
Abstract
Introduction
Many previous studies have shown consistently that HICI and HPV were largely dependent on extracellular Ca2+ (9, 10, 13, 19, 25, 29, 34, 35, 42). Apparently, the extracellular Ca2+ dependence can be interpreted as the Ca2+ source for Ca2+ influx from extracellular space stimulated by hypoxia. Extracellular Ca2+ entry may be activated by hypoxia-altered level of reactive oxygen species (ROS) (1, 18, 26, 39, 41). The suggested Ca2+ entry pathways by hypoxia include store- (4, 23, 29, 35, 42) and voltage-operated Ca2+ influx (1, 20, 43). Meanwhile, Ca2+ release channel(s) especially ryanodine receptors (RyRs) have also been broadly documented to be involved in HICI and HPV (8, 12, 15, 32, 34, 48).
Innovation
Hypoxic pulmonary vasoconstriction (HPV) matches lung ventilation with blood perfusion and also leads to pulmonary hypertension. The mechanism underlying HPV remains largely unknown. The Ca
Extracellular Ca2+ is also the ligand for extracellular calcium-sensing receptor (CaSR) (14). The dependence of HICI and HPV on extracellular Ca2+ may not be understood solely as Ca2+ influx secondary to the depletion of intracellular Ca2+ stores by hypoxia. Substantial progress in this controversial point warrants a novel attempt to elucidate how these different Ca2+ pathways orchestrate to induce [Ca2+]i elevation during hypoxia. As a matter of fact, however, it has never been studied whether extracellular Ca2+ plays any role(s) in addition to the source of Ca2+ influx from extracellular space in HICI and HPV.
CaSR is a G-protein-coupled membrane receptor that senses extracellular Ca2+ concentration and conveys this information to intracellular space through multiple signal pathways including [Ca2+]i and extracellular regulated protein kinase (14). CaSR protein is expressed in vascular smooth muscle cells (6, 21, 45). The activation of CaSR in vascular smooth muscle cells increases [Ca2+]i and induces a vasoconstriction of the gerbilline spiral modiolar artery (45). Additionally, functional CaSR appears sensitive to redox status (2, 47).
