
Abstract
Introduction
BolA homolog 2 has been shown to be cytoplasmic in S. cerevisiae and to interact with the cytoplasmic monothiol glutaredoxins Grx3 and Grx4 (23). In S. cerevisiae, it has been named Fra2 and, together with Grx3/4 and another protein (Fra1), is involved in regulation of the iron regulon via the nucleoplasmatic shuttling of the transcription factor Aft1 (23). BOLA3 has recently been implicated in the biogenesis of [Fe-S] clusters for oxidative phosphorylation complexes and 2-oxo acid dehydrogenase enzymes in mitochondria (6).
Innovation
The BolA protein family is widespread among eukaryotes and bacteria. There are reports about its role in defense against oxidative stress in bacteria, but its function in mammals is unknown. Furthermore, the mechanism behind its role in defense against oxidative stress has not been elucidated, although it has been predicted to interact with a monothiol glutaredoxin. Here we show that BOLA1, one of the three members of the family in eukaryotes, is mitochondrial in human. Knockdown of the gene increases the level of oxidation of mitochondrial thiol groups. Overexpression of the gene reduces the effect of glutathione (GSH) depletion on the oxidation of thiol groups, and nullifies the effect of GSH depletion on mitochondrial morphology. Consistent with its role in reducing thiol redox potential, we show that BOLA1 orthologs only occur in aerobic species. We supply evidence that BOLA1 interacts with a mitochondrial monothiol glutaredoxin GLRX5, providing a possible mechanism for BOLA1's role in cellular processes, such as mitochondrial morphology regulation.
Little experimental data are available on the function of Bola homolog 1. BOLA1 has been observed to be overexpressed in mononuclear cells of female patients with chronic fatigue syndrome (12), but that observation could not be reproduced in a systematic study (4). Here we set out to examine the function of BOLA1, with specific emphasis on its role in regulating mitochondrial morphology.
Results
BOLA1 orthologs only occur in aerobic species
In eukaryotes, the BolA protein family comprises three phylogenetically widespread clades (Fig. 1). Notably, none of the genomes of anaerobic eukaryotes sequenced contains a BOLA1 ortholog, whereas orthologs of BOLA2 and BOLA3 are present in the anaerobic genus Cryptosporidium and the anaerobic species Blastocystis hominis, but not in the other anaerobic species (Trichomonas vaginalis, Giardia intestinalis, Encephalitozoon cuniculi, and Entamoeba histolytica). As the last common ancestor of all eukaryotes likely was aerobic (10) and the eukaryotic anaerobes with sequenced genomes belong to five independent evolutionary lineages (T. vaginalis and G. intestinalis belong to the same lineage), BOLA1 orthologs have been lost at least five times in the transition to anaerobic life, and BOLA1's genomic presence strongly correlates with an aerobic metabolism.

BOLA1 specifically localizes in mitochondria
Mouse BolA1 contains a mitochondrial targeting signal [0.97, mitoprot (8)], and its mitochondrial localization was verified by GFP tagging and mass spectrometry (29). The mouse BolA1 protein was found in 13 out of 14 tissues examined (29) and human BOLA1 is ubiquitously expressed, but not overexpressed (at least 10-fold over the median) in any specific tissue (33). Likewise, human BOLA2 and BOLA3 have been found to be ubiquitously expressed, but not overexpressed in any specific tissue examined (33), while mouse Bola3 protein has been detected in mitochondria in 12 out of 14 tissues examined (29). A c-Myc tagged version of BOLA1, overexpressed in African green monkey kidney (Cos-7) cells, was shown to be present in discrete spots that did not encompass the Golgi, but were not further characterized (43). To determine BOLA1's intracellular localization, we transduced primary human skin fibroblasts with a baculovirus containing a fusion of BOLA1 and AcGFP1 (BOLA1-GFP) and loaded them with the mitochondrial membrane potential-sensitive fluorescent dye tetramethylrhodamine methyl ester (TMRM).
