
Abstract
Oxidative stress plays a fundamental role in the development of diabetes, which has become a great threaten for health in the whole world. In recent years, it has been found that, in addition to the effect on metabolism, insulin plays an antioxidant role. However, the effect of insulin on the whole antioxidant enzyme system, especially in vivo, is not completely understood. We note that, in vitro and in vivo, insulin administration could sequentially and transiently increase a battery of antioxidant enzymes through the activation of the key transcription factor nuclear factor erythroid 2 p45-related factor 2 (Nrf2). The sequential activation of extracellular signal–regulated kinases (ERK)-protein kinase B (Akt) pathway maybe required for insulin-induced enhancement of antioxidant defense regulated by Nrf2. Our observation leads to the hypothesis that insulin regulates the redox balance and insulin bolsters antioxidant defenses via the ERK-Akt-Nrf2 pathway. Antioxid. Redox Signal. 16, 1061–1070.
Diabetes and Antioxidant Role of Insulin
It is well-known that insulin regulates carbohydrate and fat metabolism in the body, through stimulating a complex web of intracellular downstream signaling pathways. In addition to the effect on metabolism, it is proposed that insulin may directly affect the endogenous antioxidant defense to protect against oxidative stress induced by the metabolism of elevated levels of glucose or free fatty acids. Several recent studies have demonstrated that insulin may influence redox balance through regulating antioxidant enzymes, including γ-glutamylcysteine ligase (GCL) and heme oxygenase-1 (HO-1), under different experimental conditions (2, 4). However, the effect of insulin on the whole antioxidant enzyme system is not completely understood and the possible mechanisms in vivo are still to be clarified.
Innovation
Oxidative stress has been believed to play a causal role in the pathogenesis of diabetes. Antioxidant role of insulin is likely to be involved in mediating the protective effect of insulin in diabetic animals and patients. However, the effect of insulin on the whole antioxidant enzyme system and possible molecular mechanisms, especially in vivo, are still to be clarified. In BRL-3A hepatocytes, rat, and mouse livers, we examined the effect of insulin administration on nuclear factor erythroid 2 p45-related factor 2 (Nrf2) and a series of antioxidant enzymes. Insulin sequentially and transiently increased the expression of a battery of antioxidant enzymes that was controlled by Nrf2. The ROS-ERK-Akt-Nrf2 signaling cascade maybe required for the activation of insulin-induced antioxidant defense. To the best of our knowledge, this article is the first to report that insulin could bolster antioxidant defenses via the ERK-Akt-Nrf2 pathway in vitro and in vivo. The novel results detailed and discussed in this article may better the understanding of the benefit of insulin on the treatment of diabetes and other oxidative stress–related diseases and may have important clinical implications.
Insulin and Nuclear Factor Erythroid 2 p45-Related Factor 2-Controlled Antioxidant Enzymes
Oxidative stress is defined as an imbalance between the production of reactive oxygen species (ROS) and the body's ability to detoxify ROS. In the body, there are many types of ROS, including superoxide (O2 −·), hydrogen peroxide (H2O2), hydroxyl (·OH)—which could result in the production of other free radicals, peroxyl, alkoxyl, and thiyl—and nitric oxide and peroxynitrite, also called reactive nitrogen species. Various antioxidant enzymes make up a strong defense against potential oxidative challenge. Superoxide dismutase (SOD), including SOD1 (in the cytoplasm) and SOD2 (in the mitochondria), catalyzes the dismutation of O2 −· into O2 and H2O2. Then, catalase (CAT) functions to catalyze the decomposition of H2O2 to H2O and O2. GCL is the rate-limiting enzyme of glutathione (GSH) synthesis, which is composed of a catalytic subunit (GCLc) and a modifier subunit (GCLm). Glutathione peroxidase (GPx) is the major enzyme responsible for the reduction of organic peroxides using GSH as a cofactor. HO-1 catalyzes the rate-limiting step in the degradation of free heme released upon oxidative stress, yielding equimolar quantities of biliverdin, iron, and carbon monoxide, which have potent antioxidant activity and cytoprotective effect. Peroxiredoxin (PRX) is a family of ubiquitous and abundant proteins that are important for antioxidant defense and regulate cell signaling pathways. Nuclear factor erythroid 2 p45-related factor 2 (Nrf2) is a crucial regulator of the cellular redox homeostasis. Under nonstressful conditions, Nrf2 is kept in the cytosol by Kelch-like ECH-associated protein 1 (Keap1), and upon recognition of signals imparted by oxidative and electrophilic molecules, Nrf2 is released from Keap1 and translocates to the nucleus to induce genes involved in defense and survival (3).
