
Abstract
Introduction
This essay focuses on how H2O2 is produced, metabolized, and ingeniously utilized in the endoplasmic reticulum (ER) to generate and maintain an efficient and environment friendly protein factory.
Oxidative Protein Folding and H2O2 Production in the ER
In all eukaryotic cells, the ER is responsible for the folding of membrane and secretory proteins. Unlike those destined to the cytosol or nucleus, the majority of these proteins contain disulfide bonds, the formation of which requires net oxidative power. In the late ’90s, the discovery of ER oxidoreductin-1 (Ero1) proteins provided an answer to how oxidative power can be generated in the ER (15, 43). These highly conserved flavoproteins oxidize protein disulfide isomerase (PDI) preferentially over other resident oxidoreductases (33). PDI then transfers disulfide bonds to cargo proteins. Thus, oxidative protein folding is achieved by step-by-step reactions involving several enzymes, an important feature to guarantee selectivity of oxidation. Concomitantly, electrons flow from Ero1 to molecular oxygen, yielding H2O2 as a by-product in stoichiometric amounts to the disulfides formed (54) (Fig. 2A). Two members of the family are present in mammals, Ero1α and Ero1β, with different tissue distribution and transcriptional control. The reader is referred to excellent reviews in this Forum for updated information on the evolution and regulation of the Ero1 family (3, 44). In yeast, ERO1 is an essential gene. The unexpectedly mild phenotype of Ero1α and Ero1β compound mutant mice, therefore, raises the question of what sustains oxidative folding in the absence of these flavoproteins (60). Possibly disulfide bonds can be inserted into proteins by sulfydryl oxidases as overexpressed Erv2 can rescue ero1-defective yeast cells (49). Interestingly, besides a flavin-containing domain endowed with oxidase activity, quiescin-sulfhydryl oxidase contains a thioredoxin-like domain through which it can directly transfer disulfide bonds to its substrates (Fig. 2B). Unlike Ero1 proteins, these enzymes are normally secreted by cells and are hence present in the entire exocytic compartment (25). It is presently unclear whether mammalian sulfydryl oxidases act also downstream of the ER and/or in the extracellular space.

