
Abstract
Introduction
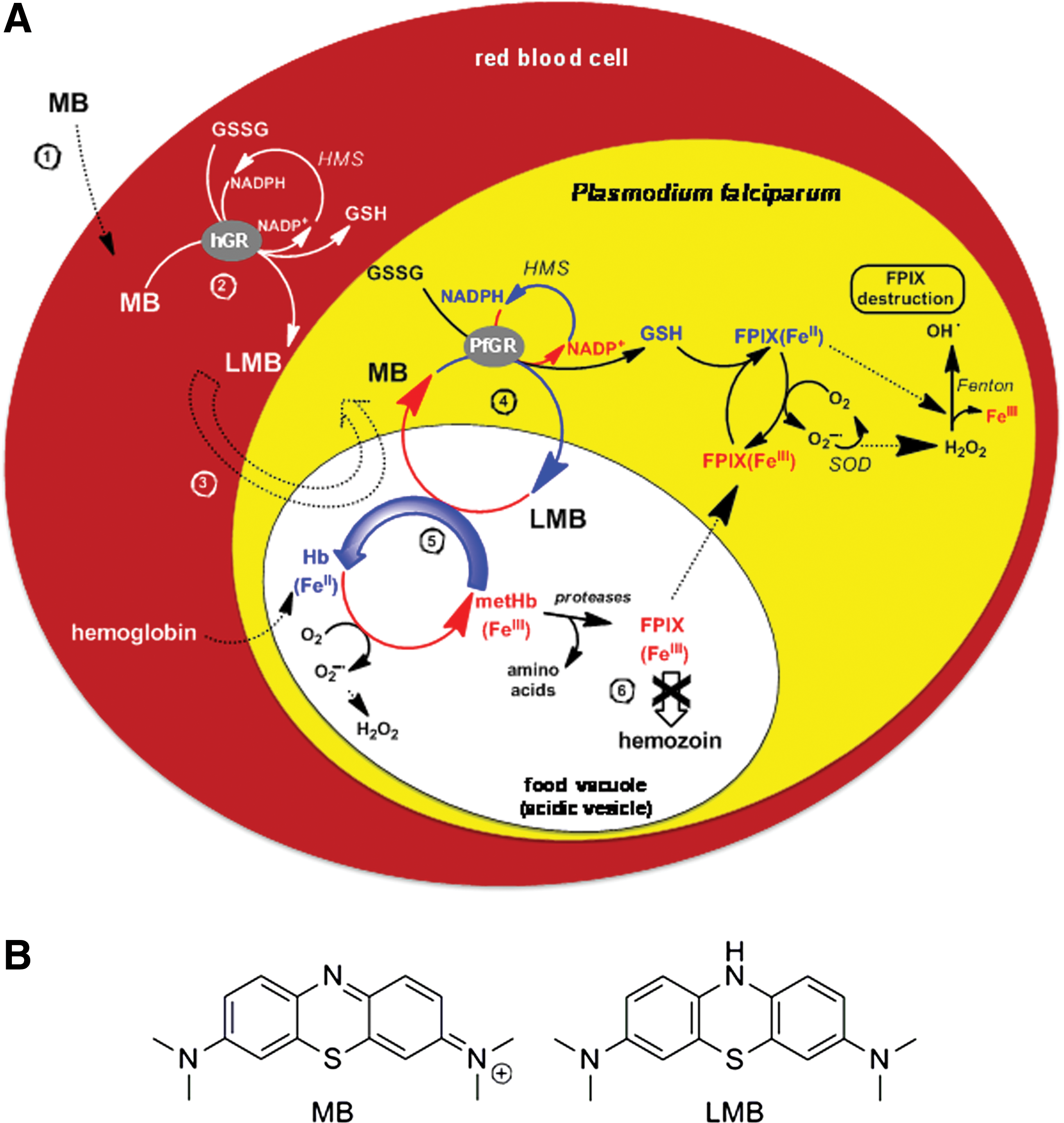
Malaria is a parasitic disease that is caused by Plasmodium species. The pathogenic agent Plasmodium infects humans and mosquitoes. The asexual period of the life cycle proceeds in different stages including those taking place in human erythrocytes. While the maturation of the parasite occurs in erythrocytes, from ring stage to trophozoite stages, acidic vesicles ensuring the transport of hemoglobin fuse into a single central digestive vacuole (also called food vacuole) where further digestion of hemoglobin proceeds at a pH around 5.5 (33). At this pH, oxyhemoglobin [oxyHb or oxyHb(FeII)] is quickly oxidized to methemoglobin [metHb or metHb(FeIII)]. The binding of hemoglobin to the hemoglobinase falcipain-2 is strictly pH dependent, and falcipain-2 preferentially binds metHb(FeIII) rather than oxyHb(FeII) (28). Inhibition of this and other proteases and its paralogues prevents parasite maturation, suggesting that drug-induced conversion of metHb(FeIII) to oxyHb(FeII) may become a strategy for the design of novel antimalarial drugs.
Innovation
The therapeutical properties of methylene blue (MB) by itself are insufficient for the rapid cure of falciparum malaria but the lack of resistance of Plasmodium falciparum to MB has renewed interest in this compound and led to drug combinations of MB with amodiaquine and artesunate. The mechanisms of interaction of MB with hematin-containing targets are yet to be fully elucidated. Here, we report that MB firmly binds to the hematin π-π dimer forming a (hematin)2:MB complex under quasi-physiological conditions. This effect is likely to contribute to the inhibition of hemozoin formation in P. falciparum. In addition, it increases the concentration of parasite-toxic heme. Furthermore, we demonstrate that the reduction of methemoglobin(FeIII) metHb(FeIII) to oxyhemoglobin(FeII) by reduced form of nicotinamide adenine dinucleotide phosphate is efficiently catalyzed by MB and glutathione reductase, the rate constant for metHb reduction being 991±44
The only part of hemoglobin not digested by Plasmodium falciparum is iron porphyrin (heme or PPIX). This heme is toxic because it can destabilize the membranes, inhibit enzymes, cause osmotic changes, and induce the Fenton reaction leading to the production of reactive oxygen species (ROS) inside the parasite. To detoxify hematin [=hydroxylated heme or FeIIIPPIX(OH) or ferriprotoporphyrin (FeIIIPPIX)] Plasmodium relies on hematin crystallization, that is the formation of hemozoin (heme biomineral or malaria pigment) (56). The stacking of iron porphyrins decreases their active surface and consequently decreases their prooxidant capacity (42). Since FeIIPPIX is an inhibitor of hematin polymerization (39), redox-active compounds displaying the ability of reducing FeIIIPPIX to FeIIPPIX can lead to the decrease of hemozoin formation and increased oxidative stress in infected red blood cells. Thus, agents with redox cycling activity able to generate FeII from FeIII states either at the hemoglobin or at the heme levels might act as putative antimalarial agents by inducing the arrest of trophozoite development. Our hypothesis is that this is the main mechanism of the antimalarial action (Fig. 1) of MB (22, 51) and of the recently reported 1,4-naphthoquinones (41).
