
Abstract
Introduction
ROS provoke oxidative damage of biomolecules, including nucleic acids, proteins, and lipids. For instance, •OH efficiently reacts with the four nucleobases, whereas 1O2 is only able to oxidize guanine bases (15, 16). During the formation of 8-hydroxyguanosine (8-OHG) by •OH, but not 1O2, a guanine radical cation (G•+, so-called electron hole) is formed with a subsequent release of an electron and proton (14, 16). Moreover, the presence of 8-OHG is considered to be a biomarker of oxidative stress (12, 13, 16, 37). It is noteworthy that G•+ can migrate long distances (hundred of Å) along DNA by a reversible hopping process before being irreversibly trapped by a reaction with H2O and O2 (14, 34). Additionally, guanine is the main target of oxidation in RNA and DNA, especially at multiple guanine sequences, that is, GG or GGG. Among nucleobases it has the lowest ionization potential that causes the migrating G•+ to briefly pause at guanine multiplets, facilitating a trapping reaction (26, 34). The formation as well as hoping and trapping of G•+ can be considered to be a kind of signal-sensing and transfer mechanism, and hence as a DNA-mediated signaling process. Over recent years researchers have indeed reported the existence of such signaling mechanisms. These results will be presented and discussed in the frame of this review.
Many oxidative base lesions in DNA are mutagenic. These can provoke structural alterations (including transversions: G→T or A→C) or overall conformational changes, which might have an effect on transcription and/or replication; ultimately, this can lead to chromosome deletions with lethal effects (18, 32). To counteract these deleterious effects organisms have evolved enzyme-based repair systems that comprise base excision repair (BER) and nucleotide excision repair (NER). During BER, a single lesion is removed via a glycosylase, while during NER a lesion-containing oligonucleotide is removed. Both DNA repair pathways are initiated by damage recognition followed by incision/excision steps, gap filling accompanied by ligation (53, 63). While these enzymatic-based processes may be rapid, they still require a finite period of time between the generation of the mutation and competition of the repair. Thus, the transfer of G•+ may provide a supporting mechanism to localize and mitigate the effects of oxidative attack to guanosine nucleotides. Taken together, these processes provide the cell with an efficient and variable DNA damage response mechanism under conditions of oxidative stress.
The main consequence of oxidative modification of messenger RNA (mRNA) is reduced protein production. Studies with human RNAs led to the proposal that RNA is more vulnerable to oxidative damage than DNA and that RNA oxidation is not random but highly selective (55). Further studies have shown that ribosomal RNAs (rRNAs) can be also oxidized via a bound redox-active metal ion and that the higher order structure of rRNA may contribute to its greater metal ion-binding ability (31). To our knowledge, RNA oxidation processes in bacteria have not yet been reported. Nonetheless, it is likely that such processes exist, in particular those in which regulatory RNA elements play a role (30, 67). Interestingly, a bacterial RNA element—designated to be a metal-sensing riboswitch—was characterized in vivo and in vitro and shown to be involved in the regulation of gene expression in a metal-dependent fashion (21).
Proteins with coordinated metal ions are also modified during oxidative stress. Metal ion (e.g., iron)-mediated generation of hydroxyl radicals occurs mostly at metal centers of proteins as •OH can travel only 4 nm within its short half-life (∼10−9 s), often inducing site-specific damage (38). With a redox potential ranging from −300 to +700 mV, iron is a versatile prosthetic component that can be incorporated into proteins either as a mono- or as a binuclear species, or in a more complex form as part of iron–sulfur clusters or heme groups (2). These redox properties provide iron with high sensitivity, and the ability to sense the redox status of the cell. This in turn provokes an appropriate cellular response. Thus, iron-based production of •OH can also be considered to be beneficial for cells, as these radicals function as redox-signaling molecules that lead to a coordinated response under oxidative-stress conditions.
Proteins exposed to •OH undergo a variety of oxidative modifications, including nitration of aromatic amino acid residues, hydroxylation of aromatic groups and aliphatic amino acid side-chains, sulfoxidation of methionine residues, and conversion of some amino acid residues to carbonyl derivatives (57). For example, oxidation of tyrosines leads to the formation of dityrosine cross-linking products, causing irreversible oxidative damage and often leading to a loss of function due to aggregation and degradation. Indeed, the presence of dityrosines has previously been used as a marker for whole-body oxidative stress (20). Cysteines are additional targets for protein oxidation, as they react easily with H2O2 and other free radicals. Reversible oxidation of cysteines are highly complex chemical reactions and can lead to multiple sulfur oxidation states, including thiols, sulfenic and sulfinic acids, thiyl radicals, and disulfide S-oxides (33).
In common with all living organisms, bacteria have evolved nonenzymatic and enzymatic pathways to prevent damage to essential macromolecules during redox-stress events (3). For example, single transcription regulators (e.g., OxyR or FurS) use cysteine thiol groups for signal sensing. Upon signal sensing a conformational change occurs within the regulator, leading to activation or de-repression of transcription of genes involved in antioxidative stress response (17, 49). SoxR is an example of a redox-sensitive transcriptional regulator that utilizes iron–sulfur [2Fe-2S] clusters. Upon oxidative stress, the cluster is oxidized, resulting in the SoxR-dependent transcriptional activation of soxS. This encodes for a further transcriptional activator, SoxS, which regulates the expression of genes involved in antioxidant production or oxidative damage repair (51).
The majority of protein-based sensory systems in bacteria are the so-called two-component systems (TCSs) consisting of a sensor kinase (SK) and a response regulator (RR) that controls cellular response by a change in gene expression. One of the best studied SKs involved in redox sensing is FixL from Rhizobium sp. FixL possesses a Per-Arnt-Sim domain that contains a heme prosthetic group. Reversible binding of oxygen to the heme group regulates its kinase activity, and thereby the phosphorylation state of its cognate RR FixJ (27). Allosteric changes of FixL upon oxygen binding lead to deactivation of kinase activity (52). Oxygen binding may induce a flattening of the heme group, promoting the formation of a hydrogen bond to the liganded oxygen (8).
