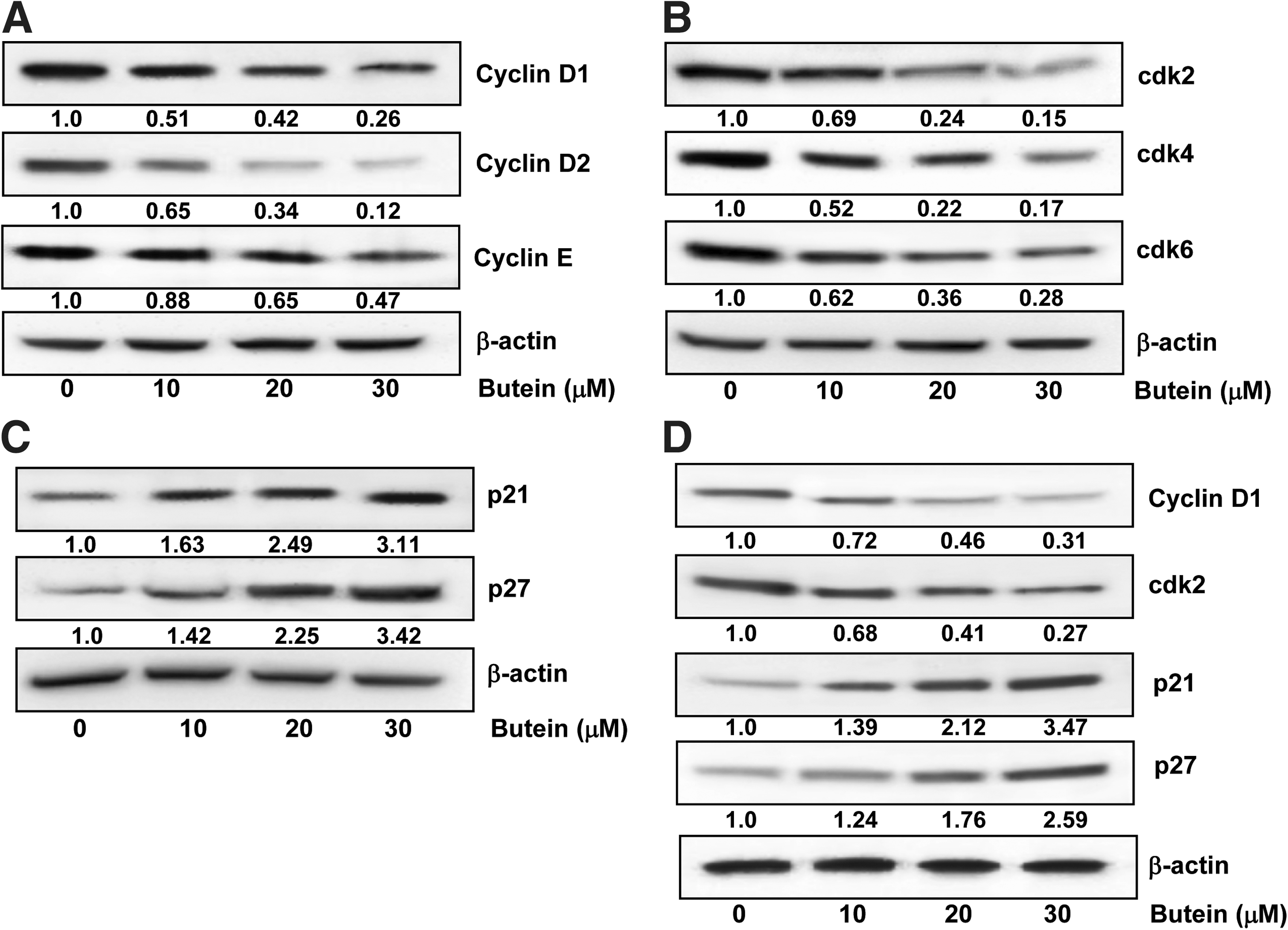
Butein caused inhibition of cyclins, cdks, and induction of WAF1/p21, KIP1/p27 in PCa cells
Cell division is uncontrolled and cyclin-dependent kinases (cdks)/cyclins act as check-points or targets in cancer cells. Cell-cycle progression is tightly regulated by interaction between cdks, cyclins, and inhibitor proteins WAF1/p21 and KIP1/p27 (1). Treatment of LNCaP cells with butein (10–30 μM; 48 h) caused a dose-dependent decrease in the protein expression of cyclins D1, D2, and E (Fig. 3A) and cdk2, 4, and 6 (Fig. 3B) by using immunoblot analysis. We next assessed the effect of butein treatment on the induction of WAF1/p21 and KIP1/27. Treatment with butein resulted in an induction of WAF1/p21 and KIP1/p27 even at the lowest concentration of 10 μM with a significant increase at the highest concentration of 30 μM (Fig. 3C). Treatment of CWR22Rν1 cells with butein (10–30 μM; 48 h) also caused an inhibition of cyclin D1, and cdk2 with increase in the protein expression of WAF1/p21 and KIP1/27 (Fig. 3D), suggesting that the effects of butein on PCa cells are not limited for one-cell type.