This study aimed at investigating the mechanism underlying extracellular Ca2+ dependence of HICI and its associations with RyR (8, 15, 34, 48), store-operated Ca2+ influx (SOC) (4, 23, 29, 35, 42), and HPV. It was found that functional CaSR, which can be sensitized by hypoxia-induced ROS, serves as a bridge between extracellular Ca2+ and [Ca2+]i elevation and, therefore, is pivotal in HPV.
Results
Expression of a functional CaSR: sensing extracellular Ca2+
Immunocytostaining showed the major localization of CaSR in the cytosol in permeabilized PASMCs and the membrane localization of the mature functional protein in nonpermeabilized PASMCs (Fig. 1A). Western blot on cell lysate simultaneously isolated from cytoplasm and membrane revealed a 170 kDa band, presumably a prematured form (24, 50) and the other 55–72 kDa one, respectively (Fig. 1B).
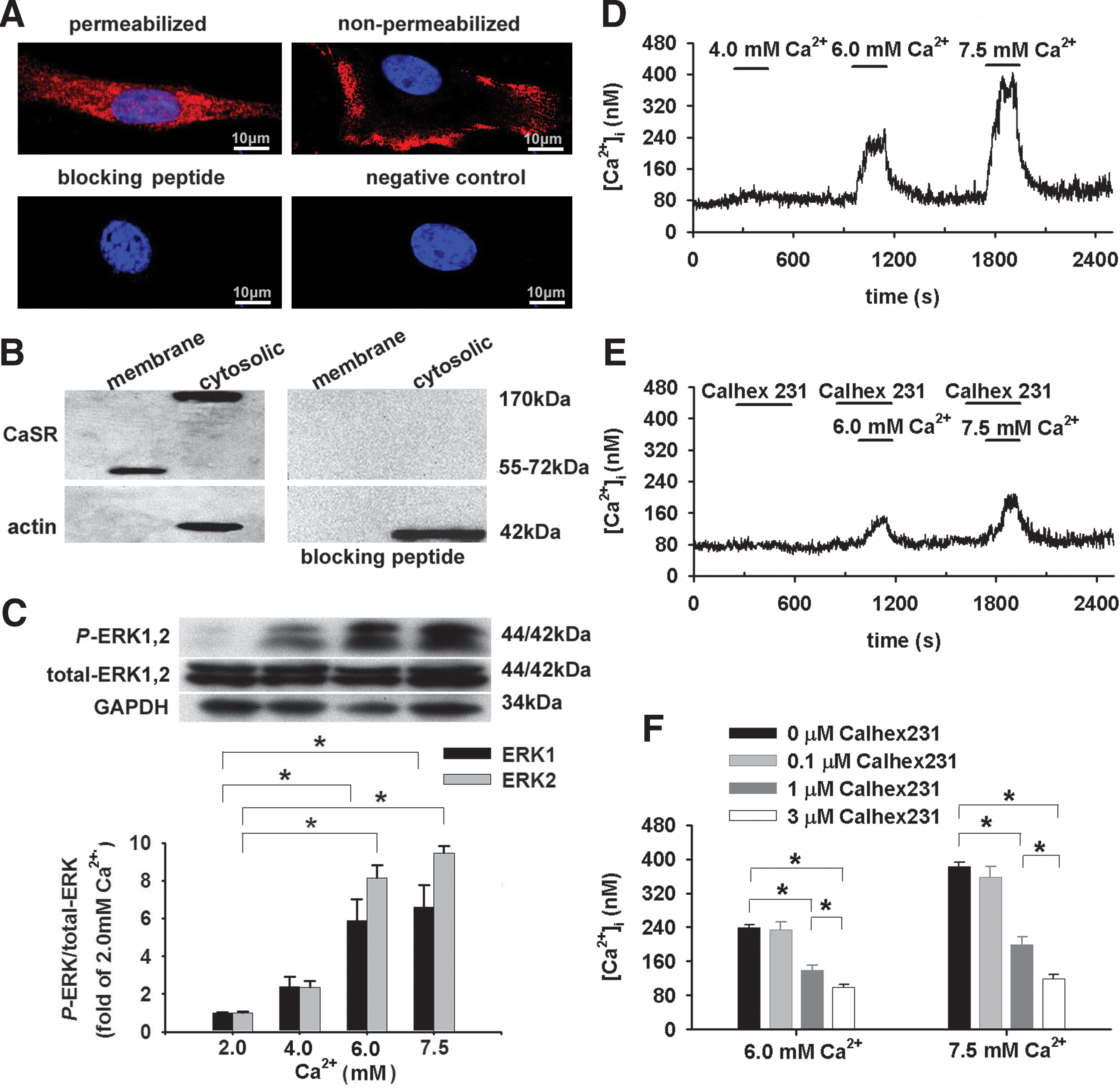
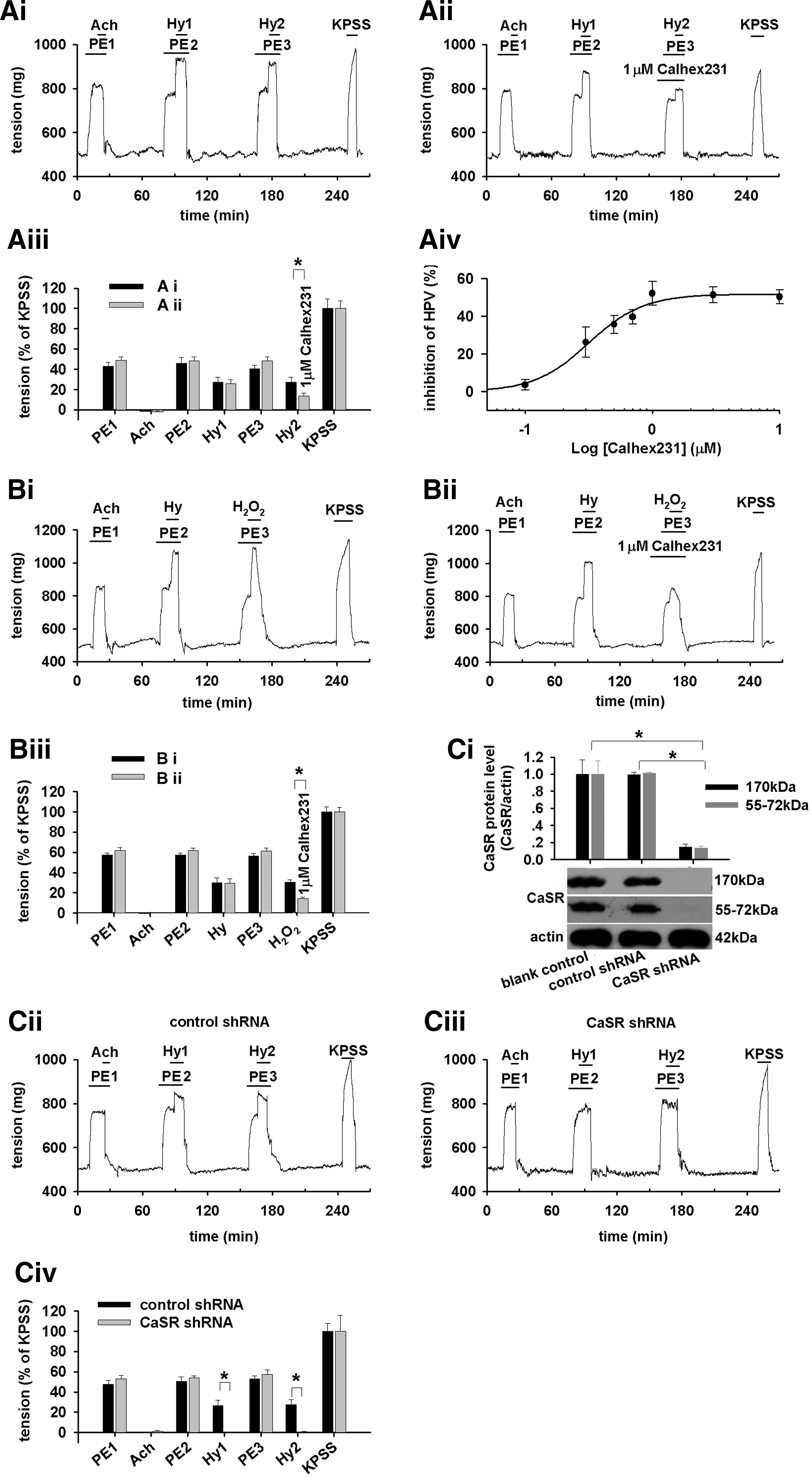
To determine whether the CaSR protein is functional, PASMCs were exposed to increased extracellular Ca2+ concentration ([Ca2+]o) (5). A 3-min [Ca2+]o elevation from 2.0 to 6.0 and 7.5 mM enhanced phosphorylation of ERK1,2 (p44/p42; Fig. 1C), an indication of CaSR activation (14) and increased [Ca2+]i (n=23, p<0.05 for each, Fig. 1D, F). (1S,2S,1′R)-N1-(4-chlorobenzoyl)-N2-[1-(1-naphthyl)ethyl]-1,2-diaminocyclo-hexane (Calhex231), the negative allosteric modulator of CaSR, dose-dependently inhibited [Ca2+]o elevation-stimulated [Ca2+]i increase (p<0.05, Fig. 1E, F). Thus, PASMCs can sense [Ca2+]o through CaSR-mediated [Ca2+]i signaling.
HICI: extracellular Ca2+ dependence, Calhex231 inhibition, and CaSR dependence