In the first part of the presented review, we describe novel studies performed on nucleic acid-mediated processes involved in redox signaling. Under these conditions specific DNA or RNA modules can be considered to be a type of signaling module. Our main aim in this first section is to provide the readers with current knowledge on novel signaling paradigms and avenues with promising research perspectives. In the second part, we focus further on a novel protein-based redox-sensing system, namely, the three component system HbpS-SenS-SenR, in which the accessory protein module HbpS directly switches on/off the corresponding signal cascade.
Nucleic Acids and Redox-Signaling
DNA-mediated electron transfer
Although DNA is not primarily an electron-transfer module, the ordered π-electron system of common DNA bases in duplex B-form DNA provides a pathway for the transfer of excess charges once generated on extended and well-defined stacks of base pairs (7, 23, 58). This concept of charge mobility is an important consideration with respect to environmentally induced DNA damage. Such excess charges can be generated in defined DNA sequences by exposure to free radicals or radiation (either ionizing [γ] or nonionizing [UV]) and a number of different studies [reviewed by refs. (25, 26)] demonstrated that long-range charge transfer occurs. Experiments have been performed showing that guanosine (G) undergoes oxidative modifications (to 8-oxoG) upon oxidative stress (12, 56). Studies have also shown that oxidation of guanosines results in a guanine radical cation (G•+), which induces an electron migration (hole transport) from a distant guanine through the base pairs of DNA (Fig. 1). Holes (electron vacancies) induced by oxidants are transferred across DNA ∼50 Å by hopping between CG base pairs and can oxidize remote G-sites, particularly sites comprising sequences of multiple GC base pairs (26). Such a mechanism would imply that upon oxidative attack of an essential DNA element, hole transport would result in oxidative damage to a less essential element. In some bacteria several genes possess G:C-rich sequences outside of the encoding area (19). These G:C-rich domains would act as sinks for the damage, and function as a driving force for hole transport. As a consequence, mutagenic damage would be concentrated in noncoding regions of the genome (Fig. 1).
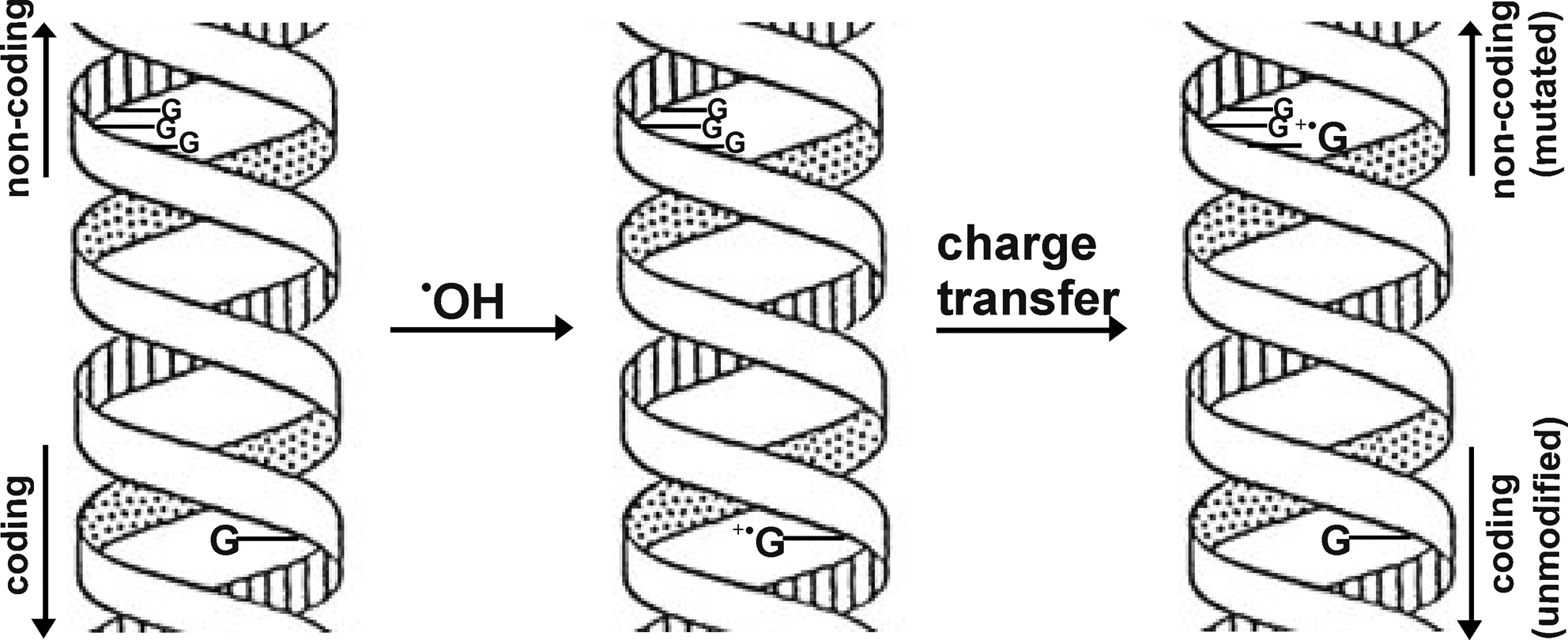
DNA-mediated redox-signaling
A common feature of protein-based redox-signaling is the modulation of the transcriptional activity of the effecter DNA-binding protein via a redox sensory protein module. In the last few years, a novel type of signaling, namely, DNA-mediated redox-signaling pathway, has been identified (4, 25). Barton and co-workers elucidated a mechanism in which DNA functions as redox sensor and mediates the oxidation of a regulatory protein, and hence the cellular response to oxidative stress. In vivo and in vitro analyses showed that ROS can rapidly migrate to regions in the chromosome that are rich in guanine multiplets (GG or GGG). Such regions are abundant near the SoxR operator within the soxS promoter. According to the above model, guanine radicals, arising from guanine multiplet regions, lead to oxidation of the SoxR protein, which subsequently causes the activation of SoxR-mediated transcription of soxS (Fig. 2). The mechanism of DNA charge transfer provides this regulator with a long-distance redox-sensing mechanism (up to 27 nm).