Furthermore, hemozoin production provides benefits to the parasite in suppressing the host's immune response by inhibiting macrophage functions through impaired phagocytosis after ingestion of P. falciparum–infected erythrocytes or isolated malarial pigment (25, 53, 54). This observation suggests that any drug able to continuously redox cycle metHb(FeIII) into oxyHb(FeII), and thus to deplete the formation of hemozoin in blood stages, should contribute to reverse the depression of cellular immunity in malaria.
The redox homeostasis of P. falciparum–infected red blood cells is challenged with high concentration of ROS produced from the immune response of the host or from hemoglobin digestion in the parasite. The parasite prevents damage by a cytosolic antioxidant thiol network that is regenerated by two reduced forms of nicotinamide adenine dinucleotide phosphate (NADPH)–dependent disulfide oxidoreductases, GR (EC 1.8.1.7), thioredoxin reductase (EC 1.8.1.9) (32, 52), and possibly by lipoamide dehydrogenase (EC 1.8.1.4) (35).
The use of MB as an antimalarial therapeutic agent has been rediscovered a few years ago (51). Since then, further studies were performed to understand the molecular interactions of MB with potential targets, including metHb (27). Historically, the appearance of more efficient antimalarial drugs and of insecticides to combat the mosquito vector of the disease halted the use of MB. The resistance of P. falciparum to drugs, and the resistance of Anopheles to insecticides have led to the revival of MB use. No clinical resistance to MB in P. falciparum has been reported so far, nor could it be induced in drug pressure experiments carried out with parasites in cultures and in animal studies (58). Furthermore, MB was found active, not only against the disease-causing asexual schizonts, but also against the disease-transmitting gametocytes of P. falciparum: this antimalarial activity was observed both in vitro with all blood stages of various parasite strains in culture with 50% inhibitory concentration (IC50) values in the lower nanomolar range (2) and in vivo (13). However, the therapeutical activity of MB alone is not sufficient to rapidly cure falciparum malaria in humans whereas a drug combination of MB with amodiaquine and artesunate was found to be very effective (62) and included a pronounced gametocytocidal activity. These effects recommend MB-based combinations for the control of transmission of P. falciparum malaria in endemic regions (13, 30). Furthermore, as early as 1891, it was reported that the phenothiazine dye MB also killed Plasmodium vivax (26).
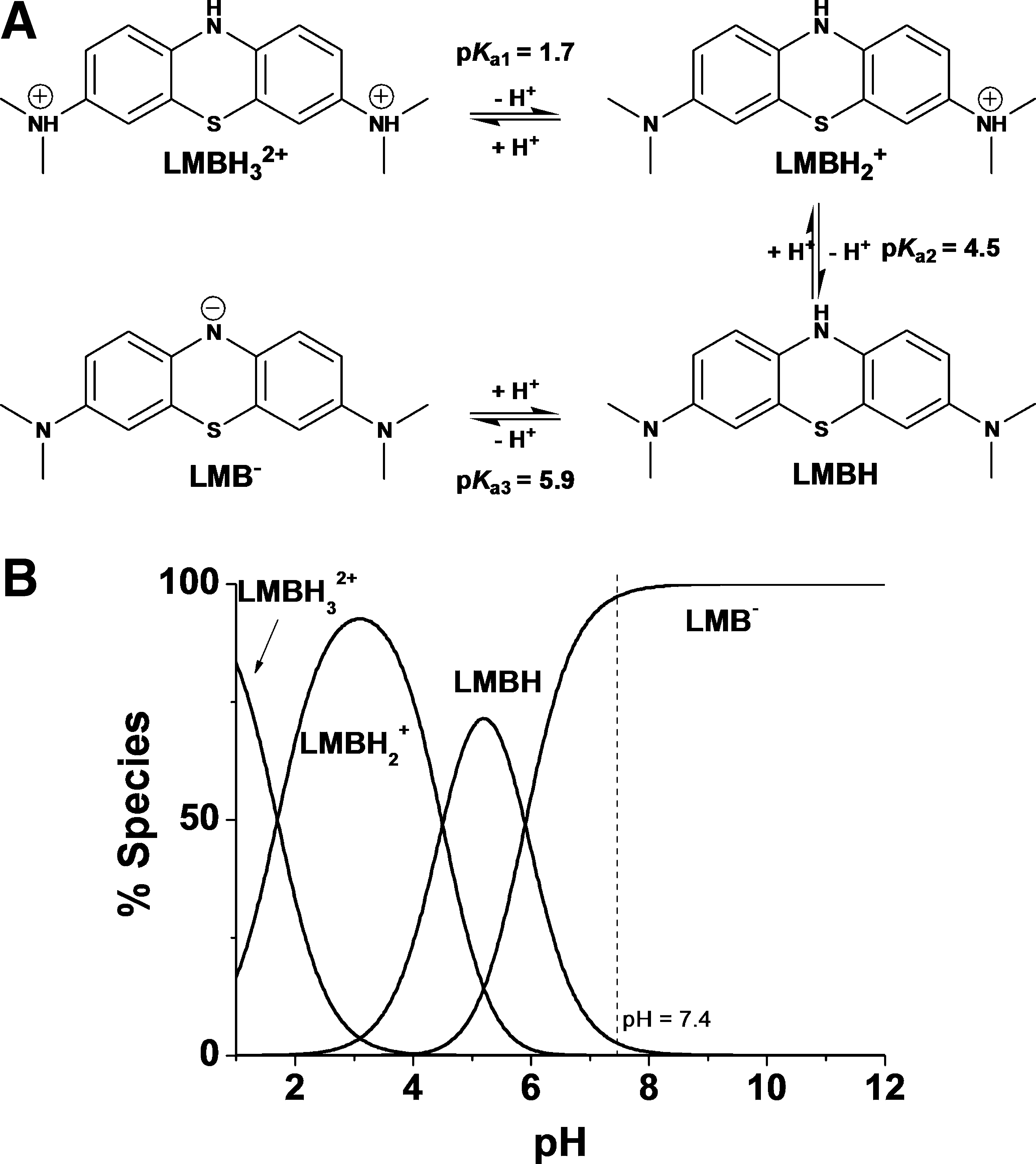
The fact that MB is excreted as unmodified form, as its two electron–reduced form leucomethylene blue (LMB, Fig. 1), or as one of its demethylated metabolites (51) points out that these compounds per se are responsible for the drug activities of MB in vivo. In addition, structure-activity relationships are of particular interest since the structural properties of the heterocycle are multifaceted: MB is involved in intricate redox processes; it has the ability to change its structure pronouncedly upon reduction to LMB and both structures can be protonated in different ways. Additionally, MB and its two electron–reduced form LMB (Fig. 1) possess different lipophilicities, and MB tends to dimerize in water. MB is reduced by various NADPH-dependent disulfide reductases (9, 10), including both glutathione reductases from Plasmodium-infected human red blood cells (22). The flow of electrons from the nicotinamide ring