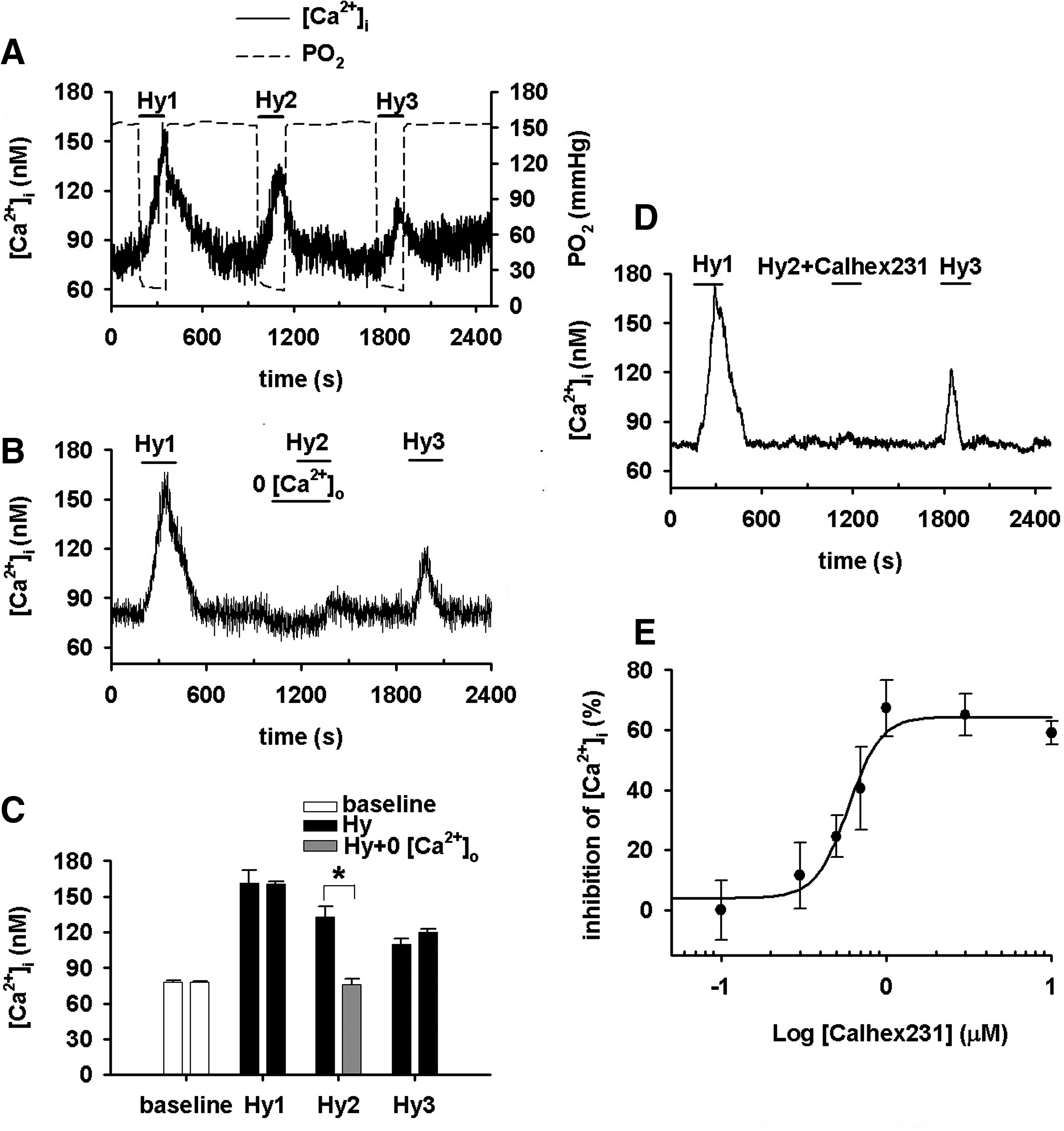
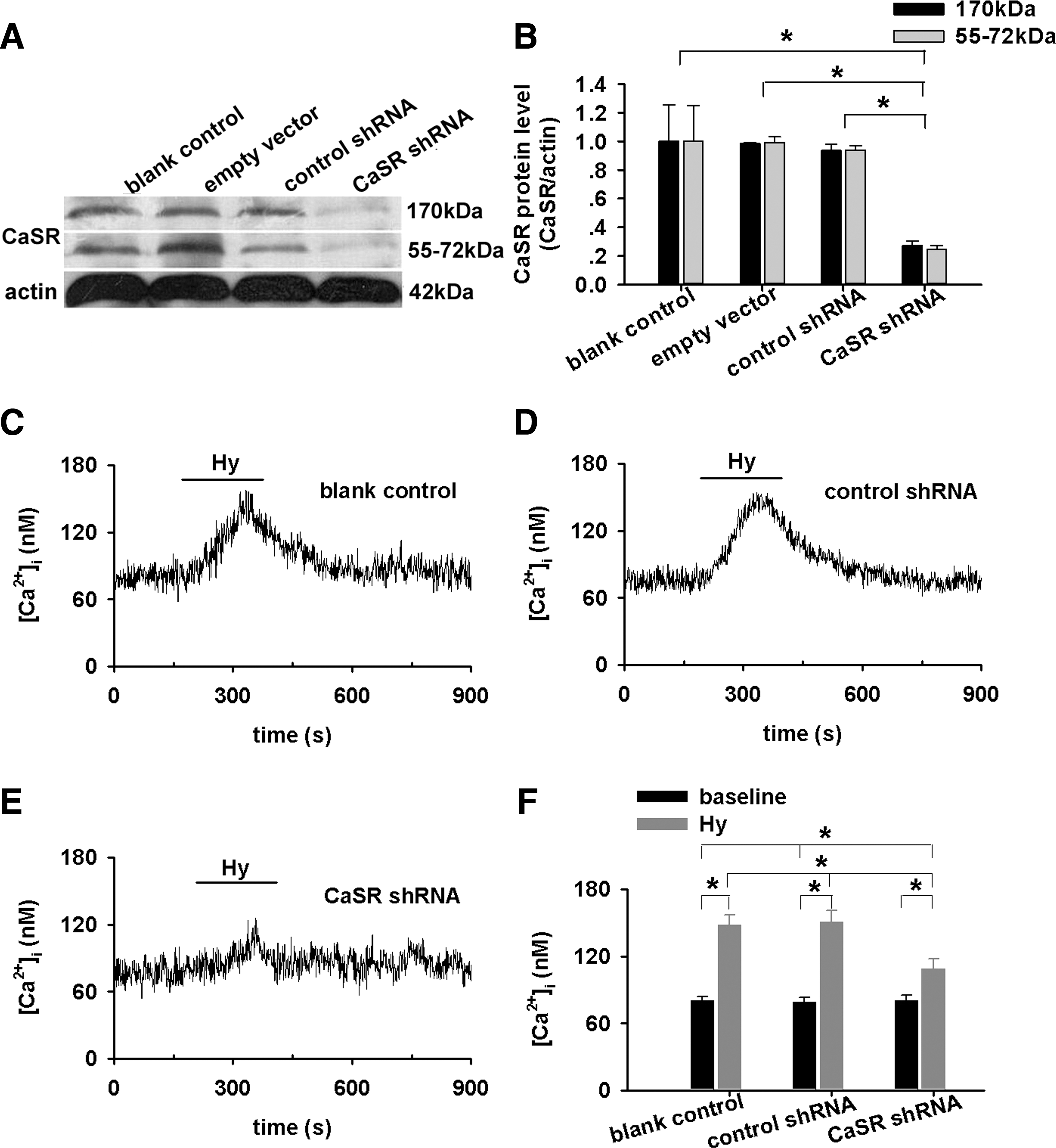
Hypoxia, by a quick superfusion of PASMCs with hypoxic medium for 3 min, increased [Ca2+]i (n=51, p<0.01, Fig. 2A, C). The HICI was completely abolished or significantly inhibited by the removal of extracellular Ca2+ (n=43, Fig. 2B, C), the presence of Calhex231 (n=8, Fig. 2D, E), or CaSR knockdown (p<0.01 vs. blank or nonspecific shRNA respectively, Fig. 3C–F). Of note, CaSR knockdown abolished/inhibited [Ca2+]i response to 6.0 and 7.5 mM [Ca2+]o, whereas it does not affect the store Ca2+ contents in PASMCs (Supplementary Figs. S1 and S2; Supplementary Data are available online at
Ca2+ sensitivity of CaSR: enhanced by ROS
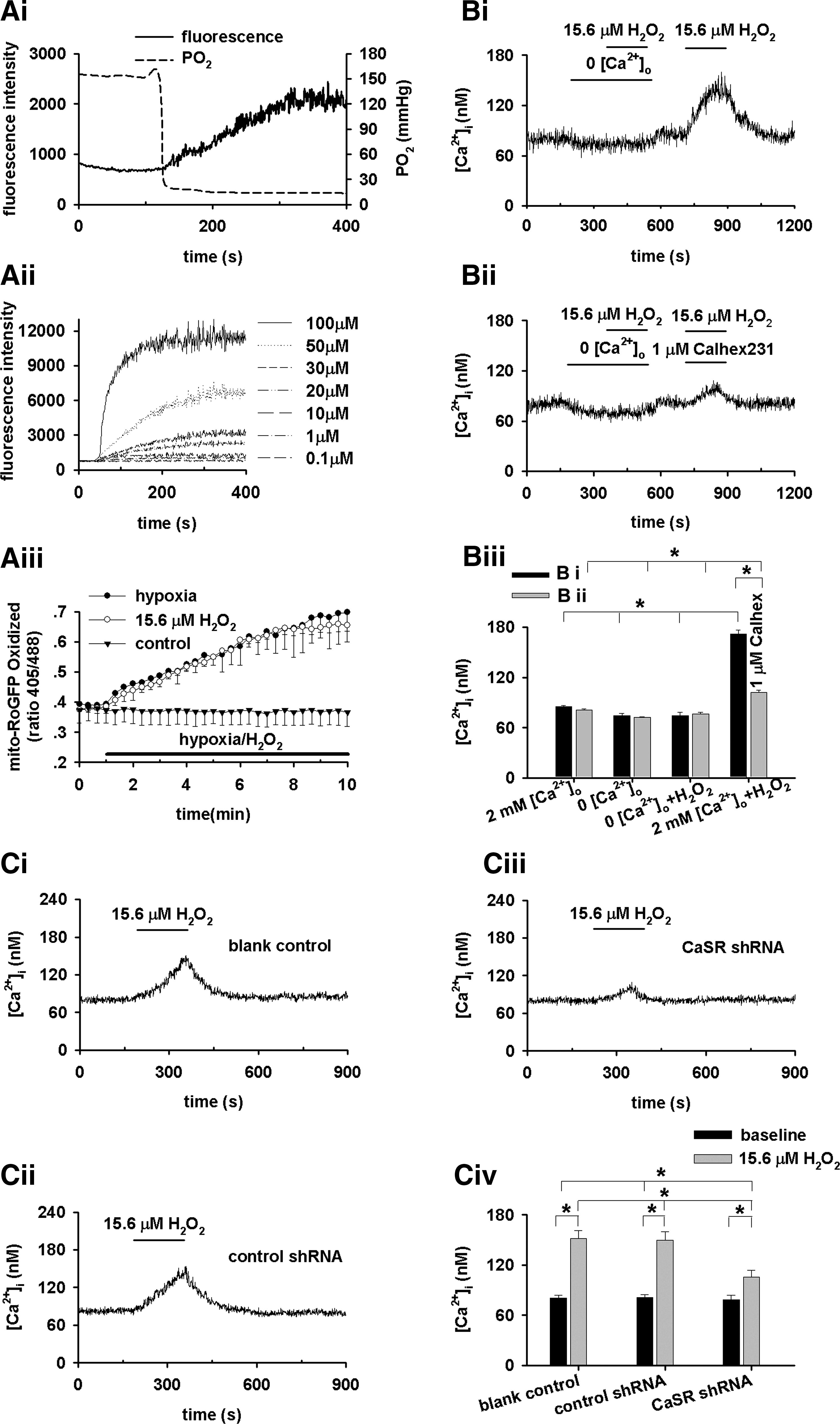
Hypoxia, as indicated by a quick decline in PO2 from 154±4 to 19±1 mmHg (n=4), stimulated an abrupt increase in ROS (n=22, Fig. 4Ai), estimated (30) to a level equivalent to 15.6 μM extracellular H2O2 (n=21–26 for each dose, Fig. 4Aii). The hypoxia-stimulated ROS generation in PASMCs was confirmed using a redox-sensitive green fluorescent protein (GFP)-RoGFP (Fig. 4Aiii) (40). Hypoxia-stimulated ROS generation is not affected by Calhex231 (Supplementary Fig. S3). About 15.6 μM H2O2, not a lower concentration of 13.0 μM (Supplementary Fig. S4), which did not induce any [Ca2+]i transient in the absence of extracellular Ca2+, triggered a [Ca2+]i transient in the presence of 2 mM [Ca2+]o (n=5, Fig. 4Bi, Biii). These results are consistent with previous studies showing that H2O2 increased [Ca2+]i in smooth muscle cells (18, 46), and the [Ca2+]i increase by low μM H2O2 depended on extracellular Ca2+ (46). 1 μM Calhex231 and CaSR-knockdown significantly inhibited 15.6 μM H2O2-induced [Ca2+]i transient (n=13, Fig. 4Bii, Biii, C). [Ca2+]o elevation-induced [Ca2+]i signal in PASMCs is enhanced by the concomitant presence of 15.6 μM H2O2 or under hypoxia (Supplementary Fig. S5). Thus, the Ca
