
Abstract
Introduction
Innovation
We show that thioredoxin (Trx), an important redox regulator, binds to the white spot syndrome virus (WSSV) immediate early gene #1 (IE1) under oxidizing conditions and rescues IE1's DNA binding ability. Trx-IE1 binding is mediated via Trx cysteine 62. IE1's CXXC motif and cysteine 55 are critical for its DNA binding activity. Double-strand RNA knockdown of shrimp Trx leads to reduced mortality and lower virus copy numbers in WSSV-infected shrimp, which suggests that viral pathogenicity is enhanced by WSSV's ability to hijack host Trx.
Our approach was first to use a cDNA microarray and real-time quantitative reverse transcriptase–polymerase chain reaction (RT-PCR) to confirm the increased transcription level of Trx in WSSV-infected P. monodon (Supplementary Protocols S1–4; Supplementary Data are available at
WSSV, white spot syndrome virus; ORF, open reading frame.
Results
PmTrx and WSSV IE1 protein expression levels after WSSV infection
After confirming that WSSV infection increased the transcription levels of PmTrx (Supplementary Fig. S1) and determining the full length sequence of the PmTrx gene (Supplementary Fig S2; GenBank Acc. No. HQ427879), we used immunoblotting to monitor PmTrx and WSSV IE1 protein levels after WSSV infection.
We found that PmTrx expression was up-regulated at 48 h after WSSV infection (hpi), while expression of the viral protein IE1 was first detected at 24 hpi (Fig. 1).

PmTrx binds directly to WSSV IE1 via PmTrx Cys62 (cysteine 62)
The direct protein–protein interaction between PmTrx and WSSV IE1 was investigated by a pull-down assay in the presence of an oxidizing agent or a reducing agent (diamide and dithiothreitol [DTT]), respectively. Immunoblotting showed that there was only a direct interaction between PmTrx and WSSV IE1 in the presence of the oxidative reagent diamide (Fig. 2A). After confirming the expression of the five PmTrx point mutants used in the assay (Fig. 2B, C), immunoblotting showed that PmTrx cysteine 62 was critical for IE1 binding (Fig. 2D). We conclude that a direct protein–protein interaction occurs between PmTrx and WSSV IE1 in the presence of diamide, a thiol-specific oxidant, and that this interaction is through the cysteine 62 of PmTrx.

Oxidative stress promotes PmTrx binding to WSSV IE1 in Sf9 cells