The above flavoprotein-driven reactions sustaining disulfide bond formation are all associated with the production of luminal H2O2. Moreover, other systems can generate or supply H2O2. For instance, NADPH oxidase-4 (Nox4) has been reported to reside in the ER membrane: this enzyme produces H2O2 without generating O2•− (Fig. 2C) and has been implicated in activating apoptosis after prolonged ER stress (58). Another potential source of oxidants might be mitochondria, where the respiratory chain can produce large amounts of H2O2. Close contacts are established between ER and mitochondria, at the level of the mitochondrial-associated membranes (MAM) (Fig. 2D) (4, 16). However, ero1-deficient murine cells did not show clear sensitivity to attenuation of mitochondrial activity (61).
H2O2 is Scavenged and Metabolized by ER Peroxiredoxins
Particularly in professional secretory cells, oxidative folding can become a major source of H2O2. A single plasma cell, for instance, inserts up to 105 disulfides per second into nascent immunoglobulin (6). What is the fate of the H2O2 generated in the process of oxidative folding, a potentially dangerous by-product? An important hint came when it was shown that H2O2 can promote disulfide bond formation in vitro with minimal oxidative damage to the substrate protein (24). It was then found that peroxiredoxin-4 (Prx4), a protein residing primarily in the early secretory compartment, can utilize H2O2 to catalyze PDI oxidation in vitro and in vivo (52, 61). This property seems to be shared also with glutathione peroxidases-7 and -8 (GPx7/8), two enzymes that like Prx4 reside in the ER (37). The physiological and structural aspects of these peroxidases are addressed in more details below.
The Prx Protein Family
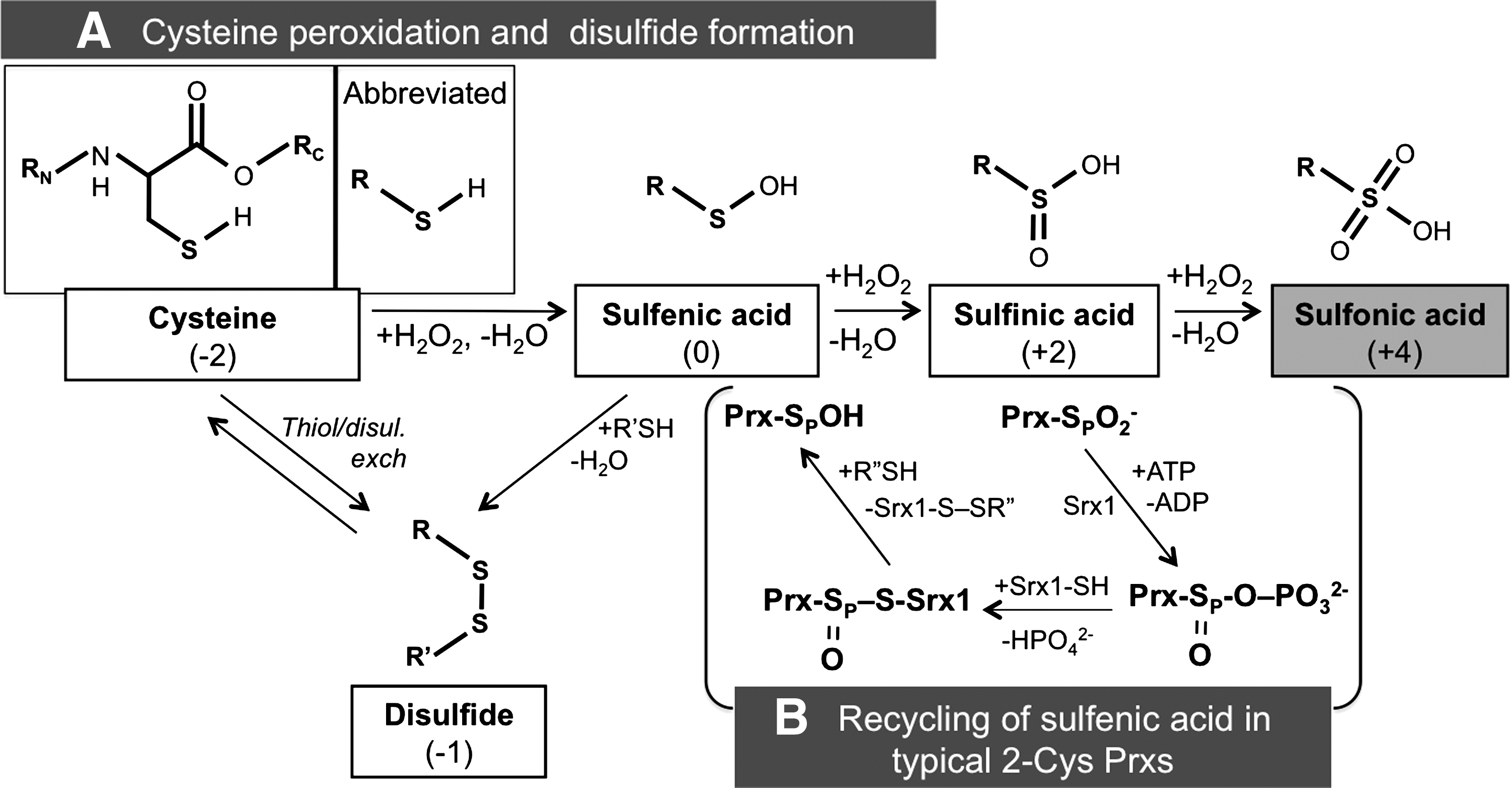
The first member of the Prx family proteins was identified in yeast in 1994 (11). Prxs are ubiquitous enzymes: in almost all organisms they can reduce different peroxides at very high rates (106–107 s−1M−1) (18). They are hence one of the main lines of defense that cells deploy against excess peroxide production. There are six members of the Prx family in the human genome, residing in different compartments (46). Prxs can be divided into three classes: typical 2-cysteine (2-Cys), atypical 