CaSR-dependent [Ca2+]i increase by hypoxia: role of mitochondria-derived H2O2
Mitochondria depletion (7, 39) using ethidium bromide almost completely diminished ROS generation by hypoxia (n=27, p<0.01 vs. control with or without pyruvate (Py) and uridine (Ur), Fig. 5A). Hypoxia triggered similar [Ca2+]i increase in control PASMCs cultured in the presence (n=8, p<0.01, Fig. 5Bi, Bv) and absence of Py and Ur (Fig. 2A). Mitochondria depletion completely abolished HICI, and this was reversed by exposure of PASMCs to 15.6 μM H2O2 (Fig. 5Bii, Bv). The exposure of H2O2-reversed [Ca2+]i increase during hypoxia in mitochondria-depleted PASMCs was prevented by 1 μM Calhex231 (n=15, p<0.01 vs. co-exposure of H2O2 in the absence of Calhex231, Fig. 5Biii, Bv) and by CaSR knockdown (Fig. 5Biv, Bv), suggesting that mitochondria-derived ROS sensitizes CaSR and thereafter triggers HICI.

PEG-Catalase, not PEG-SOD loaded into PASMCs (7), abolished HICI (Fig. 5Ci–Ciii), clarifying the specific role of intracellular H2O2 in HICI.
HICI: RyR and SOC dependence
To determine the downstream target of CaSR activation by hypoxia, we manipulated the intracellular Ca2+ release and SOC pathways. The inhibition of RyR by 100 μM ryanodine, but not the inhibition of IP3 receptor by 20 μM XeC, significantly attenuated HICI (p<0.05 vs. control, Fig. 6Ai, Aiv for ryanodine; p=nonsignificance vs. control, Fig. 6Aii, Aiv for XeC). These are consistent with previous studies using either genetically manipulated PASMCs (48) or pharmacological inhibitors (8, 11, 15, 34). Being accordant with other investigators (4, 23, 35), SOC inhibitor, 50 μM SKF96365 significantly inhibited HICI (p<0.05 vs. control, Fig. 6Aiii, Aiv). Similarly, Ryanodine and SKF96365, not XeC, inhibited H2O2-induced [Ca2+]i increase (Fig. 6B, C).