Combined digital-imaging microscopy of TMRM and GFP fluorescence revealed that 70% of the fibroblasts expressed BOLA1-GFP. The clear overlap of its fluorescence with that of TMRM demonstrated BOLA1-GFP's mitochondrial localization in human cells (Fig. 2A). BOLA1-GFP overexpression did not detectably alter the mitochondrial appearance (Supplementary Fig. S1A, fifth panel; Supplementary Data are available online at
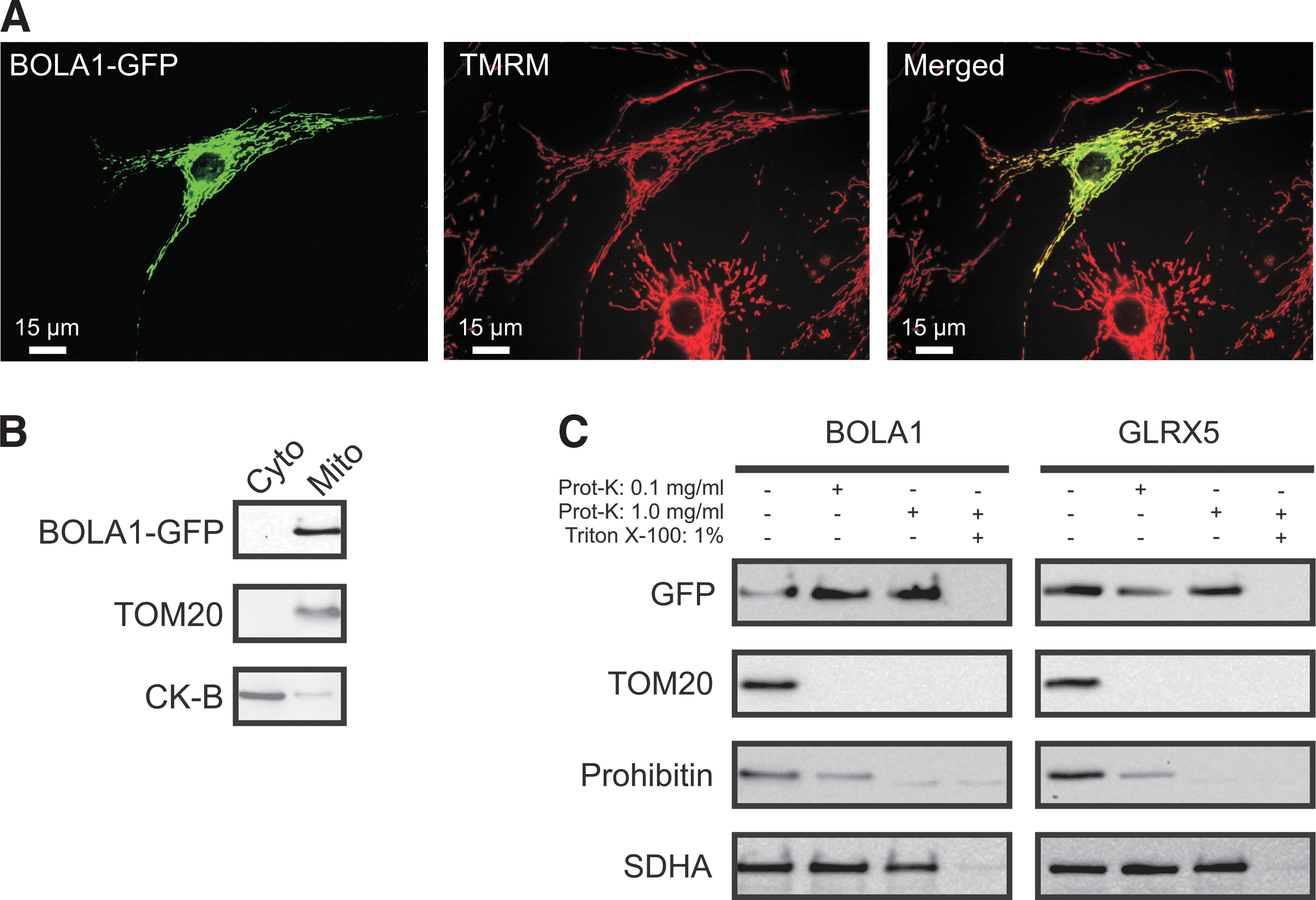
Fractionation of HEK293 cells induced to express BOLA1-GFP confirmed its mitochondrial localization (Fig. 2B). In mitochondria with digitonin-permeabilized outer membranes, both BOLA1-GFP (Fig. 2C, left panel) and GLRX5-GFP (Fig. 2C, right panel), like subunit A of mitochondrial matrix succinate dehydrogenase (SDHA), appeared resistant to proteinase K, whereas this enzyme readily digested the import receptor (TOM20) of the outer-membrane translocator and prohibitin of the inner membrane, indicating that BOLA1 is located within the limits of the inner-membrane.
BOLA1 is complexed with GLRX5
The BolA-like protein family displays a phylogenetic distribution that is very similar to that of the monothiol glutaredoxins (9, 16), and yeast BolA2 was indeed demonstrated to interact with cytosolic monothiol glutaredoxins Grx3 and Grx4 (23). As human mitochondria contain the monothiol glutaredoxin GLRX5 (30) (Fig. 2C, right panel), we expected BOLA1 to interact with it, although the phylogenetic co-occurrence of BolA1 and Grx5 is not perfect (Supplementary Fig. S3). Supporting an interaction between BOLA1 and GLRX5 is that the S. cerevisiae BolA2 His-103 is conserved in the BolA1 orthologous group (Fig. 1 and Supplementary Fig. S4). This histidine binds the [Fe2S2] cluster that has been implicated in the BolA2-Grx3/4 complex in S. cerevisiae (25).