2-Cys, and 1-cysteine (1-Cys) Prx. Their mode of action includes similar cycles, which are schematized in Figure 3. The peroxidatic cysteine (CP) initially reacts with H2O2 generating a water molecule and a sulfenic (SOH) intermediate. In 2-Cys Prxs, the cysteine sulfenic acid can then react with a resolving cysteine (CR) to form a disulfide bond. In atypical 2-Cys enzymes an intra-chain disulfide is formed, while in typical 2-Cys Prx the resolving cysteine is located in an adjacent molecule (Fig. 3). The covalent Prx dimers are eventually reduced by thioredoxin or other reductases through thiol/disulfide exchange reactions. In the presence of excess H2O2, however, the SOH moiety can be further modified to the sulfinic and sulfonic derivatives (SO2H or SO3H). While the SO2H form can be reversed to sulfenic acid by the catalysis of sulfiredoxin-1 in an ATP-dependent manner (8), SO3H species are irreversibly inactivated. This mechanism, known as the floodgate model (17), may ensure that sufficient amounts of H2O2 locally accumulate to allow productive signaling.
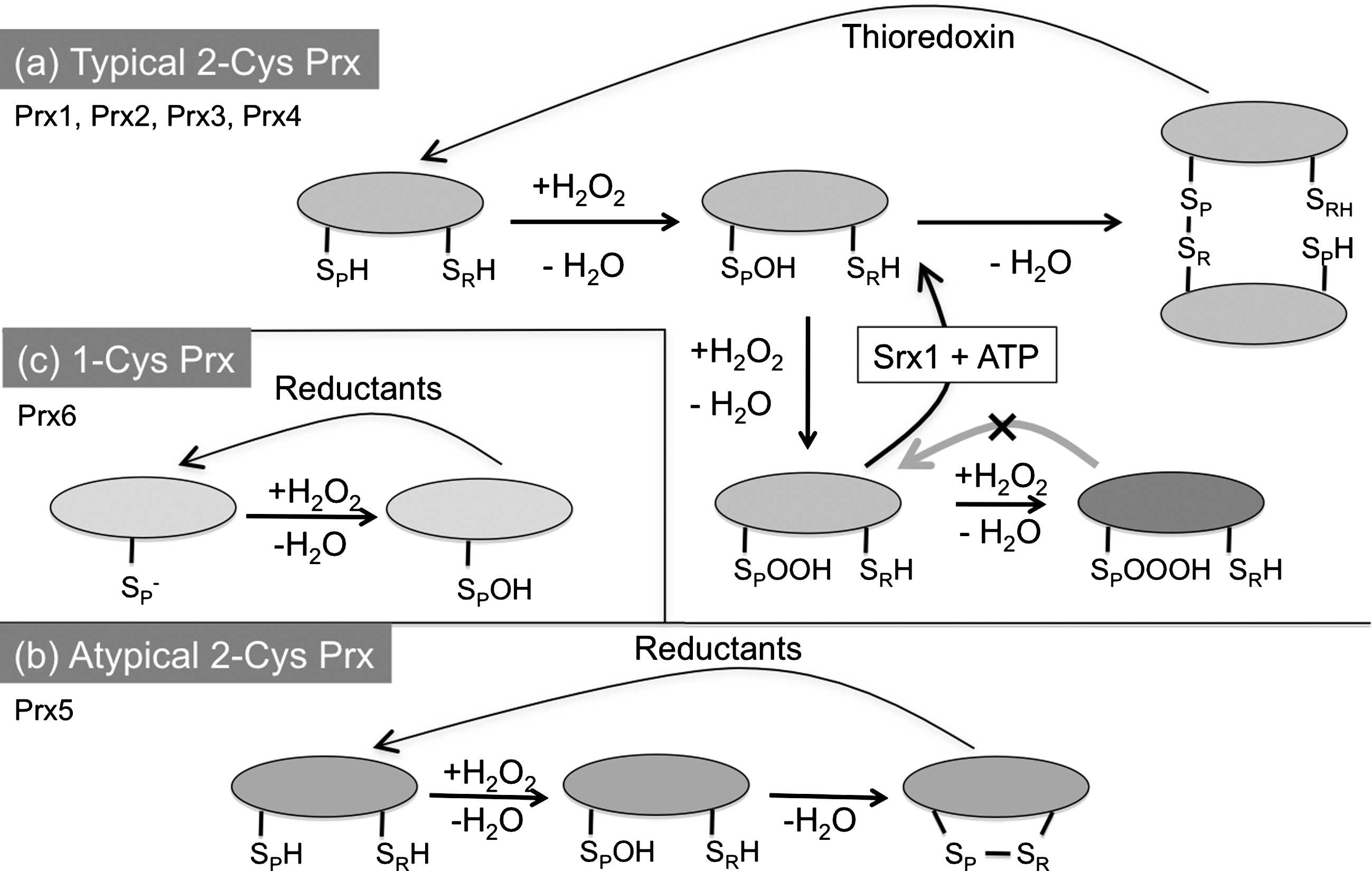
Upon hyper-oxidation, Prxs may form decamers or higher order assemblies, and have been reported to lose their peroxidase activity and acquire chaperone activity (17). In this way, Prx2 can induce cell cycle arrest (42). In view of the importance of H2O2 in signaling, it is not surprising that many eukaryotic 2-Cys Prxs contain two motifs, the GGLG and YF sequences that decrease the flexibility of the peroxidatic cysteines, making them more susceptible to hyper-oxidation (57).
Prx4, a Typical 2-Cys Prx Residing in the Secretory Pathway
Prx4, also known as AOE372, AOE372-2, or TRANK (thioredoxin peroxidase-related activator of NF-κB and c-Jun N-terminal kinase), is a typical 2-Cys Prx, comprising catalytic and noncatalytic cysteines (51). Prx4 has several interesting and unique features. First, it differs from other 2-Cys Prxs in that it also utilizes reduced glutathione (GSH) as a reducing agent in addition to Trx (39). This is an important feature, because Trx is not present in the secretory pathway. However, oxidized Prx4 clearly transfers disulfide bonds much more efficiently to PDI (52, 61), ERp46 (10) and possibly other ER oxidoreductases than to GSH (Fig. 4A). This feature places Prx4 in the dual role of ER resident antioxidant and promoter of oxidative folding. Notably, an oxygen molecule can produce two disulfide bonds in the ER without H2O2 release, through the sequential oxidation of two oxidoreductases, that is, Ero1 and Prx4 (Fig. 4B).