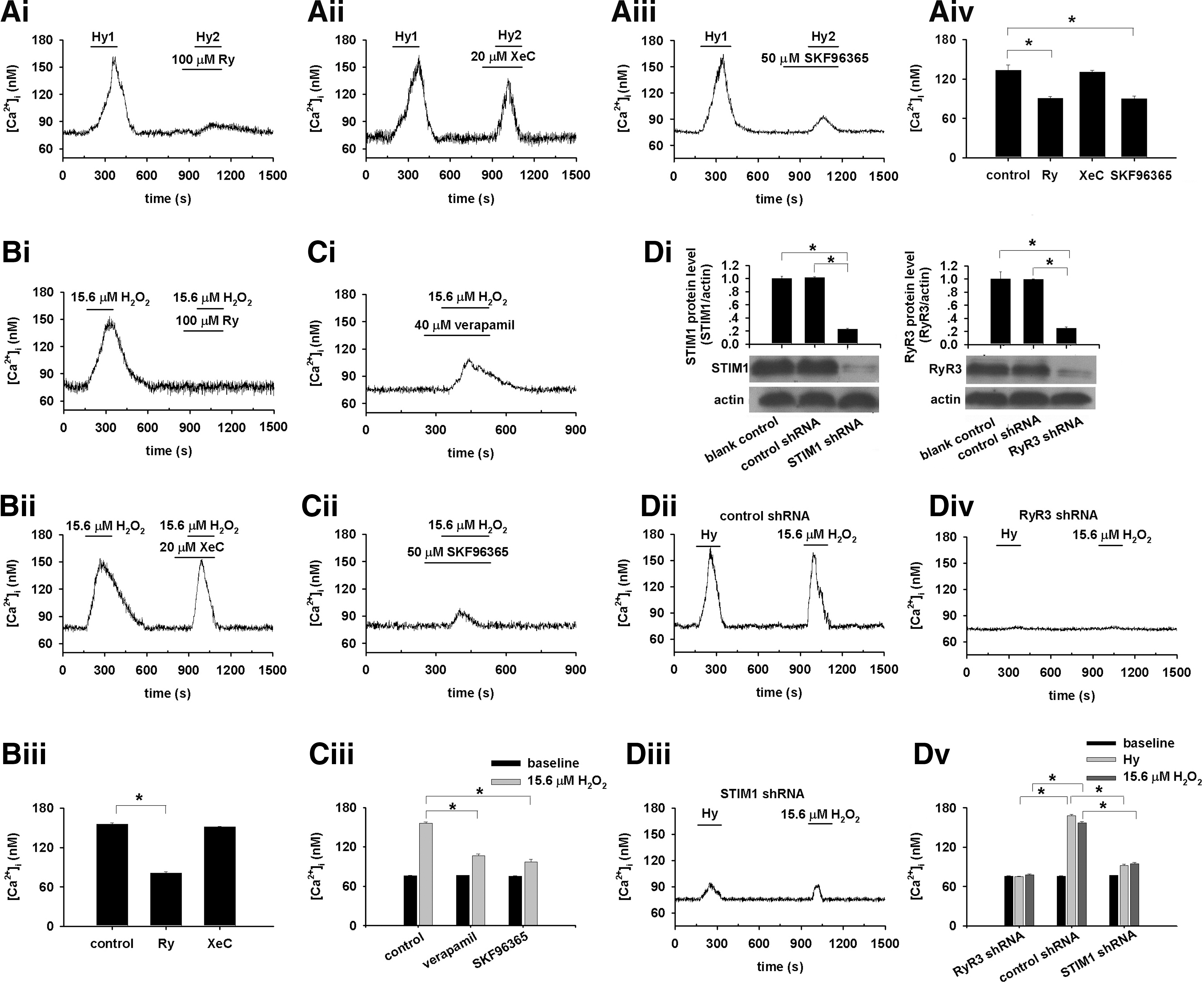
RyR3 (48) and STIM1 knockdown block or profoundly inhibited hypoxia- and H2O2-induced [Ca2+]i increase, respectively (Fig. 6D), revealing that RyR3 and STIM1 can be the major molecular targets of CaSR activation or sensitization upon hypoxia and H2O2 stimulation. However, this study does not exclude the involvement of voltage-sensitive pathways (1, 25, 43), as verapamil also inhibited H2O2-induced [Ca2+]i elevation (n=12, p<0.05 vs. control, Fig. 6C).
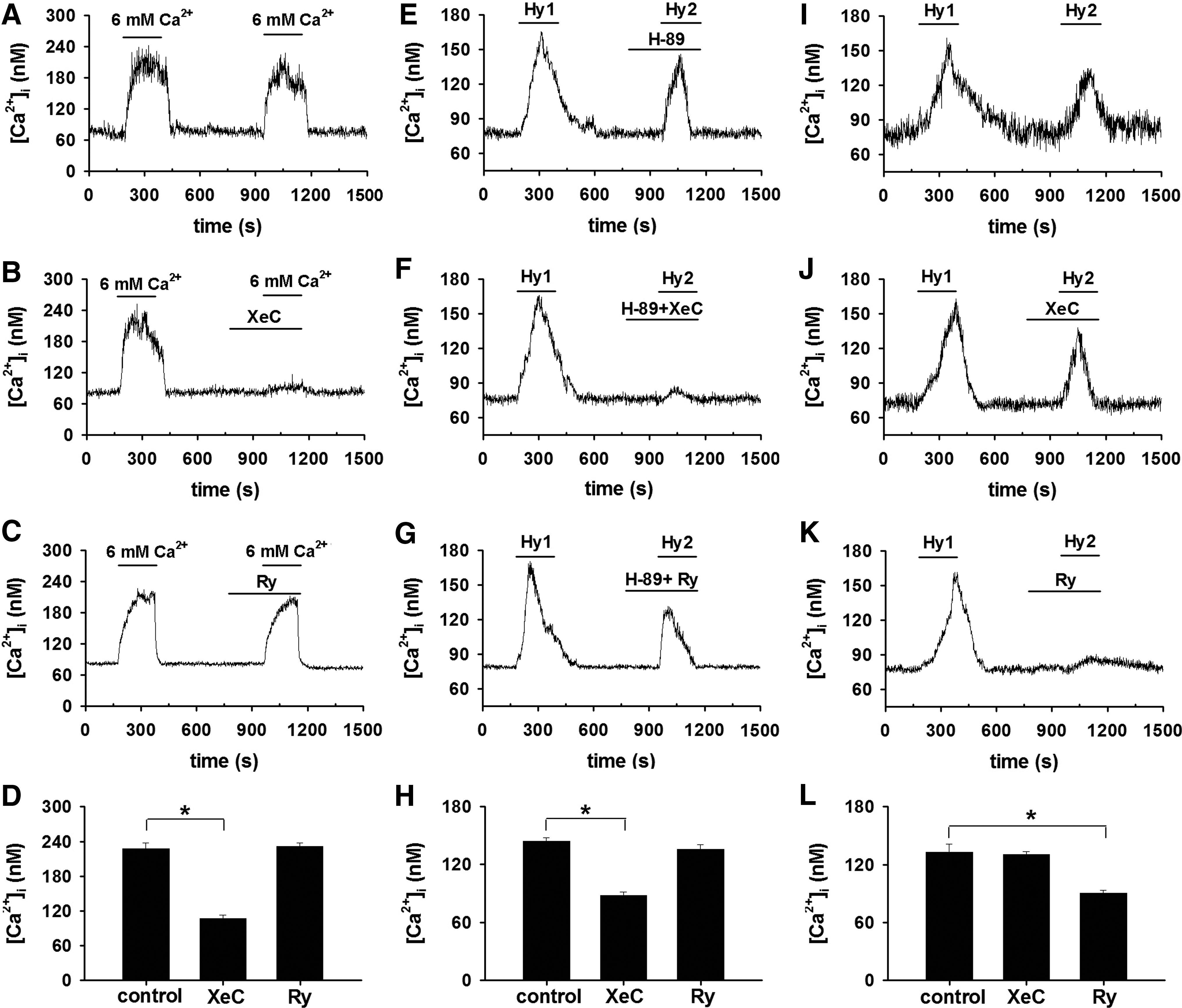
Under normal or normoxic conditions, CaSR activation (by elevating [Ca2+]o to 6.0 mM) -induced [Ca2+]i signal in PASMCs was inhibited by XeC (Fig. 7A–D), indicating that CaSR activation-induced [Ca2+]i signal is mediated by IP3 receptor, the well-established downstream target of CaSR (14). However, the downstream target turns to be RyR under hypoxic condition in PASMCs (Fig. 6). To reveal the discrepancy, the role of protein kinase A (PKA) pathway was explored. XeC did (Fig. 7F, H), whereas ryanodine failed to (Fig. 7G, H), inhibit HICI in the presence of H-89, a PKA inhibitor, indicating that IP3 receptor mediates HICI, while PKA signaling pathway is blocked. On the contrary, ryanodine did, whereas XeC failed to, inhibit HICI in the absence of H-89 (Fig. 7I–L), implying that rynodine receptor mediates HICI, while PKA signaling pathway is not blocked.