RNA-based processes in redox-signaling
The main consequence of oxidative modification of mRNA is reduced protein synthesis, due to the oxidative degradation of mRNA (55). Recently, the discovery of RNA elements within mRNAs, riboswitches, led us to hypothesize a role for RNA in the sensing of redox events. In general, riboswitches directly sense and bind cellular metabolites. This binding induces a conformational change within the RNA element or directly participates in a chemical reaction that modulates riboswitch activity. These events affect transcriptional initiation and termination, as well as translation (30, 67). Interestingly, these metabolites include the redox-active compound riboflavin (from which both flavin adenine dinucleotide and flavin mononucleotide [FMN] are derived). The FMN-mediated riboswitch is exclusively located upstream of genes involved in riboflavin biosynthesis and transport (66). FMN can accept electrons and exists in three different states, all possessing different chemical properties (oxidized, singly reduced, and doubly reduced) (40). Recently, structural analyses of FMN riboswitches (with and without bound FMN) from Fusobacterium nucleatum have shown that FMN-free RNA element may adopt a bound-like structure, suggesting that structural rearrangements occurring upon binding are restricted to a few of the loop regions that form the binding pocket of the riboswitch (54, 64). Studies on the FMN riboswitch from Bacillus subtilis demonstrated that there are different parameters, that is, kinetic of RNA polymerase, concentration of FMN, and thermodynamic equilibrium that together govern the highly complex mode of action of this riboswitch in vivo (65). However, these studies do not provide direct evidence for the redox state of FMN. We believe that under certain oxidative-stress conditions FMN may adopt different redox states, which would result in further conformational changes of the riboswitch, fine-tuning regulatory response.
Interestingly, in vivo and in vitro studies allowed the identification and characterization of a metal-sensing riboswitch in B. subtilis that interacts with Mg2+ and is involved in Mg2+ homeostasis (21). These studies revealed that Mg2+ can induce a compacted tertiary architecture for the so-called M-box RNAs that ultimately facilitate the accessibility of nucleotides involved in genetic control. Dann et al. suggest that additional RNA-based metal sensors exist and that these may have a high relevance during metal ion-mediated signaling (21). In this context it is noteworthy to mention that a human rRNA has been shown to be oxidized via a bound redox-active metal ion and that the higher order structure of rRNA may contribute to its greater metal ion-binding ability (31). Therefore, it will be challenging to search within bacterial genomes for riboswitches that are specific for redox-active metal ions, that is, iron or copper, and to investigate the genetic response in these organisms upon exposure to oxidative stress. We assume in general that redox-active ligands (i.e., ions or metabolites) from riboswitches may sense the redox state of the cell environment and subsequently undergo conformational or electrostatic changes that ultimately result in changes of the riboswitch structure, eliciting the appropriate response in terms of inhibition or activation of gene expression (Fig. 3).
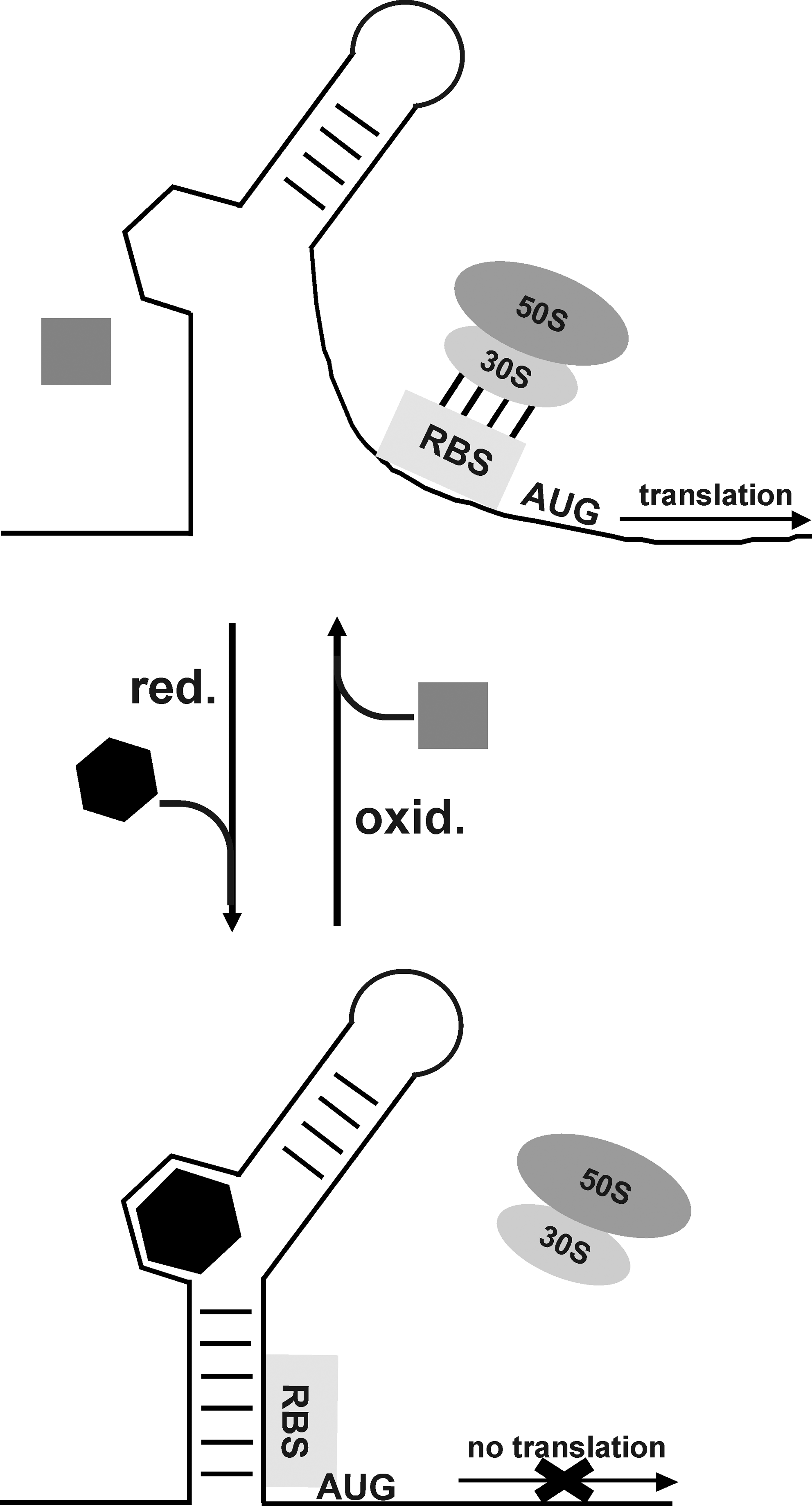