Considering the multiforms and high reactivity of ROS, we could not lay hope on activating a single antioxidant enzyme to fight against oxidative stress. In the study, we measured the protein expression of various antioxidant enzymes after the exposure to insulin in vitro and in vivo. BRL-3A cells were exposed to 100 nM insulin for 0–16 h. During the culture period, Nrf2 expression was transiently increased, with the highest expression at 1–4 h that returned to basal level after 8 h (Fig. 1A) (results expressed as means±SD were shown in Supplementary Fig. S1A; Supplementary Data are available online at
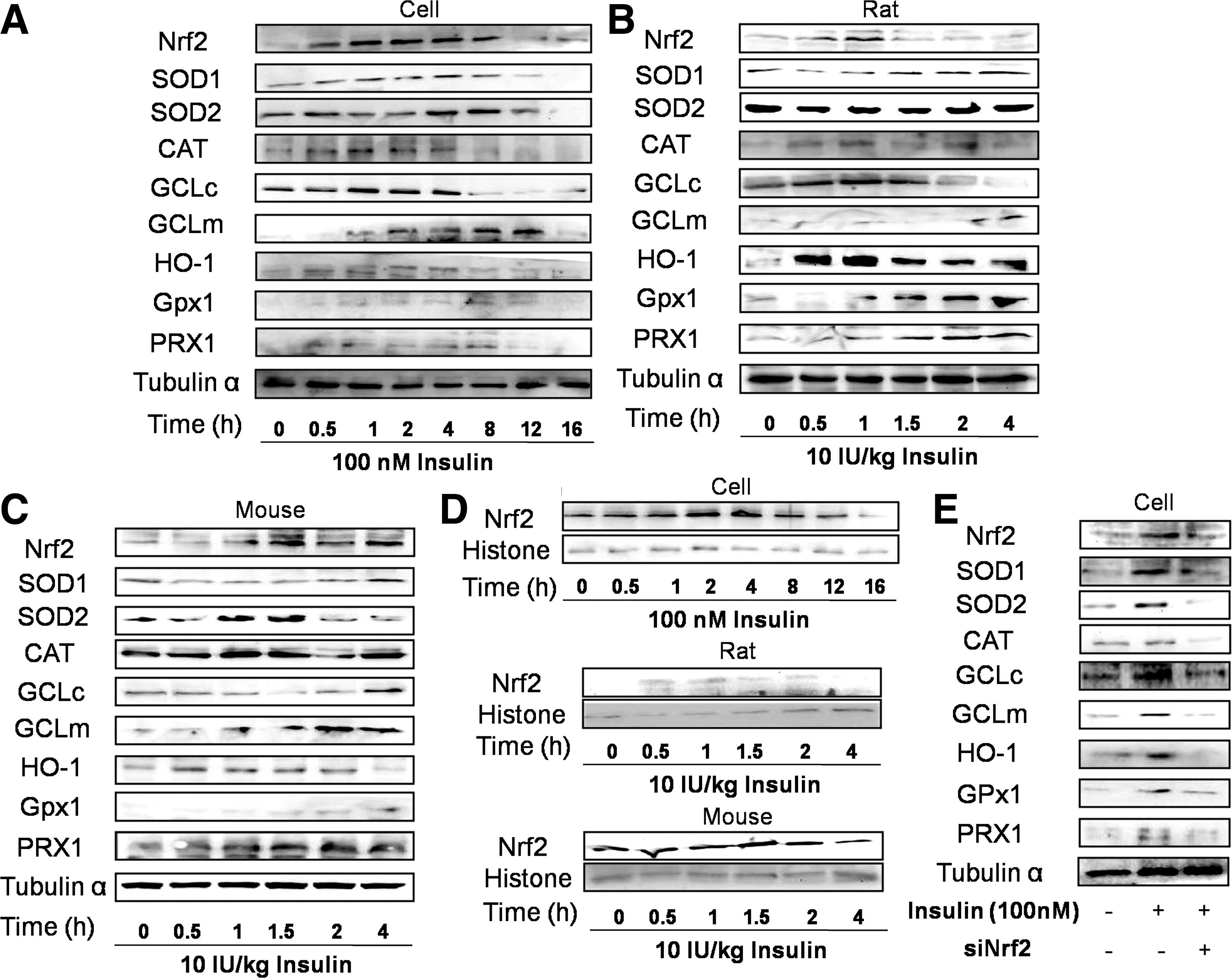
Then, we examined nuclear expression of Nrf2. In BRL-3A cells, rat and mouse livers, insulin administration notably induced a transient increase of nuclear expression of Nrf2 (in Fig. 1D and Supplementary Fig. S1D). To further detect whether the enhancement of antioxidant enzymes was attributed to Nrf2 activation, small interfering RNA of Nrf2 (siNrf2) was used. In the presence of