Butein caused an inhibition of NF-κB and IκB kinaseα and phosphorylation and degradation of IκBα in PCa cells
NF-κB forms a family of transcription factors that contributes in numerous biological processes, including immune response, inflammation, cell growth, survival, and development. The NF-κB proteins are usually sequestered in the cytoplasm by a family of inhibitors, including IκBα and other related ankyrin repeat-containing proteins. The degradation of IκBα is mediated through its phosphorylation by the IκB kinase (IKK), a trimeric complex composed of two catalytic subunits, IKKα and IKKβ, and a regulatory subunit, IKKγ (8). We found a significant dose-dependent inhibition in the phosphorylation of IκBα protein on treatment of LNCaP cells with butein (Fig. 4A). We measured the IKKα protein level to evaluate the possible inhibitory mechanism of butein on IκBα protein degradation. Immunoblot analysis showed that pretreatment of LNCaP cells with butein inhibited IKKα in a dose-dependent manner (Fig. 4A). We also investigated whether treatment with butein inhibits NF-κ
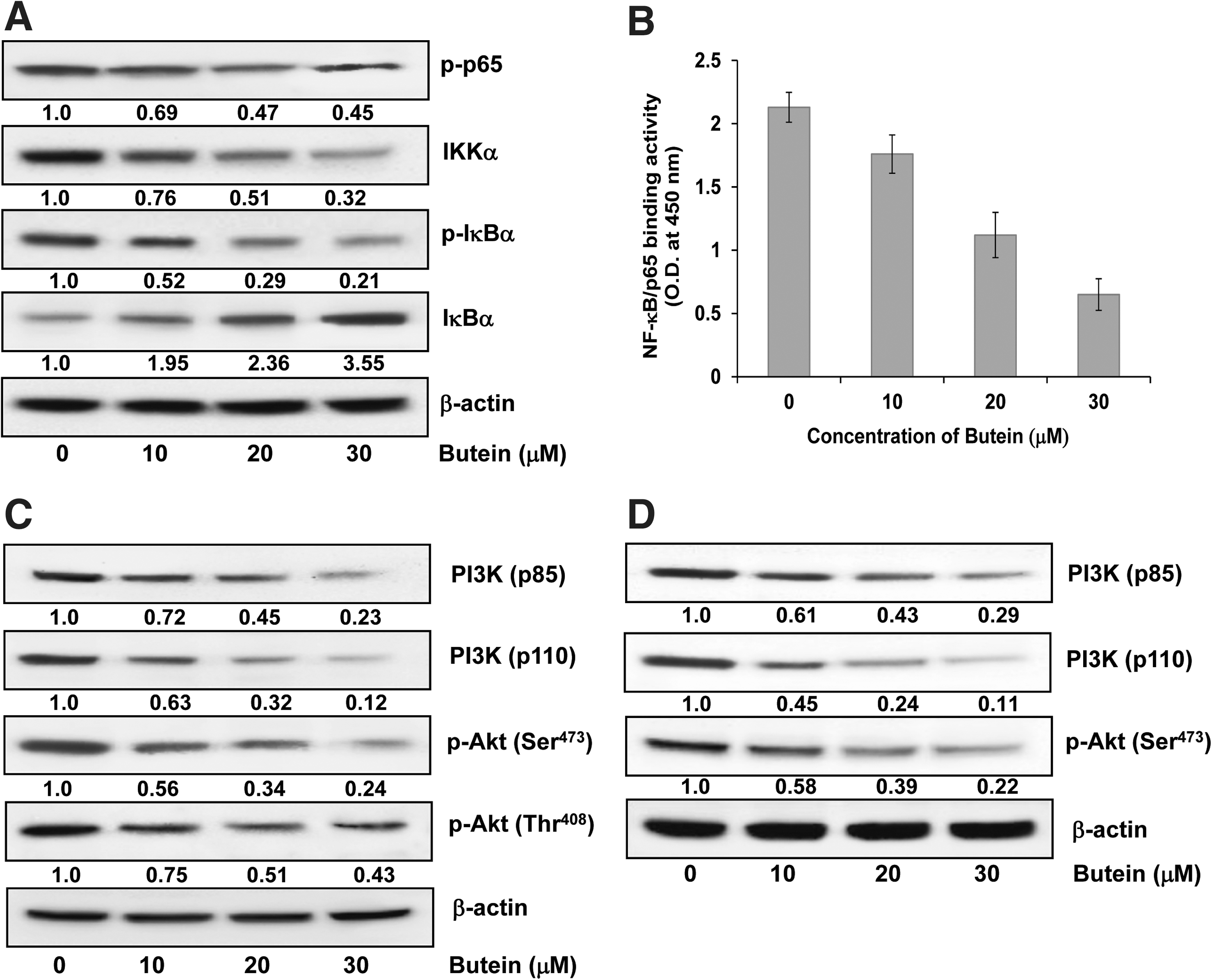
Butein caused inhibition of PI3K and phosphorylation of Akt protein expression in PCa cells
The PI3K control key signaling pathways in cancer cells, leading to cell proliferation, survival, motility and angiogenesis. Activation of PI3K results from gain of function or over-expression of PI3K and/or hyperactivity of up- or downstream players in the pathway in many cancer types (5). Treatment of LNCaP and CWR22Rν1 cells with butein caused a dose-dependent inhibition in the expression of both regulatory (p85) and catalytic (p110) subunits of PI3K (Fig. 4C, D).
Akt is a component of an essential pathway for cell survival and growth during development and carcinogenesis. It regulates cell cycle and proliferation by directly targeting cdks-inhibitors WAF1/p21 and KIP1/p27 and by indirectly modulating levels of cyclin D1 (39). Since we observed a decrease in the levels of cdks-inhibitors and cyclin D1 on treatment of cells with butein, we determined the effect on phosphorylation of Akt. We found that treatment of LNCaP cells with butein caused a dose-dependent inhibition in the phosphorylation of Akt at Ser473 and Thr308 (Fig. 4C), and treatment of CWR22Rν1 cells with butein caused an inhibition in the phosphorylation of Akt at Ser473 (Fig. 4D).
Butein caused cleavage of poly (ADP-ribose) polymerase and modulation of Bax/Bcl2 ratio in PCa cells