of NADPH proceeds via the flavin ring of GR-bound FAD to the phenothiazine of MB/leucoMB. Many flavin-binding disulfide reductases show this diaphorase activity catalyzing the reduction by NAD(P)H of various dyes (MB) and quinones that act as hydrogen acceptors. Based on cysteine alkylation or Cys→Ala mutations at the active site dithiol, the NADPH-reduced flavin was shown to be the redox center for this diaphorase activity (6, 10). In particular, the deficiency of metHb(FeIII) reductase (NADH diaphorase) activity in human erythrocytes responsible for hereditary or acquired methemoglobinemia (57)—characterized by an elevated level of metHb(FeIII) in erythrocytes—is corrected by MB administration (51). Redox cycling and the ability of MB to modify its lipophilicity upon electron exchange are therefore likely to form the basis for its biological effects. Revisiting the physicochemical properties and the mechanisms of action/interactions of MB toward its relevant targets might contribute to a better understanding of the physiopathology of the disease and to corroborate the concept of new redox-active agents (16, 41) against malaria.
Results
Structural and redox properties of MB
MB is based on a tricyclic phenothiazine chromophore and is an intensely colored blue cationic dye (ɛ660=105
Protolytic properties of MB and LMB
MB is a positively charged cation in a wide pH range (pK a=0) (29). Under basic conditions (pH>10–11), MB is not stable and can undergo stepwise and slow demethylation (55). By contrast, LMB could be represented by LMBH3 2+, LMBH2 +, LMBH, and LMB− protonated species (Fig. 2). Above pH 7, LMB exists in water mainly as a deprotonated anion, LMB− (29).
Dimerization of MB
The dimerization of MB in aqueous solution may appear surprising, since it possesses a positive charge, but many dyes and organic molecules display this phenomenon (38). These interactions occur in aqueous solution and have both a π-π stacking and a charge-transfer (CT) complex character (40). The nature and the concentration of electrolytes, temperature, and pH have also significant influence on the dimerization processes (45). The dimerization mechanism of MB is a two-step process involving the very fast formation of a complex from the diffusion controlled interaction of two MB molecules (k
1f=1.76×109
To evaluate the K
Dim value under physiological conditions (pH=7.4, T=37°C), ultraviolet/visible (UV-Vis) spectra of MB were recorded at different concentrations (Fig. 3). Within the concentration range of 2.73 μM to 175 μM, a single equilibrium 2MB ⇄ (MB)2 has been considered and higher oligomerization processes were neglected (see Supplementary Data available online at

The absorbances at any wavelength can then be correlated to the absorptivities of both the monomer and the dimer, which are related by K
Dim (24).
The use of equation 4 resulted in the determination of K
Dim=6.8×103
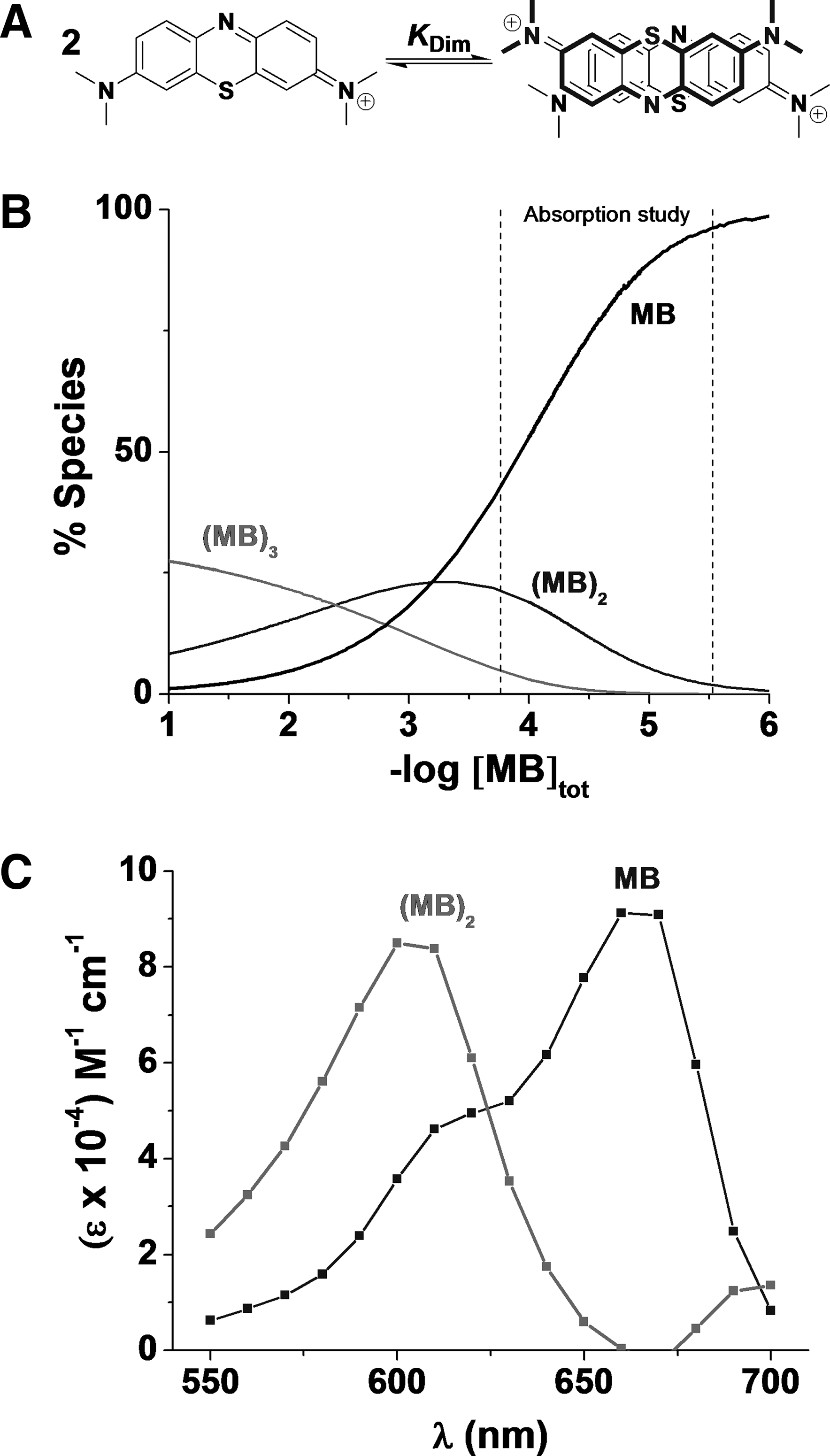
The distribution diagrams (Fig. 4) demonstrate the predominance of MB and (MB)2. This range of concentration from 2.73 to 175 μM is relevant since MB accumulated inside the erythrocytes (36, 43). The formation of (MB)3 or oligomers of higher degree is unlikely to occur in vitro and in vivo since the concentrations required in water for such processes is >1 mM. Polymerization reactions, however, can occur in solvents of low dielectric constants such as hexane. In addition, no pH dependence on the dimerization of MB is expected because of the very low pK a value of MB (pK a=0) (29) unless at high pH where demethylation may occur.