siNrf2, the increased expression of all these antioxidant enzymes was inhibited (Fig. 1E and Supplementary Fig. S1E), indicating that Nrf2 was required for insulin-stimulated regulation on antioxidant enzymes. In agreement with previous studies, Nrf2 activation becomes an efficient strategy to solid antioxidant defense through inducing various antioxidant enzymes and our study suggests that insulin acts as a potent stimulus of this pathway.
Are There Relationships Between Moderate Generation of ROS and Insulin-Induced Activation of Nrf2?
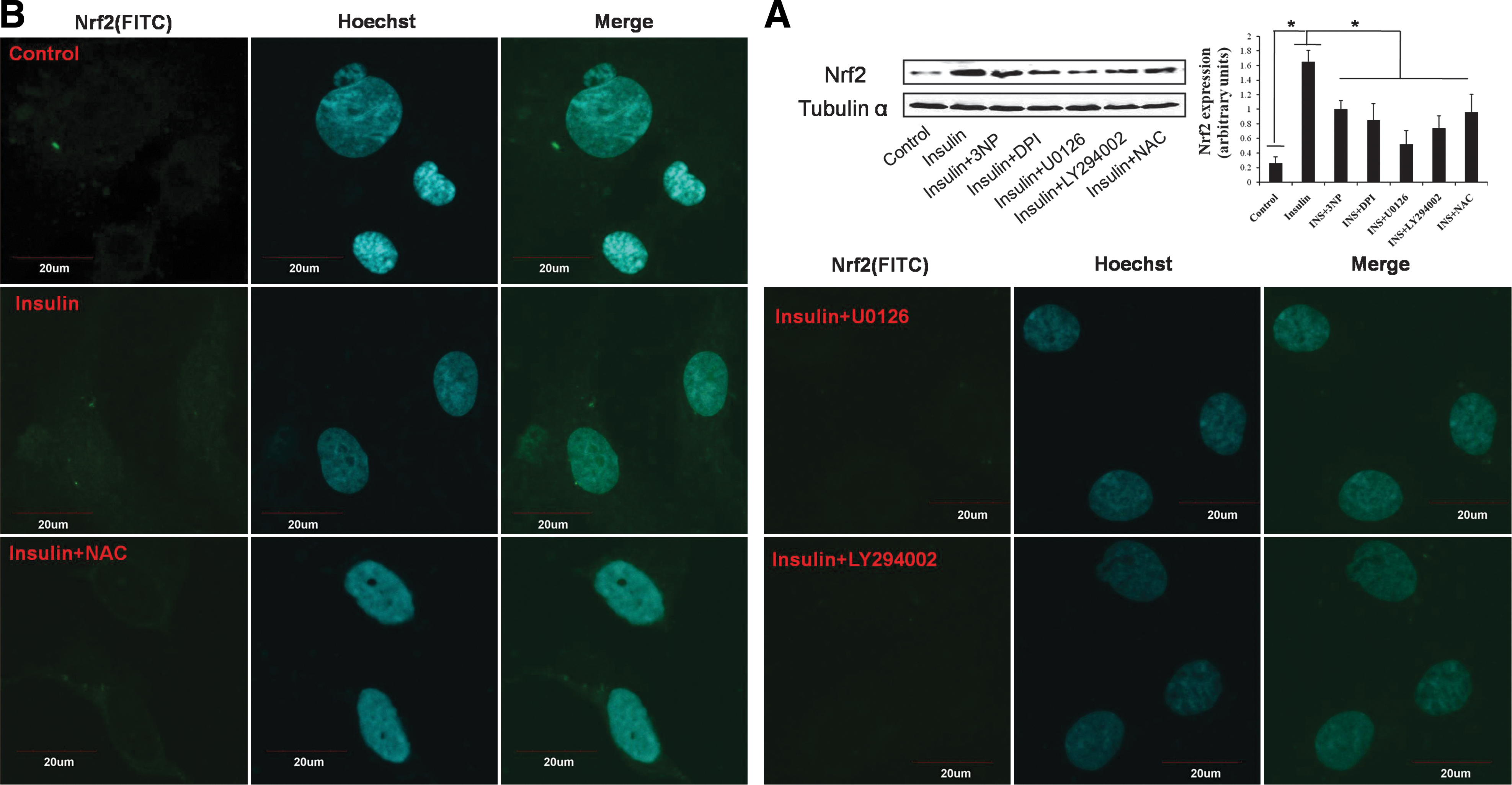
Low concentration of ROS is a definite stimulus for the activation of Nrf2. In the current study, 100 nM insulin resulted in a dynamic alteration of fluorescence intensity, with the highest fluorescence intensity at 30 min (Fig. 2). In addition, intracellular and extracellular ROS generation was also detected separately, which confirmed that insulin significantly increased ROS production transiently (Supplementary Fig. S2). Mitochondria and nicotinamide adenine dinucleotide phosphate (NADPH) oxidase are the main sources of ROS in a cell. As shown in Figure 2 and Supplementary Fig. S2, ROS induced by insulin was significantly inhibited by 3 nitropropionic acid (3NP, an inhibitor of mitochondria), diphenyliodonium (DPI, an inhibitor of NADPH oxidase), and N-acetyl-L-cysteine (NAC), indicating that ROS mainly derived from mitochondria and NADPH oxidase under the present condition. In addition, in BRL-3A cells, incubation with 3NP, DPI, and NAC could significantly inhibit insulin-stimulated expression of Nrf2 (Fig. 3A, B). Furthermore, in mouse livers, 3NP, DPI, and NAC could significantly inhibit the increased expression of Nrf2 induced by insulin (Fig. 4). These results indicated that moderate ROS generation derived from mitochondria and NADPH oxidase was required for insulin-stimulated activation of Nrf2 and thus the downstream antioxidant enzymes.


Is Extracellular Signal–Regulated Kinases-Protein Kinase B Signaling Pathway Required for the Antioxidant Role of Insulin?