Poly (ADP-ribose) polymerase (PARP) has several cellular functions including DNA recombination and repair, cellular proliferation, apoptosis in ischemic conditions, and necrotic cell death. On treatment of LNCaP cells with butein, we found that there was an increase in the protein expression of the 85 KD fragment of PARP, while there was a decrease in the protein expression of full-size PARP (116 KD) protein (Fig. 5A).

The deregulation of apoptosis is studied by the expression of Bax and Bcl2. Bax is proapoptotic, whereas Bcl2 is antiapoptotic. There was a significant increase in the protein expression of Bax with a dose-dependent decrease in the protein expression of Bcl2 on treatment of LNCaP and CWR22Rν1 cells with butein as shown in Figure 5B and C, respectively. We also observed a significant dose-dependent shift in the ratio of Bax to Bcl-2 after treatment with butein, indicating the induction of apoptosis (Fig. 5D, E).
Butein caused induction of apoptosis in PCa cells
For the detection of apoptotic and necrotic cells, annexin-V/PI staining was performed after treatment of LNCaP cells with butein. When LNCaP cells were stained with annexin-V and examined under a fluorescence microscope, apoptotic cells were found to be increased in butein-treated cells. Treatment with general caspase-inhibitor Z-VAD-FMK caused an inhibition of apoptosis, which was further increased when the cells were treated with the combination of Z-VAD-FMK and butein (20 μM) for 48 h (Fig. 6A). We found no necrotic cells on treatment with butein, suggesting that butein caused an induction of apoptosis in LNCaP cells (Fig. 6A).

Butein caused induction of active caspases-3, -8 and -9 in PCa cells
Caspases are a family of evolutionally highly conserved cysteine proteases that act as common effector molecules in various forms of cell death. Caspases are synthesized as inactive proenzymes and become activated either by oligomerization in a large multimeric complex, which is the case for the initiator caspases-8 and -9, or alternatively via proteolytic cleavage, which applies for effector caspases such as caspase-3. Once activated, they cleave various substrates in the cytoplasm or nucleus, causing characteristic morphological features of apoptotic cell death (7). Treatment of LNCaP cells with butein (10–30 μM) caused an increase in the activation of caspases-3, -8, and -9 dose dependently (Fig. 6B).