The dimerization was taken into account in further experiments with MB in aqueous solution, and was considered for further models for the interpretation of UV spectra. In contrast to the dimer formation in water, no dimerization was observed in dimethylsulfoxide (DMSO).
FeIIIPPIX in aqueous solution
In water, ferriporphyrins such as hematin can undergo various equilibria (dimerization and oligomerization) and protolytic processes depending on the physicochemical conditions. The nature of the hematin dimer in solution has been elucidated and reviewed in the past few years (17, 31, 44). The μ-oxo dimer is the dominant species in aqueous mixtures of aprotic solvents or in detergent solutions (Fig. 5). It is characterized by an absorption band in the visible region centered at ∼575 nm (4).
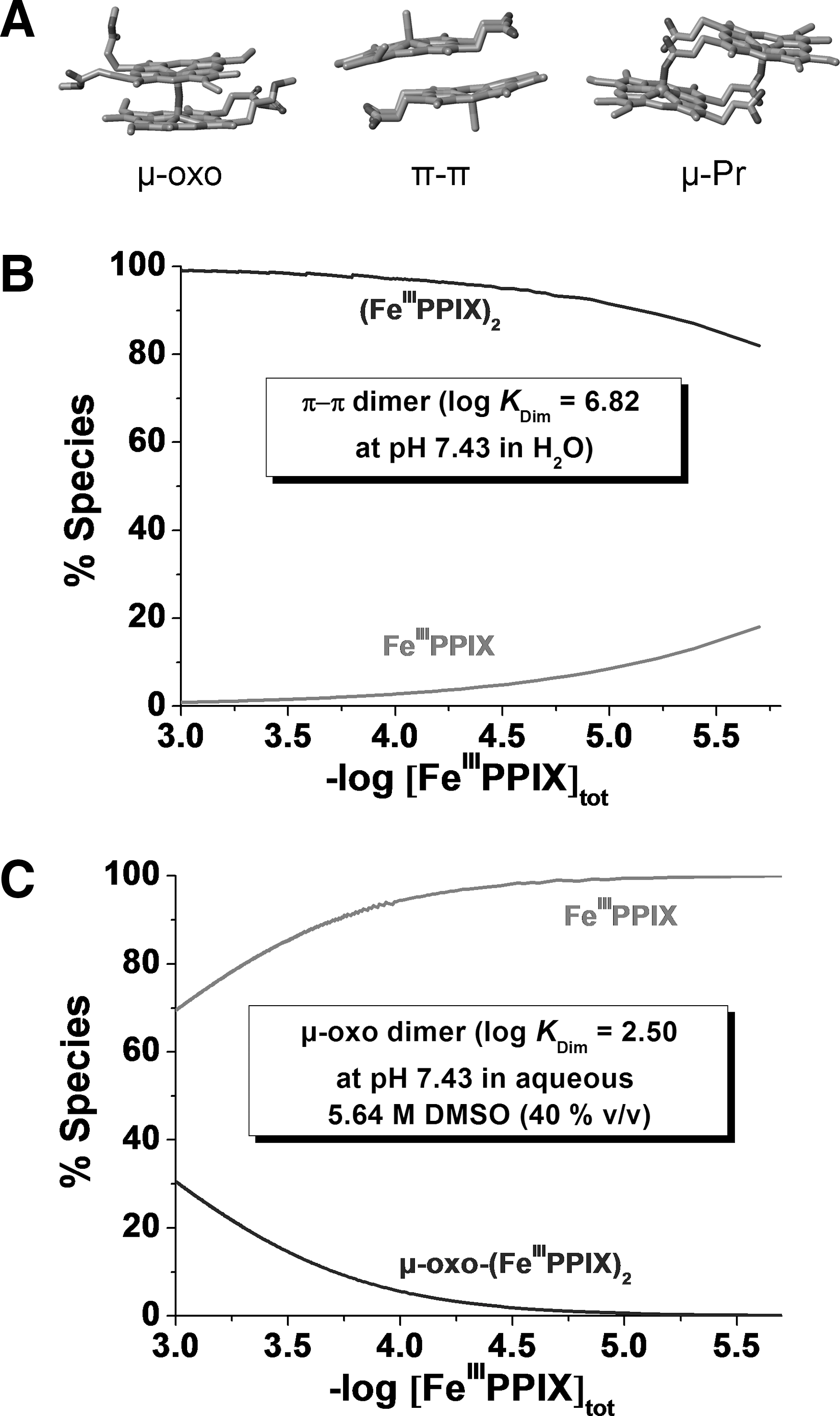
At pH 7.4, the π-π (FeIIIPPIX)2 dimer represents the major species in a wide range of [FeIIIPPIX]tot (Fig. 5). The (FeIIIPPIX)2 dimer is characterized by broad Q bands in the visible region and a CT band, (band III, a2u→d yz ) (19, 61), at ∼630 nm (4). The direct overlap of the π orbitals of the porphyrins results in an excitonic coupling, which breaks the degeneracy of the excited state B, giving rise to two Soret bands lying at 385 and 360 nm. These broad and split Soret bands are spectroscopic signatures of the presence of the π-π dimer (4).