Over the last decades researches were able to in vitro synthesize small RNA molecules, called RNA aptamers, via the SELEX (systematic evolution of ligands by exponential enrichment) procedure (62). These aptamers show high specificity for macromolecules (i.e., proteins, metabolites, or redox cofactors). For instance, Burgstaller et al. obtained an RNA aptamer that displayed high affinity to FMN (11). Binding studies revealed that the aptamer exclusively recognizes the flavin portion of the cofactor with a Kd of ∼500 nM. Nevertheless, a detailed analysis of redox-dependent modulation of these aptamers has yet to be performed. Interestingly, Lauhon and Szostak synthesized an anti-NMN+ (nicotinamide mononucleotide) aptamer that was shown to be able to discriminate between NMN+ and NMNH in solution by ∼15-fold. Their studies additionally show that there are specific parts in the aptamer that are responsible for the binding activity and stability of the aptamer–ligand interaction (39).
Despite the fact that naturally occurring redox riboswitches and in vitro synthetized aptamer-redox cofactor exist (61), an open question remains: are these cofactors used as a prosthetic group for RNA-mediated redox signaling?
RNA-binding redox sensor proteins
Most systems identified to date rely on DNA-binding proteins to control differential gene expression upon redox stress. However, increasing information is available on proteins that interact with RNA to regulate cellular response on a translational level, thereby exerting additional levels of redox regulation. Careful regulation of mRNA half-lives is a fundamental mechanism that allows cells to quickly respond to changing environmental conditions. Examples are the iron responsive proteins (IRP1 and IRP2), which function under low iron conditions as RNA-binding proteins to modulate the translation of mRNAs. These proteins are regulated by iron-mediated oxidative stress in which the reactivity of H2O2 with iron directly connects oxidative stress and intracellular iron metabolism (50). In IRP1 there is a further link between the presence of iron and metabolism. Under high iron conditions IRP1 contains an intact 4Fe-4S cluster and functions as aconitase within the citric acid cycle. Under low iron conditions the protein incorporates a 3Fe-4S cluster and functions as an RNA-binding translational regulator. Iron responsive elements (IREs)-like sequences are present at the untranslated regions of open reading frames that are involved in either iron or oxygen metabolism of the cell (6, 29). IRP2 is also redox regulated under low iron conditions. Two cysteine residues of the RNA-binding cleft form a disulfide bridge and close off the terminal-loop binding pocket preventing insertion of the IRE terminal loop (69). However, in contrast to IRP1 it does not contain an iron–sulfur cluster under high iron conditions but is polyubiquitylated before proteosomal degradation (41).
Accessory Proteins-Mediated Redox Signaling
TCSs and accessory proteins
Sensing of environmental signals in bacteria is often coupled to TCSs (48). Recently, the existence of accessory proteins modulating the activity of certain TCSs has been reported (42, 48, 59). For instance, the activity of the TCS YycGF from B. subtilis seems to be modulated by two proteins, YycH and YycI, which are tethered to the periplasmic side of the membrane via a single N-terminal helix. Theoretical calculations combined with mutagenesis studies indicate that the SK YycG forms a complex with YycH and YycI. It is proposed that subtle alterations in the structure of the transmembrane complex influence YycG kinase activity (59). Because the predicted interface between YycH and YycI is distinct from their interface with YycG the implication of the proposed model is that the two proteins tether the cellular YycG pool in a large complex (59). However, the exact mechanism of interaction and signaling between YycH and YycI with YcgGF remains unclear. Szurmant and co-workers hypothesize that YycH and YycI are involved in sensing the redox state of the electron transport over the membrane and link this to the YycGF system (60).