Based on the vectors pDHSP70-V5-His and pDHSP70-Flag-His (17), we constructed the V5-tagged EGFP-PmTrx plasmid pDHsp70-EGFP-PmTrx-V5-His and the Flag-tagged IE1 plasmid pDHsp70-IE1-Flag-His. After Sf9 cells were cotransfected for 16–18 h with these two plasmids, oxidative stress was applied for 30 min using 5 mM H2O2. When the cell lysates were subjected to a coimmunoprecipitation assay, the enhanced green fluorescent protein (EGFP)-PmTrx fusion protein only produced a clear signal under the conditions of oxidative stress (Fig. 3, upper panel).
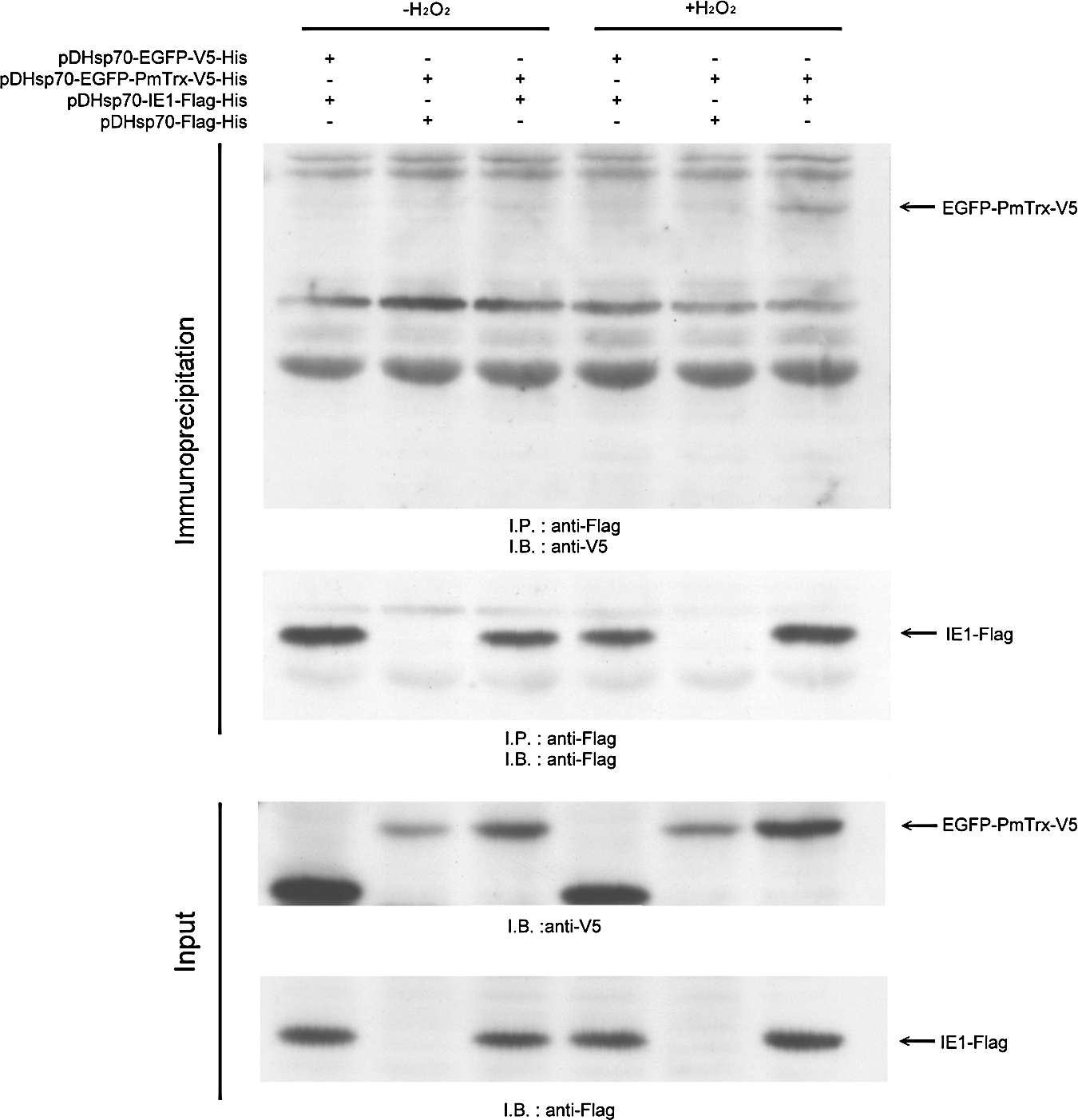
Redox controls the DNA binding activity of WSSV IE1
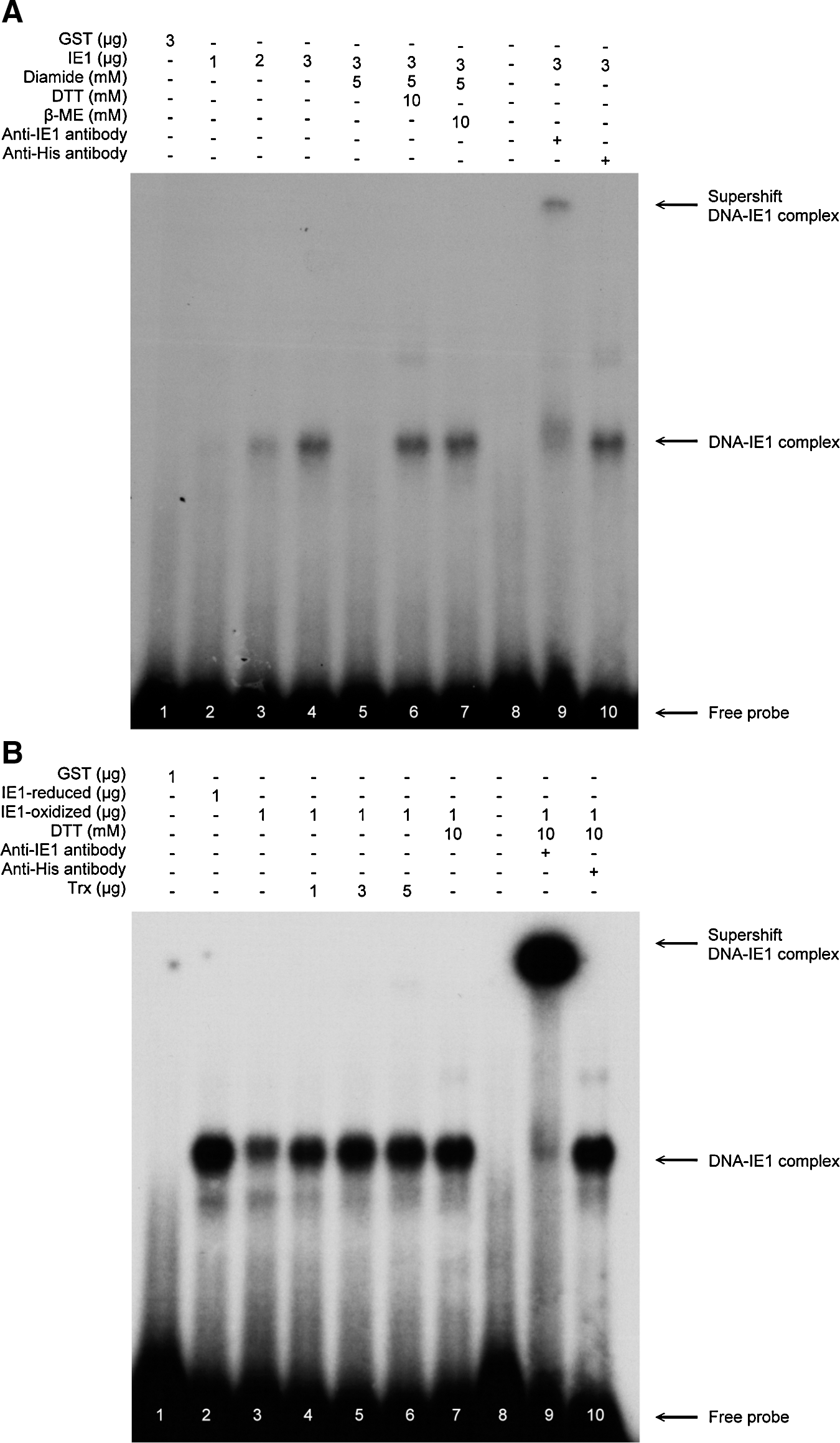
Our previous study showed that WSSV IE1 has DNA binding activity (25). Since the DNA binding activity of some transcriptional factors is known to be regulated by changes in their redox status, we speculated that the DNA binding activity of WSSV IE1 might also be controlled by redox and further that PmTrx might restore the DNA binding activity of inactived, oxidized WSSV IE1 by converting it into the reduced form. An electrophoretic mobility shift assay (EMSA) was performed to investigate this hypothesis. As found previously (25), the DNA binding activity of IE1 increased with increasing IE1 concentration (Fig. 4A, lanes 2–4). This binding activity was abolished by the addition of the thiol-specific reagent diamide in lane 5. However, with the further addition of the reducing agents DTT and beta mercaptoethanol (β-ME), IE1's DNA binding ability was restored (Fig. 4A, lanes 6, 7). We next investigated whether PmTrx could restore the DNA binding activity of oxidized WSSV IE1. Preliminary studies showed that Trx itself was oxidized by diamide and thus lost its disulfide reducing activity (data not shown). We therefore used a centricon device to remove the diamide (or DTT) after it had been used to produce the oxidized (or reduced) form of WSSV IE1. Figure 4B, lanes 2 and 3 show that, as expected, the oxidized form of WSSV IE1 had a lower DNA binding activity than the reduced form. The figure also shows that the DNA binding activity of the oxidized IE1 was recovered by incubating with increasing concentrations of PmTrx (Fig. 4B, lanes 4–6). These results strongly suggest that the redox status of WSSV IE1 controls its DNA binding activity and that PmTrx is likely to be involved in this control mechanism.