Potentiometric and absorption spectrophotometry were used to analyze the protonated species of FeIIIPPIX present under physiological conditions (see Supplementary Data). Figure 6 displays the potentiometric titration of a 1.83 mM (FeIIIPPIX)2 solution. Under these conditions, the (FeIIIPPIX)2 dimer is the exclusive heme species in solution. Below pH 3, the dimer is soluble in water (dark brown color). Above pH 3 and up to pH 5–6, a brown precipitate, attributed to β-hematin (crystalline heme), is formed spontaneously. This precipitate could be resolubilized above pH 6.0. A color change from brown to green was observed when the pH was further increased to ∼11.5–12, thus indicating deprotonation of the diaquo(FeIIIPPIX)2 species. The statistical processing of the data allowed determining two pK a values (pK a1=7.0 and pK a2=8.06). These data are in reasonable good agreement with the literature values (pK a1=6.2 and pK a2=8.5) (17).
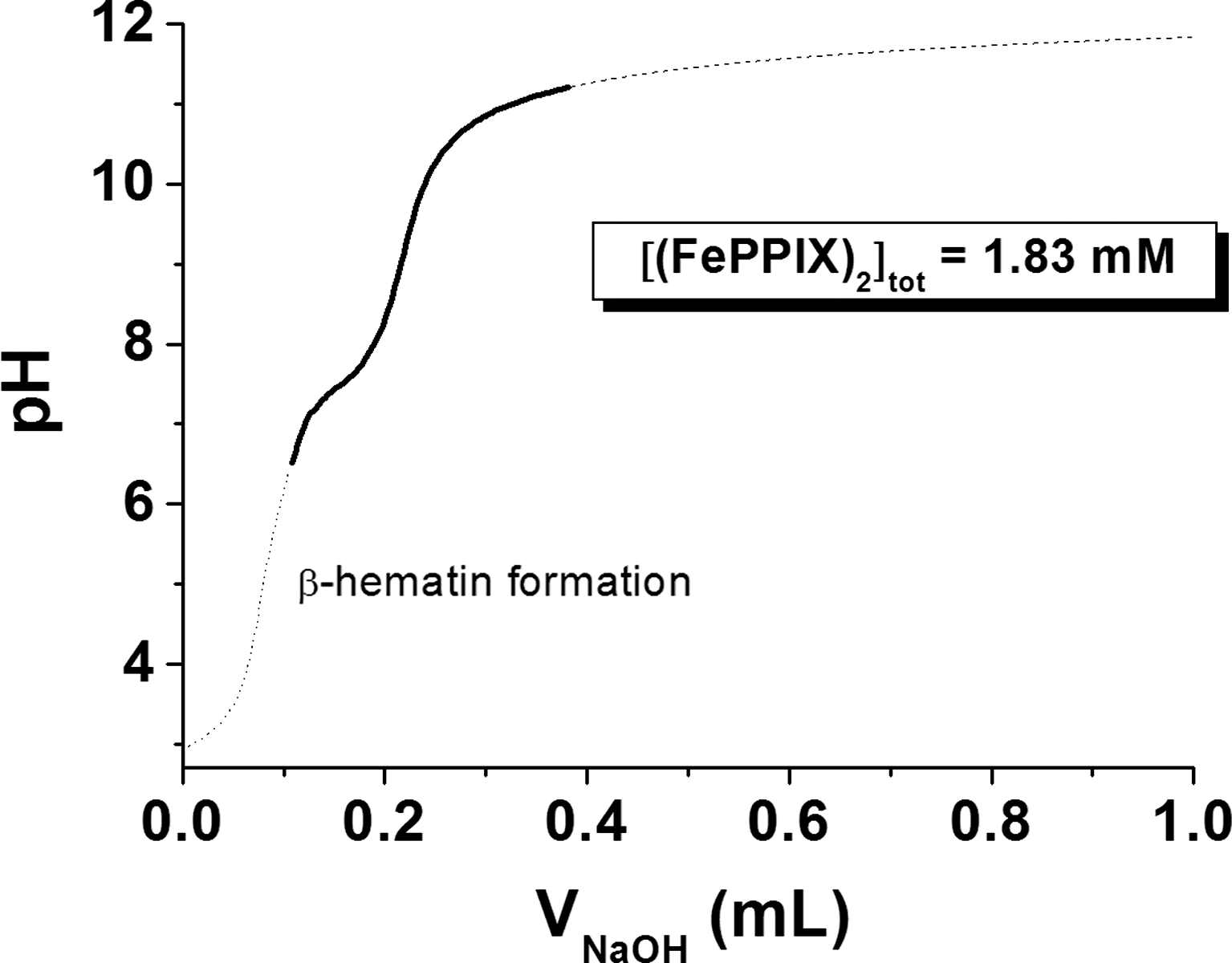
Under basic conditions, the absorption spectrum of the (FeIIIPPIX)2 dimer is characterized by a split B band centered at 385 and 360 nm and a CT absorption at ∼611 nm. Upon protonation, marked spectral changes were observed with a red shift of ∼19 nm of the CT band and a pronounced change of the relative intensities of the two Soret absorptions at ∼360–385 nm. The statistical processing of these data allowed characterizing two protonated species, (FeIIIPPIX(OH))2 and (FeIIIPPIX(OH2))2, which are related by a global protonation constant log β2=14.34. This value is in agreement with the β2 (equation 7) that can be deduced from potentiometry (log β2=pK
a1+pK
a2=14.7, equations 5 and 6).
Electronic spectra are given in Figure 7. Deprotonation of the two water ligands induces a bathochromic shift of the CT absorption and a hyperchromic shift of the Soret bands additionally to the change in the relative absorptivities. These spectral observations can be rationalized by stronger binding of these exogenous ligands that pull outward the two ferric centers and thereby alter the π-π interactions as well as the overlap between the porphyrin, e.g., π* excited state orbitals and the FeIII d yz and d xz orbitals.