Inhibition of butein-induced induction of caspases by Z-VAD-FMK in PCa cells
Since we found that treatment of Z-VAD-FMK blocked butein-induced activation of caspases (Fig. 6A), we confirmed the results by immunoblot analysis. Pretreatment of LNCaP cells with Z-VAD-FMK (40 μM) for 2 h before treatment with butein (20 μM) resulted in a significant decrease in the protein expression of active caspases-3, -8, and -9 as compared with treatment of cells with butein alone (Fig. 6C).
Butein caused inhibition of the growth of human prostate carcinoma CWR22Rν1 cells and PSA secretion in athymic nude mice
In athymic nude mice implanted with AR-positive CWR22Rν1 human PCa cells, treatment with butein resulted in a significant inhibition of tumor growth. The appearance of small solid tumors was observed in animals of control group 7 days after cell inoculation. This latency period was prolonged to 14 days in animals receiving butein day 1 after tumor cell implantation (Fig. 7A). In the control group, the average tumor volume of 1200 mm3 was reached in 28 days after tumor cell inoculation. At this time point, the average tumor volume in the butein-treated group was 483 mm3. After tumor cell inoculation, the average tumor volume of 1200 mm3 in the butein-treated group was achieved in 42 days (Fig. 7B). Tumor data were analyzed for survival probability by Kaplan–Meier analysis, which indicated that treatment with butein to prostate tumor bearing athymic nude mice resulted in increased survival (p<0.0001, log-rank test), with a median survival of 42 days in butein-treated group mice compared with 28 days in control group mice (p<0.0001, log-rank test; data not shown). Quantitative sandwich ELISA was used to determine circulating PSA levels in mouse serum secreted by CWR22Rν1 tumor xenografts. At 28 days postinoculation, there was 52% decrease in the secreted PSA levels in the butein-treated group as compared with the control group normalized to tumor volume (Fig. 7C). Therefore, our results show that treatment of mice with butein caused a significant decrease in the serum PSA of athymic nude mice.

Butein caused inhibition of Ki-67, vascular endothelial growth factor, and CD31 in tumors of athymic nude mice
We examined the expression of molecules associated with cell proliferation and angiogenesis in xenografts by immunohistochemistry. Ki-67 is a cell proliferation marker and is expressed in all phases of the cell cycle, except in resting cells. By immunohistochemical analysis of the tissues from athymic nude mice, we found a significant increase in the number of Ki-67 positive cells in the control group as compared with the group treated with butein (Fig. 8). Solid tumors recruit new blood vessels for their growth, maintenance, and metastasis. Discovering drugs that suppress tumor-induced development of new blood vessels is an important strategy for cancer treatment, and many current clinical therapies mainly target vascular endothelial growth factor (VEGF) and CD31. Expression levels of VEGF and CD31 were higher in tissues of control mice than in mice treated with butein (Fig. 8). The intensity and degree of staining was markedly reduced in tissue sections of butein-treated groups. Morphologically, tumor tissues showed more solid cords, sheet-like cells, and glandular structures.

Discussion
PCa is a heterogeneous disease; while most cases behave indolently and are undiagnosed during life, it is still the second most common cause of cancer death in men. Many men are being diagnosed with PCa than die from it over the past two decades. This upsurge in diagnosis has led to an aggressive treatment of indolent disease in many persons and has been the motivation for finding a means of reducing the risk of PCa. Various dietary and lifestyle factors are involved in the multifactorial etiology, development, and progression of PCa (15, 33).
In this study, we have shown that treatment of PrEC cells with butein caused minimal effects, while there was a decrease in cell viability in PCa cells. The ability of the cells to divide, in turn, is mainly attributed to the presence of two classes of molecules, Cdks, a family of serine/threonine kinases and their binding partners, cyclins. Cells progress through the various phases of the cell cycle via the interactions of different cyclins with their respective cdk subunits (28). Quiescent cells enter the cell cycle after mitogenic stimuli and upregulate the cyclins D and E during G1 phase of the cell cycle (23). Cyclin D is associated with cdk-4 and -6 (2), while cyclin E is associated with cdk2 (20). Cyclins and cdks play important roles in synthesis of DNA and cell division. Inhibitors of cyclin-cdks can modulate the cell cycle by preventing or limiting cyclin-cdks from phosphorylating their normal substrates (36). Treatment of PCa cells with butein resulted in a decrease in cyclins D1, D2, E, and cdks-2, 4, and -6 with an increase in the protein expression of WAF1/p21 and KIP1/p27.
