
Abstract
Introduction
Innovation
The in vivo study of the metazoan mitochondrial thioredoxin system function(s) is severely hampered by the lack of an appropriate model at the level of a complete organism. This critical limitation is illustrated by both the murine and Drosophila knock-out models that display embryonic or larval lethal phenotypes, respectively. The present work describes the functional characterization of the mitochondrial thioredoxin system of Caenorhabditis elegans. Our findings show that this nematode model is the first reported metazoan in which the absence of the mitochondrial thioredoxin system does not cause any lethal phenotype. Thus, the availability of a viable metazoan model lacking a functional mitochondrial system combined with the genetic amenability of C. elegans provide an ideal scenario to further investigate the in vivo functions of this system as well as to identify other redundant enzymatic systems functionally overlapping with the mitochondrial thioredoxin system.
Mitochondrial dysfunction and alterations in redox homeostasis have been unequivocally linked to the progression of many neurodegenerative diseases. Using Caenorhabditis elegans, we have uncovered a protective role of the mitochondrial thioredoxin reductase 2 against the paralysis phenotype caused by the aggregation of the β-amyloid peptide in the muscle cells of the nematode, a well-established invertebrate model of Alzheimer's disease (AD). This unexpected finding paves the way to use the strains generated in this work in pharmacological screens and other studies of clinical significance aimed at evaluating the role of mitochondria, and more specifically, that of the mitochondrial thioredoxin system, in the pathophysiology of AD.
Being an important source of ROS (4, 28), mitochondria are well protected against the deleterious effects of ROS by a set of antioxidant defenses such as superoxide dismutases, glutathione peroxidases, glutathione transferases, peroxiredoxins and specific mitochondrial thioredoxin and glutaredoxin systems (27). In addition to its key role as antioxidant defense (5, 44, 60), the mitochondrial thioredoxin system has been shown to be an important mediator of many processes where the mitochondrial function is crucial, such as apoptosis (55), angiogenesis (9), transcription factor DNA binding activity (46) or cell growth and differentiation (25, 38), among others. Interestingly, the mitochondrial thioredoxin system has been shown to functionally interact with the mitochondrial glutaredoxin system by directly reducing mitochondrial glutaredoxin (22). Furthermore, both mitochondrial systems display overlapping functions as, for example, in the reduction of the mitochondrial peroxiredoxin (18).
Mitochondrial thioredoxin systems are, in most cases, composed of dedicated thioredoxin reductase and thioredoxin proteins encoded by nuclear genes, different from those that encode the cytoplasmic thioredoxin system (36). An exception to this generality is found in Drosophila melanogaster, where the same gene (trxr-1) encodes both the cytoplasmic and the mitochondrial thioredoxin reductase isoforms by alternative splicing (37). Surprisingly, a mitochondria-specific thioredoxin has not yet been identified in this organism. In yeast, null mutants of the mitochondrial thioredoxin system (composed of trxr-2 and trx-3 genes) are viable but show enhanced sensitivity to oxidative stress (43). In contrast, inactivation of the mouse mitochondrial thioredoxin system trxr-2 and trx-2 genes results in early embryonic lethality (7, 40) and null mutants of the Drosophila trxr-1 gene die at the second instar larval stage (37). These lethal phenotypes in several model organisms entail important difficulties to genetically dissect the pathways in which the eukaryotic mitochondrial thioredoxin system participates.
In this context, we focused our attention on C. elegans, a well-established, genetically tractable model, in order to study the functions of the mitochondrial thioredoxin system in vivo and in the framework of a complete organism. Surprisingly, C. elegans mutants of the mitochondrial thioredoxin system are viable and do not show any apparent phenotype even under stress conditions. However, we have uncovered an unexpected protective role of the mitochondrial thioredoxin reductase trxr-2 gene in a worm model of Alzheimer's disease (AD) that shows an age-dependent progressive paralysis phenotype upon overexpression of the human β-amyloid peptide (31).
Results
C. elegans trx-2 and trxr-2 genes form a functional thioredoxin system
Previous in silico genomic analyses have demonstrated the presence of two different thioredoxin reductases in C. elegans encoded by the trxr-1 and trxr-2 genes, respectively (16). We have recently demonstrated that thioredoxin reductase 1 (TRXR-1) has a cytoplasmic localization (51), while TRXR-2 is assumed to be a mitochondrial protein based on the presence of a putative mitochondrial targeting sequence (MTS) at its N-terminus (Supplementary Fig. S1A; Supplementary Data are available online at
Two deletion alleles, tm2047 and ok2267, are available for the trxr-2 gene, which is composed of 6 exons and spans about 3 kb on linkage group III (

Similarly, two deletion alleles, tm2720 and ok1526, are also available for the trx-2 gene, which is composed of 3 exons and spans about 1.1 kb on linkage group V (
TRX-2 and TRXR-2 are mitochondrial proteins
To demonstrate TRX-2 and TRXR-2 subcellular localization in mitochondria as well as to identify the cells and tissues expressing these proteins within the nematode, we generated transgenic animals expressing transcriptional (promoter only) and translational (promoter plus gene) green fluorescent protein (GFP) fusion reporters. Noteworthy, trxr-2 is the third gene in an operon composed of the unc-32, tpk-1 and trxr-2 genes (
A transcriptional construct Ptrx-2::GFP was expressed only in AIYL/R and ASEL neurons as determined by the position of their cell bodies, morphology of their axons and dendrites and colocalization experiments with specific markers for these neurons (Fig. 2A–C and Supplementary Fig. S2). In some, but not all, transgenic animals, a fourth neuron identified as ASER also displayed fluorescent signals (data not shown). The unc-32 gene, the first gene of the trxr-2 operon, has been previously reported to be expressed in several cells and tissues such as the ventral nerve cord, nerve ring and spermathecal-uterine valve, among others (41). We confirmed most of these expression patterns in transgenic animals harboring a Punc-32::GFP construct and, in addition, we also detected expression in muscle cells, several neurons in the tail, intestinal cells and coelomocytes (Fig. 2D–F). This expression pattern was complemented with that of the trxr-2 internal promoter where a transcriptional construct Ptrxr-2::GFP was found to be expressed in the intestine plus additional neurons in the head (Fig. 2G–I). Interestingly, in L1 animals, only two (unidentified) neurons are marked, but the number of neurons showing fluorescence increases as the animal grows to adulthood (Fig. 2G–I). As a whole, the combined expression pattern of unc-32 and trxr-2 internal promoters suggests a basically ubiquitous expression of TRXR-2, which strongly contrasts the restricted expression for TRX-2 in only three to four head neurons.