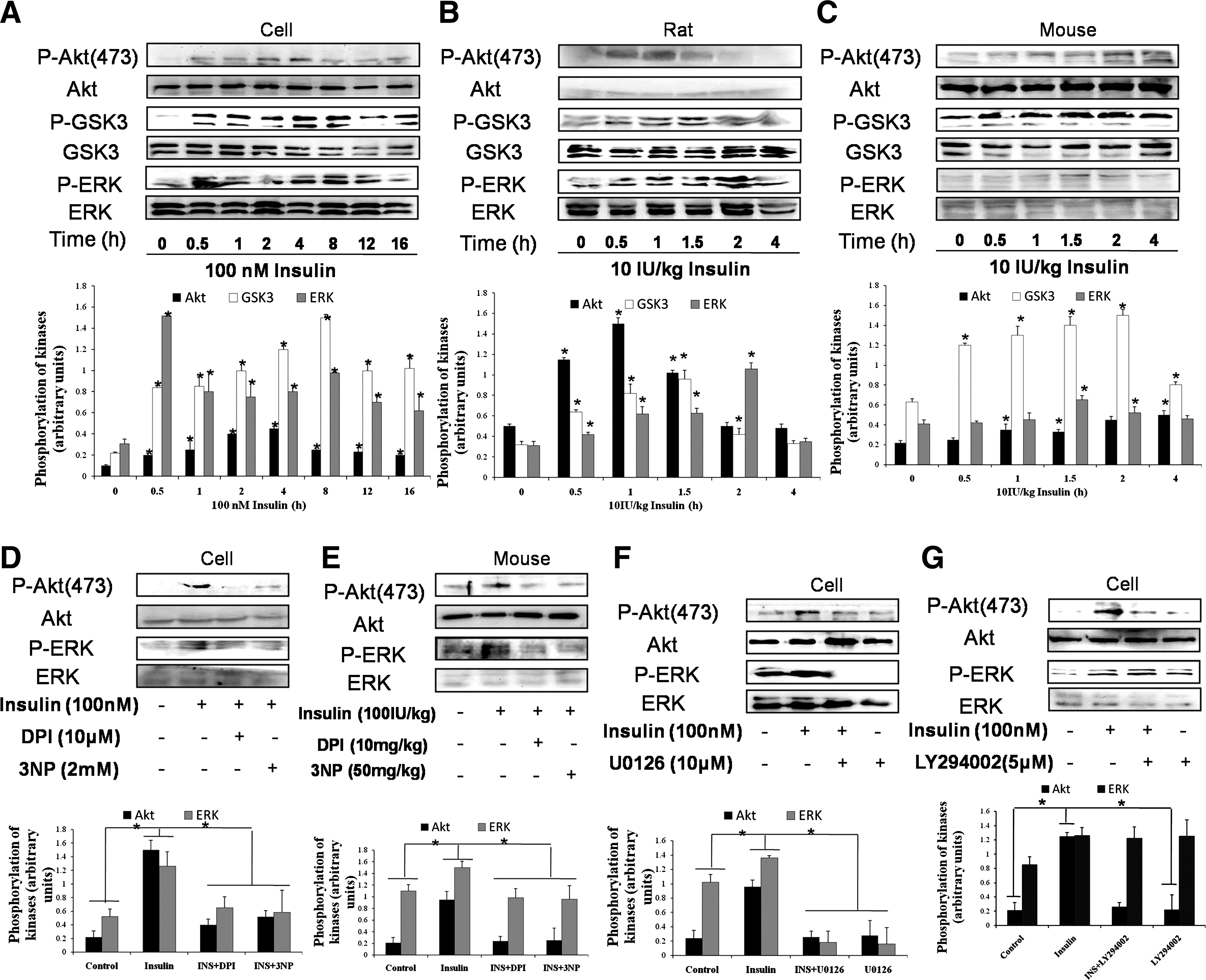
Our previous results suggest that the activation of protein kinase B (Akt) and the extracellular signal–regulated protein kinases (ERK) maybe involved in Nrf2 activation induced by several other agents (7 –9). We then determined possible role of Akt and ERK in insulin-induced effect on Nrf2-controlled antioxidant enzymes. In hepatocytes and rat livers, insulin induced a transient increase of the phosphorylation of Akt, ERK, and glycogen synthase kinase 3α/β (GSK3α/β), downstream target of Akt (Fig. 5A, B). In mouse livers, insulin time-dependently induced the phosphorylation of Akt and GSK3α/β and induced a transient increase of the phosphorylation of ERK (Fig. 5C). DPI and 3NP significantly inhibited insulin-induced phosphorylation of Akt and ERK both in vitro and in vivo (Fig. 5D, E). Then, the sequence of Akt and ERK phosphorylation was also evaluated, using U0126, an inhibitor of MEK/ERK, and LY294002, an inhibitor of PI3K/Akt. U0126 significantly inhibited both of ERK and Akt phosphorylation (Fig. 5F). However, inhibition of Akt by LY294002 had no effect on insulin-induced phosphorylation of ERK (Fig. 5G). These results demonstrated that ERK may act as an upstream kinase of Akt in the current experiment. We further detected the effect of U0126 and LY294002 on Nrf2 expression in the presence of insulin. U0126 and LY294002 significantly decreased insulin-induced expression (Fig. 3A) and nuclear accumulation (Fig. 3B) of Nrf2 in BRL-3A cells. Moreover, in mouse livers, U0126 and LY294002 remarkably inhibited insulin-induced Nrf2 expression (Fig. 4). These results demonstrated that the activation of ERK-Akt signaling induced by ROS was responsible, at least partly, for the enhancement of Nrf2-controlled antioxidant defense induced by insulin.
Does Insulin Protect Against Tert-Butyl Hydroperoxide–Induced Oxidative Damage?
In the following experiments, we tested the effect of insulin on tert-butyl hydroperoxide (tBHP)–induced cytotoxicity. When 10 and 100 nM insulin was present, the decrease of cell viability induced by tBHP was significantly inhibited (Fig. 6A). siNrf2 significantly inhibited the protective effect of insulin on cell viability (Fig. 6B). U0126 and LY294002 notably inhibited the insulin-induced protective effect on cell viability (Fig. 6C). In addition, insulin significantly inhibited tBHP-induced apoptosis (Fig. 6D). These data suggested that insulin could protect cells against cytotoxicity through activating Nrf2-controlled antioxidant defense via ERK/Akt signaling cascade.