NFκB can modulate the transcriptional activation of genes associated with cell proliferation, angiogenesis, metastasis, tumor promotion, inflammation, and suppression of apoptosis (3, 11). Tumor cells cease proliferation and become more sensitive to the action of antitumor agents by blocking of NF-κB (13). Hence, NFκB is the subject of intense study, and agents capable of suppressing NF-κB activation have therapeutic promise and potential to inhibit carcinogenesis. Our results showed that treatment of human PCa cells with butein inhibited NF-κB, IKKα, and phosphorylation and degradation of IκBα protein. Our study suggests that the effects of butein on NF-κB occurred through inhibition of phosphorylation and subsequent proteolysis of IκBα as butein blocked phosphorylation and degradation of IκBα.
PI3K pathway plays a pivotal role in cell growth, proliferation, and survival and signaling via this pathway is upregulated in cancer (4, 24). PI3 kinases are heterodimers comprising of a regulatory subunit (p85) and a catalytic subunit (p110). The Akt kinase represents the primary downstream mediator of the effects of the PI3K pathway, and plays a central role in both normal and pathological signaling (32). Akt occupies a key regulatory node in the PI3K pathway, below which the pathway branches significantly to influence a wide range of cellular processes that promote cell-cycle progression, cell growth, energy metabolism, and resistance to apoptosis (9). We have demonstrated that treatment of human PCa cells with butein resulted in a decrease in the expression of PI3K (p85 and p110) and phosphorylation of Akt at Ser473 and Thr408. Apoptotic pathways are regulated by the members of the Bcl2 family of proteins with individual members that can suppress (e.g., Bcl2) or promote (e.g., Bax) apoptosis. We found that butein treatment caused a decrease in the protein expression of Bcl2 and an increase in Bax; therefore, the ratio of Bax to Bcl-2 was altered in favor of apoptosis, which was also confirmed by cleavage of PARP. The process of apoptosis is controlled by multiple and interrelated pathways, ensuring that caspases, the proteolytic initiators, and executioners of apoptosis are triggered only in cells requiring termination. We have shown in the present study that caspases-3, -8, and -9 were activated during butein-mediated apoptosis and that addition of a general caspase inhibitor decreased butein-mediated apoptosis. These data provide evidence that butein-mediated apoptosis was caspase dependent in human PCa cells.
To establish the relevance of these in-vitro findings to in-vivo situations, we implanted athymic nude mice with human PCa cells. Treatment with butein significantly decreased PCa tumor growth in mice and caused a significant reduction in serum levels of PSA. Serum butein levels were not studied, because butein was administered intraperitoneally to mice, and the in-vivo effects were comparable to the in-vitro effects, suggesting that the cells and tumor growth inhibition was related to butein. We also found that there was an increase in the expression of cell-proliferation marker (Ki-67) and markers of angiogenesis (VEGF and CD31) in tissues of control mice than in mice treated with butein. The present study demonstrates the effect of butein, a plant polyphenol, on human PCa cells growth in in-vitro and in-vivo preclinical models. We suggest that butein could be a valuable chemopreventive and/or chemotherapeutic agent against human PCa.
Materials and Methods
Materials
The mono and polyclonal antibodies cdks 2, 4, and 6, WAF1/p21, KIP1/p27, Bcl2, VEGF, and CD31 were obtained from Santa Cruz Biotechnology Inc. Anti-cyclins D1, D2, E, Bax, Bcl2, active caspases-3, 8, and 9, Akt, phospho-Akt (Ser473 and Thr308), PI3K (p85 and p110), IKKα, IκBα, and phospho-IκBα antibodies were obtained from Cell Signaling Technology. PARP (116) and NF-κB/p65 were procured from Upstate. PARP (85) was obtained from Promega. Ki-67 antibody was obtained from Epitomics. Butein was purchased from Enzo Life Sciences. ELISA kit for NF-κB/p65 DNA binding activity was purchased from Active Motif. Human PSA Elisa kit was procured from Anogen. Annexin-V-Fluos staining kit was from Roche Diagnostics Corporation. Anti-mouse and anti-rabbit secondary antibody horseradish peroxidase (HRP) conjugate was obtained from Amersham Life Science Inc. BCA Protein assay kit was obtained from Pierce. Novex precast Tris-glycine gels were from Invitrogen.