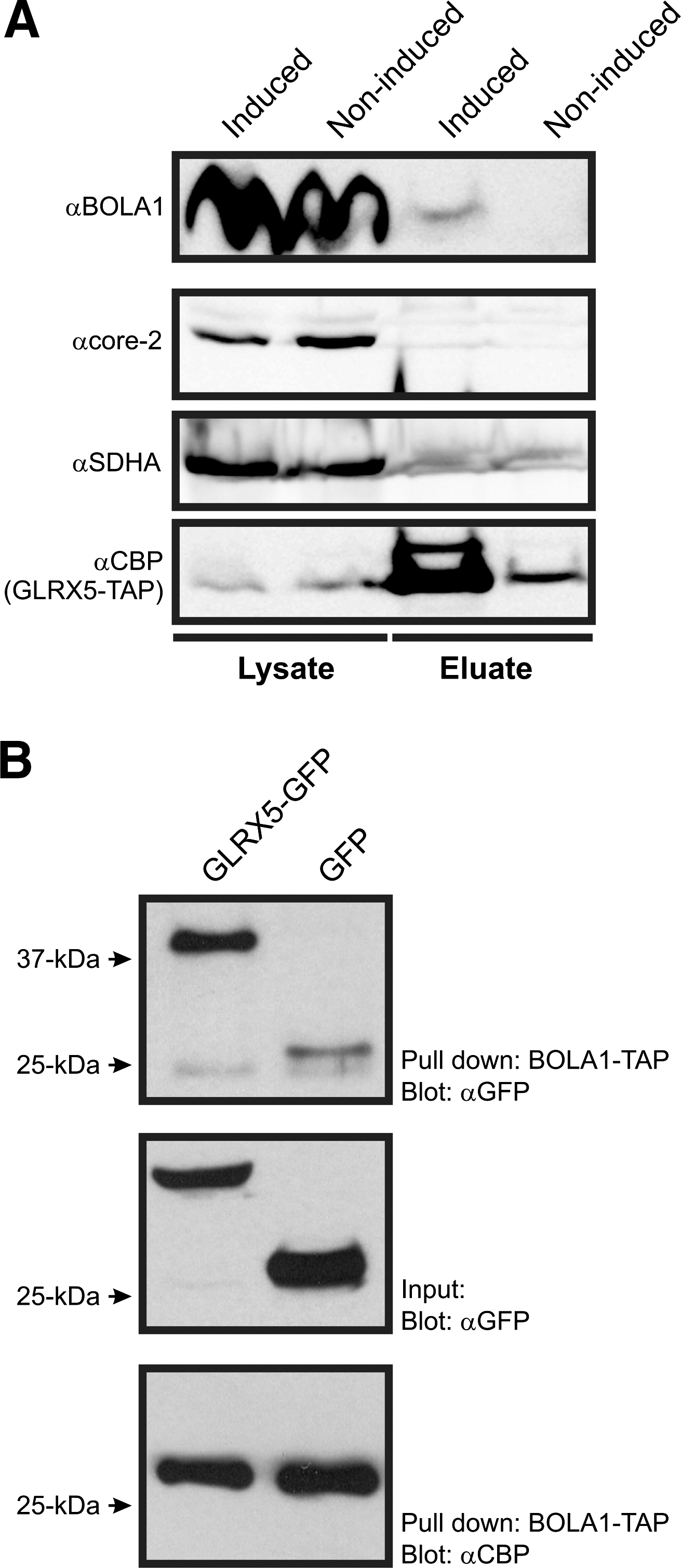
To demonstrate a physical interaction between BOLA1 and GLRX5, we generated HEK293 T-REx cells that inducibly express these proteins with a C-terminal TAP tag. At 24 h after induction, cell lysates were affinity purified and proteins were identified by nanospray ionization liquid chromatography tandem mass spectrometry (nLC-MS/MS). With BOLA1-TAP as a bait, GLRX5 was among the 3 copurified mitochondrial proteins that were specific to BOLA1-TAP, in the sense that they were not also observed in noninduced cells, or in affinity purifications of seven other mitochondrial proteins (Supplementary Table S1a). Conversely, BOLA1 was among the eight copurified mitochondrial proteins that were only copurified with GLRX5-TAP (Supplementary Table S1b). To verify the nLC-MS/MS data, the affinity purifications of BOLA1 and GLRX5 were repeated, and subsequently tested by Western blotting for copurification of GLRX5 and BOLA1, respectively. BOLA1 was copurified with GLRX5-TAP (Fig. 3A). Nevertheless, purification of BOLA1-TAP did not yield detectable amounts of copurified GLRX5 (data not shown). To substantiate the BOLA1-TAP GLRX5 interaction, we mixed lysates of BOLA1-TAP-expressing cells with those of either GLRX5-GFP- or GFP-expressing cells and incubated these with streptactin beads. Possible copurification of GLRX5-GFP by BOLA1-TAP was verified by Western blotting using anti-GFP. This approach resulted in a significant amount of BOLA1-TAP copurified GLRX5-GFP with respect to the GFP control (Fig. 3B, upper panel) strengthening the evidence for an interaction.