Three IE1 cysteine residues are critical for its DNA binding activity
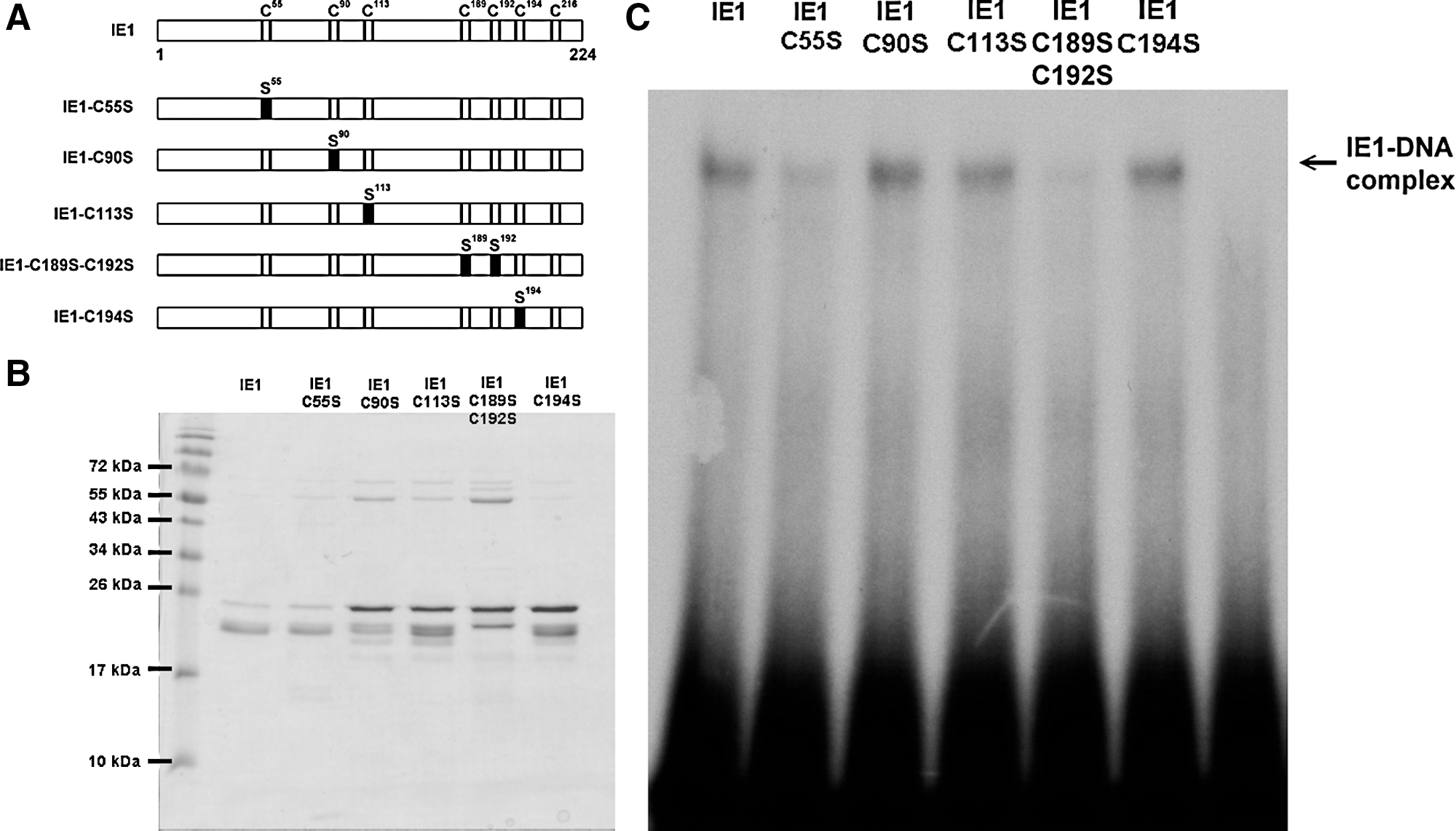
Although we tried to produce point mutants for all seven of the cysteine residues in IE1, we were only able to express and purify four single mutants (C55S, C90S, C113S, and C196S) and one double mutant (C189S-C192S) in sufficient quantity (Fig. 5A, B). Our EMSA results show that the DNA binding activity of C55S and the CXXC motif double-mutant IE1-C189S-C192S was markedly decreased (Fig. 5C).