Transgenic animals expressing translational Ptrx-2::trx-2::GFP, Punc-32::trxr-2::GFP and Ptrxr-2::trxr-2::GFP constructs all showed an expression pattern consistent with mitochondrial localization (Fig. 2J–P). Thus, Ptrx-2::trx-2::GFP was clearly expressed in muscle cells in the head, vulva or body wall where mitochondria display their typical tubular morphology (Fig. 2J, K). Indeed, the expression in head muscle cells precluded in most cases the identification of the AIYL/R and ASEL neurons labeling (Fig. 2J). The fact that muscle cells are marked only when using the translational construct Ptrx-2::trx-2::GFP but not the transcriptional construct Ptrx-2::GFP indicates that additional regulatory sequences directing muscle expression are present within the trx-2 genomic open reading frame. The translational Punc-32::trxr-2::GFP construct was found to be expressed in a punctate pattern in the head neurons and ventral nerve cord, while in body wall muscle cells and intestine, the mitochondria distribution was found in a tubular/reticular pattern excluding the nucleus (Fig. 2L, M). Finally, the Ptrxr-2::trxr-2::GFP translational construct expression demonstrated perinuclear punctate fluorescence in several head neurons and reticular labeling in the intestinal cells when transcribed from its internal promoter (Fig. 2N–P).
To unequivocally demonstrate in vivo that the patterns obtained with the translational fusion constructs do indeed pertain to mitochondria, we first generated expression constructs in which the putative TRXR-2 and TRX-2 MTS (Supplementary Fig. S1A and B, respectively) were directly fused to GFP. Transgenic animals harboring Ptrx-2::MTS::GFP, Punc-32::MTS::GFP and Ptrxr-2::MTS::GFP all confirmed that the punctate/reticular patterns obtained with the translational GFP constructs are indeed driven by their respective putative MTS (Fig. 3A–G). Next, we performed mitochondrial colocalization experiments in the worm body wall muscle cells, because this tissue expresses both TRX-2::GFP and TRXR-2::GFP fusion proteins from the trx-2 and unc-32 promoters (Fig. 2J–L). As mitochondrial marker, we used a fusion protein of C. elegans mitochondrial import receptor subunit TOMM-20 and monomeric red fluorescent protein (mRFP), whose expression is driven in body wall muscle cells by the myo-3 promoter (Supplementary Table S1, a kind gift of Dr. Amir Sapir and Dr. Paul Sternberg). As shown in Figure 3H and I, the mitochondrial reporter TOMM-20::mRFP colocalizes with both TRX-2::GFP and TRXR-2::GFP fusion proteins expressed in body wall muscle cells from their respective trx-2 and unc-32 promoters. Taken together, all these experiments demonstrate that both trx-2 and trxr-2 genes encode the C. elegans mitochondrial thioredoxin system.

The mitochondrial thioredoxin system is dispensable for C. elegans development
Next, we set out to evaluate the phenotypic effect of the lack of trx-2 and trxr-2 genes during C. elegans development. To our surprise, and in sharp contrast with the early embryonic lethal phenotypes displayed by their mammalian orthologs (7, 40), C. elegans trxr-2 and trx-2 single mutants were viable, grew normally to adulthood with no apparent embryonic or postembryonic defects, were fertile and had normal mitochondrial morphology (Supplementary Table S2 and Supplementary Fig. S3). Similarly, double mutants trx-2 (tm2720); trxr-2 (ok2267) and trx-2 (tm2720); trxr-2 (tm2047) did not display any obvious phenotype.
Apoptosis is a developmental process where the mammalian mitochondrial thioredoxin system has been shown to play a key role (10, 42). In fact, trx-2 knock-out mice embryos die by massive apoptosis, while liver apoptosis has been shown to partially underlie the lethal embryonic phenotype of trxr-2 knock-out mice (7, 40). During C. elegans embryogenesis, the apoptotic program is invariant with 113 cells undergoing apoptosis out of 671 cells generated from zygote to newly hatched larva (54). During postembryonic development, 18 additional cells die by programmed cell death, thus making up a total of 131 somatic cells undergoing apoptosis in C. elegans (53). Furthermore, in the adult nematode germ line, another round of cell death takes place to reduce the number of cells that complete oogenesis (17). Given the important role of the mitochondrial thioredoxin system in the apoptotic process in other organisms, we investigated whether C. elegans trx-2 and/or trxr-2 mutants exhibited defective regulation of apoptosis. To this purpose, we used four-dimensional differential interference contrast (DIC) timelapse video microscopy in trx-2 and/or trxr-2 mutant embryos to determine the kinetics of apoptosis of the first 13 programmed cell death events that occur in the AB lineage (39). As shown in Figure 4, we found no significant alteration of cell death kinetics of the mitochondrial thioredoxin system single and double mutants compared with wild-type control embryos. We also measured the penetrance of germ line cell death in adults of the same set of mutants and again found no significant difference in the kinetics of apoptosis in this tissue [wild type 1.00±1.17; trx-2 (tm2720) 0.90±1.07; trxr-2 (tm2047) 0.95±0.88 and trx-2 (tm2720); trxr-2 (tm2047) 1.05±0.94 (Numbers indicate the average±standard deviation of apoptotic corpses per gonad arm, n=20, p>0.05 in all cases by unpaired two-tailed t-test)]. Together, these results demonstrate that in C. elegans, the mitochondrial thioredoxin system is dispensable for normal development under standard growth conditions and does not regulate apoptosis.