BOLA1 knockdown causes an oxidative shift of the mitochondrial thiol redox potential
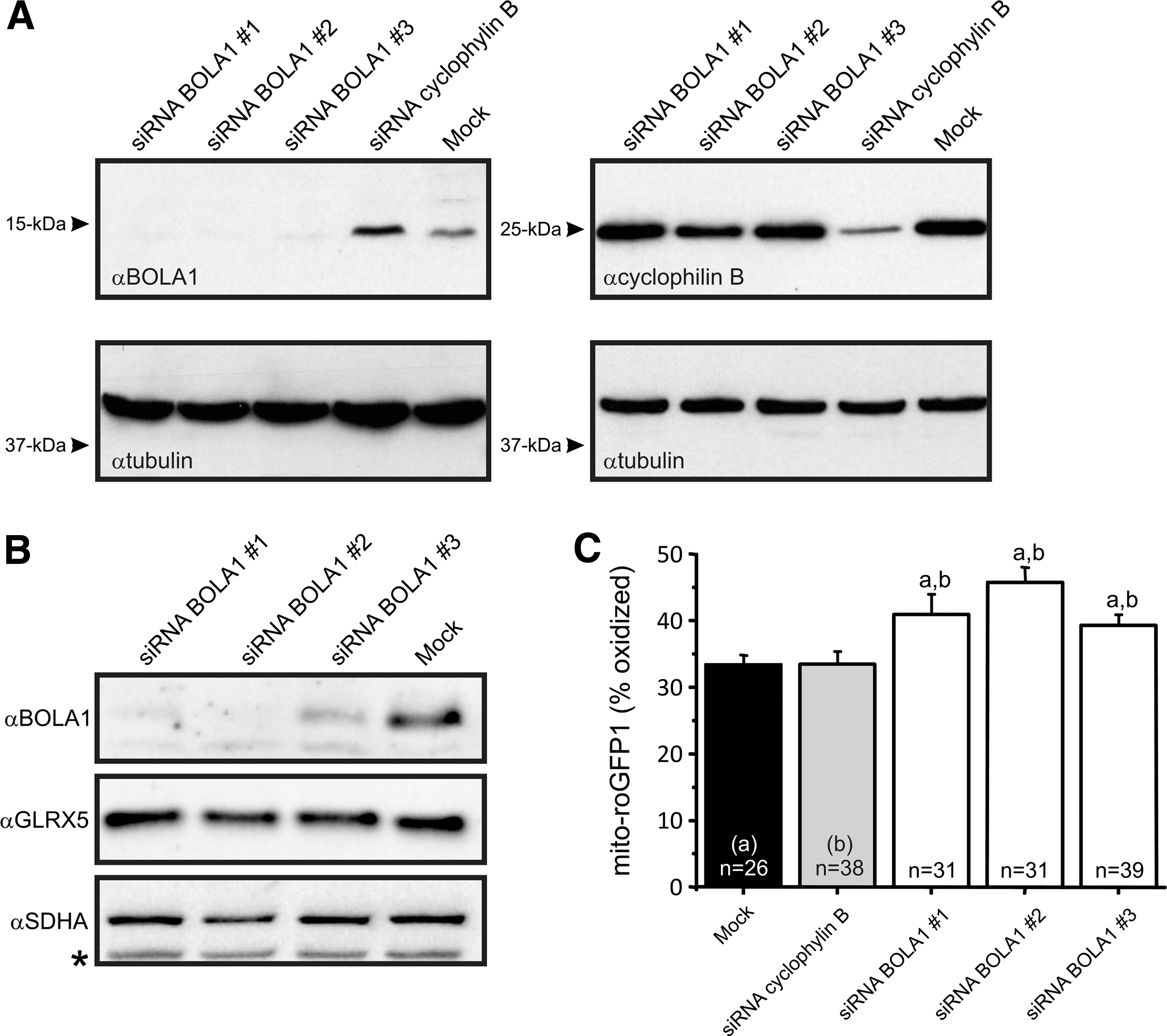
Because a bacterial homolog of BOLA1, BolA, is overexpressed under oxidative stress conditions (31), we investigated whether BOLA1 plays a role in the maintenance of the mitochondrial thiol redox potential. To this end, HeLa cells were transfected with small interfering RNAs (siRNAs) against BOLA1 mRNA. All three siRNAs effectively knocked down BOLA1 (Fig. 4A). BOLA1 knockdown did not alter the amount of GLRX5 (Fig. 4B). Digital-imaging microscopy of cells expressing mito-roGFP1, revealed that BOLA1 knockdown caused an oxidative shift of the mitochondrial thiol/disulfide redox status (Fig. 4C). This shift was not paralleled by a detectable change in the production of hydroethidine (HEt)-specific oxidants as revealed by digital-imaging microscopy of cells loaded with HEt (Supplementary Fig. S5). HEt oxidation measures superoxide production, whereas mito-roGFP1 measures thiol oxidation, processes that have been shown to be rather independent from each other, for example, L-buthionine-(S,R)-sulfoximine (BSO)-induced thiol oxidation can occur in parallel with decreased HEt oxidation (35).
BOLA1 overexpression attenuates the effect of glutathione depletion on the mitochondrial thiol redox potential
To investigate a putative role of BOLA1 in the maintenance of the mitochondrial thiol redox potential, fibroblasts were cotransduced with a baculovirus for expression of BOLA1-RFP and a baculovirus for expression of mito-roGFP1. Although BOLA1-GFP overexpression tended to cause a reductive shift of the mitochondrial thiol/disulfide redox status, this effect did not reach statistical significance (Fig. 5A).