Concluding Remarks and Future Directions
These data support the hypothesis that insulin bolsters antioxidant defenses via the ERK-Akt-Nrf2 pathway through moderate generation of ROS signal. There are still several questions requiring us to answer. (i) What is the essence of insulin effect? Insulin possesses both of “good” (antioxidant) and “bad” (prooxidative) effect. A proper understanding of this phenomenon could be that insulin is a systematic hormone, which is responsible for the maintenance of redox balance of the whole body (8). (ii) Is concentration the determinant of the biological effect of ROS? It appears paradoxical that insulin bolsters antioxidant defense via promoting ROS generation. In this case, ROS generated by insulin stimulation may act as required signal (1, 5) for the activation of downstream cascades. The phenomenon further reflects that ROS acts as a double-edged sword (6) and the concentration may serve as an “on-off” factor, determining the directions of the biological effect of ROS. (iii) How to fight against potential oxidative stress? There are a series of antioxidant enzymes and antioxidant proteins, serving as antioxidant defense against oxidative stress. Various antioxidant enzymes may constituent an “antioxidant enzyme chain” (8), which could be activated sequentially under the regulation of key transcription factors, including Nrf2, to resist potential oxidative challenge. (iv) How to utmostly and clinically utilize the benefit of insulin? In type 2 diabetic patients, under insulin-resistant condition, insulin signaling cascade is disturbed and the structure of insulin maybe modified under oxidative stress condition. A proper strategy is needed to utilize the redox-regulatory role of insulin. Answering these questions will undoubtedly help find out the essence of the biological effect of insulin and provide new insight into the mechanisms underlying the role of insulin in keeping redox balance.