The mitochondrial thioredoxin system is not a major defense system against stress but might play a role in the mitochondrial unfolded protein response
The mammalian mitochondrial thioredoxin system functions as a key defense system against oxidative stress and other cellular insults either directly (19, 50) or by reducing other antioxidant enzymes such as peroxiredoxins (18). To determine whether this protective function is also conserved in worms, we evaluated the sensitivity of trx-2 and trxr-2 mutants exposed to different agents and treatments that induce oxidative stress. No significant differences in sensitivity were found with arsenite, juglone, sodium azide, paraquat or thermal stress treatments (Fig. 5A–E), although the trx-2 (tm2720) mutant was slightly more resistant to thermal stress and trx-2 (tm2720) single and trx-2 (tm2720); trxr-2 (tm2047) double mutant showed a slightly higher resistance to paraquat treatment. Consistent with a dispensable role as a major defense system against oxidative stress, trx-2 and trxr-2 single and double mutants do not show increased ROS production either under basal unstressed conditions or in the presence of 1 mM sodium azide or 0.8 mM paraquat (Supplementary Fig. S4). Besides, efficient RNA interference (RNAi) downregulation of C. elegans trx-2 and trxr-2 gene expression (Supplementary Fig. S5A) in animals carrying integrated GFP fusion transgene markers of different types of cellular stress such as DAF-16, SKN-1, SOD-3, GST-4, HSP-16.2, HSP-4 and HSP-6 (Supplementary Table S1) did not modify the expression levels or subcellular localization of these stress markers (data not shown).
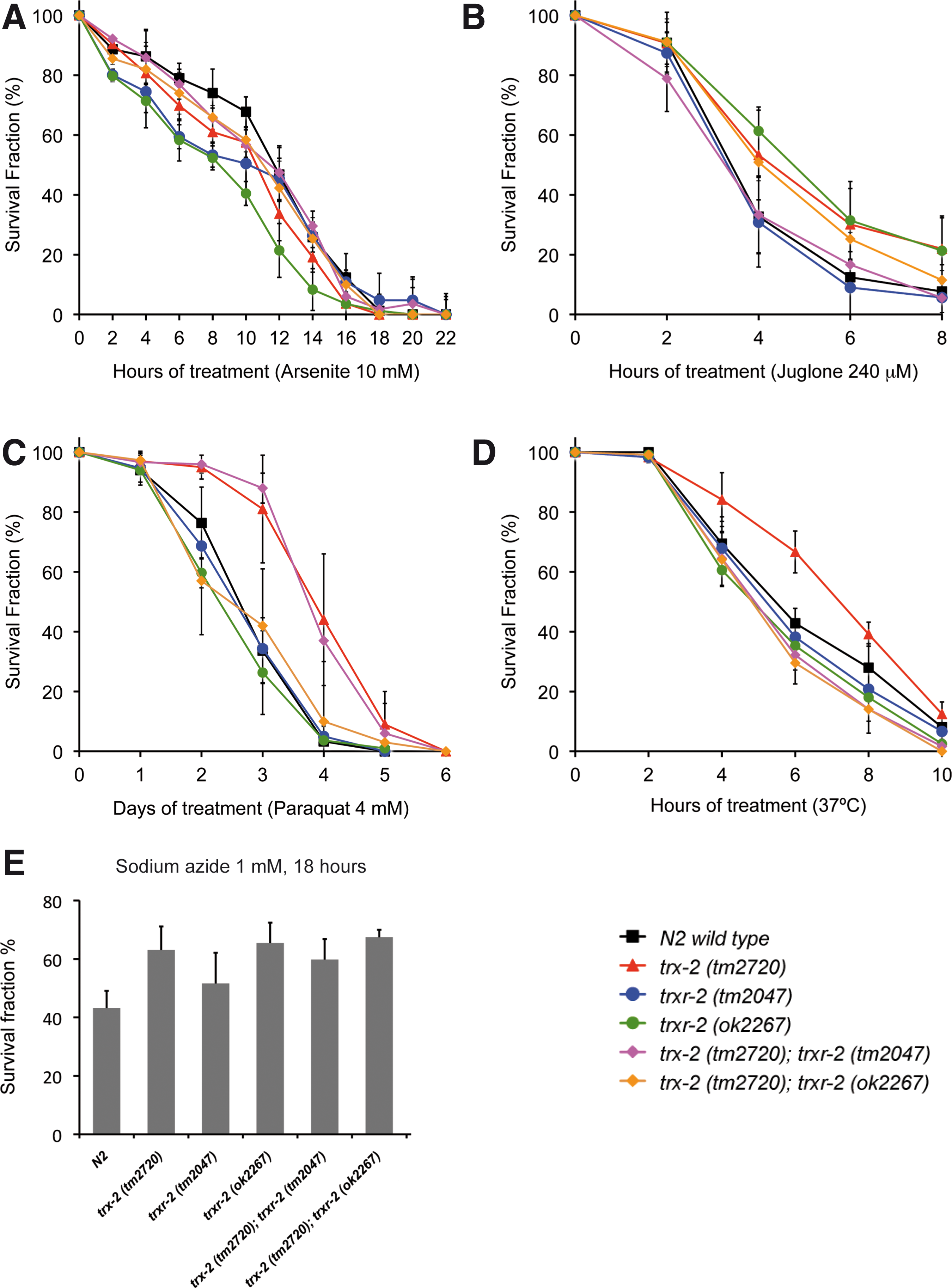
Next, we asked whether the expression levels of trx-2 and trxr-2 GFP reporters were modified under different stress conditions such as food deprivation, heat shock, sodium azide or paraquat treatments as well as induction of the mitochondrial unfolded protein response (UPRmito) resulting from spg-7 RNAi (59). Interestingly, out of all these treatments, we only found a clear induction of GFP expression in transgenic worms grown in spg-7 RNAi (Fig. 6). RNAi downregulation of other genes known to induce UPRmito such as hsp-6 and phb-2 (1, 59) confirmed that the trx-2 and trxr-2 GFP reporters were upregulated under conditions that activate UPRmito (Fig. 6). Unexpectedly, we also found some induction of trxr-2, trx-2 and hsp-6 GFP mitochondrial reporters when using ero-1 RNAi as negative control for UPRmito [ero-1 RNAi activates UPRer (59)] (Fig. 6) This result adds further support to the increasing evidence of an important crosstalk between ER and mitochondria (45). Despite trx-2 and trxr-2 genes being upregulated on UPRmito induction, trx-2 and trxr-2 mutants did not activate the UPRmito reporter HSP-6::GFP nor further enhanced the UPRmito-dependent developmental arrest phenotypes caused by spg-7, phb-2 and hsp-6 RNAi (59). Consistent with these results, overexpression of TRX-2 and TRXR-2 did not alleviate the developmental arrest phenotypes just mentioned that are caused by UPRmito induction (data not shown).