The HbpS-SenS-SenR system: HbpS as an accessory module
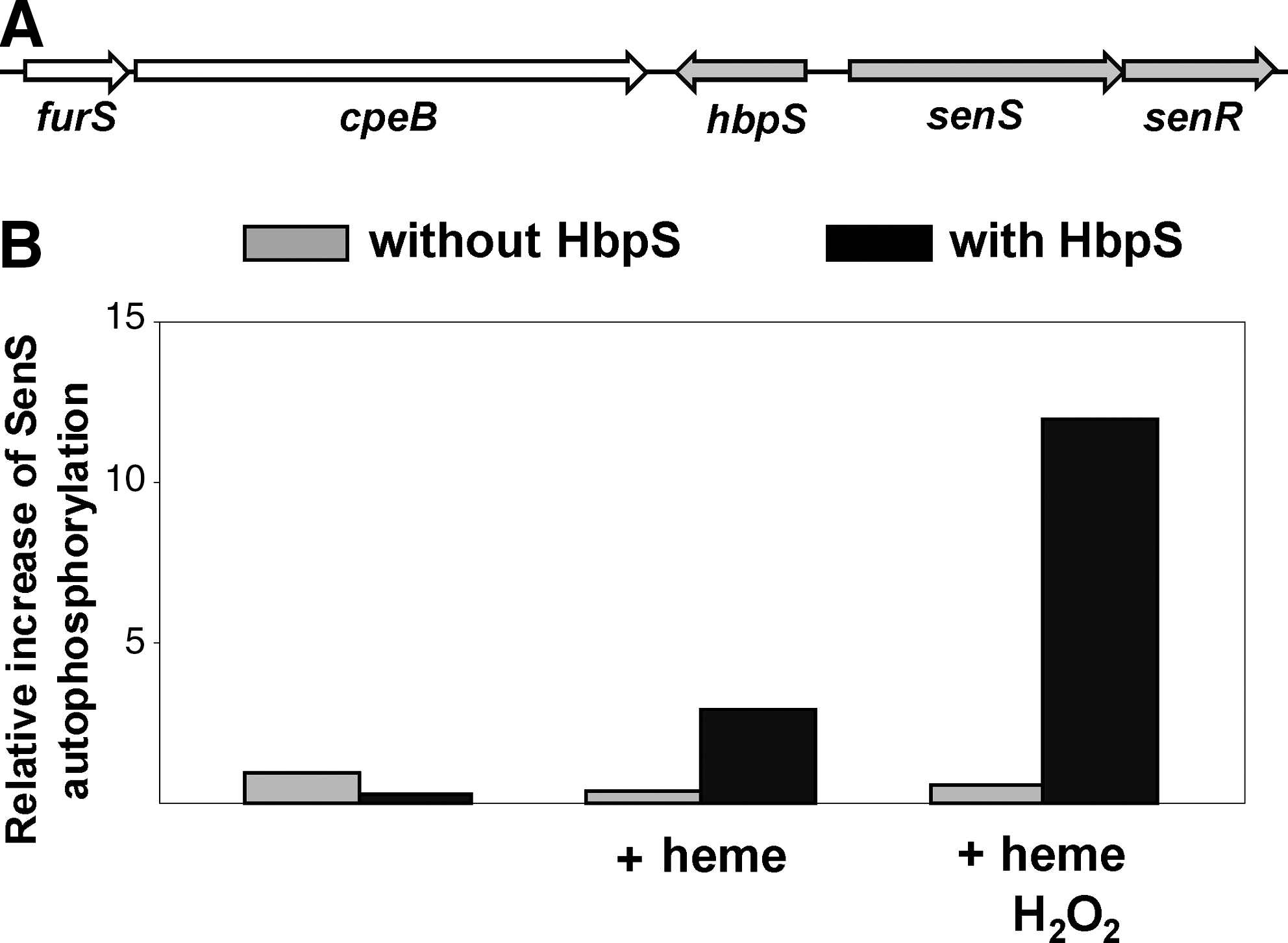
The soil bacterium and cellulose degrader Streptomyces reticuli possesses a TCS composed of SenS-SenR (10, 47). SenS is an SK that transfers the phosphate group to its cognate RR, SenR. Dependent upon its phosphorylation state the RR SenR regulates the transcription of the senS-senR operon, the gene hbpS encoding for a heme-binding protein (HbpS), the furS-cpeB operon encoding for the redox regulator (FurS), and the mycelia-associated catalase-peroxidase CpeB (9, 44, 68). Physiological studies have demonstrated that CpeB protects S. reticuli against redox-cycling compounds (35), indicating that CpeB plays an important role in minimizing reactions caused by ROS. The production of two additional proteins that have features of redox-active proteins (xylanase-like and tyrosinase-like) has also been shown to be under the control of SenS-SenR (10). Thus, we have proposed SenS-SenR to be a redox-sensing system involved in antioxidative stress response. Indeed, sensitivity and viability assays showed that an S. reticuli senS-senR disruption mutant is more sensitive to redox-active compounds, including iron ions, heme, H2O2, or plumbagin (47).
We have characterized the hbpS gene that is located between the operons furS-cpeB and senS-senR (Fig. 4A). Immunological studies combined with mutagenesis have demonstrated that HbpS is located extracellularly (46). Interestingly, comparative physiological studies revealed that the presence of HbpS increases the synthesis of the catalase-peroxidase CpeB and provides S. reticuli with resistance against redox-cycling compounds. Therefore, we have proposed that HbpS interacts with extracellular, membrane-associated, or membrane-embedded proteins involved in a signal transduction cascade, regulating cpeB transcription and resistance to redox stress. As the membrane-embedded SK SenS together with the RR SenR regulates cpeB transcription, an interaction between HbpS and SenS has been postulated. Further in vivo and in vitro analyses showed that in fact HbpS specifically interacts with SenS, and subsequently modulates its autophosphorylation: while HbpS alone inhibits the autophosphorylation of SenS, in the additional presence of iron ions or heme and redox stress considerably enhances SenS autokinase activity (Fig. 4B) (10, 44).