Notes
Materials and reagents
P-ERK, P-GSK, GSK, Akt, ERK, HO-1, SOD1, and SOD2 antibodies were purchased from Epitomics. P-Akt, GCLc, and tubulin α antibodies were purchased from Bioworld Technology. Nrf2, CAT, PRX1, and GPx1 antibodies were purchased from Santa Cruz Biotechnology. Insulin, DPI, 3NP, LY294002, U0126, H2DCF-DA, and NAC were purchased from Sigma–Aldrich. Most of the other chemicals and reagents used in this study were also procured from Sigma.
Animal experiments
Animal experiments were approved by University Ethics Committee. Sprague–Dawley rats (180–220 g) and Kunming mice (20–22 g) were purchased from the Experimental Animal Center of The Fourth Military Medical University. They were maintained in a standard environmental condition and fed with a standard pellet diet and water ad libitum. The rats and mice were housed in stainless steel cages in a room kept at 22°C±1°C with a 12-h light/12-h dark cycle.
In insulin-treating groups, rats and mice were intraperitoneally injected with 10 IU/kg insulin. Control rats and mice were administrated with the vehicle (saline). About 0, 0.5, 1, 2, and 4 h after the injection of insulin, rats and mice were sacrificed and livers were collected for detection. In some experiments, several mice received an intraperitoneal injection of DPI (10 mg/kg), 3NP (50 mg/kg), U0126 (10 mg/kg), LY294002 (10 mg/kg), and NAC (50 mg/kg) 30 min before the injection of insulin to investigate the signaling pathways responsible for the effect of insulin. At the end, the rats were anesthetized with sodium pentobarbital (60 mg/kg) and then liver samples were harvested for the assays.
Cell culture and treatment
The rat normal hepatocyte cell line BRL-3A was purchased from Shanghai Cell Biology, Chinese Academy of Sciences. The cells were cultured in RPMI 1640 supplemented with 10% fetal bovine serum (Gibco BRL). The experiments were conducted using serum-free RPMI 1640. After growing into confluence, cells were exposed to 100 nM insulin in the presence or absence of several inhibitors and antioxidant for indicated time periods. At the end, cells were harvested for the detection of western blotting and other analyses.
Western blotting
Western blotting was conducted as described previously (7). Briefly, liver and cell lysates were prepared by incubation on ice with lysis buffer and total protein was extracted. Protein extractions were separated by using SDS-PAGE on 10% polyacrylamide gels, and transferred to nitrocellulose membranes (Millipore). After blocking for 1 h with 8% skimmed milk in TBS buffer (10 mM Tris and 150 mM NaCl), the membrane was incubated with indicated primary antibodies overnight at 4°C. After the membrane was washed four times for 15 min each with TBST buffer (10 mM Tris, 150 mM NaCl, and 0.1% Tween-20), it was incubated in the appropriate HRP-conjugated secondary antibody (diluted 1:5000 in TBST) at 37°C for 30 min. The protein bands were visualized using chemiluminescent reagents according to the manufacturer's instructions and quantified using an image analyzer Quantity One System (Bio-Rad). All protein quantifications were adjusted for the corresponding tubulin α or histone (nuclear protein) level, which was not consistently changed by the different treatment conditions.
Immunostaining
For immunofluorescence histochemistry, nonspecific antibody-binding sites were first blocked with 5% BSA, and then slides were incubated with anti-Nrf2 antibody (diluted 1:50) overnight and then incubated with fluorescein isothiocyanate (FITC)–conjugated anti-rabbit (diluted 1:50) for 30 min at 37°C. After washing for three times, slides were incubated with Hoechst for 5 min at room temperature. Specimens were examined with a laser scanning confocal microscope (Olympus).