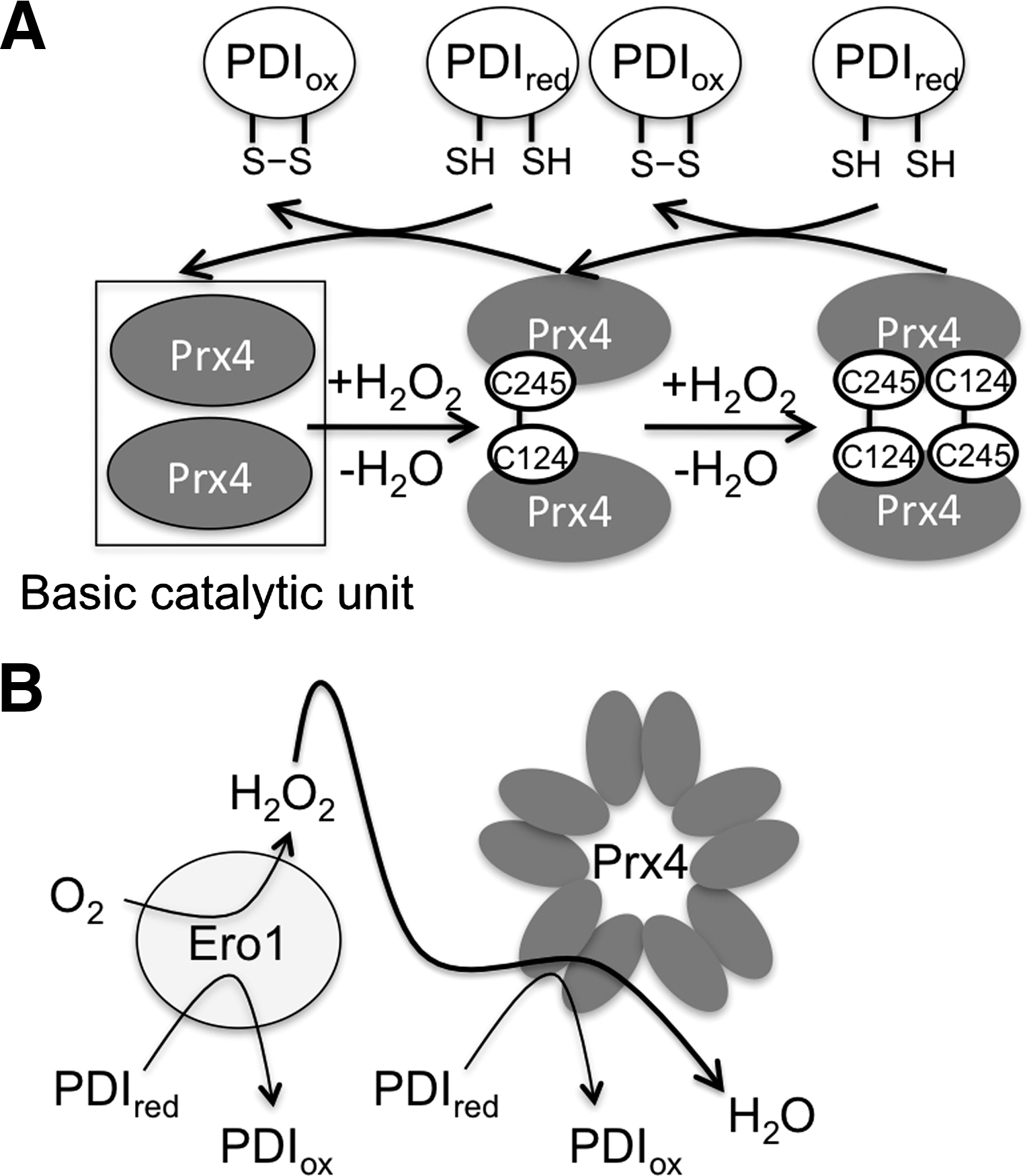
Structure and Oligomerization of Prx4
Unlike other members of the Prx family, Prx4 has an additional N-terminal loop, putatively directed toward the center of the decamer rings (10, 55) (Fig. 5). A cysteine residue (Cys51) in the loop can form interchain disulfide bonds (51), which, in conjunction with those formed between the peroxidatic and resolving cysteines (Cys124 and Cys245, respectively) as part of the catalytic cycle (Fig. 4A), generate covalent decamers. Also Prx1, another typical 2-Cys Prx, forms a noncatalytic interchain disulfide bond that involves Cys83 and stabilizes decamers. In this conformation, Prx1 switches from peroxidase function to a molecular chaperone activity (26). However, Cys83 of Prx1 shows more similarity to Cys148 of Prx4, which was found irrelevant in covalent oligomer formation, than to Cys51 (51). The crystal structures of Prx4 in three different redox states confirmed that the N-terminal loop is important for stabilization of decamers, independently from interchain bonding via Cys51 (55). Accordingly, replacing Cys51 for alanine did not prevent decamer formation (10).
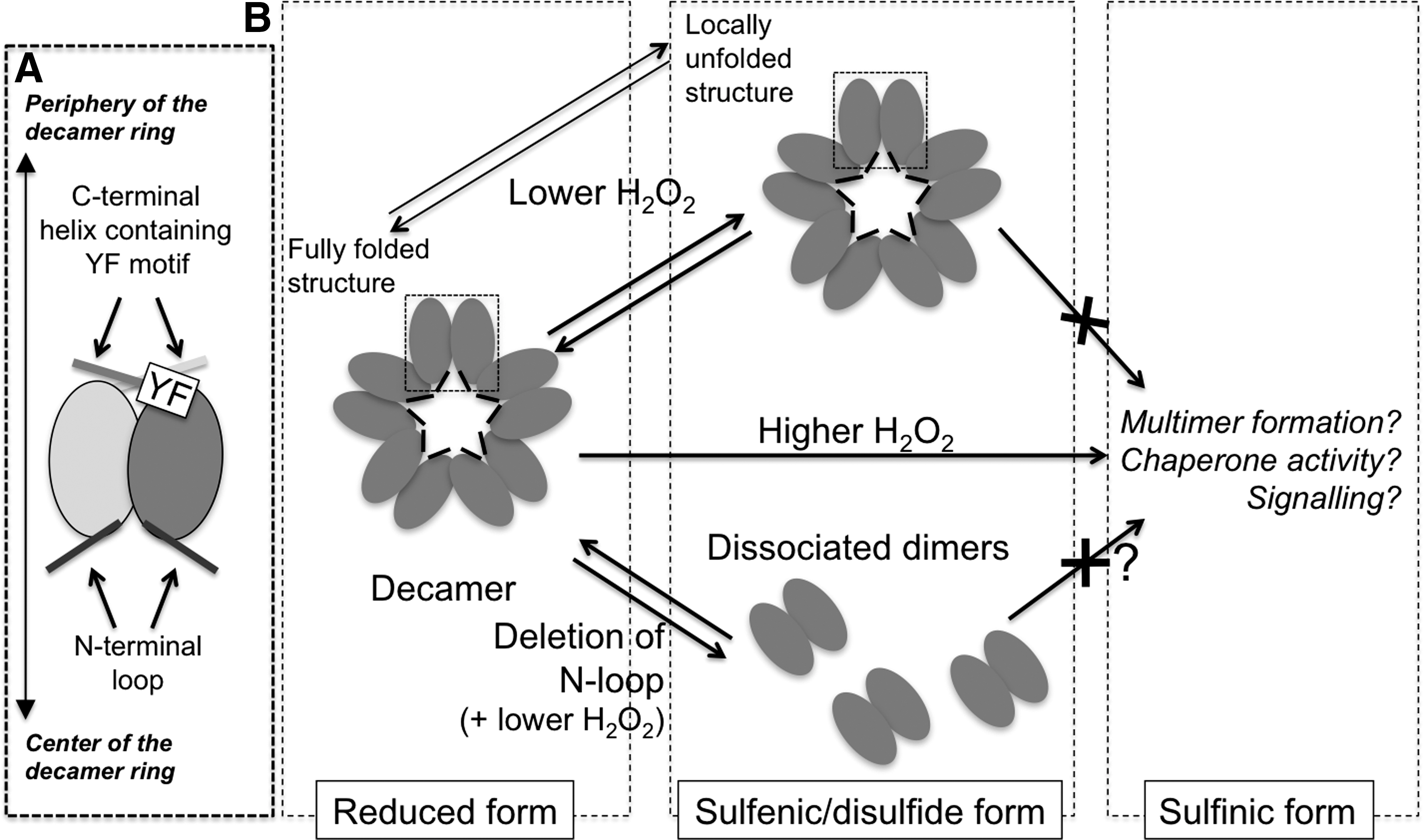
Regulating Prx4 Hyper-Oxidation
In cytosolic Prxs, sulfiredoxin 1 reverts sulfinylated subunits restoring their antioxidant properties. Is Prx4 hyper-oxidation differently regulated in the ER, where sulfiredoxin 1 is absent? The oxidized species accumulating after reaction with one H2O2 molecule, Prx4-SOH or Prx4-Prx4 (via C124-C245), undergo conformational changes that yield a locally unfolded structure (Fig. 5B, upper). Of note, and somehow counterintuitively, a Prx4 mutant lacking the resolving cysteine (C245A) was found to be resistant to hyper-oxidation even when exposed to high concentrations of H2O2 (10). Cao et al. found that this locally unfolded structure reacts less efficiently with H2O2, limiting hyper-oxidation (10) (Fig. 5B). It was also shown that either deleting the YF motif or mutating a single amino