Domain and sequence alignments of HbpS yielded a number of putative homologs encoded within ecologically relevant bacteria [i.e., Streptomyces coelicolor A3(2), Streptomyces kasugaensis, Arthrobacter aurescens, Pseudomonas fluorescens, Pseudomonas putida, and Sphingomonas aromaticivorans] and medically relevant bacteria (i.e., Vibrio cholera, Yersinia enterolitica, Leifsonia xyli, Acinetobacter baumani, Photorabdus luminiscens, and Bordetella avium). The majority of the corresponding hbpS-like genes are clustered with genes encoding for SKs and RRs (48). Interestingly, the genes of the accessory proteins YycH and YycI as well as those of the TCS YycGF are also clustered within their corresponding bacterial genomes (60).
HbpS binds and degrades heme
Domain analysis using the Pfam Protein Families Database revealed that HbpS displays homologies with small stretches within the N-terminal regions of heme-binding proteins belonging to the NapC/NirT cytochrome c-family, suggesting that HbpS might interact with heme. The existence of an HbpS-heme complex was colorimetrically demonstrated within the extracellular proteins from S. reticuli. In addition, a typical heme spectrum (Soret peak between 409 and 412 nm) was recorded using purified HbpS proteins (44, 46). As HbpS does not possess any known heme-binding signature (e.g., C-X-X-C-H), it was suggested to be a novel heme-binding protein.
Electron density analysis from the three-dimensional structure of HbpS elucidated from crystals growing in the presence of heme revealed the presence of bound iron ions at the protein surface, potentially as a result of heme degradation. Subsequent spectroscopic and biochemical studies confirmed that HbpS degrades heme through a nonenzymatic and H2O2-dependent mechanism known as coupled oxidation (44).
HbpS has an octomeric assembly
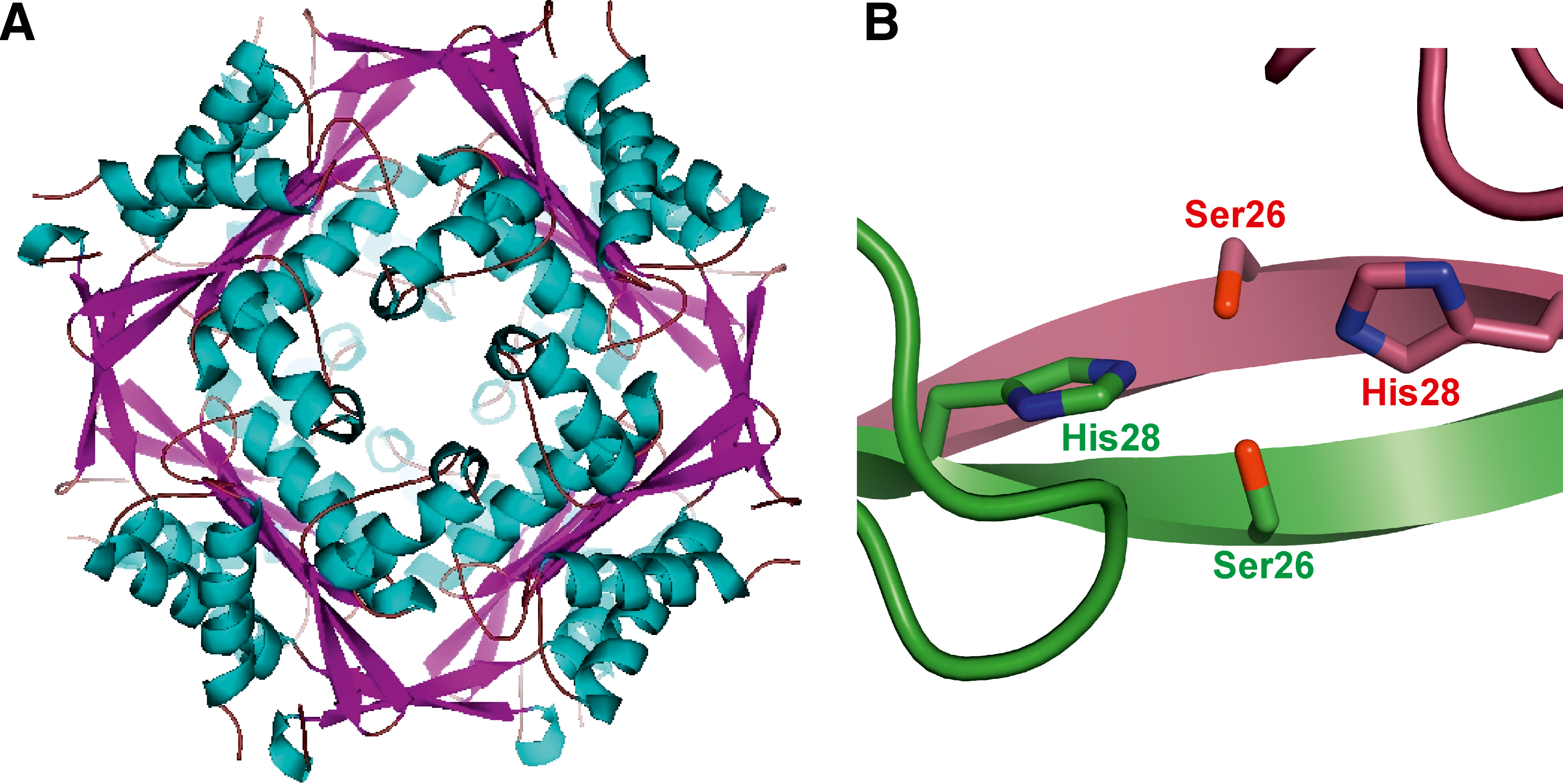
Size exclusion chromatography combined with static light scattering analyses allowed the identification of HbpS as an octamer (44). Moreover, HbpS was successfully crystallized either in the presence or in the absence of heme. Both crystal structures (1.6 and 2.3 Å, respectively) shared this octomeric assembly (Fig. 5A). The extensive interactions between N-terminal residues within the HbpS octamer are suggested to control the oligomerization process. A detailed analysis allowed the identification of two N-terminal residues (Ser26 and His28) as essential for octomeric assembly. Ser26 from one subunit interacts via hydrogen bonds with His28 from an immediately adjacent subunit, and vice versa (Fig. 5B). The octomeric assembly was additionally demonstrated in solution using site-directed spin labeling combined with pulse electron paramagnetic resonance (SDSL EPR) spectroscopy (35). Further protein–protein interaction studies combined with phosphorylation assays revealed that the octomeric assembly in HbpS is required for efficient interaction with SenS, and the modulation of SenS autophosphorylation (10, 48).