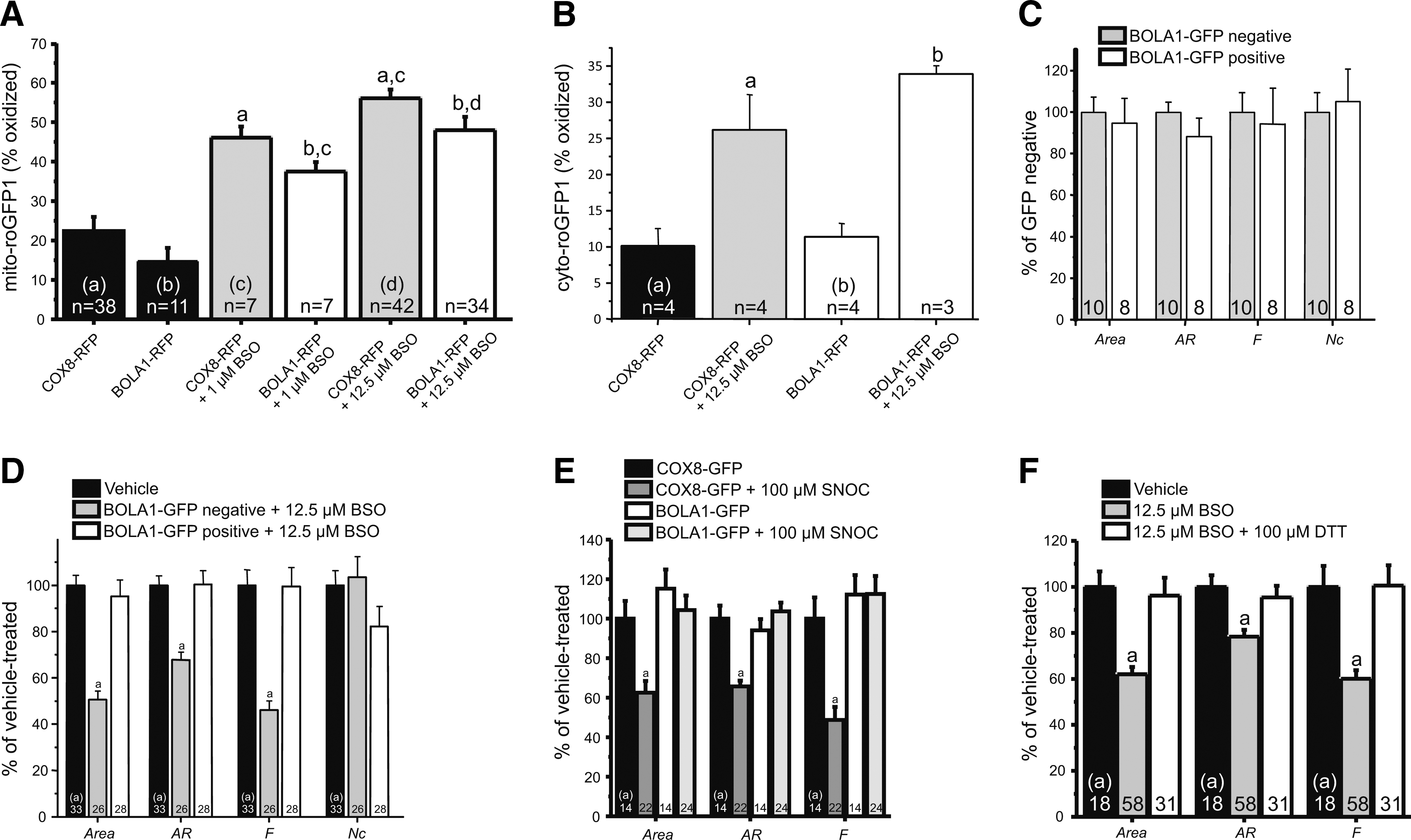
We previously published that 27-h treatment of fibroblasts with 12.5 μM BSO, an inhibitor of the glutathione (GSH)-synthesizing enzyme γ-glutamylcysteine synthetase, caused an oxidative shift of both the cytosolic and mitochondrial thiol/disulfide redox status (35). Here we show that BOLA1-RFP overexpression significantly reduced this effect of BSO in cells treated for 72 h with either 1 μM or 12.5 μM BSO of this inhibitor (Fig. 5A). In agreement with its mitochondrial localization, BOLA1-RFP overexpression did not alter the effect of BSO on the cytosolic thiol redox potential (Fig. 5B), suggesting a compartmentalized regulation of this parameter.
Digital-imaging microscopy of BOLA1-GFP overexpressing fibroblasts loaded with monochlorobimane, a fluorescent indicator of intracellular GSH, revealed that BSO decreased the amount of GSH equally potently in BOLA1-GFP-negative and -positive cells present on the same coverslip, with more than 80% decrease in GSH at 12.5 μM BSO (Supplementary Fig. S6A). This result indicates that BOLA1-GFP overexpression has no effect on BSO-induced GSH depletion.
BOLA1 overexpression prevents the change in mitochondrial shape under GSH depletion
To address a possible effect of BOLA1 on the mitochondrial shape and number, we applied our method for quantitative analysis of these parameters (22) to fibroblasts overexpressing BOLA1-GFP. Mitochondria were visualized by using TMRM. Our analysis revealed that the mitochondrial area, the aspect ratio (AR, a measure of mitochondrial length), the form factor (F, a measure of mitochondrial length and degree of branching) and the number of mitochondria per cell (Nc) were not significantly different between BOLA1-GFP-negative and -positive cells lying next to each other on the same coverslip (Supplementary Fig. S1A and Fig. 5C).