acid (T155E) prevented Prx4 hyper-oxidation by enhancing the mobility of C124 or the dissociation of Prx4 decamer to dimers, respectively (55). These findings suggest that the susceptibility of human Prx4 to hyper-oxidation depends on multiple factors, such as (i) the stability of the enzyme active site, (ii) decamerization and (iii) the C-terminal YF motif limiting the mobility around C124. They also indicate that Prx4 retained structural features conductive to its hyper-oxidation, even though no sulfiredoxin activity has been demonstrated so far in the exocytic compartment. This raises intriguing questions as to the fate and possible significance of sulfinylated or sulfonylated forms in the ER, presumably inactive as H2O2 scavengers and oxidants of PDI. Do these species display increased chaperone activity? Are they degraded upon restoration of redox homeostasis in the ER? Or are they perhaps preferentially secreted, acting as stress or damage associated molecules?
Isoforms, Topology, and Physiological Aspects of Prx4
Contrasting reports appeared in the literature concerning the subcellular localization of Prx4. Prx4 was initially identified as a cytosolic protein that regulates NF-κB (23). Soon thereafter, however, Haridas et al. described it as a secreted protein with a cleaved N-terminal signal sequence (19). Much remains to be understood on the precise Prx4 localization, which has been shown to depend on alternative transcription initiation and proteolytic processing. Thus, the 27-kDa species is a soluble protein accumulating in the secretory compartment of most cell types, derived from cleavage of the N-terminal signal peptide encoded in exon 1B (Fig. 6). The 31-kDa form instead is a membrane-anchored protein retaining the signal sequence. This isoform is expressed in testis after puberty, being particularly abundant in the acrosomal membrane of spermatids but not in differentiated sperm. A recent study described yet an additional 29.5-kDa isoform, in which exon1-B is replaced by exon 1-A (59). The latter exon does not encode a hydrophobic N-terminal signal peptide. Accordingly, the 29.5-kDa variant was found to accumulate in the soluble fraction of mice testis homogenates, suggesting a cytosolic localization. Its functional role remains to be established.