HPV: role of CaSR in vitro and in vivo
Phenylephrine (PE)-preconstricted PA rings showed no response to acetylcholine confirming the removal of endothelium; however, they contracted when each hypoxic challenge was applied to the vessels (Fig. 8Ai). Calhex231 dose-dependently inhibited the hypoxia-induced constriction of the vessel (four parameter logistic regression, p<0.05, Fig. 8Aiv). 15.6 μM H2O2 contracted rings in a way similar to hypoxia (Fig. 8Bi) that was also inhibited by 1 μM Calhex231 (n=5, p<0.01, Fig. 8Bii, Biii), suggesting the mechanistic role of H2O2 in sensing CaSR and the subsequent constriction of PA by hypoxia. Furthermore, the hypoxia-induced constriction of PA isolated from CaSR knockdown rats was profoundly inhibited as compared with control (n=3, p<0.01, Fig. 8C).
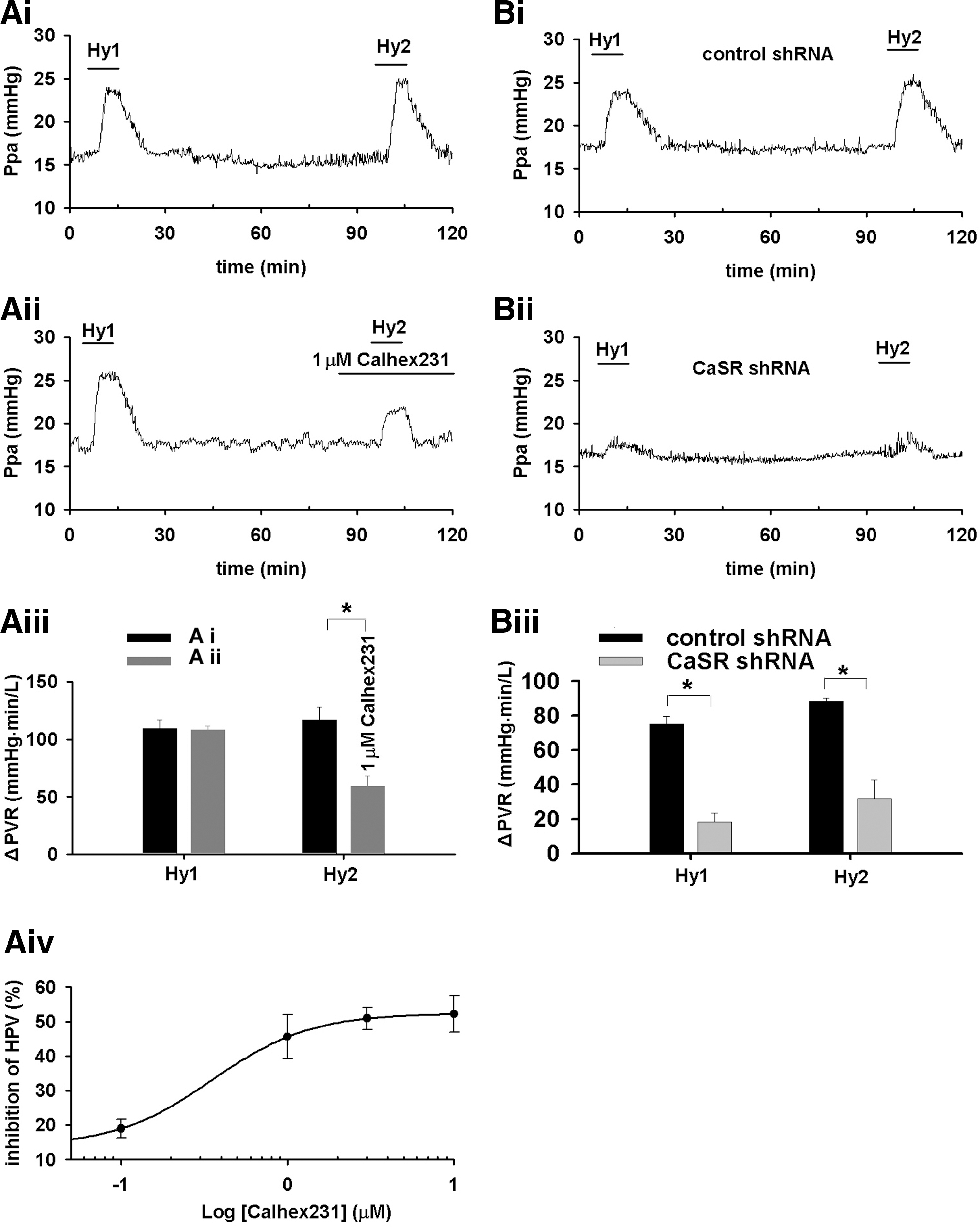
The 10 min episodes of hypoxia successively triggered typical HPV in rats (n=4, Fig. 9Ai), which was dose-dependently inhibited by Calhex231 (four-parameter logistic regression, p<0.05, Fig. 9Aiv). HPV was found to be typical in control rats treated with nonspecific shRNA (n=3, Fig. 9Bi), whereas significantly inhibited in CaSR knockdown rats (n=4, p<0.01 vs. control, Fig. 9Bii, Biii). CaSR knockdown did not affect PE-induced pulmonary vascular constriction in rats (not shown) or plasma Ca2+ level (Supplementary Fig. S6).
Discussion
The basal function of CaSR in PASMCs was first evaluated by elevating the extracellular Ca2+ to supraphysiological levels, the direct stimulus of CaSR (14) (Fig. 1). In the subsequent experiments revealing the actual role of CaSR in HPV, the extracellular Ca2+ was maintained at a physiological level.
Hypoxia induces an [Ca2+]i increase (HICI) in PASMCs that depends on extracellular Ca2+ (Fig. 2). These results are broadly consistent with most of the previous observations well documented in literature including those on isolated or cultured PASMCs (9, 25, 34, 35), isolated pulmonary arteries (PAs) (13, 19, 29), and perfused lung or lung slices (10, 42). In the absence of extracellular Ca2+, however, hypoxia was reported to cause a small and transient increase followed by a sustained decrease in [Ca2+]i below baseline in canine PASMCs (22, 23), a small increase in rat PASMCs (25) or transient(s) in 34% of rat PASMCs examined (27). A transient contraction of rat PA rings by hypoxia in Ca2+-free medium was also noted (12). This discrepancy can be due to differences in experimental preparations (3, 32) and heterogeneity of [Ca2+]i response to hypoxia in PASMCs (27). Extracellular Ca2+ was shown to differentially affect hypoxic vasoconstriction in different species (32), and hypoxia also differentially affected [Ca2+]i in PASMCs isolated from different regions of PA tree (3). Additional and potential reasons can be the spontaneous Ca2+ release in the absence of extracellular Ca2+ reported in a variety of cell types including vascular smooth muscle cells (31, 44) and endogenous peroxidase activity. Generally, the extracellular Ca2+ dependence of HICI was understood as the importance of Ca2+ influx in all previous studies. The present study, however, showed further that both Calhex231 and CaSR knockdown inhibited HICI in PASMCs (Figs. 2 and 3). This provides, to the best of our knowledge, the first clue of the CaSR involvement in HICI and HPV. These novel findings further imply that the extracellular Ca2+ dependence of HICI cannot be simply or merely interpreted as the resource of Ca2+ influx as presumed. An additional role of extracellular Ca2+, the ligand of CaSR can be an initial trigger for the activation of CaSR during hypoxia and the subsequent [Ca2+]i elevation and constriction.