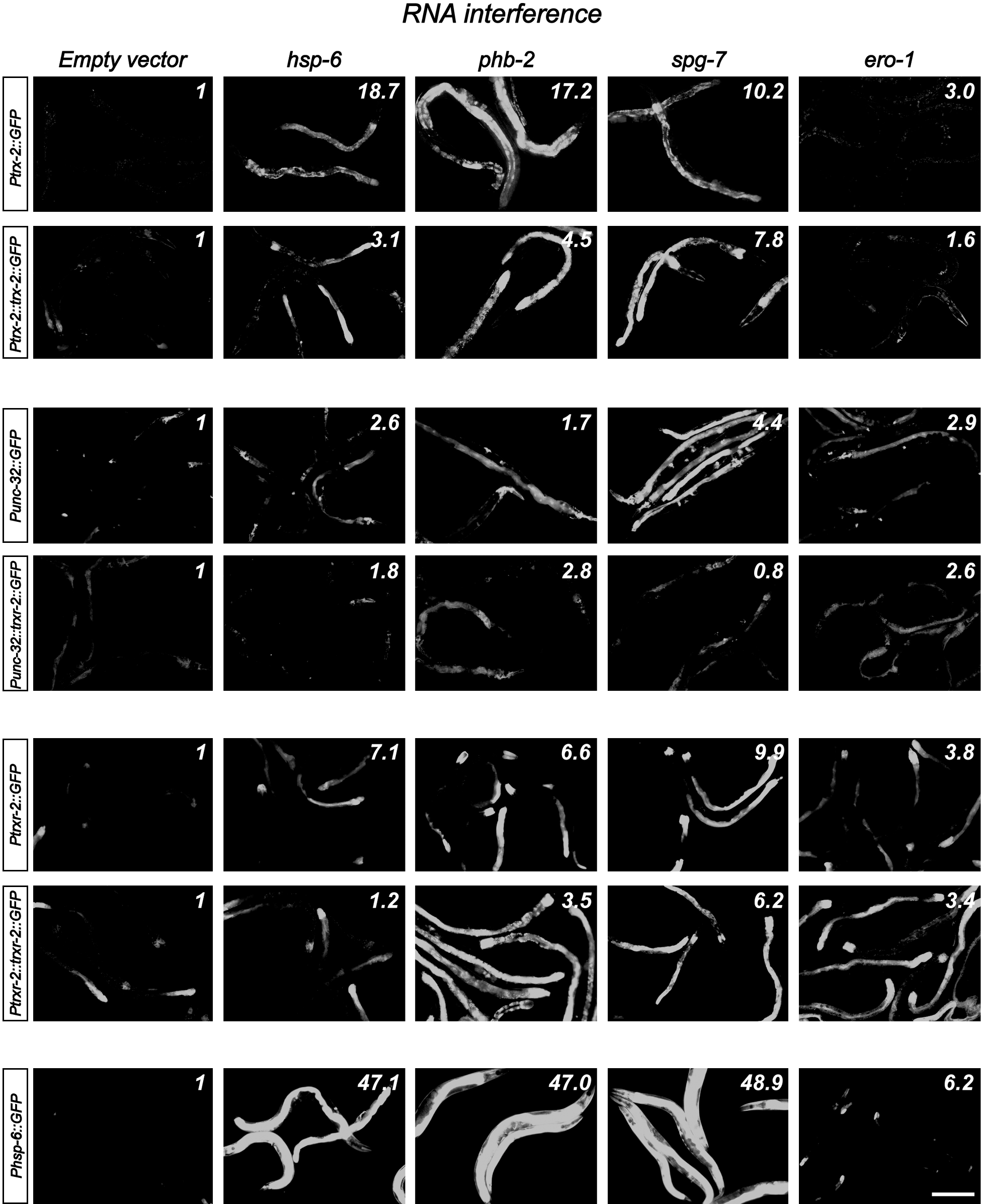
Collectively, these data indicate that the mitochondrial thioredoxin system is not required to maintain general redox homeostasis in C. elegans even under different stress conditions, but it might play a role when mitochondrial protein folding homeostasis (mitochondrial proteostasis) is compromised.