Irreversible sulfhydryl modifying reagent inhibits the DNA binding activity of WSSV IE1
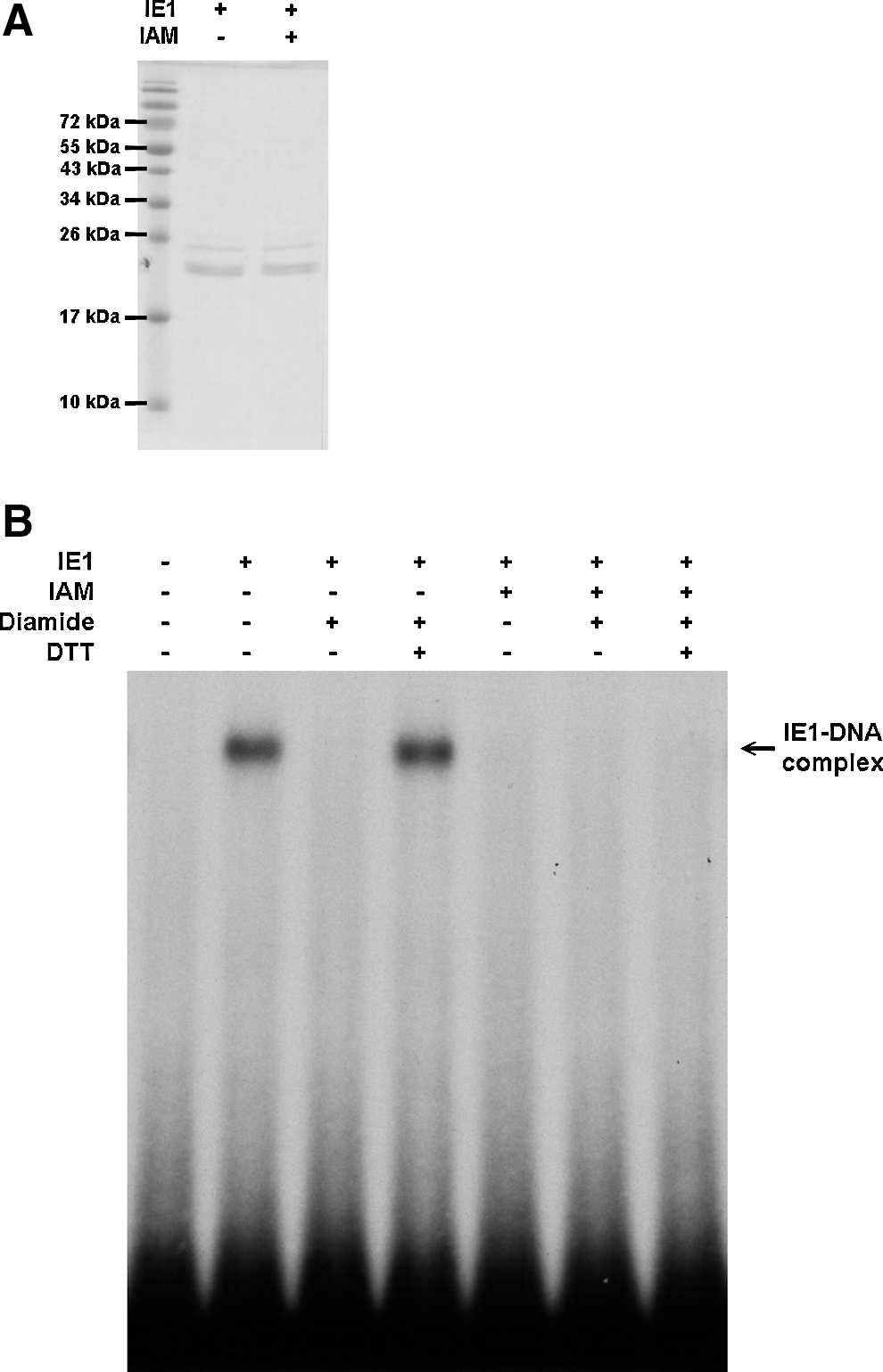
As a more practical alternative to the simultaneous mutation of all seven IE1 cysteine residues, we used the irreversible SH-modifying agent iodoacetamide (IAM) to alkylate all of the free cysteine residues on IE1. After IAM treatment, the DNA binding activity of IE1 was totally lost and no longer under redox control (Fig. 6).