Cell culture and treatment
The LNCaP, CWR22Rν1, and PC-3 cells were obtained from ATCC. LNCaP, CWR22Rν1, and PC-3 cells were grown in RPMI 1640 (Invitrogen) supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin. Human PrEC were obtained from Cambrex Bioscience and grown in PrEC basal cell medium (Cambrex Bioscience) according to the manufacturer's instructions. The cells were maintained under standard cell culture conditions at 37°C and 5% CO2 in a humid environment. Butein dissolved in dimethyl sulfoxide (DMSO; final concentration 0.1% v/v) was used for the treatment of cells. The cells (60%–70% confluent) were treated with butein (10–30 μM) for 48 h in complete growth medium.
Cell viability
The effect of butein on the viability of cells was determined by 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazoliumbromide assay. LNCaP, CWR22Rυ1, PC-3, and PrEC cells were plated at 1×104 cells per well in 200 μl of complete culture medium containing 10–30 μM concentrations of butein in 96-well microtiter plates for 24 and 48 h. After incubation for specified times at 37°C in a humidified incubator, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazoliumbromide (5 mg/ml in PBS) was added to each well and incubated for 2 h, after which the plate was centrifuged at 1800×g for 5 min at 4°C. The absorbance was recorded on a microplate reader at the wavelength of 540 nm. The effect of butein (10–30 μM) for 24 and 48 h on growth inhibition was assessed as percent cell viability where DMSO-treated cells were taken as 100% viable. DMSO at the concentrations used was without any effect on cell viability.
ELISA for NF-κB/p65
This was done by using commercially available Trans-AM kit as previously described (18).
Apoptosis assessment by annexin-V staining
LNCaP cells were treated with butein (10, 20, and 30 μM) 48 h. The annexin-V-Fluos staining kit was used for the detection of apoptotic cells according to vendor's protocol. The fluorescence was measured by Nikon Eclipse Ti system (Nikon Instruments, Inc.). Images were captured with an attached camera linked to a computer.
Protein extraction and western blotting
This was performed as previously described (17, 18). Densitometric measurements of the band in Western blot analysis were performed using digitalized scientific software program Image J (National Institutes of Health).
In vivo tumor xenograft model
Athymic (nu/nu) male nude mice were obtained from NxGen Biosciences and housed under pathogen-free conditions with a 12-h light/12-h dark schedule, and fed with an autoclaved diet ad libitum. We selected AR-positive CWR22Rν1 cells for determining the in vivo effects of butein based on the fact that these cells form rapid and reproducible tumors in nude mice and secrete significant amounts of PSA in the bloodstream of the host. CWR22Rν1 tumor xenografts were generated as described in our earlier study (16). Animals were divided into two groups consisting of eight animals in group 1 and 16 animals in group 2. Mice in group 1 (control) received an intraperitoneal injection of DMSO (30 μl). Mice in group 2 received an i.p. injection of butein (1 mg/animal) in 30 μl of DMSO twice weekly. Tumor size was measured weekly as previously described (34). When tumors reached a volume of 1200 mm3 in the control group, all mice of group 1 and 8 animals from group 2 were sacrificed. Remaining mice of group 2 were sacrificed when tumors reached a volume of 1200 mm3. All procedures conducted were in accordance with the guidelines laid down for the use and care of laboratory animals. At the end of the study, blood samples were collected by the mandibular bleed. The sera were separated by centrifuging blood for 5 min at 4°C and then stored at −20°C until assayed for secreted PSA.
ELISA for PSA
The human PSA ELISA kit from Anogen was used for the quantitative determination of PSA levels in serum.
Immunohistochemical analysis
Sections (5 mm thick) were cut from paraffin-embedded lung tissues. Immunostaining was performed using specific antibodies with appropriate dilutions and was replaced with either normal host serum or block for negative controls, followed by staining with appropriate HRP-conjugated secondary antibodies. The slides were developed in diaminobenzidine and counter stained with a weak solution of hematoxylin stain. The stained slides were dehydrated and mounted in permount and visualized on Nikon Eclipse Ti system (Nikon Instruments, Inc.). Images were captured with an attached camera linked to a computer.
Statistical analysis
Results were analyzed using a two-tailed Student's t-test to assess statistical significance, and p values<0.05 were considered significant.
Footnotes
Acknowledgments
This study was supported by National Institutes of Health, National Cancer Institute Grants R01CA120451 (to HM) and R03CA153961 (to NK).
Author Disclosure Statement
No competing financial interests exist.