C. elegans trx-2 and trxr-2 mutants have normal longevity
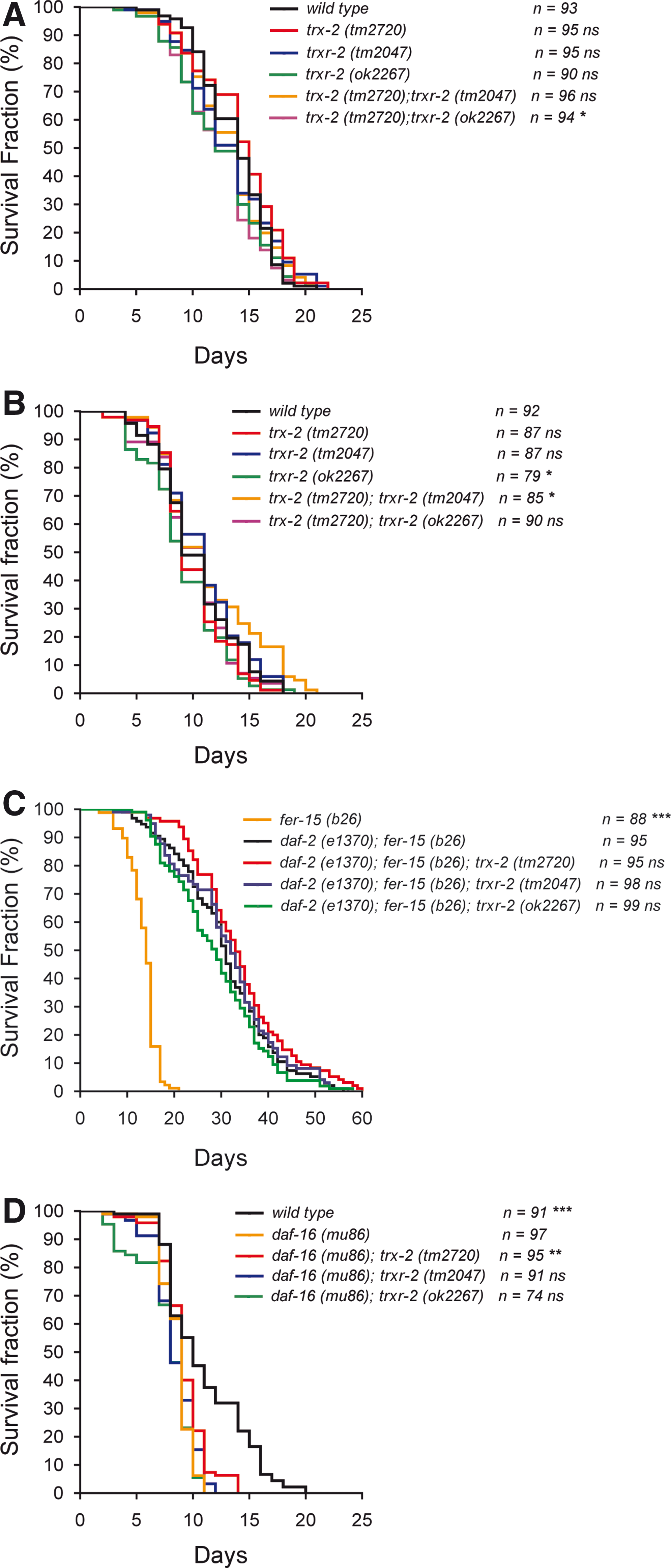
Proteostasis is critical for survival and, therefore, eukaryotic organisms integrate a complex network of chaperones, proteases and assembly factors across all cellular compartments, known as UPR, to ensure a proper balance between protein folding, assembly and degradation (3). In this context, it has been recently proposed that mitochondrial function in cellular proteostasis is critical for worm longevity (26). The finding that trx-2 and trxr-2 gene expression is induced when UPRmito is activated prompted us to investigate their role in C. elegans longevity. First, we determined C. elegans trx-2 and trxr-2 single and double mutants lifespan at both 20°C and 25°C (Fig. 7A, B) and found no major differences in mean and maximum lifespan when compared with N2 wild-type control animals. Since the insulin pathway is a major regulator of longevity (24) and because reduced insulin signaling protects against proteotoxicity (6), we next asked whether the mitochondrial thioredoxin system regulates the lifespan of mutants in the insulin pathway. For this purpose, we made double mutants combining the trx-2 and trxr-2 deletions with mutations in the genes for the insulin receptor daf-2 or the FOXO transcription factor daf-16, which are key determinants of insulin-pathway-dependent longevity (24). Again, no significant differences were observed in any mutant combination with either daf-2 or daf-16 mutants at 25°C, except for a slightly higher maximum lifespan of the trx-2 mutant in the daf-16 background (Fig. 7C, D). Overall, these data indicate that the mitochondrial thioredoxin system is not involved in regulating C. elegans longevity.
Mitochondrial TRXR-2 protects against the paralysis phenotype of a transgenic worm model of AD expressing human β-amyloid peptide
Given the increased expression of trx-2 and trxr-2 genes upon UPRmito activation, we decided to study whether these genes play a role in protein aggregation and proteotoxicity. For this purpose, we used different transgenic worms that overexpress human β-amyloid peptide in body wall muscle cells (hereafter called Aβ worms), resulting in muscular dysfunction and age-dependent progressive paralysis as a consequence of intracellular Aβ aggregation (31, 34). These Aβ worms are commonly considered a C. elegans model of AD (32).
First, we evaluated the impact of trx-2 and trxr-2 RNAi downregulation on the paralysis phenotype of the CL2006 strain, which overexpresses the Aβ peptide in a constitutive manner from the dvIs2 (Punc-54::Aβ) integrated array (31). As shown in Figure 8A, decreased levels of trxr-2 resulted in increased paralysis onset, while, in turn, trx-2 downregulation did not affect paralysis. To validate the RNAi results, we aimed at generating derivatives of dvIs2 Aβ worms in trx-2 and trxr-2 mutant backgrounds. trx-2 (tm2720); dvIs2 (Punc-54::Aβ) animals reproduced the trx-2 RNAi result and had similar paralysis onset as the dvIs2 control (data not shown). However, to our surprise, we were unable to generate animals carrying the dvIs2 integrated array in a trxr-2 mutant background. This unexpected synthetic lethal interaction could be explained by either a positional effect of the dvIs2 array insertion locus or, given the enhanced paralysis caused upon trxr-2 downregulation, by worms being unable to withstand Aβ production in the complete absence of trxr-2. To discriminate between these two alternatives, we made use of the strain CL2750 (C.D. Link, unpublished results), which, similar to CL2006, produces the Aβ peptide constitutively in muscle cells from the integrated array dvIs100 (Supplementary Table S1). Interestingly, unlike dvIs2 worms, we managed to generate Aβ dvIs100 animals in a trxr-2 (tm2047) mutant background and confirmed their increased paralysis when compared with its respective control (Fig. 8B). To rule out that variations in Aβ content might underlie the differences between the CL2006 and CL2750 strains, we examined the total Aβ levels in these two strains and found that CL2006 has a little higher amount of Aβ compared with that of CL2750 (Supplementary Fig. S5B). Based on these results, we support the notion that the failure to obtain animals expressing the dvIs2 array in a trxr-2 mutant background is due to a positional effect of the locus where the dvIs2 array is inserted (which we have independently mapped at LG II) rather than to the higher Aβ levels in CL2006 as compared with those of CL2750, although we cannot completely rule out that this slight difference might play a role.