Oxidative stress was observed in WSSV-infected shrimps at 48 hpi
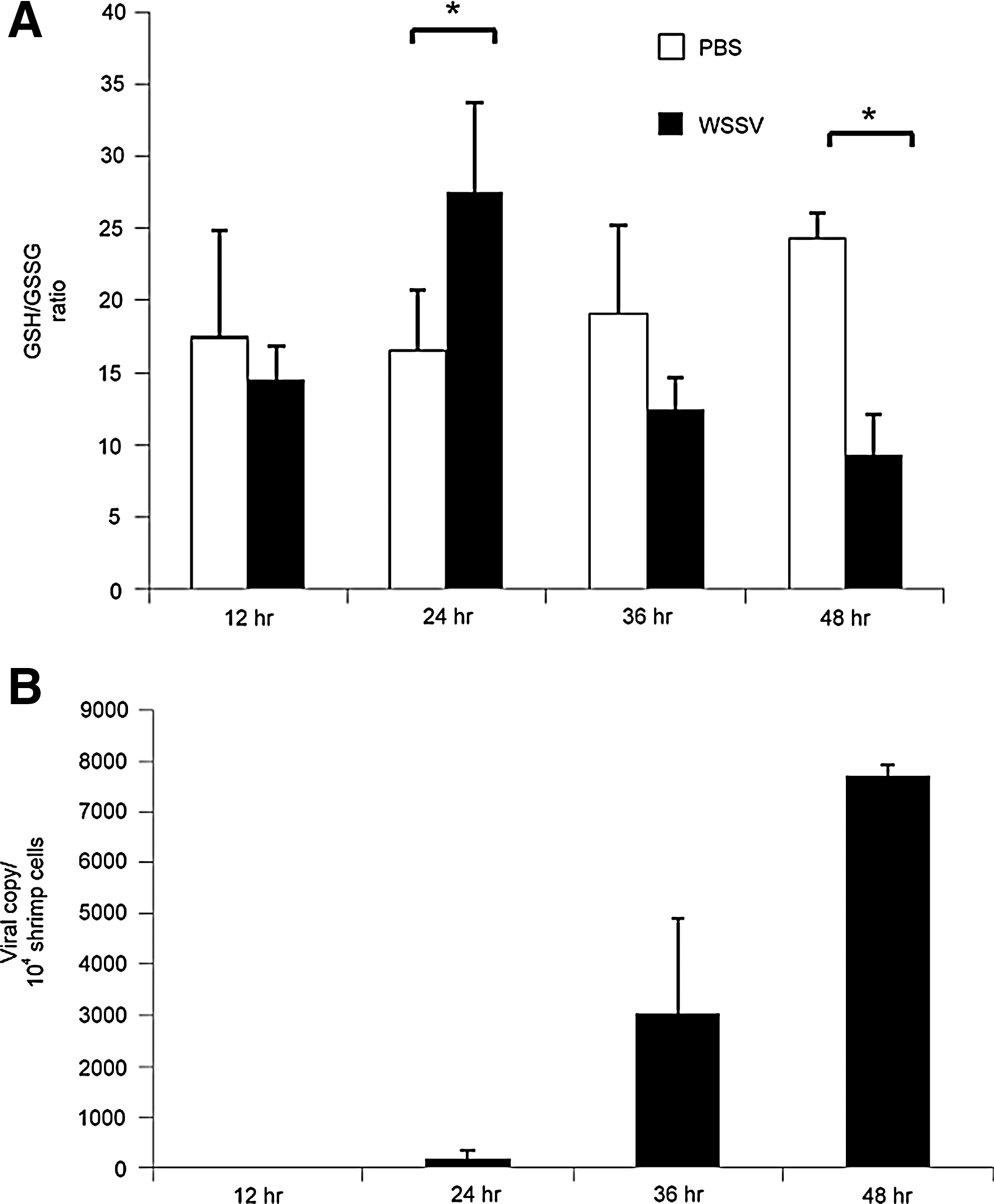
To monitor the cellular redox state during WSSV infection, the ratio of reduced glutathione to oxidized glutathione (GSH/GSSG) was measured using an LC/ESI/MS system. In phosphate-buffered saline (PBS)-injected shrimps, the GSH/GSSG ratio ranged from 10 to 25, but in WSSV-infected shrimps, the ratio was significantly up-regulated at 24 hpi, and significantly down-regulated at 48 hpi (Fig. 7A). Figure 7B shows the corresponding increase in viral copy number in the sampled WSSV-infected groups at the same time points.
Suppression of the shrimp Trx gene significantly reduces the mortality rate after challenge with WSSV
To explore the in vivo role of Trx during a WSSV infection, Trx expression was suppressed by a dsRNA-mediated RNA interference (33). Two days after injection, an RT-PCR confirmed that Trx was suppressed (Fig. 8A). When the dsTrx-treated shrimps were challenged with WSSV, we found that their viral copy numbers were lower than those in the dsEGFP control group at day 2, and this reduction reached statistical significance at day 3 (Fig. 8B). In the dsTrx-WSSV group, the cumulative mortality rate was also significantly lower at 3, 4, and 5 days postinfection (dpi) (Fig. 8C).

Discussion
Trx acts as a hydrogen donor that breaks disulfide bonds by transferring reducing equivalents to the disulfide groups in target proteins such that the oxidized disulfide (–S2) form is converted to the reduced dithiol [–(SH)2] form (11). Sequencing and alignment of the cDNA sequence of Trx from P. monodon showed that although PmTrx shares more than 90% identity with Litopenaeus vannamei Trx and Fenneropenaeus chinensis Trx, it is only around 50% identical to human Trx-1 (Supplementary Fig. S2B). Although none of the shrimp Trxs is highly similar to human Trx-1, they still exhibit disulfide-reducing activity (1). Sequence analysis further showed that like the other shrimp Trxs, PmTrx has four cysteines (Cys32, Cys35, Cys62, and Cys73), including the highly conserved catalytic site (-Cys32-Gly-Pro-Cys35-).
However, PmTrx-WSSV IE1 binding is evidently not mediated by the catalytic site because, as Figure 2 shows, the wild-type PmTrx and the single and double mutants are still able to bind to WSSV IE1. Binding is only prevented in the two mutants, where Cys62 has been replaced by serine. Wang et al. (44) found that the binding between human Trx1 and actin is likewise independent of the catalytic site of Trx and that it is mediated via Cys62 instead. We propose that a similar mechanism involving a disulfide bond at PmTrx Cys62 is also responsible for the interaction between PmTrx and WSSV IE1. Figure 2 also shows that PmTrx only binds to WSSV IE1 in the presence of oxidizing reagent diamide, which suggests that binding is under redox control.