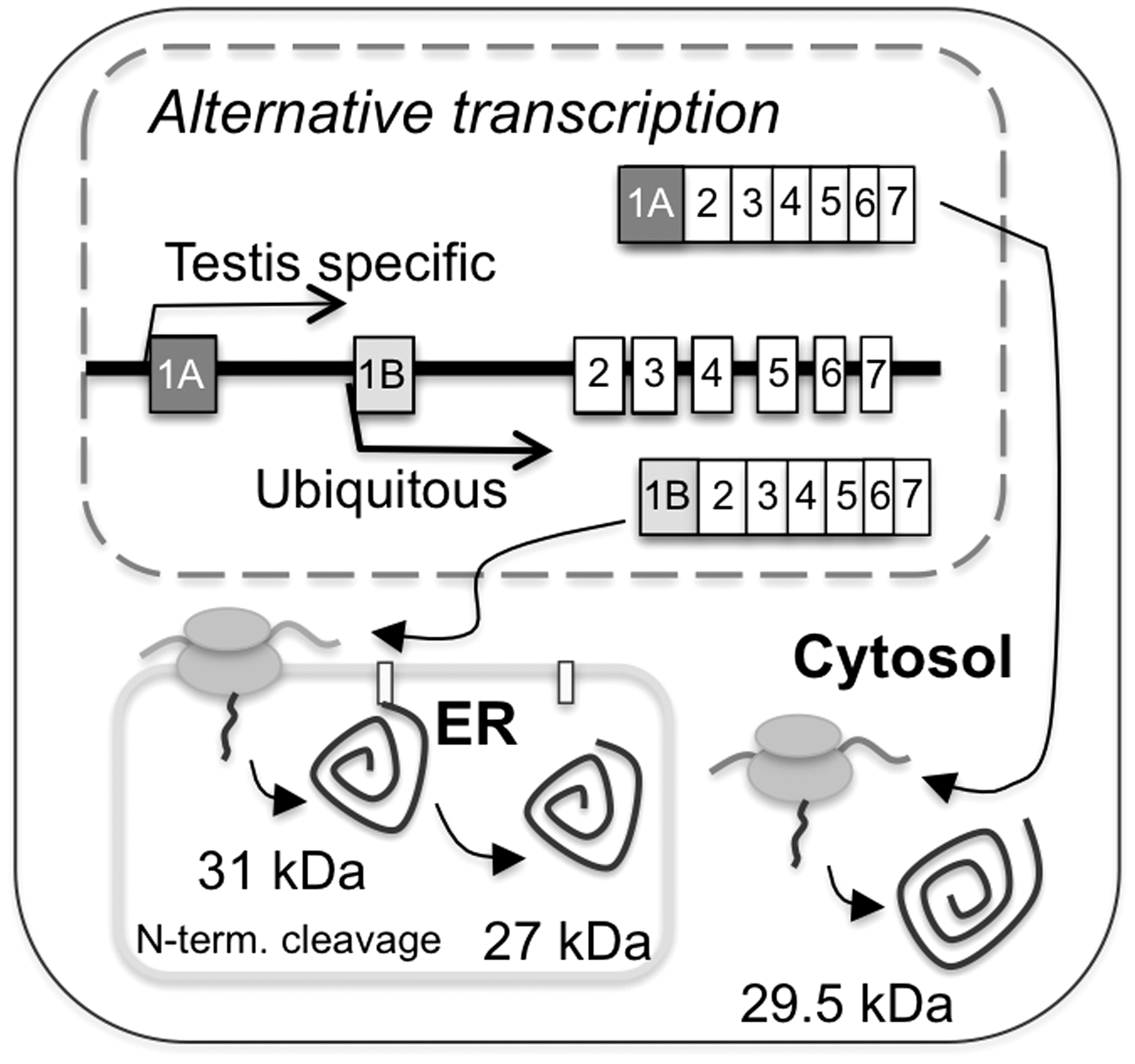
The presence of membrane-bound and soluble isoforms seems to be a feature common to various ER-resident enzymes. For instance, ER degradation-exhancing alpha-mannosidase-like-1 (EDEM1), a lectin-like molecule accelerating ER-associated degradation, exists as a type II transmembrane protein in addition to the signal peptide-cleaved, soluble isoform. Interestingly these two isoforms have different substrate specificities (50). Similarly, transmembrane PDI family members show preferences for other client proteins than soluble PDIs (48). They may derive oxidative power from Ero1α and Ero1β, a fraction of which associates with membranes too (41). It will be of interest to determine whether membrane bound Prx4 serves a discrete role in ER redox homeostasis. In this connection, the ubiquitous, soluble 27-kDa Prx4 isoform lacks a C-terminal ER retrieval motif. Interestingly, mammalian Ero1 molecules also lack known ER localization signals. Being intracellularly retained by virtue of their interactions with ER oxidoreductases, primarily PDI and ERp44, Ero1α and Ero1β are secreted when overexpressed (40). Further experiments are needed to determine how Prx4 maintains its subcellular localization, and whether its retention/secretion can be regulated in living cells.
Differentiating B cells strongly upregulate Prx4 as they embark on massive antibody production (6). Surprisingly, however, mice lacking the soluble 27-kDa Prx4 isoform had only minor defects in polymerization and secretion of IgM, a molecule harboring many disulfide bonds. These mice could produce the 29.5-kDa isoform; however, this presumably cytosolic Prx4 variant is thought to be specifically expressed in the testis (55). Therefore, the subtle phenotype of these mice implies redundancy in the ER antioxidant systems. GPx7 and GPx8 are good candidates to replace Prx4 in scavenging luminal H2O2 in a productive way.
GPx-7/-8, Members of Nonselenium GPxs
Discovered in 1957 by Mills (36), GPx orthologs have been found in archaea, prokaryotes, and eukaryotes (53). The activity of many GPxs depends on selenocysteine, a rare amino acid residue similar to cysteine, in which a selenium (Se) replaces sulfur in its selenothiol group. Recent reports identified also nonselenium GPxs with a cysteine at the position normally occupied by a selenocysteine residue. Biochemical studies revealed that most nonselenium GPxs depend on thioredoxin rather than glutathione (21). The working cycle of seleno-GPx and nonselenium GPx are schematized in Figure 7.