We next asked whether the protective effect of trxr-2 on the paralysis of Aβ (constitutive) worms is a consequence of its requirement during embryogenesis and early development, when body wall muscle cells are generated (53, 54), or it can also be implemented at later developmental stages, when muscle cells are fully formed. For this purpose, we used the CL4176 strain that overproduces the Aβ peptide from the dvIs27 integrated array in an inducible manner after temperature upshift at the L3 larval stage (34). Our results show that trxr-2 also protects against the paralysis even when Aβ is produced in an inducible fashion (Supplementary Fig. S5C), indicating that the protective effect of trxr-2 in Aβ-dependent paralysis is associated to muscle function but not to muscle biogenesis.
Since the lack of trxr-2 enhances the Aβ-dependent paralysis phenotype, we wondered whether high levels of TRXR-2 would improve the paralysis onset. To this aim, we generated transgenic strains expressing high levels of TRXR-2 in both constitutive dvIs2 and inducible dvIs27 Aβ worms (Supplementary Fig. S5F). Surprisingly, while decreased levels of TRXR-2 worsen paralysis, transgenic Aβ worms expressing high levels of TRXR-2 did not improve the paralysis phenotype when compared with their corresponding nonoverexpressing control siblings (Fig. 8C and Supplementary Fig. S5D). These data indicate that restoring endogenous TRXR-2 levels is sufficient to rescue the enhanced paralysis of Aβ worms caused by dowregulation of trxr-2 and that increased amount of TRXR-2 does not provide additional protection against Aβ-dependent paralysis.
To determine whether the effect of TRXR-2 on paralysis onset was associated with differences in Aβ aggregation or oligomerization, we first performed Aβ immunoblotting on constitutive dvIs2 Aβ worms using the Aβ-peptide specific antibody 6E10. This antibody detects all forms of Aβ (amyloid deposits, amorphous Aβ aggregates and soluble Aβ oligomers and monomers) (14). As shown in Figure 8D, and except for a slight increase in Aβ oligomeric species (range 75–150 kDa), no major differences were found in Aβ worms when trxr-2 was downregulated by RNAi. Unexpectedly, we found a dramatic decrease in total Aβ amount in TRXR-2 overexpressing Aβ worms, an effect that was abolished upon trxr-2 RNAi when Aβ oligomeric species reappeared (Fig. 8D). Similar results were obtained with inducible dvIs27 Aβ worms, although the reduction of total Aβ in TRXR-2 overexpressing worms was not as strong as in the constitutive Aβ strain. (Supplementary Fig. S5E). This difference in Aβ reduction between the constitutive and inducible strains most likely reflects saturation of the (unknown) clearance mechanism as a consequence of the rapid and robust Aβ production upon temperature upshift in the inducible strain.
Immunohistochemistry with the 6E10 antibody or the fluorescent amyloid vital dye X-34 (33) on dvIs2 Aβ worms corroborated the immunoblot data, as no differences were found in either the total Aβ load or amyloid deposits formation between control and trxr-2 downregulated animals (Fig. 8E). On the other hand, dvIs2 Aβ worms overexpressing TRXR-2 showed a robust reduction in both total Aβ species and amyloid deposits (Fig. 8F).
Together, our data demonstrate an in vivo role of TRXR-2 on Aβ-dependent paralysis and aggregation in C. elegans, suggesting a possible protective function of TRXR-2 in AD.
Discussion
Given the difficulty of studying the function of the mitochondrial thioredoxin system in the context of a complete animal using the mouse or Drosophila models (due to embryonic or larval lethal phenotypes, respectively), we turned our interest to the nematode C. elegans, an excellent genetically tractable model where the mitochondrial thioredoxin system is highly homologous to that of mammals.
We first demonstrated that C. elegans trx-2 and trxr-2 genes encode proteins that are targeted to mitochondria in vivo, driven by putative MTS at their respective N-terminus. Interestingly, the rather ubiquitous expression pattern of the trxr-2 gene deeply contrasts with the highly restricted one of the trx-2 gene in AIYL/R and ASEL neurons and muscle cells, which suggests that TRXR-2 might have additional substrates other than TRX-2. We identified TRX-2 on the basis of its conserved active site sequence WCGPC and it cannot be ruled out that other thioredoxin family members with a more divergent active site sequence are also present in mitochondria. Furthermore, it has been reported that human mitochondrial glutaredoxin 2 is a substrate for mitochondrial thioredoxin reductase (22), a function that could also be conserved in worms (at least four glutaredoxin genes have been reported in C. elegans, see below). Likely, other TRXR-2 substrates not functionally or structurally related to thioredoxins and glutaredoxins might also exist in mitochondria. It is worth noting that TRX-2 is not detected in the intestine under unstressed conditions (Fig. 2A–C and J, K), but it is expressed in this organ upon UPRmito induction (Fig. 6), indicating that TRX-2 expression might be turned on in additional tissues depending on stress conditions.