Previous evidence suggests that fibroblast mitochondria become longer and more branched under conditions of increased HEt oxidation (21). Here we show that fibroblast mitochondria become shorter and less branched upon treatment with BSO (Supplementary Fig. S1C and Fig. 5D). The effect of BSO on the mitochondrial area, AR and F was completely absent in BOLA1-GFP-positive cells present on the same coverslip (Supplementary Fig. S1D and Fig. 5D). This result was not observed with BOLA3, the other mitochondria-localized member of the BolA family (6) (Supplementary Fig. S6B). Note that BSO treatment did not alter the amount of mitochondria per cell, suggesting mitochondrial shrinkage rather than fragmentation. BOLA1 overexpression did not counter all the effects of BSO, as it failed to restore NAD(P)H levels, which are significantly increased in BSO-treated cells (Supplementary Fig. S6C) (35).
Similarly to BSO, the NO donor S-nitrosocysteine (SNOC; 100 μM, 72 h) that nitrosylates thiol groups and reduces the amount of GSH in the cell (3), caused a marked decrease in area, AR and F, which was again completely nullified by overexpression of BOLA1-GFP (Fig. 5E). Conversely, and similarly to BOLA1-GFP overexpression, the sulfhydryl-reducing agent dithiothreitol (DTT; 100 μM, 72 h) prevented the BSO-induced change in the mitochondrial shape (Fig. 5F).
Discussion
To date, experimental data on the biological function of BOLA1 are lacking. Here we show that BOLA1 is a mitochondrial protein that, in agreement with comparative genomics analyses, appears to interact with the mitochondrial monothiol glutaredoxin GLRX5. Recent work in human (5) and zebrafish (39) has linked GLRX5 deficiency to decreased haem synthesis in erythroid cells, and it has been linked to protection against oxidative stress in osteoblasts (26). It has a single glutaredoxin domain, and although it can reduce GSH mixed disulfides, it does so at a rate 100 times lower than the dithiol glutaredoxin GLRX2 (18). Furthermore Grx5 in S. cerevisiae is only inefficiently reduced by GSH (34). One explanation for the low catalytic efficiency of Grx5 and GLRX5 is that they interact with a protein that increases the rate at which they reduce GSH mixed disulfides and/or at which they are reduced by GSH. Here we provide first evidence that BOLA1 might exert such a role and that one of the target proteins of the BOLA1/GLRX5 complex is involved in maintaining the normal mitochondrial shape. It should thereby be noted that, although BolA's 3D structure is similar to that of the bacterial reductase OsmC, suggesting a reducing activity of BolA itself, BolA1 does not contain any conserved cysteines, rendering any reducing function dependent on an external source of reducing equivalents (16).
Knockdown of endogenous BOLA1 significantly increased the amount of oxidized mito-roGFP1. Because mito-roGFP1 senses the redox potential of the mitochondrial GSH buffer, this indicates that BOLA1 plays a role in keeping this buffer in a reduced state. As evidence has been provided that glutaredoxins act as mediators of reversible electron flow between GSH and mito-roGFP1 (27), changes in the affinity of a glutaredoxin toward GSH and/or its endogenous protein substrates might lead to alterations in the degree of oxidation of these latter molecules. Conversely, when fibroblasts were treated with BSO to decrease the amount of GSH, BOLA1 overexpression attenuated the BSO-induced increase in mito-roGFP1 oxidation without interfering with the BSO-induced decrease in GSH. These observations are compatible with a model in which the observed physical interaction between BOLA1 and GLRX5 leads to an increase in GLRX5 activity and, therewith, to maintenance of the reduced state of mito-roGFP1 at decreased GSH concentrations. Nevertheless, GLRX5 protein levels do not depend on BOLA1 expression, and we have no direct evidence that BOLA1's effect on mitochondrial morphology and thiol redox potential depends on an interaction with GLRX5.
We provide a first indication for a possible physiological role of BOLA1. Both BSO and SNOC caused a marked decrease in mitochondrial length and degree of branching, which was completely prevented by overexpression of BOLA1. The effect of BOLA1 was mimicked by DTT, indicating that a putative target protein of the BOLA1 has to be in the reduced state to perform its effect. Our observation that BOLA1 could also prevent the SNOC-induced change in the mitochondrial shape can be explained by a SNOC-induced increased oxidation of GSH, thus reducing the amount of GSH available, for example, for GLRX5, to keep the putative target protein in the reduced state. SNOC has been shown to cause a fragmented mitochondrial phenotype, in part, through S-nitrosylation of the mitochondrial fission protein Drp1 (7). Here we show that BSO treatment did not alter the amount of mitochondria per cell, suggesting mitochondrial shrinkage rather than fragmentation. The effect of SNOC on the mitochondrial shape was completely reversed by overexpression of BOLA1. An alternative explanation, that BOLA1 would directly prevent SNOC-induced S-nitrosylation of Drp1, rather than reduce the effect of SNOC-induced increase of GSH oxidation, remains to be established. This might be of critical importance because beta-amyloid protein found in brains of patients with Alzheimer's disease was found to induce S-nitrosylation of Drp1 and S-nitrosylated Drp1 was shown to be increased in brains of these patients, suggesting that S-nitrosylation of Drp1 might contribute to the pathogenesis of neurodegeneration (7).