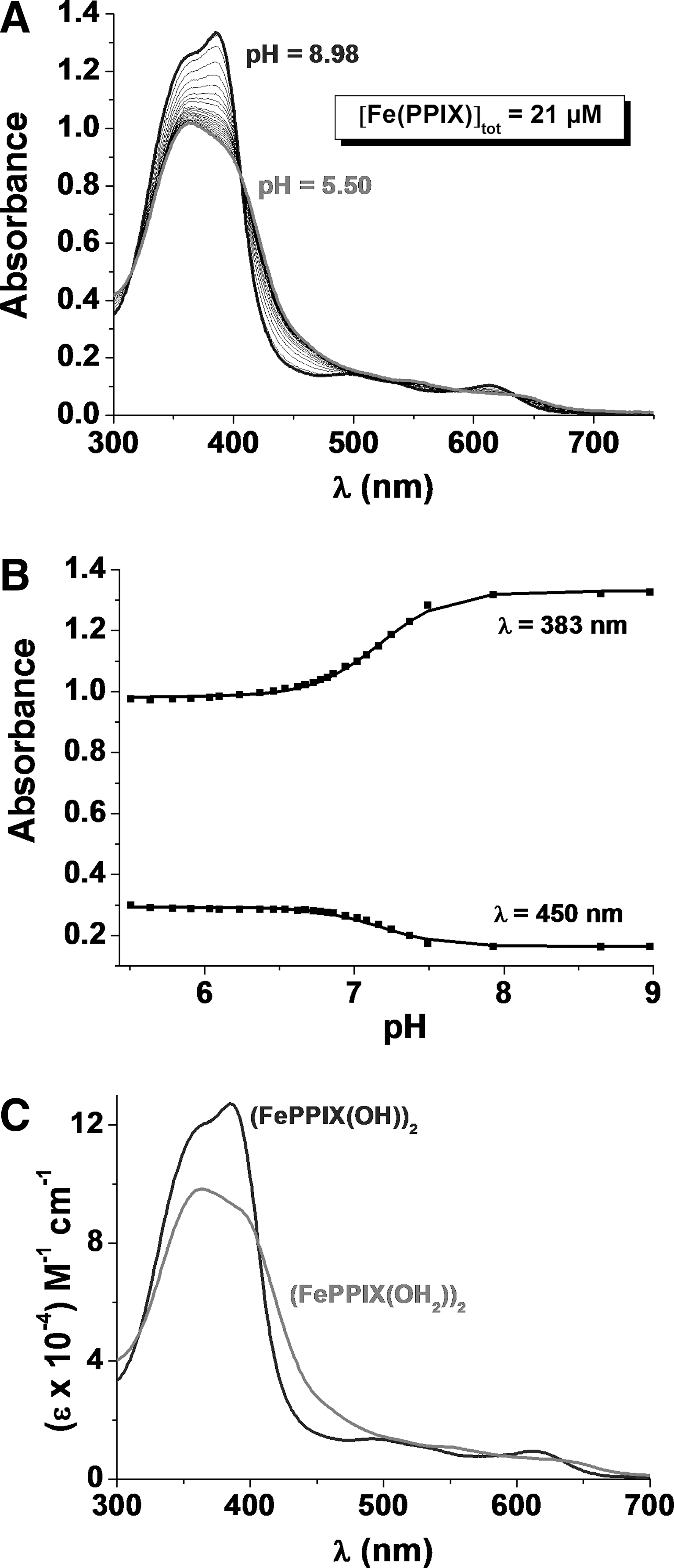
At pH 7.4, the mixed aquo/hydroxo complex (FeIIIPPIX(OH2))·(FeIIIPPIX(OH)) dominates, while the bisaquo (FeIIIPPIX(OH2))2 species is the major species below pH 6 (4), that is under conditions where β-hematin precipitates spontaneously. Our potentiometric study shows that the polymerization and crystallization processes may therefore be related to the formation of (FeIIIPPIX(OH2))2. The water molecules are more labile ligands than the hydroxyl groups. At pH<6, the propionic side arms are deprotonated and are therefore capable to substitute the labile coordinated water ligands and to trigger the biomineralization process. The pH of the acidic digestive vacuole of P. falciparum is well suited for hemozoin nucleation and growth, which contributes to detoxification of the large concentration of heme produced by digestion of metHb. Under more acidic conditions, dissociation can be related to the protonation of the propionates (14).
Hematin:MB speciation in aqueous solution
The inhibition of hemozoin formation is one of the targets of antimalarial therapy. The interactions between (FeIIIPPIX)2 and MB at pH ∼7.4 were first analyzed by UV-Vis absorption spectroscopy. The stoichiometry of the MB:hematin complex was determined with the method of continuous variations. Series of HEPES-buffered solutions of MB and FeIIIPPIX subject to the condition that the sum of the total MB and FeIIIPPIX concentrations is constant (84.1 μM) were prepared. Absorption spectra were measured for each of the solutions and the extremum x max value (∼0.31) indicates the predominant formation of a 2:1 stoichiometry complex ((FeIIIPPIX)2:MB).
To evaluate the affinity of MB for the hematin dimer and to get further structural and spectroscopic information, spectrophotometric titrations in the UV-Vis region were then performed at pH 7.4 (Fig. 8).