C. elegans is, to date, the only metazoan in which the mitochondrial thioredoxin system is not essential for survival. More surprisingly, trx-2 and trxr-2 single and double mutants do not show enhanced sensitivity to different stressors. Collectively, this lack of phenotype supports the idea that a redundant system exists to counterbalance the absence of the mitochondrial thioredoxin system under both unstressed and stressed conditions. One alternative could be that the C. elegans cytosolic thioredoxin reductase gene trxr-1 might produce an isoform targeted to mitochondria as it happens in other organisms (48). However, we have ruled out this possibility, as trxr-1; trxr-2 double mutants are also fully viable (51). The viability of the trxr-1; trxr-2 double mutants raises the interesting question of which enzyme(s) maintain the different worm thioredoxins in their reduced active state in the absence of both thioredoxin reductases. An obvious candidate is the glutaredoxin system based on the numerous reports of functional redundancy of both thioredoxin and glutaredoxin systems (either cytoplasmic or mitochondrial) in several organisms (12, 58). We have recently uncovered an example of such functional redundancy in C. elegans as trxr-1 animals feeding on glutathione reductase (gsr-1) RNAi bacteria display a highly penetrant larval arrest phenotype due to defective cuticle ecdysis (51). However, RNAi downregulation of gsr-1 or any of the four worm glutaredoxins (glrx-5, glrx-10, glrx-21 and glrx-22) in trx-2 and trxr-2 single and double mutant backgrounds did not result in any obvious synthetic phenotype (Supplementary Table S3). Since RNAi feeding penetrance can be highly variable depending on genetic backgrounds and tissue expression, combinations of glutaredoxin system mutants with those of trx-2 and trxr-2 will be needed to unequivocally identify the redundant system for the mitochondrial thioredoxin system. Other plausible explanations for the lack of phenotype of the mitochondrial thioredoxin system mutants could be that the system is dispensable under normal laboratory growth conditions while being essential in the wild or that, alternatively, the mitochondrial thioredoxin system plays a modulatory role in nonessential ROS-mediated signaling mechanisms.
The induction of trx-2 and trxr-2 upon UPRmito activation suggests that the mitochondrial thioredoxin system is a part of the chaperone machinery required to cope with the load of unfolded/misfolded proteins that accumulate and aggregate when mitochondrial proteostasis is compromised. Most likely, the function of the mitochondrial thioredoxin system under UPRmito is aimed at reducing incorrect disulfide bonds in these misfolded proteins to facilitate either native refolding or their export from mitochondria and subsequent degradation by proteasome (56). The maintenance of cellular and subcellular proteostasis is essential for organismal survival and its imbalance dramatically affects the function of all cellular organelles, including mitochondria, eventually leading to enhanced progression of aging and development of neurodegenerative diseases such as Alzheimer's, Parkinson's or Huntington's disease (26).
In this work, we have found a protective role of the trxr-2 gene on the aging-dependent progressive paralysis phenotype caused by Aβ aggregation in worm muscle cells, a well-established model for AD (31). This protective role is not shared by the trx-2 gene despite it is also expressed in muscle cells, reinforcing the notion that additional substrates for TRXR-2 might mediate such a protective function. We have shown here that while trxr-2 downregulation clearly enhances paralysis, increased TRXR-2 expression does not alleviate the paralysis onset. On the other hand, trxr-2 downregulation does not substantially modify the total levels of Aβ species nor amyloid deposits, which, in turn, are dramatically diminished in TRXR-2 overexpressing animals. The lack of correlation between total amyloid loads and the paralysis rate has been previously reported in this AD worm model (15, 32), although no molecular mechanism has been proposed yet to clarify this fact. A plausible explanation relates to the recent finding that muscle mitochondria of Aβ worms are highly fragmented (C.D. Link, unpublished data). Our preliminary unpublished results indicate that decreased TRXR-2 levels further enhance the mitochondrial fragmentation of Aβ worms, which might account for the increased paralysis phenotype in trxr-2 mutants or trxr-2 RNAi treated Aβ worms without provoking noticeable changes in total Aβ load or amyloid deposits. Alternatively, despite the fact that higher levels of TRXR-2 strongly reduce both total Aβ load and amyloid deposits, it is possible that the residual amount of Aβ species detected in both immunoblots and immunostainings of overproducing TRXR-2 Aβ worms might still be sufficient to induce paralysis, thus explaining why overexpressing TRXR-2 does not improve paralysis onset.
Elucidating the mechanism(s) by which TRXR-2 overexpression degrades most Aβ load is of enormous interest. For instance, it is conceivable that TRXR-2 might interact with proteins implicated in the specific degradation of the Aβ peptide and amyloid deposits in mammals, such as angiotensin-converting enzyme, insulin-degrading enzyme or neprilysin (2), all of which have close orthologs in worms. Alternatively, an interesting possibility relates to the finding that reduced insulin signaling improves paralysis of Aβ worms by promoting autophagic degradation of Aβ (13). Thus, it could be possible that TRXR-2 overexpression triggers a decrease of insulin signaling, resulting in Aβ autophagic-dependent degradation. As a whole, it will be very important to gain a deeper insight into the molecular mechanisms underlying the dual role of TRXR-2 in alleviating paralysis and decreasing amyloid deposits in Aβ worms and the potential application of this knowledge to the human scenario in AD.
In conclusion, the characterization of the C. elegans mitochondrial thioredoxin system reported in this work provides a new tool to further investigate the function of this redox system in metazoan physiology and pathology. The genetic amenability of C. elegans paves the way to use the mitochondrial thioredoxin mutants in genetic screens to identify other systems involved in the maintenance of organismal redox homeostasis and, particularly, opens new possibilities to understand the role of mitochondrial redox systems in the pathology of neurodegenerative diseases.
Materials and Methods
C. elegans strains and culture conditions
The standard methods used for culturing and maintenance of C. elegans were as described previously (52). The strains used in this work are described in Supplementary Table S1. All experiments were performed at 20°C unless otherwise noted. All VZ strains were 6× backcrossed with N2 wild type.
RNA extraction and RT-PCR analysis
RNA extraction (from embryos of the different genotypes) and cDNA generation were performed by standard methods using commercial kits. Details on the nested RT-PCR procedures are found in Supplementary Materials and Methods.