PmTrx was also found to interact with WSSV IE1 in the absence of diamide in Sf9 cells exposed to oxidative stress (Fig. 3). These in vitro oxidizing conditions were designed to emulate those produced by a respiratory burst, which is an innate immune response in shrimp that causes the release of reactive oxygen species, including hydrogen peroxide and superoxide radicals. Indirect experimental evidence has already suggested that a respiratory burst is produced in shrimp in response to WSSV infection (47), and our new data show that oxidative stress does indeed occur in WSSV-infected shrimp at 48 hpi (Fig. 7A). We hypothesize that under the conditions of Figure 3, H2O2 probably oxidizes an IE1 thiol group to produce a sulfenic acid (–SOH), which then interacts with a thiol group on PmTrx to form a disulfide bond. If no PmTrx is present, then the IE1 sulfenic acid would presumably form an intramolecular disulfide bond.
The replication cycle of WSSV is completed within 24 h (45), and at this time, lysis will start to occur in some infected cells. However, at 24 hpi, Figure 7A shows an increased GSH/GSSH ratio, suggesting that reducing conditions are maintained in the cytosol of most cells at this time. In the intact cells, reducing conditions would provide a good environment for protein synthesis, which would be favorable to both viral progression and the host defenses. As the infection progresses through a second viral replication cycle, the number of infected cells increases exponentially (2), and these favorable conditions can no longer be maintained; consequently, oxidative stress is seen in Figure 7A at 48 hpi, which is close to the moribund stage of WSSV infection (i.e., when almost all of the cells will have lysed). [We should note that when Mohankumar and Ramasamy (31) monitored redox conditions using different methods, they reported a steady increase in oxidative stress in WSSV-infected Fenneropenaeus indicus. The reason for the discrepancy between their data and Fig. 7A is not yet known.]
The DNA binding activity of IE1 appears to depend on three of its cysteine residues, two of which (Cys189 and Cys192) comprise a CXXC motif (Fig. 5). The third residue important for DNA binding activity, Cys55, is located in the transactivation domain of IE (25), and we hypothesize that this cysteine residue might be essential because of some conformational change that it induces in IE1's structure.
The PmTrx/WSSV IE1 pull-down results (Fig. 2) suggest a redox-dependent interaction that is mediated by a disulfide bond. Figure 4 shows further evidence of a redox-dependent interaction between these two proteins. Specifically, PmTrx is able to reduce the oxidized form of IE1 and restore its DNA binding activity. In addition to these in vitro results, our dsRNA experiment shows that knockdown of shrimp Trx reduces both viral copy number and mortality in WSSV-challenged shrimp (Fig. 8). We hypothesize that these effects are mediated by the same interactions we demonstrated in vitro and that they are under redox control. In further support of this hypothesis, we note that a redox-controlled interaction between Trx and WSSV IE1 would presumably be of benefit to the virus in the oxidizing conditions that are usually induced during viral infection (34). As already shown (Fig. 7A), oxidizing conditions do indeed exist in shrimp at 48 h post challenge. In the absence of sufficient PmTrx, this means that IE1 would be oxidized and has no DNA binding activity. However, since PmTrx is up-regulated at this time (Fig. 1), it might restore (or maintain) IE1 to its reduced form and thus restore its DNA binding activity. [We note that WSSV IE1 is unlike most other immediate early genes in that it is highly expressed throughout the infection cycle (24), even though IE1's actual functions–if any–after the immediate early stage are presently unknown.]
Materials and Methods
Virus inoculum, shrimp, and virus challenge
The virus used in this study was the WSSV Taiwan isolate (AF440570) (3). Virus stock was prepared as described previously (39) and was stored at −80°C until use. The WSSV inoculum was prepared from the supernatant of the virus stock by centrifugation at 3000 g (10 min) and further dilution (10−2) with PBS as described previously (39).
The shrimps used in the immunoblotting assay and in the supplemental experiments were WSSV-free P. monodon (35–55 g) obtained from Tung-Kung Marine Laboratory, Taiwan Fisheries Research Institute. Shrimps in the experimental groups were challenged by intramuscular injection with 100 μl of virus inoculum (100× dilution of the viral stock), while shrimps in the control groups were injected with 100 μl PBS. All shrimps were cultured in 500 L tanks containing seawater (33‰ salinity) with a mean ambient temperature of 25°C. At various time points (0, 6, 24, and 48 h) after injection, the gills of the experimental shrimps (n=3) and control shrimps (n=3) were collected and frozen immediately in liquid nitrogen and stored at −80°C.
The shrimps used in the redox monitoring experiment were WSSV-free L. vannamei (∼10 g body weight) obtained from a commercial farm located in Pingtung, Taiwan. Shrimps in the experimental groups were challenged by intramuscular injection with 50 μl of virus inoculum (200× dilution of the viral stock), while shrimps in the control groups were injected with 50 μl PBS. All shrimps were cultured in 500 L tanks containing seawater (33‰ salinity) with a mean ambient temperature of 25°C. At various time points (12, 24, 36, and 48 h) after injection, the stomachs of the experimental shrimps (three groups; each group contained four shrimps) and control shrimps (three groups; each group contained four shrimps) were collected and frozen immediately in liquid nitrogen and stored at −80°C
The shrimps used in the dsRNA-mediated Trx knockdown experiment were WSSV-free L. vannamei (∼5 g body weight) obtained from the Terrestrial Animal Experimental Center, National Taiwan Ocean University, Keelung, Taiwan. After specific gene suppression, shrimps in the experimental groups were challenged by intramuscular injection with 100 μl of virus inoculum (2000× dilution of the viral stock), while shrimps in the control groups were injected with 100 μl PBS. Shrimps were cultured in 100 L tanks containing seawater (33‰ salinity) with a mean ambient temperature of 25°C.