In bacterial operons, BolA tends to occur not only with a monothiol glutaredoxin, but also with proteins involved in defense against oxidative stress: glutathione-S-transferase and Toluene exporters (16). We found, however, no evidence for upregulation of BOLA1 under conditions of oxidative stress in human gene expression data (Supplementary Table S2). Nor did we detect increased oxidation of HEt, which measures superoxides, in BOLA1 knockdowns, suggesting that BOLA1 is specific for thiol oxidation. Alternatively, to directly affecting the thiol oxidation state of target proteins, BOLA1 could have an indirect effect, for example, by being involved in the assembly of Fe-S clusters, like its mitochondrial paralog BOLA3 (6). Interactions between Grx5 and the Fe-S assembly protein ISA1 have been measured in S. cerevisiae (36) and in Schizosaccharomyces pombe (19), implicating Grx5 in Fe-S assembly, and GLRX5 has been implicated in Fe-S cluster assembly in human erythroblasts (41). The nonassembly of Fe-S clusters in mitochondria will lead to free iron, which via Fenton chemistry can lead to the formation of hydroxyl radicals, and therewith to increased oxidation of biomolecules. The increased thiol redox potential that we observe in BOLA1 knockdowns could thus be an indirect effect of the inability to assemble Fe-S clusters. Also, a fusion protein of BolA1 and the chloroplastic Fe-S assembly protein SufE in plants (40) argue for a role of BolA1 in Fe-S assembly or repair. Nevertheless, mitochondrial Fe-S assembly proteins, including Grx5 (Supplementary Fig. S3) have a wider phylogenetic distribution than only aerobic species. BolA1's strictly aerobic genomic occurrence, its reducing effect on the mitochondrial thiol redox potential, its interaction with Grx5 and the role in Fe-S assembly of the latter, suggest that BolA1 functions by increasing an aspect of Grx5's activity in Fe-S assembly or another process that is specifically required under aerobic, thiol-oxidizing conditions.
Materials and Methods
Sequence analysis and phylogenetic analysis
Eukaryotic orthologs of the BolA protein family were collected using PSI-Blast (default parameters) with BOLA1 as query and iterating until convergence against the NCBI nonredundant protein database in August 2011. To the eukaryotic sequences, E. coli BolA representatives (YrbA and BolA) were added. Alignment for the BolA domain was derived with ClustalX (24) followed by manual adjustments. Representatives from the eukaryotic crown groups, including all the anaerobic species and the proteins with experimental data, were selected for alignment (Supplementary Fig. S4) and phylogeny (Fig. 1). The maximum likelihood phylogeny was computed with PhyML (13), using the LG amino acid substitution model with a Gamma distribution approximated by 4 discrete-rate categories (4G). The proportion of invariant sites (I) and the gamma shape parameter (alpha) were estimated from the data. The Akaike Information Criterion implemented in ProtTest (v2.4) (1) was used to choose the evolutionary model and its parameters. The same procedure was followed for the monothiol glutaredoxins (Supplementary Fig. S3). We refer to the human proteins with BOLA1, BOLA2, BOLA3, and GLRX5, while we use BolA1, BolA2, BolA3, and Grx5 to refer to the orthologous groups that contain these proteins.
Primary human skin fibroblasts and inducible HEK293 and HeLa cell lines
Fibroblasts were obtained according to the relevant Institutional Review Boards from a skin biopsy of a healthy subject and cultured in the HEPES (25 mM)-buffered M199 medium (Invitrogen) supplemented with 10% (v/v) fetal calf serum (FCS), 100 IU/ml penicillin (Gibco), and 100 IU/ml streptomycin (Gibco). Generation of HEK293 cell lines for inducible expression of AcGFP1- and TAP-tagged BOLA1 and TAP-tagged GLRX5 is described in Supplementary Materials and Methods. HEK293 and HeLa cells were cultured in Dulbecco's modified Eagle medium (DMEM; Biowhitaker) with the same additions. T-REx™ Flp-In™ HEK293 cells were cultured in the presence of 5 μg/ml blasticidin (Invitrogen) and 300 μg/ml zeocin (Invitrogen).
Baculoviruses for transient overexpression of proteins of interest in human skin fibroblasts
Baculoviruses for cyto- and mito-roGFP1 are described elsewhere (35). Viruses for AcGFP1-tagged BolA homolog 1 (BOLA1-GFP; BC_063405), 2 (BOLA2-GFP; AF060511), and 3 (BOLA3-GFP; NM_21552) and cytochrome c oxidase subunit VIII (COX8) targeting sequence (COX8-GFP) and for TagRFP-tagged BOLA1 (BOLA1-RFP) and COX8 targeting sequence (COX8-RFP) were generated as reported previously (15). The BOLA protein entry vectors were purchased from Open Biosystems. Briefly, cDNA of the protein of interest was cloned into a modified pFastBacTMDual vector behind a CMV promoter and adjacent to a C-terminal GFP or TagRFP sequence. The obtained constructs were used to generate infectious recombinant baculoviruses by site-specific transposing-mediated insertion into baculovirus genome (barmaid). Isolated recombinant barmaids were used for virus amplification in Spodoptera frugiperda 9 insect cells.