Thioredoxin activity assays
The enzymatic activity of the recombinant His-CeTRX-2 and His-CeΔTRX-2 proteins was tested by their ability to reduce bovine insulin A and B chains (Sigma) using either DTT or NADPH (Sigma) and rat thioredoxin reductase-1 (IMCO) as electron donors, as previously described, with slight modifications (20). Details on recombinant protein purification and enzymatic assays are found in Supplementary Materials and Methods.
GFP expression constructs and transgenesis
All transcriptional and translational GFP fusion constructs were generated using the pPD95.77 vector backbone. Information on primer sequences, cloning sites and the size of the respective inserts will be provided upon request. Details on transgenic strains generation are found in Supplementary Materials and Methods.
Expression pattern analysis
GFP transgenic animals were mounted in a 5 μL drop of 10 mM levamisole (Sigma) on a 3% agarose pad covered with a 24×24 mm coverslip. DIC and fluorescence imaging was performed on a Zeiss AxioImager M2 ApoTome fluorescence microscope equipped with an AxioCam MRn (Zeiss). Images were captured with the AxioVision 4.8 Software (Zeiss) and equal adjustment of brightness and contrast on control and matched experimental images was done using Adobe Photoshop 10 Software (Adobe Systems).
Analysis of embryonic and germ line apoptosis
Apoptotic corpses were detected by their refractile shape using a Leica DM6000 microscope equipped with DIC optics and scored as described (47). For embryonic apoptosis, three embryos of each genotype were mounted at a very early stage (2–4 cell stage) and the recording of their development was carried out at 25°C. The 4-D microscopic analysis was carried out by recording 30 focal planes of the embryos for 10 h. As a result, four-dimensional (4D) movies (3D of the embryo+time) were obtained for each genotype. The SIMI Biocell software (SIMI GmbH) allowed tracing each and every cell of the embryo in time and space as described (39). Time of birth and onset of apoptosis were measured to define the kinetics of cell death. Time to die is defined as the time period from which a cell is born until it begins to show morphological characteristics of apoptosis, such as rounding of the cell, cytoplasmic condensation or nuclear swelling (23). For germ line apoptosis, 20 gonad arms per genotype were analyzed and the apoptotic corpses detection and scoring were performed as described above.
Stress assays
For paraquat, sodium arsenite, sodium azide (Sigma) and heat-shock treatments, L4 larval stage hermaphrodites were used to initiate the assays. For juglone treatment, young adult hermaphrodites were utilized. The viability of each animal was assessed at the indicated times by microscopic examination of pharyngeal pumping and movement in response to prodding with a pick. Dead worms were counted and removed from the plates. Details are found in Supplementary Materials and Methods. The percentage of surviving animals for each treatment was calculated from three or more independent experiments.
RNAi and UPRmito induction quantification
Feeding RNAi was performed as previously described (57). Details are found in Supplementary Materials and Methods. Fluorescence quantification of UPRmito induction was done on images (acquired under identical settings) of 5–10 worms for each RNAi condition, using the ImageJ Software (National Institutes of Health).
Longevity assays
Life-span assays were performed at 20°C or 25°C as previously described (30) with slight modifications. Details are found in Supplementary Materials and Methods.
Paralysis phenotype, Aβ peptide expression and β-amyloid deposits quantification
For strains CL2006, CL2750 and derivatives, synchronous populations of Aβ worms were generated by time-limited (2 h) egg lay at 16°C. Parents were removed and progeny was grown continuously at 20°C. Paralysis scoring was initiated at the first day of adulthood and determined daily, whereby paralyzed worms were removed from plates. For strains CL4176 and VZ296, synchronous populations of Aβ worms were generated by time-limited (2 h) egg lay at 16°C. Parents were removed and progeny was grown continuously at 16°C for 48 hours followed by a temperature upshift to 23°C or 25°C to induce Aβ production. Paralysis scoring was initiated 24 h after a temperature upshift and determined every hour, whereby paralyzed worms were removed from plates. Aβ immunoblotting and immunostaining was carried out using the 6E10 monoclonal antibody (Covance Inc.). The quantification of β-amyloid blots was performed using the ImageJ Software, while the quantification of β-amyloid deposits using the fluorescent amyloid dye X-34 (a kind gift from Prof. William Klunk) was performed as described, with brief modifications (15, 33). Details are found in Supplementary Materials and Methods.
Footnotes
Acknowledgments
We thank the Caenorhabditis Genetics Center, the Japanese National Bioresource Project, Ellen A. Nollen, Piali Sengupta, Amir Sapir, Paul Sternberg, Simon Tuck, Oliver Hobert, Alexander van der Bliek, Andrew Fire, Cesar Santiago and Julian Cerón for sharing worm strains and plasmids. We are grateful to Pilar Alarcón, María Jesús Rodríguez and Francisco José Naranjo for excellent technical assistance, Lizett Rodríguez for help with enzymatic assays and Fernando Calahorro for his expert assistance with sequence analysis. We also thank Elias Arnér for critical reading of the article. Continuous support from Peter Askjaer's and Manuel Muñoz's groups is deeply acknowledged. A.M.-V. was supported by the Instituto de Salud Carlos III (Projects PI050065 and PI080557, co-financed by the Fondo Social Europeo, FEDER) and Junta de Andalucía (Projects P07-CVI-02697 and P08-CVI-03629), Spain. B.C.-V. was supported by a fellowship from the Consejo Nacional de Ciencia y Tecnología (CONACYT) from the Government of Mexico. P.N. was supported by the Instituto de Salud Carlos III (project PI080500, co-financed by the Fondo Social Europeo, FEDER) and Junta de Andalucía (project P08-CTS-03988). Work in the laboratory of P.S., a member of the NordForsk Nordic C. elegans network, was supported by a grant from the Swedish Research Council. J.C. was supported by the Spanish Ministry of Science and Innovation Grant BFU2010-21794 and the RiojaSalud Foundation.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.