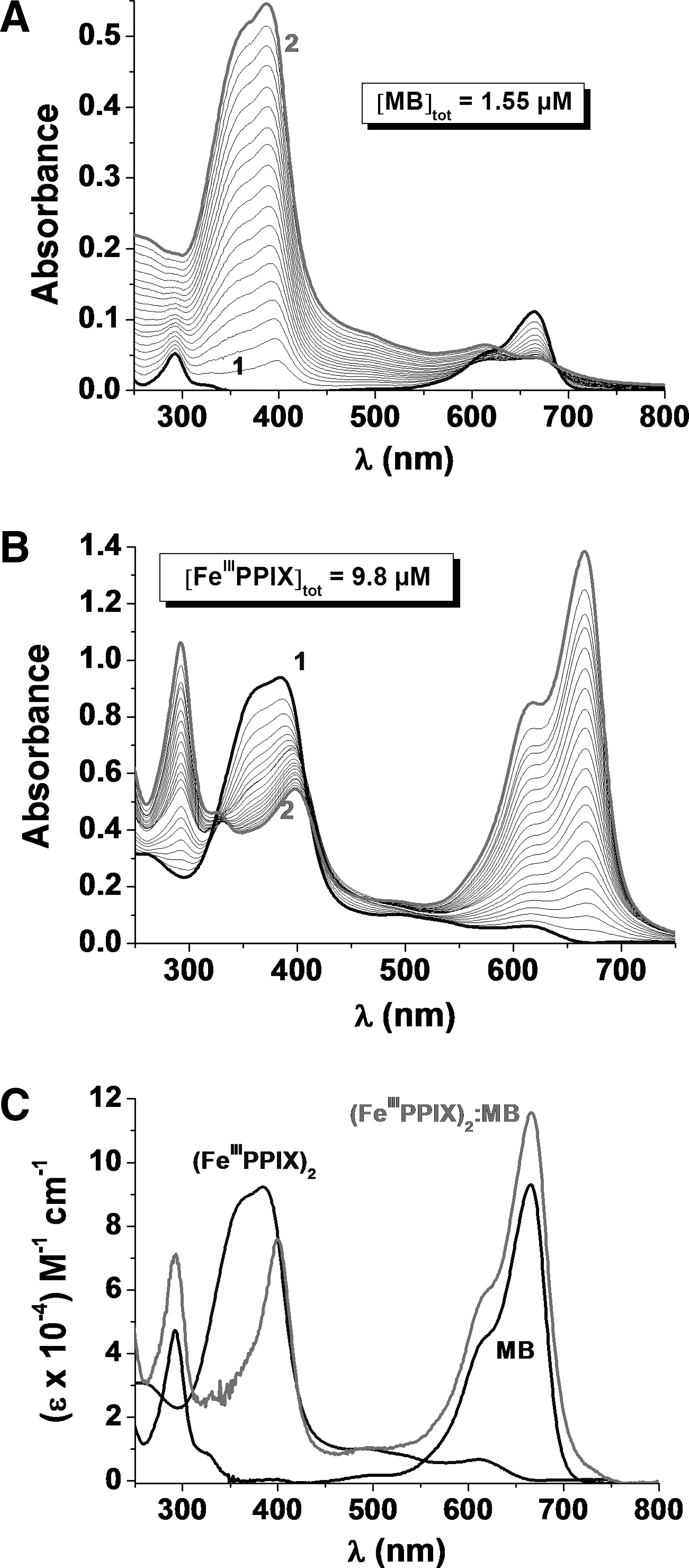
The statistical processing of the spectrophotometric data, taking into account K
Dim=6.8×103
Binding of MB induces a significant narrowing and red shift of the Soret band of hematin. This spectroscopic finding suggests strong interactions between MB and FeIIIPPIX and demonstrates a loss of excitonic coupling between the two ferriporphyrins, which indicates that MB is intercalated between two FeIIIPPIX in a sandwich-type arrangement.
Since MB is a fluorescent compound (S2→S0 radiative deactivation at λ=685 nm), the measurement of fluorescence spectra of the MB–hematin titration was the method of choice for an additional proof of the complex formation. The formation of (FeIIIPPIX)2:MB was characterized by an inhibition of the MB-centered emission in agreement with π-π stacking and/or a CT between the two types of chromophores in the sandwich-like complex. The global association constant was determined to be logβ((FeIIIPPIX)2:MB))=13.1 and is in good agreement with that determined from absorption.
Methemoglobin reduction coupled assay with hGR/NADPH
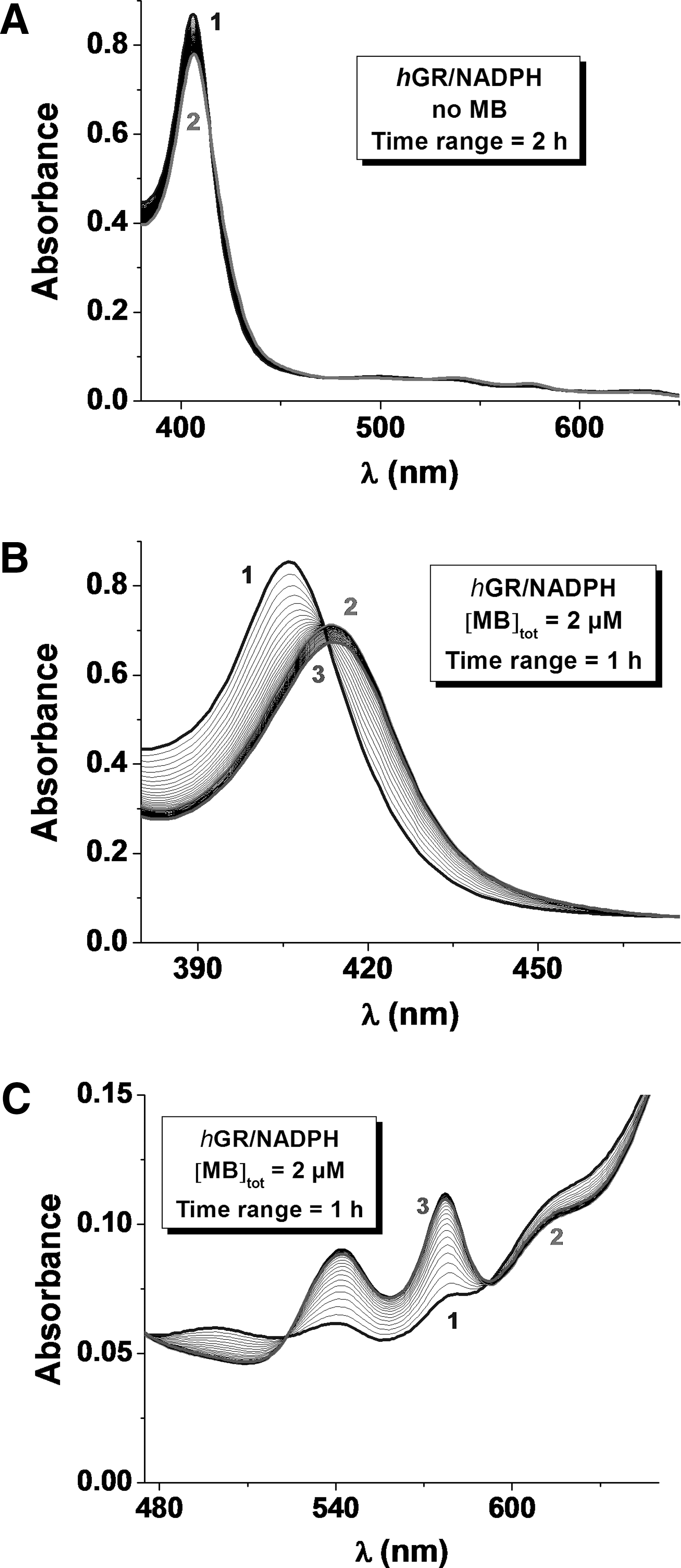
Methemoglobin reduction by MB was reported by clinicians many years ago; its reduced metabolite LMB is the mediator in vivo (7). A reduction assay coupled to the human GR (hGR)/NADPH system in vitro was recently established as a relevant in vivo model in our laboratory (16). The reduction process of metHb(FeIII) can be observed due to characteristic changes in the UV-Vis region (see Supplementary Data). The absorption spectrum of metHb(FeIII) is characterized by a maximum absorption at ∼405 nm (Soret band of the pentacoordinated FeIII heme in a high spin state) and a broad band centered at ∼603 nm. Upon metHb(FeIII) reduction by LMB (T=25°C; pH=6.9), which is generated from MB by hGR, the formation of the reduced Hb(FeII) species was associated with a bathochromic shift of the λmax of the Soret band from 405 to 410 nm (Fig. 9B). This corresponds to oxyHb(FeII) with the metal cation being hexacoordinated in a low spin state. We also observed the appearance of two new absorption bands at ∼536 and 576 nm (Fig. 9C).