Molecular switches governing the HbpS-mediated cascade
HbpS inhibits SenS autophosphorylation under nonstressing conditions. However, in the presence of heme or iron ions and redox-stress this activity is highly enhanced. Using fluorescence resonance energy transfer (FRET) a decrease of the distance between Trp90 and Ser139 (from 21 to 16.2 Å) under iron-based oxidative-stressing conditions was recorded (45), indicating that HbpS undergoes a conformational change upon exposure to oxidative stress.
The FRET data are in line with those obtained by SDSL EPR, which also show an iron-based oxidative stress-dependent distance decrease between Ser139 and Trp90 within the HbpS octamer (35). Moreover, analyses of secondary structures by circular dichroism spectroscopy in the presence of increasing levels of iron-mediated oxidative stress showed a reduction in α-helical structures (from 42% to 20%) combined with an increase of β-sheets (from 15% to 25%) after addition of higher concentrations of iron ions (45).
Biochemical analyses revealed that iron-catalyzed oxidative stress led to the formation of sodium dodecyl sulfate (SDS)-resistant cross-linked forms of HbpS. The formation of intra- or inter-protein cross-linked derivatives can be induced by several different mechanisms: formation of dityrosines, direct interaction of two carbon-centered radicals, or interactions of the carbonyl groups of oxidized proteins with the primary amino groups of lysine residues in the same or a different protein (57). Carbonyl groups have been identified in iron-stressed HbpS proteins using specific antibodies. Furthermore, the formation of dityrosines in HbpS has also been demonstrated by fluorescence spectroscopy. In combination with mutational analyses it was shown that Tyr77-Tyr77 bridges between neighboring subunits within the octamer are formed. Additional in vivo analyses showed that upon iron-mediated oxidative stress the levels of cpeB as well as of hbpS transcripts are highly elevated in an HbpS-SenS-SenR-dependent manner. Thus, oxidative modifications accompanied by structural and conformational changes in HbpS are responsible for the up-regulation of the activity of the HbpS-SenS-SenR system (Fig. 6).
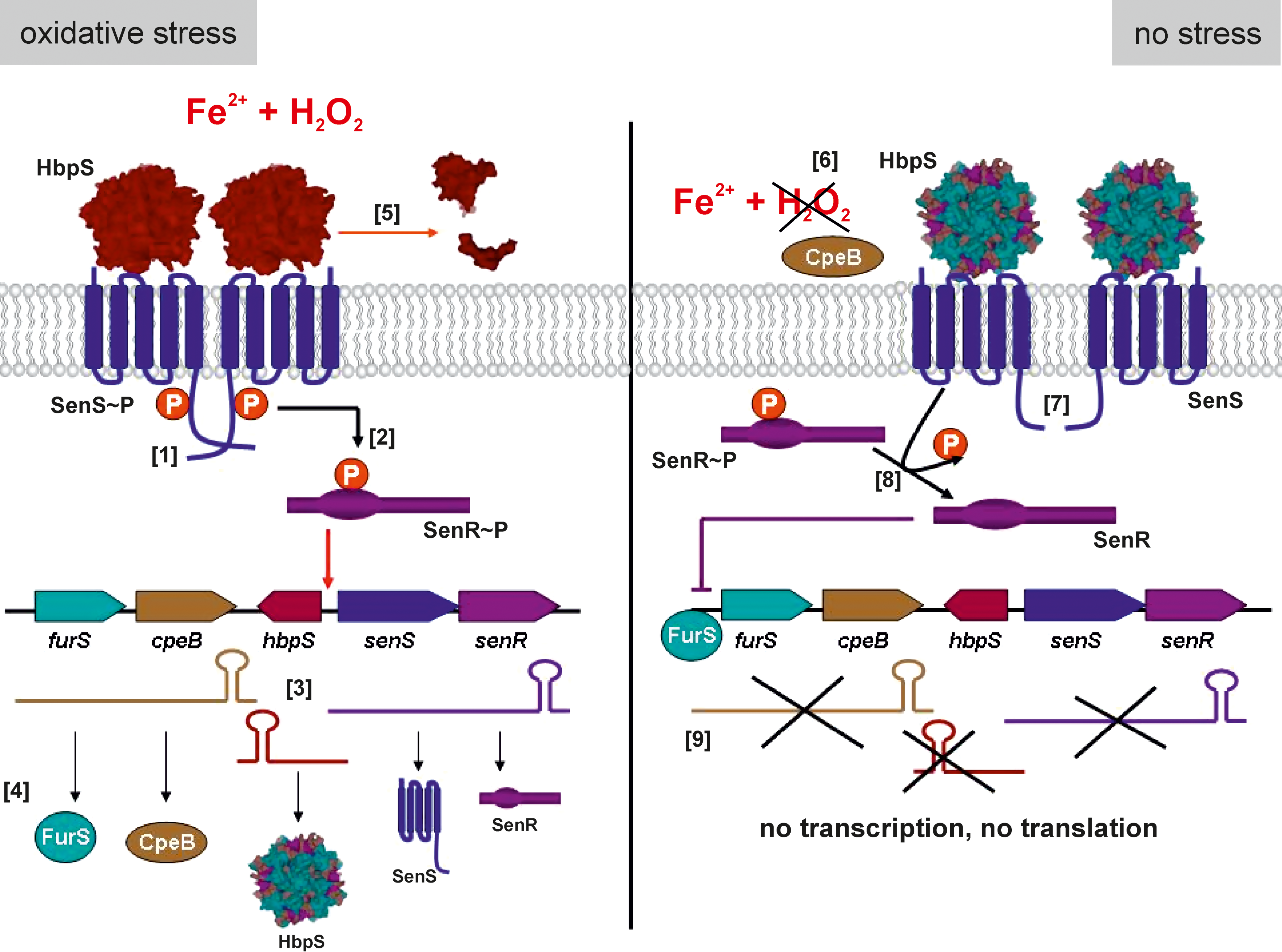
In vitro and in vivo studies demonstrated that oxidatively modified HbpS is degraded, either autonomously or enzymatically through action of naturally occurring extracellular Streptomyces proteases (35, 45). Recently obtained results showed that the mycelia-associated catalase-peroxidase CpeB protects HbpS in vivo and in vitro from iron-mediated oxidative modifications. In this manner, a freshly synthesized HbpS remains unmodified and inhibits SenS autophosphorylation, providing negative feedback for the HbpS-SenS-SenR signaling cascade (9, 35, 45) (Fig. 6).