Anti-Trx and anti-WSSV IE1 antibodies
For the immunoblottings, anti-Trx and anti-WSSV IE1 antibodies were prepared by cloning the PmTrx or WSSV ie1 coding sequence into the pET28b vector (see Table 3 and 4 for primers). The resulting constructs, pET28b-Trx and pET28b-WSSVIE1, respectively, were transformed into Escherichia coli strain BL21 Codon Plus cells. Recombinant proteins were purified on an Amicon Ultra-30 column (Millipore) and then separated on a sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis (PAGE). Protein concentration was determined using a Bio-Rad protein assay kit. Anti-Trx antibody and anti-WSSV IE1 antibodies were raised in rabbit using the purified recombinant PmTrx or WSSV IE1 as the antigen according to the conventional methods (15).
sm, single mutation.
dm, double mutation.
tm, triple mutation.
qm, quadruple mutation.
Restriction enzyme cutting sites and mutated sites are underlined.
Immunoblotting
Total protein was extracted from the excised P. monodon stomachs using a three-fold dilution of PBS. Samples of extracted protein (20 μg) were separated on 15% SDS-PAGE, and the separated proteins were transferred to a PVDF membrane (Millipore). PmTrx was detected with rabbit anti-Trx primary antibody and goat anti-rabbit secondary antibody (Chemicon). WSSV IE1 was detected with rabbit anti-WSSV IE1 primary antibody and goat anti-rabbit secondary antibody (Chemicon). P. monodon β-tubulin was used as the internal control and detected with rabbit anti-β-tubulin primary antibody and goat anti-rabbit secondary antibody (Chemicon). Target signals were visualized by an ECL system (Millipore) according to the manufacturer's instructions.
Preparation of recombinant IE1 and IE1 mutants
GST-IE1 fusion proteins were produced by transforming Escherichia coli BL21 Codon Plus cells with the plasmids pGEX5T-1 and pGEX5T-1-IE1 as described previously (25) with slight modifications. IE1 protein was obtained from the purified GST-IE1 by using a protease, factor Xa (10 units/1 mg GST fusion protein; Novagen), to remove the GST as described previously (25).
For the production of IE1 mutants, plasmids to express four single mutants (Cys55→Ser55, pGEX5T-1-IE1-C55S; Cys90→Ser90, pGEX5T-1-IE1-C90S; Cys113→Ser113, pGEX5T-1-IE1-C113S; and Cys196→Ser196, pGEX5T-1-IE1-C196S;) and one double mutant (Cys189,192→Ser189,192, pGEX5T-1-IE1-C189-192S) were then constructed by rolling-circle PCR using pGEX5T-1-IE1 as a template. After transformation of these constructs into Escherichia coli strain BL21 Codon Plus cells, the recombinant IE1 mutants were purified as described above. Primersets are listed in Tables 3 and 4.
Expression and purification of recombinant PmTrx and PmTrx mutants
For the production of recombinant PmTrx, the PmTrx coding sequence was cloned into the pET28b vector by PCR amplification. Plasmids to express the four mutants (Cys32→Ser32, pET28b-smTrx; Cys32,35→Ser32,35, pET28b-dmTrx, Cys32,35,62→Ser32,35,62, pET28b-tmTrx-C62S; Cys32,35,73→Ser32,35,73, pET28b-tmTrx-C73S; Cys32,35,62,73→Ser32,35,62,73, pET28b-qmTrx) were then constructed by rolling-circle PCR using pET28b-Trx, pET28b-dmTrx or pET28b-tmTrx-C62S as a template. After transformation of these constructs into Escherichia coli strain BL21 Codon Plus cells, the recombinant Trx, smTrx, dmTrx, tmTrx-C62S, tmTrx-C73S, and qmTrx proteins were purified without the use of reducing agents. Primers are listed in Tables 3 and 4.
Pull-down assays
To test whether PmTrx binds to WSSV IE1, purified recombinant Trx was mixed with IE1 in the presence of 1 mM DTT or 1 mM diamide. To identify which Trx cysteine residues are necessary for IE1 binding, Trx and its mutants were mixed with purified IE1 in the presence of 1 mM diamide. The His-tagged Trx and Trx mutants were pulled down using nickel-agarose beads (Qiagen) according to the manufacturer's instructions. Anti-Trx antibody was used to confirm pull down, and anti-IE1 antibody was used to detect the target protein.
Coimmunoprecipitation assay in Sf9 cells
Plasmids to express WSSV-IE1 and PmTrx proteins were constructed, respectively, from Drosophila heat shock protein 70 promoter-based expression vectors pDHsp70-Flag-His and pDHsp70-V5-His (17). To express IE1, the Flag-tagged WSSV IE1 expression plasmid, pDHsp70-IE1-Flag-His, was constructed by PCR using WSSV genomic DNA as a template. Expression of PmTrx was slightly more complicated: because the PmTrx coding regions are so short, the EGFP sequence was isolated from a commercial plasmid pEGFP-N1 (Clontech) by PCR and cloned into the pDHsp70-V5-His vector to produce pDHsp70-EGFP-V5-His. The V5-tagged EGFP-PmTrx expression plasmid pDHsp70-EGFP-PmTrx-V5-His was then constructed by PCR using pET28b-rTrx as a template.