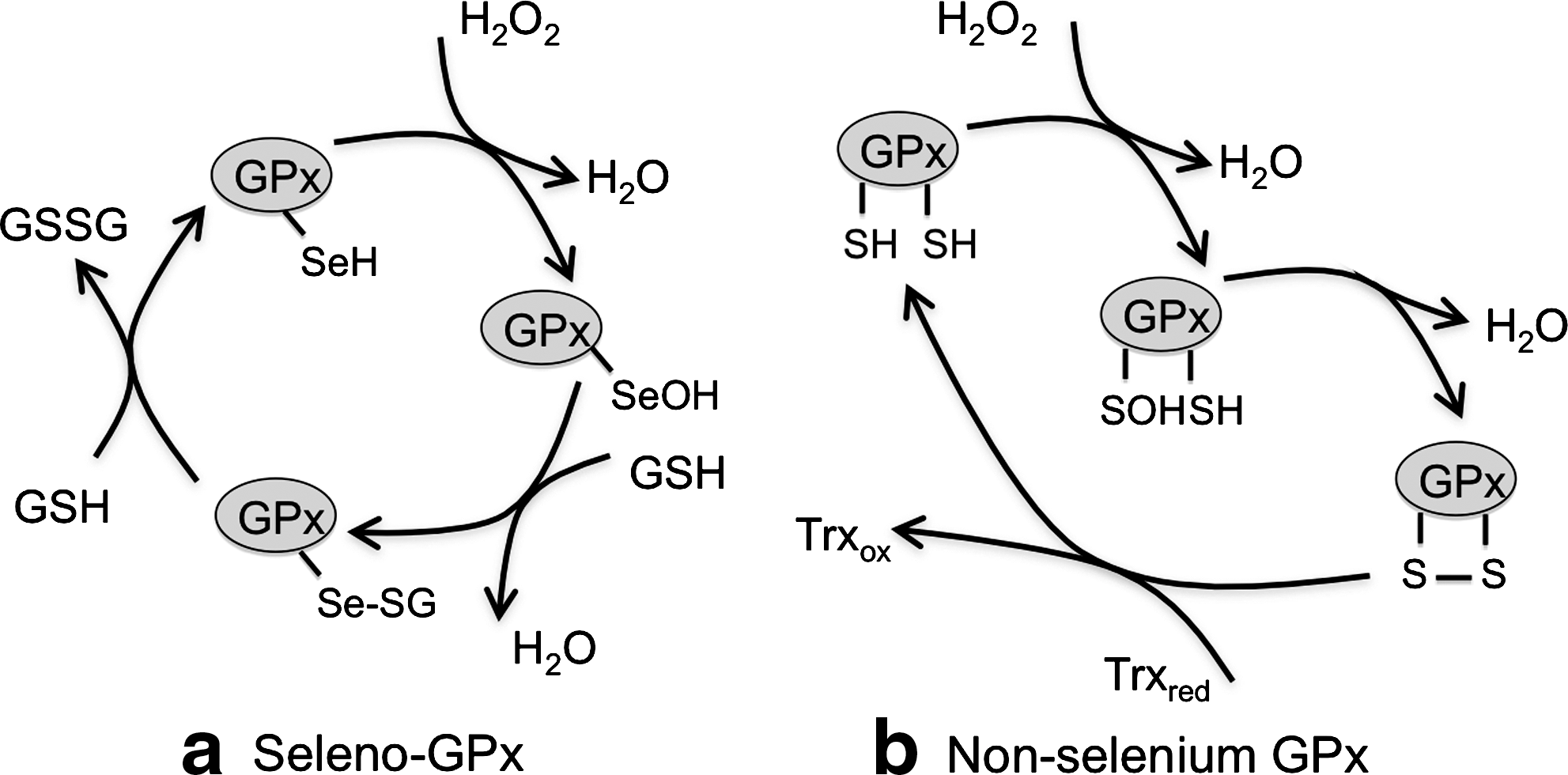
There are eight GPxs in humans. Two of them, GPx7 and GPx8 are bona fide ER resident proteins. The former is a soluble protein while the latter is anchored to the ER membrane (37). Both efficiently promote oxidative refolding of reduced cargo proteins in the presence of PDI and H2O2. GPx7 and GPx8 interact with Ero1α. GPx7 was shown to increase oxygen consumption by Ero1α. Therefore, GPx7 could promote oxidative folding, utilizing the peroxides produced by vicinal Ero1α molecules, as demonstrated for Prx4 (52, 61). Both GPx7 and GPx8 have cysteine residues in their active site. However, it remains to be established whether they share the working cycle with other nonselenium GPxs or not.
Physiology and Pathology of ER Luminal H2O2
The above findings suggest a scenario in which a dangerous by-product of Ero1 and other ER pathways, H2O2, is scavenged by Prx4, GPx7, and GPx8 which further promote oxidative folding by introducing disulfide bonds into oxidoreductases (9). However, part of the H2O2 generated in the ER could also enter anterograde transport vesicles to be secreted, or cross the ER membrane to reach the cytosol, potentially modulating various redox sensitive intra- and intercellular signaling pathways (12, 24, 31), including the activation and proliferation of certain cell types, such as B lymphocytes (6). An interesting question deserving more investigation is whether and to what extent biological membranes are permeant to H2O2 (2). Recently, evidence has accumulated for aquaporins being implicated in transmembrane H2O2 transport in yeast and mammalian cells (7, 35). The identification of molecules that facilitate or control H2O2 transport across membranes might shed a different view on redox signaling.