The actual role of ROS in HPV remains controversial, and even both increased and decreased ROS by hypoxia were documented as recently reviewed (33, 37, 38). The present study is consistent with those showing increased ROS by hypoxia (10, 26, 39 –41). The oxidation of mitochondria-localized RoGFP observed here also suggests the origin of ROS, and this is generally consistent with the previous study showing increased oxidation of RoGFP in the intermembrane space but decreased oxidation of RoGFP in the mitochondrial matrix (40). H2O2 at the level achieved by hypoxia increased [Ca2+]i in the presence of extracellular Ca2+, which was diminished by the removal of extracellular Ca2+, the presence of Calhex231, or CaSR knockdown (Fig. 4). These results suggest that ROS elevated by hypoxia cannot directly stimulate intracellular Ca2+ release by itself and that H2O2 induces [Ca2+]i elevation through CaSR in PASMCs. It also implies that the Ca2+ sensitivity of CaSR should be enhanced sufficiently by the H2O2 level or oxidized status achieved by hypoxia for mediating [Ca2+]i elevation in the presence of constant extracellular Ca2+ level (without elevating [Ca2+]o as the stimulus of CaSR). To better describe this process mechanistically, the present study adopts the expression of “sensing/sensitization,” a concept usually used to indicate an altered activity of ion channel under oxidized/reduced status underlying HPV (33, 43) Thus, H2O2 at the level achieved by hypoxia induces [Ca2+]i signaling in PASMCs through sensitizing CaSR. The mechanism underlying how hypoxia/ROS sensitizes CaSR remains unclear. One speculation can be the regulation of CaSR dimerization by disulfide bridge(s), although it is not an absolute requirement (2, 47).
The mitochondria-derived H2O2 was validated to sensitize CaSR and trigger HICI using mitochondria depletion strategy (Fig. 5). Ryanodine at the concentration inhibiting RyR or SOC inhibitor, but not IP3 receptor inhibitor, significantly attenuated HICI (Fig. 6) that confirms previous reports (4, 8, 11, 15, 23, 34, 35, 48). An additional finding from the present study is that ryanodine/RyR3 knockdown and SOC inhibition/STIM1 knockdown similarly attenuated 15.6 μM HICI in PASMCs (Fig. 6), suggesting that RyR3 and STIM1 are likely the key components or downstream targets underlying CaSR activation or sensitization-triggered [Ca2+]i signaling during hypoxia. In other words, the pivotal role of sensitized CaSR is to arrange or direct the mitochondria-derived H2O2, extracellular Ca2+, the intracellular Ca2+ release pathway of RyR3, and the STIM1-participated extracellular Ca2+ influx pathway of SOC to orchestrate the [Ca2+]i elevation during hypoxia in PASMCs. However, this study does not exclude the role of voltage-sensitive channels and myofilament Ca2+ sensitivity in HICI and HPV (1, 25, 28, 43). The inhibitory effect of ryanodine on HICI was swapped to XeC by the presence of a PKA inhibitor (Fig. 7), indicating that the switch of downstream target of CaSR activation from IP3 receptor under normal/normoxic condition (14) to RyR under hypoxia is possibly related with PKA.
HPV is typically biphasic, an initial and transient one followed by a sustained constriction (4, 11, 16, 28, 29). The critical role of CaSR revealed in the current study appears applicable mainly to the initial phase and may also be involved in the sustained constriction (Supplementary Fig. S7).
Calhex231 and CaSR knockdown inhibited hypoxia-induced constriction of PA (Fig. 8). This is consistent with that CaSR activation in vascular smooth muscle cells can induce [Ca2+]i elevation (17) and cause constriction of the gerbilline spiral modiolar artery (45) and that hypoxia- and peroxide-induced increase in [Ca2+]i and contraction are intimately related in PASMCs (28, 29). Calhex231 also inhibited H2O2-induced PA constriction (Fig. 8), functionally confirming the signaling role of H2O2 in sensing CaSR, which, in turn, triggers PA constriction during hypoxia. The dose-dependent inhibition of HPV by Calhex231 was noted in rats, and the critical role of CaSR was further validated by the attenuated HPV in CaSR knockdown rats (Fig. 9). The effect of Calhex231 revealed in the current study appears to be specific, as it does not affect PE, high K+, or angiotensionII-induced PA constriction (Supplementary Figs. S8 and S9). Taken together, these results provide functional evidence for the pivotal role of CaSR-mediated Ca2+ signaling in HPV, as illustrated in the mechanistic flow chart next (Fig. 10).