Concluding Remarks and Perspectives
Over the last years studies have shown that oxidative attack on macromolecules, including proteins as well as nucleic acids, results in specific damage events. A number of systems have been described by which living organisms can sense and react to an increase in oxidative damage, which may be as a result of environmental changes, or through the presence of competing organisms. Some of these systems have evolved to efficiently recognize this oxidative damage, using the modified molecules as mediators in signaling cascades that control the expression of genes involved in antioxidative stress responses. DNA-based protection can be generated by the transport of charges across the DNA, facilitating the transport of a mutagen (guanine radical) from more (coding) to less (noncoding) important DNA regions. Studies on the redox regulator SoxR have also demonstrated that guanine radicals generated via guanine multiplets, near the SoxR-binding site, can oxidize SoxR, modulating its activity. Thus, analyses of DNA-binding sites of other redox transcriptional regulators in order to identify neighboring guanine multiplets may provide supporting data for a general mechanism of DNA-based signaling. Interestingly, guanine multiplets (GG and GGG) are to be found near to the DNA-binding site of the iron-dependent redox regulator FurS from S. reticuli (49). This transcriptional repressor undergoes a H2O2-mediated intramolecular disulfide bond formation that leads to conformational changes, resulting in a loss of FurS-binding to its own DNA operator sequence within the regulatory region of the target operon, furS-cpeB. In future, studies will be pursued in order to establish the extent by which DNA-mediated signaling co-regulates the activity of FurS under oxidative stress. It should be noted that the furS-cpeB operon is also under the control of the redox-sensing system HbpS-SenS-SenR that mediates the response to extracellular iron-based oxidative stress.
It will be also challenging to analyze whether a comparable RNA-based signaling exists. Such a process might provide a redox-based mechanism to modulate the activity of RNA-binding regulators (e.g., IRPs). In support of this a regulatory function of RNA has been demonstrated for small RNAs and riboswitches can regulate gene expression, at transcriptional or translational levels, in a ligand-dependent manner. As redox metabolites and metal-ion-containing riboswitches have been identified, it will be interesting to explore the ability of such RNA elements to sense redox stress-based signals that would result in changes of the oxidation state of the ligand. Such changes may subsequently be able to induce conformational changes of the riboswitch, resulting in a fine-tuning of regulatory response.
Protein-based sensory and regulatory systems are well known. Most of these systems possess a sensory and regulatory module. An exception is the HbpS-SenS-SenR system in which the extracellular protein HbpS acts as an accessory module, interacting with the membrane-embedded SK SenS. The octomeric HbpS senses extracellular iron-mediated oxidative stress signals, which lead to oxidative modifications accompanied by conformational and structural changes of HbpS, activating SenS. The activity of the SK SenS leads to the expression of antioxidative genes (i.e., those coding for the mycelium-associated catalase-peroxidase CpeB) via the RR SenR. The CpeB enzyme has been shown to protect HbpS from oxidation; and unmodified HbpS inhibits SenS autophosphorylation, resulting in the down-regulation of the HbpS-SenS-SenR signaling cascade (Fig. 6). Given the presence of homologous proteins in the genomes of other organisms, it is highly likely that other accessory-based TCSs sensing systems are present. However, further work is necessary to elucidate the exact nature of the signals that are detected.
Despite the fact that the molecular mechanisms involved in HbpS-based activation/inhibition of SenS are understood, further aspects merit study. As HbpS does not possess any known heme-binding signature, it was postulated to be a novel type of heme-binding protein. Therefore, the elucidation of heme-binding site(s) within the octomeric assembly of HbpS using structural biology will deepen the understanding of the heme-mediated regulation of HbpS. Moreover, it will give further insights into the heme-degradation processes and the ability of HbpS to transport and coordinate the released iron. The coordination of extracellular iron ions or sources of iron is highly relevant, as these processes, together with other HbpS-independent ones, may ensure that under iron-rich conditions these potentially damaging ions are maintained in the extracellular space and are not transported in to the cytoplasm.
Footnotes
Acknowledgments
We are very grateful to all our collaborators—in particular to Prof. Dr. H. Schrempf (Applied Genetics of Microorganisms, University of Osnabrueck). This work was supported by Grant OR 224/1-3 of the Deutsche Forschungsgemeinschaft (DFG).