BOLA1 knockdown and analysis of its effect on the cytosolic and mitochondrial thiol redox potential and the rate of HEt oxidation
Three BOLA1 siRNAs were designed using online software provided by the Whitehead Institute for Biomedical Research (42): #1 antisense strand: 5′-UUAACAUGGAACAUCCGGGdTdT, #2 antisense strand: 5′-UUGUUUCCAACUCAUCAGGdTdT, #3 antisense strand: 5′-ACUGUAUCAAAGGGAAGGCdTdT. The siRNAs were synthesized by Biolegio. Transfection of HeLa cells with siRNA as described before (37). Briefly, HeLa cells, plated in DMEM supplemented with 10% (v/v) FCS (without antibiotics), were transfected with siRNA using Dharmafect 1 transfection reagent (Dharmacon) and Optimem (Gibco). Forty-eight hours later, the cells were splitted 1:5, plated on glass bottom dishes (Willco Wells), and incubated overnight. After a second round of transfection, DMEM was replaced with M199 (without antibiotics) containing mito- or cyto-roGFP coding baculovirus (35). Ninety-six hours later, M199 was changed to HEPES–Tris containing 132 mM NaCl, 4.2 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 5.5 mM D-glucose, and 10 mM HEPES (pH 7.4) and cells were subjected to digital-imaging microscopy of roGFP and HEt oxidation products fluorescence (for details, see Supplementary Materials and Methods). Immunoblot analysis (see below) with anti-BOLA1 was used to demonstrate the effectiveness of the siRNAs.
BOLA overexpression and analysis of its intracellular localization and effect on BSO- and SNOC-induced changes in mitochondrial morphology and cytosolic and mitochondrial thiol redox potential
To determine the intracellular localization of BOLA1, 2, and 3 and the effect of BOLA1 on mitochondrial morphology, fibroblasts were seeded on glass bottom dishes, cultured for 24 h, and transduced with the appropriate baculovirus for expression of their GFP-tagged version. At 96 h after transduction, mitochondria were stained with TMRM and cells were subjected to digital-imaging microscopy of GFP and TMRM fluorescence. Quantitative analysis of mitochondrial morphology was performed on TMRM images as described before (20). To assess the effect of BOLA1 on the cytosolic and mitochondrial thiol redox potential, cells were cotransduced with a baculovirus for BOLA1-RFP and a baculovirus for either cyto- or mito-roGFP1. For further details on roGFP measurements, see Supplementary Materials and Methods. BSO and SNOC were added at 24 h after transduction.
Single-step affinity purification and nanospray ionization liquid chromatography tandem mass spectrometry and immunoblot analysis
Single-step affinity purification and nano-LC-MS/MS and immunoblot analysis were performed as described previously (38). For single-step affinity purification of BOLA1-TAP and GLRX5-TAP, cells were resuspended in the lysis buffer provided with the InterPlay TAP Purification Kit (Agilent Technologies) and subjected to three cycles of freeze–thawing followed by 10-min centrifugation at 10,000 g. The supernatant was incubated for 2 h under rotation at 4°C in the presence of Strep-Tactin-Superflow beads (IBA). Next, the beads were washed six times with the streptavidin-binding buffer provided with the kit, supplemented with 0.1% (w/v) lauryl maltoside, and bound proteins were eluted with the Strep-tag elution buffer containing D-Desthiobiotin (IBA). Before being processed for nLC-MS/MS analysis or immunoblotting, eluates were concentrated by passing through a 3 kDa cutoff filter (Millipore). For details on nLC-MS/MS analysis, see Supplementary Materials and Methods. Proteins identified in noninduced cells or in TAP-purifications of background, mitochondrial proteins AURKAIP1, C12orf62, C7orf44, PET100, PET117, ES1, and C10orf65 for which we have no indication that they are functionally or physically linked to BOLA1 or GLRX5, were excluded. Immunoblotting of the concentrated eluates was done as described previously. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels were run with 80–300 μg of protein and resolved proteins were transferred onto a nitrocellulose or PVDF membrane for immunostaining with primary antibodies. The GLRX5 antibody was kindly provided by Dr. Wing-Hang Tong (Molecular Medicine Branch, National Institute of Child Health and Human Development, Bethesda).
Footnotes
Acknowledgments
This work was supported by an equipment grant of NWO (Netherlands Organization for Scientific Research, No: 911-02-008). R.S. is supported by CSBR (Centres for Systems Biology Research) from the Netherlands Organisation for Scientific Research (NWO; CSBR09/013V), B.F.J.W. by a Horizon grant (050-71-053), U.K. and J.E. by the Dutch Ministry of Economic Affairs (IOP Grant #IGE05003), I.D. by the Portuguese Foundation for Science and Technology-FCT (SFRH/BD/32959/2006), and by “BolsasRui Tavares 2010.”
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.