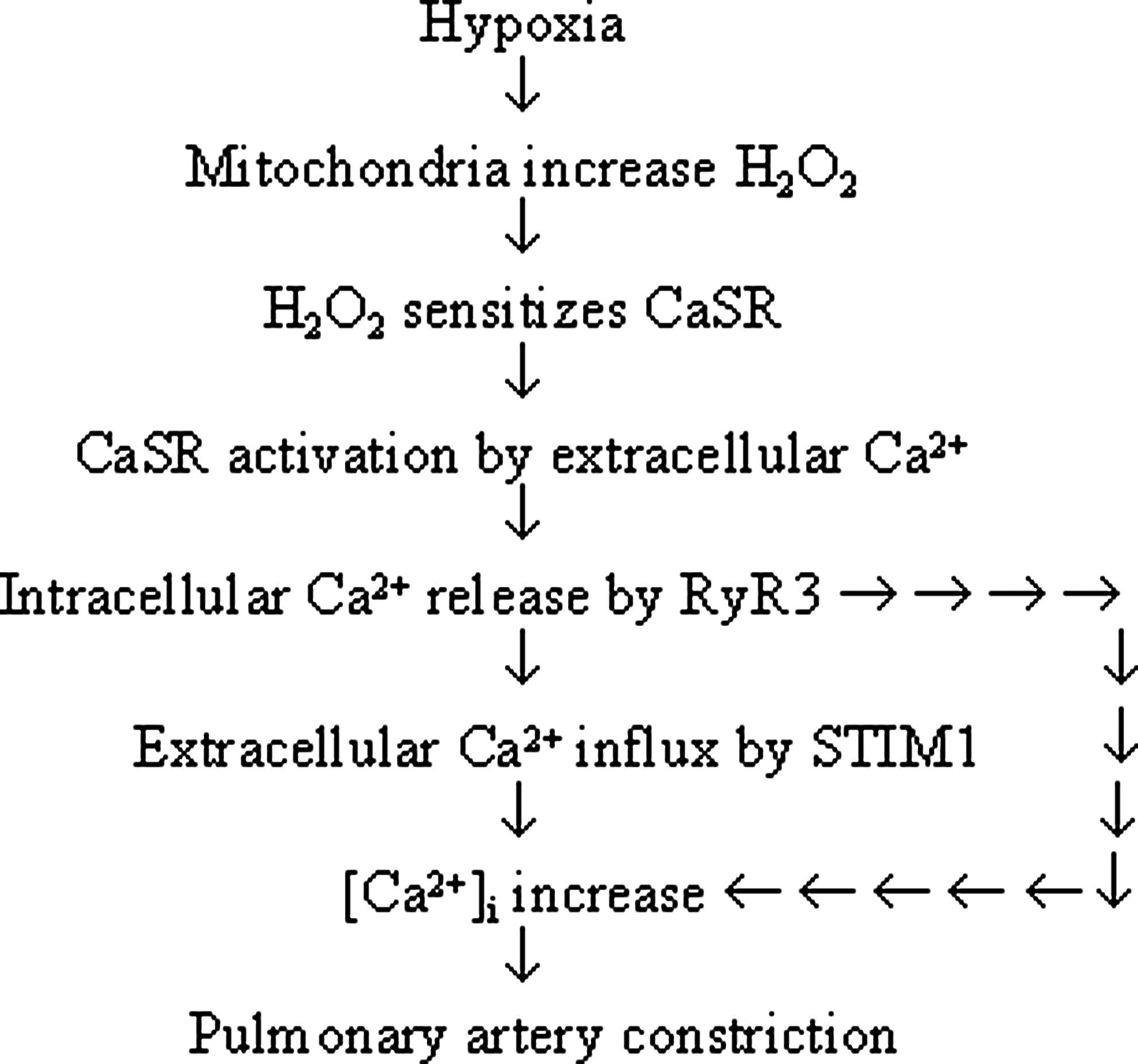
The blockage of hypoxia-triggered Ca2+ signaling in PASMCs by loading PEG-catalase and the essential role of CaSR in hypoxia-triggered Ca2+ signaling and HPV (Figs. 2 –4, 5C, and 6 –9) may warrant future studies to determine whether CaSR activity can be modulated by endogenous peroxidase and whether CaSR manipulation can potentially serve as a therapeutic target for HPV-related diseases such as hypoxic pulmonary hypertension and edema.
Materials and Methods
Cell culture
All experiments involving Sprague-Dawley rats were approved by the Institutional Animal Care and Use Committee.
Rat PASMCs were cultured from the third branches of intralobar artery explants and maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (7, 36). The PASMCs at ∼70% confluence in 1–2 passages were employed in experiments. To deplete mitochondrial DNA, PASMCs were treated with 110 μg/ml sodium Py, 50 μg/ml Ur, and 200 ng/ml ethidium bromide as fully described earlier (7). PASMCs were isolated from at least three rats for each kind of experiment.
[Ca2+]i measurement
[Ca2+]i measurements were performed in 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid-buffered saline (HBS) containing (mM): NaCl 135, KCl 5.0, CaCl2 2.0, MgSO4 1.2,
Intracellular ROS measurement
The detection of intracellular ROS using H2DCFDA and its calibration exploiting the DCFDA oxidation speed versus extracellular application of a series of H2O2 were previously described in detail (7, 30, 49). The mitochondrial targeting RoGFP was generated from pAcGFP1-Mito vector (Clontech) by incorporating four mutations (C48S, Q80R, S147C, and Q204C) in GFP coding region (40) using a QuikChange Multi Site-directed mutagenesis kit (Stratagene), and the mutations were confirmed by sequencing. After transfection with Lipofectamine 2000, RoGFP images in PASMCs were obtained under a confocol microscope (OLYMPUS IX71) with excitation of 405 or 488 nm and emission at 515±10 nm corresponding to the oxidized and reduced status, respectively. The ratios of fluorescence intensity from excitation at 405 and 488 nm were analyzed using FLUOVIEW V.5.0 software to quantify the extent of oxidative status.
Delivery of shRNA
Effective shRNA specifically against CaSR, STIM1, RyR3, and nonspecific control (Origene) were transfected into PASMCs using Lipofectamine 2000 as previously described (7). The shRNA constructs simultaneously encode GFP for identification of transfected PASMCs individually. The oligo-DNA encoding the control and the effective shRNA targeting CaSR were subcloned into the Lenti-X shRNA expression vector (Clontech), respectively, confirmed by sequencing and packaged into lentiviral particles using Lenti-X HTX packaging system and 293T cells (Clonetech). 72 h before hemodynamic experiments, 1 ml lentivirus (∼106 TU/ml) was intravenously injected into each rat.
Immunochemical staining and western blot
To determine the expression of CaSR in PASMCs and endothelium-removed PAs, immunostaining and western blot were conducted as previously described in detail (7, 50) using an anti-CaSR antibody (Pierce Biotechnology).
Measurement of force
The endothelium-removed rings of intralobar PA were mounted in a wire myograph immersed in individual water-jacketed chambers containing physiological salt solution (118 NaCl, 24 NaHCO3, 1 MgSO4, 0.44 NaH2PO4, 4 KCl, 5.5 glucose, and 1.8 CaCl2 mM, and pH 7.35–7.45), bubbled continuously with 95% O2-5% CO2 at 37°C, and stretched to a predetermined optimal passive tension of 500 mg. Force displacement was recorded using a PowerLab/4SP data acquisition system (ML118, AD Instruments). Each ring was initially contracted by 0.1 μM PE and subsequently challenged by 0.01 μM acetylcholine to confirm its contractility and endothelium nonintegrity, respectively. Since a small pretone by agonist can facilitate the hypoxic response in rat intralobar PA (16, 28), the rings were contracted by 0.1 μM PE 5 min before and during the hypoxic challenge. In some experiments, oxygen tension in the myograph chamber was simultaneously monitored via an isolated dissolved oxygen meter and electrode (ISO2, World Precision Instrument). Hypoxia was induced to a stable PO2 level of 33 mmHg in PSS close to the rings within ∼1.7 min by continuously bubbling with 95% N2-5% CO2.
Hemodynamic measurements
Rats were anesthetized with urethane (1.2 g/kg ip), and a catheter was inserted in the trachea and connected to a ventilator supplying room air or a 10% O2-90% N2 gas mixture. To continuously monitor pressure of pulmonary artery, a PE-50 tubing with a curved tip was passed from the right jugular vein into the main PA. The catheterization was also performed for the measurement of left ventricular end-diastolic pressure, and cardiac output (CO) was measured using the thermodilution method with the CO pod (ML313C, AD Instruments) and the PowerLab/4SP data acquisition system. A 200 μl of room temperature 0.9% NaCl solution was injected into the left jugular vein catheter with its tip near the right atrium, and changes in blood temperature were detected by a T-type ultra-fast thermocouple probe (MLT1402, Physitemp) positioned in the aortic arch from the right carotid artery.
Statistical analysis
Data are reported as means±standard error. The Student's t-test and one-way analysis of variance analysis were made for two or multiple group comparisons, respectively. A difference was considered significant at p<0.05.
Footnotes
Acknowledgments
The research was supported by 973 program-2012CB518201, NSFC grants (81170048, 30971162, 30670926 and 30770940), and a grant from key laboratory of high altitude medicine of MOE.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.