Control experiments clearly showed that in the absence of the mediation by MB and its reduced form LMB, the hGR/NADPH couple is not able to reduce metHb(FeIII) directly (Fig. 9A). The redox cycling of MB is based on NADPH oxidation by hGR, which in turn catalyzes the reduction of MB to LMB (k
NADPH/hGR
red=4760 M−1·s−1) (8). NADPH per se does not efficiently reduce MB to LMB (k
NADPH
red=6.6 M−1·s−1 at pH 6.9) (10) without the enzyme hGR and no reduction of metHb(FeIII) is observed in the absence of MB (LMB). This indicates that the hGR/NADPH couple is not the prevailing system but only the reductant for MB (Fig. 9). Thus, LMB is the active reducing intermediate for the reduction of metHb(FeIII) to oxyHb(FeII). The splitting of the electron pair could occur at the GR-bound flavin; hence the electrons are transferred to the phenothiazine of MB and then from LMB to the heme group of metHb. Since the amount of MB used in the experiments was in some cases in the substoichiometric range {2.52<[MB]tot/[metHb(FeIII)]tot
mon<0.12}, and the process of metHb reduction ran to completion, the data clearly demonstrate that the redox cycling of metHb(FeIII) to oxyHb(FeII) is catalytically mediated by MB through LMB generated in hGR-catalyzed NADPH reactions (Fig. 9). We studied metHb(FeIII) reduction by varying [MB]tot at pH 6.9 in the presence of fixed [hGR]tot, [NADPH]tot, and [metHb(FeIII)]tot. The apparent reduction rate (k
obs, s−1) varied linearly with [MB]tot and the bimolecular reaction rate of the LMB-mediated reduction of metHb(FeIII) was calculated to be k
metHb
red=991±44
Discussion
The determination of the physico-(bio)chemical properties of MB in vitro was focused on its features as an antimalarial drug. Our results led to the conclusion that MB has various profiles of action: (i) MB firmly interacts with hematin though a 2:1 sandwich complex formation and thus competes in terms of binding affinity for hematin dimers with the most potent drugs of the 4-aminoquinoline series (20, 21); (ii) MB undergoes a catalytic redox cycling that leads to efficient reduction of metHb to oxyHb through MB reduction in NADPH-dependent diaphorase-catalyzed reactions mediated by hGR (Figs. 1 and 9). Typical diaphorases are moonlighting flavoenzymes such as dihydrolipoamide dehydrogenase and glutathione reductase. These interactions are likely to enhance the oxidative stress for the parasite through inhibition of hemozoin formation and elevation of the hematin(FeIII) concentration in the acidic digestive vesicles and vacuole.
While the MB-induced reduction of metHb(FeIII) is harmful to the parasites, it is potentially beneficial for the host. The amount of metHb(FeIII) is abnormally elevated in severe malaria, forming up to 16.4% of circulating hemoglobin, and also in nonparasitized human red blood cells during P. vivax infections (3). As reported by Anstey et al. (3), the degree of methemoglobinemia correlates with disease severity and severity of anemia. A decreased oxygen-carrying capacity of blood due to anemia was observed to be exacerbated by reduced oxygen-carrying capacity of metHb(FeIII) (even in the presence of a low amount) contributing to tissue hypoxia in patients with severe falciparum malaria. Falciparum malaria is a major cause of morbidity and mortality in African children with severe anemia who live in malaria endemic areas. The high frequency of the sickle-cell hemoglobinopathy and of glucose-6-phosphate dehydrogenase (G6PD) enzyme deficiency in malaria endemic regions is believed to be due to a natural genetic protection against fatal malaria (1, 12, 34, 50). Evidence of G6PD deficiency in Mediterranean countries, Asia, and Sub-Sahara Africa conferring resistance against severe malaria can be explained by biochemical and clinical observations. At the biochemical level, G6PDH is the first enzyme of the pentose phosphate pathway responsible for providing the reducing power of the cells in the form of NADPH. As NADPH is the main reductant of GR in erythrocytes, G6PD deficiency in red blood cells can be regarded as hGR insufficiency (23). The deficiency is not lethal for humans but prevents severe attack of malaria since the oxidative stress released in erythrocytes creates a milieu hostile for Plasmodium. Inherited hGR deficiency has similar effects (23). Consequently, in these enzyme deficiency diseases and most of hemoglobinopathies, a rapid elimination of the red blood cells from the circulation occurs by enhanced phagocytosis via complement activation (5, 11, 23). Increased oxidative stress in red blood cells induced by redox-active compounds like MB (23) or 1,4-naphthoquinones (41) is also expected to protect from malaria by triggering enhanced ring-stage phagocytosis rather than by impairing parasite growth directly. While MB at high dose was reported to induce hemolysis, it is important to mention that a recent clinical study in Burkina-Faso showed both efficacy and safety of the redox-active MB in G6PD-deficient patients with uncomplicated falciparum malaria (62).
Since the malaria treatment is faced with resistance of the parasites to various drugs, it is important to identify new targets and to reconsider old orphan drugs. MB appears to be an excellent candidate to be applied in drug combinations and to serve as the basis for the design of new redox-active substrates as novel antimalarial agents. Optimization and development of new antimalarial lead redox-active drug candidates into preclinical studies will however involve several challenges concerning the safety of patients with hemoglobinopathies and malaria.
Footnotes
Acknowledgments
The Centre National de la Recherche Scientifique (CNRS) and the University of Strasbourg (UMR 7509 CNRS-UdS) partly supported this work. This work was also funded by the ic-FRC (International Center for Frontier Research in Chemistry) in Strasbourg. O. B. thanks the French Embassy/Affaires Etrangères/in Berlin for granting her a postdoctoral fellowship. The authors also express their gratitude to Dr. Didier Belorgey for his careful reading of the manuscript.
Author Disclosure Statement
No competing financial interests exist for any of the authors.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.