For the coimmunoprecipitation assay, Sf9 cells (8×105 cell/well) were cotransfected with the above plasmids using an Effectene transfection kit (Qiagen) according to the manufacturer's protocol. After transfection (16–18 h), exposure to 5 mM H2O2 or PBS for 30 min and harvesting, proteins were pulled down with 15 μl anti-FLAG M2 agarose gel (Sigma), separated on 17.5% SDS-PAGE, and transferred to a PVDF membrane (Millipore). A V5-tagged protein was detected with rabbit anti-V5 primary antibody (Sigma) and goat anti-rabbit (Chemicon) secondary antibody. Flag-tagged protein was detected with rabbit anti-Flag primary antibody (Sigma) and goat anti-rabbit (Chemicon) secondary antibody. Targets signal were visualized using an ECL system (Millipore) according to the manufacturer's instructions.
Electrophoretic mobility shift assay
EMSAs used a 79 mer oligonucleotide (5′-GTCGCTCGA GCGGTATGACGAGATCTA(N)25TAGATCTGCGTCACTA GTCTAGACTAG-3′) as a random probe and were performed exactly as described previously (25) except that the quantity of poly(dI-dC) was reduced from 200 to 50 ng.
GSH/GSSG ratio measurement
The GSH/GSSG ratio was monitored because it serves as a good indicator of the cellular redox state (7, 32). Total metabolic materials were extracted from the L. vannamei stomach using cold 100% MeOH (tissue weight [mg]: buffer [μl] 1:20). The extracted lysate was centrifuged at 12,000 g for 10 min, and the supernatant was dried using a centrifugal evaporator (CVE-2000, EYELA). The pellet was suspended in a volume of deionized water that was equal to the original volume of the buffer. The reduced and oxidized forms of glutathione (GSH and GSSG) were measured by a LC/ESI/MS system in positive electrospray ionization.
WSSV copy number quantification
WSSV copy number quantification was performed using the commercial real-time PCR kit for viral load quantification (GeneReach Biotechnology Corp.) as described previously (22). Results were presented as means±standard deviation. Quantified data were analyzed using the Student's t-test. p<0.05 was considered statistically significant.
The effect of dsRNA-mediated Trx knockdown on WSSV-infected shrimps
Double-strand shrimp Trx RNA (dsTrx RNA) was generated by using the T7 RiboMAX Express Large Scale RNA System (Promega) following the manufacturer's instructions. Briefly, the DNA template of L. vannamei Trx and T7 promoter was generated by PCR amplification with the appropriate primers (LvTrx-ds-F/LvTrx-ds-R, 5′-ATGGTTTACCAAGTGAAAG-3′ and 5′-CTTGTTCTTCTCAACGAGTTC-3′; Lv-Trx-T7ds-F/Lv-Trx-T7ds-R 5′-TAATACGACTCACTATAGG GAGAATGGTTTACCAAGTGAAAG-3′ and 5′-TAATACGA CTCACTATAGGGAGACTTGTTCTTCTCAACGAGTTC-3′) for the single-strand RNA. The dsEGFP RNA was obtained as described previously (43). The experimental shrimps were treated with dsTrx or dsEGFP RNA at a dosage of 1 μg/g shimp in 100 μl PBS by intramuscular injection. A negative control group was injected with 100 μl PBS only.
To monitor the silencing effect, 2 days after dsRNA injection, the gills of the experimental group (dsTrx) and control groups (dsEGFP and PBS) were collected. RNA was extracted, and cDNA was obtained using the Trizol reagent (Invitrogen) and Superscriptase III reverse transcriptase (Invitrogen) according to the manufacturer's instructions. RT-PCR was used to confirm the gene suppression effect with the primers sets (LvTrx-F/LvTrx-R, 5′-ACTGCTCTCTCTGGCACCAAG-3′ and 5′-ATTCACCTGAAATAATTAGC-3′; EF-1α-F/EF-1α-R, 5′-ATGGTTGTCAACTTTGCCC-3′ and 5′-TTGACCTCCTTGATCACACC-3′).
For the WSSV copy number experiment, the pleopods of the experimental shrimps (dsTrx-WSSV injected) and control shrimps (PBS-WSSV injected and dsEGFP-WSSV injected) were collected at various time points (1, 2, and 3 dpi; three shrimps in each sample), frozen immediately in liquid nitrogen, and stored at −80°C. To determinate the WSSV copy number, the viral load was quantified using a real-time PCR as described above.
For the cumulative mortality experiment, shrimps (n=30 in each group) were injected with dsTrx, dsEGFP, or PBS only by intramuscular injection 2 days prior to WSSV challenge. Mortality was monitored every 12 h and recorded every day. Data were processed with the GraphPad Prism 5 (GraphPad Software) and presented using a Kaplan–Meier plot. Statistical significance was determined using the log rank χ 2 test.
Footnotes
Acknowledgments
This investigation was supported financially by National Science Council grants (100-2311-B-002-005-MY3 and 100-2321-B-002-025) and a National Taiwan University grant (CESRP-10R70602B5). The WSSV-free P. monodon shrimps were kindly provided by Kuan Fu Liu from Tung-Kung Marine Laboratory, Taiwan Fisheries Research Institute. We thank Chun Han Ho and Yu-Hsuan Tung for their helpful comments. We thank Paul Barlow for his helpful criticism of the article.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.