The unfolded protein response (UPR) is a multidimensional signaling pathway that adjusts the ER capacity to its folding demands, but becomes maladaptive and ultimately activates apoptosis if the underlying causes persist or are excessive (45). The UPR is activated by conditions that induce ER stress, such as the synthesis of aberrant proteins or of normal ones in excess, or altered Ca2+ homeostasis, and has been linked to many pathological conditions, including cancer, neurodegenerative diseases, or obesity. Therefore, a fundamental question is what determines the adaptive to maladaptive transition (22). In yeast, the UPR entails robust activation of Ero1p, and this oxidase has been implicated as an important determinant of the reduced growth of ER stressed cells (20). Similarly, worms expressing low Ero1 levels accumulated less ROS when exposed to ER stressors, and were more resistant to their toxic effects (30). These findings suggest that Ero1-derived H2O2 can sustain the maladaptive transition in ER stress responses, at least in lower eukaryotes. In mammalian cells, however, Ero1 molecules seem to have little role in sustaining oxidative stress and this may correlate with the appearance of Prx4, GPx7, and GPx8 as efficient H2O2 scavengers during evolution (3). Nevertheless, Ero1α −/− murine cells are less sensitive to ER stress-dependent apoptosis: the underlying mechanism seems to involve diminished Ca2+ release through inositol(1,4,5)triphosphate receptors around MAM (see (27) and references therein). It has been recently shown that Ero1α preferentially accumulates in MAM and regulates Ca2+ fluxes (1, 16). Interestingly, in hypoxic conditions Ero1α translocates from MAM to the ER (16).
Measuring H2O2 Production and Signaling
So far, our understanding on the role of oxidants is limited because of their rapid interconversion in the crowded cell environments and the difficulties in establishing reliable measurement methods (56). Different strategies have been followed to circumvent the difficulties in detecting H2O2 in cells and tissues (47). Chemical probes have been developed containing a boronate-based cage that releases a fluorochrome upon reacting with H2O2 (34). These probes are highly specific, but suffer two limitations: (i) they react rather slowly with H2O2 and (ii) they are not reversible, impeding dynamic measurements. On the other hand, protein-based sensors were obtained by fusing a permutated fluorescent protein with the bacterial transcription factor OxyR the activation of which is regulated by H2O2. The H2O2-dependent formation of a disulfide bond in the OxyR moiety causes conformational changes in HyPer sensors that lead to a ratiometric switch between two fluorescence excitation peaks (32).
HyPer sensors rapidly and reversibly react with H2O2 but not with O2 •−, GSSG, NO, or ONOO− (5), and allow the measurement of H2O2 production and fluxes within and between living cells. For instance, transgenic zebrafish expressing HyPer allowed to demonstrate a signaling role for extracellular H2O2 in recruiting inflammatory cells to the vicinity of wounds (38). Organelle targeted HyPer have been successfully engineered: several labs produced ER-localized HyPer (HyPer-ER) appending a signal sequence and a KDEL motif onto the HyPer N- and C-termini, respectively (14, 29, 58). By using HyPer-ER, Wu et al. provided evidence implicating the ER membrane localized Nox4 and downstream Ras/ERK signaling in the production of H2O2 in response to tunicamycin or HIV-tat treatment of endothelial cells (58).
However, a limit in the utilization of HyPer-ER is that its wavelength shifts depend on the formation of a disulfide bond. In the secretory compartment, therefore, HyPer could report on the efficiency of oxidative folding, besides on the H2O2 levels, by directly reacting with oxidized PDIs. Indeed, in reducing compartments such as the cytosol, HyPer readily reacts with thioredoxins for its disulfide recycling. It will be of interest to identify the system(s) that can oxidize/reduce the ER luminal HyPer sensors. These studies will likely be relevant to determine the role and regulation of ER peroxidases during the UPR.
Concluding Remarks
As the mechanistic dissection of Ero1 molecules proceeded, the ER emerged as a potential source of abundant H2O2 particularly in the process of de novo disulfide bond formation (54). However, it became clear that a potential foe, H2O2 can act in a friendly and useful way, further fueling oxidative folding (Fig. 8). The presence of several ER-resident peroxidases, Prx4, GPx7, and GPx8, couples the scavenging of H2O2 to oxidation of PDI, and in turn of cargo proteins (37, 52, 61). Mammalian cell and animal models tolerate well the absence (or extremely low levels) of molecules that are essential in lower eukaryotes (3). On the one hand, redundancy contrasts with the notion that specific protein-based relays—rather than a general redox poise—are active in the ER to allow the coexistence of disulfide bond formation, isomerization, and reduction. On the other hand, redundancy implies the evolution in metazoan of parallel pathways devoted to form disulfides without eliciting oxidative stress (9). Indeed, a lower sensitivity to Ero1 hypo- or hyperactivity coincides with the appearance of enzymes capable of productively scavenging peroxides. Since metabolism and redox signaling are associated with key pathophysiological events including cancer and degenerative disorders, further investigation on the handling of H2O2 in and from the ER could lead to novel approaches for therapy.

Footnotes
Acknowledgments
We thank Christian Appenzeller-Herzog, Kazutaka Araki, Milena Bertolotti, and Anna Rubartelli for their most helpful suggestions and critical reading of the manuscript; all the members of our laboratories for helpful discussions and Tina Scacciante and Tamami Ishida for secretarial assistance. This work was supported by grants from Associazione Italiana per la Ricerca sul Cancro (AIRC), Regione Lombardia and Telethon (to R.S), and by a Grant-in Aid for Creative Scientific Research and for Scientific Research on Priority Area from MEXT (to K.N). We apologize to the many colleagues whose pioneering work could not be cited owing to the strict space limits imposed by the editors.