For Nrf2 subcellular localization, BRL-3A cells were fixed in a freshly prepared solution of 4% paraformaldehyde, rinsed, and permeabilized with 0.1% Triton X-100 in phosphate-buffered saline (PBS). Nonspecific binding was blocked and cells were incubated overnight at 4°C with anti-Nrf2 antibody and then incubated with FITC-conjugated anti-rabbit antibody (diluted 1:50). Hoechst was used to label nuclear DNA. Specimens were examined with a laser scanning confocal microscope (Olympus).
Preparation of nuclear extracts
Nuclear protein preparation was conducted according to the manufacturer's instructions. Briefly, cells were rinsed with ice-cold PBS, homogenized on ice in 1 ml PBS, and centrifuged for 3 min (800 g, 4°C). The pellet was resolved in 600 μl of buffer A. The tissues were lysed in the presence of 0.2% Nonidet P-40 (NP-40) on ice for 15 min with occasional vortexing. The lysates were centrifuged for 5 min at 16,000 g at 4°C and the supernatant was saved as cytoplasmic extract. The nuclear pellet was resuspended in nuclear extraction buffer B on ice, with gentle shaking for 40 min, and then centrifuged at 16,000 g for 10 min at 4°C. The supernatant (nuclear extract) was either stored at −80°C or quantified freshly using BCA protein assay kit. The nuclear extract was used for western blotting.
Dynamic determination of ROS generation
Dynamic measurement of ROS generation in cells for 1.5 h was conducted by the oxidation-sensitive fluorescent probe H2DCF-DA. In brief, after the addition of insulin, inhibitors, and H2DCF-DA (10 μM), the culture dish was immediately inserted into a laser scanning confocal microscope (Olympus). Dynamic production of ROS was evaluated by recording the trend of fluorescent intensity.
siRNA preparation and transfection
Nrf2-SiRNAs targeting three positions (SiRNA-1: Sense: 5′-GGCUCAUCUCACAAGAGAU-3′, Antisense: 5′-AUCUCUUGUGAGAUGAGCC-3′; SiRNA-2: Sense: 5′-GCAAGACUUGGGCCACUUA-3′, Antisense: 5′-UAAGUGGCCCAAGUCUUGC-3′; SiRNA-3: Sense: 5′-GCAAGAAGCCAGAUACAAA-3′, Antisense: 5′-UUUGUAUCUGGCUUCUUGC) of the rat Nrf2 mRNA were synthesized at Genechem. In brief, BRL-3A cells were plated the day before transfections and grown up to 30%–50% confluence. Transfections were carried out with Lipofectamine 2000 (Invitrogen) following the manufacturer's instructions. Transfected cells were grown for 48 h in a 37°C incubator with 5% CO2 and changed to serum-free medium just before treatments were conducted.
Cell viability
Cell viability was measured using microculture tetrazolium (MTT) assay. Briefly, cells were seeded in 96-well plates. After growing into 50%–60% confluence, cells were pretreated with or without siNrf2 for 48 h, and then exposed to 600 μM tBHP and insulin in the presence or absence of two inhibitors, including DPI and 3NP, for 12 h; then, the supernatant was discarded and cells were rinsed with PBS. Then the cells were treated with 0.5 mg/ml MTT (dissolved in PBS and filtered through a 0.2-mm membrane) at 37°C. Four hours later, the formazan crystals were dissolved in DMSO, and the absorption values were determined at 492 nm on an automated Bio-Rad microtiter plate reader.
Detection of apoptosis
Annexin V FITC/PI kit (Beyotime Institute of Biotechnology) assay was used to evaluate cell apoptosis, according to the manufacturer's instructions. In brief, after the experiment, cells were harvested and washed twice with cold PBS. Then, cells were suspended in 195 μl binding buffer at a concentration of 1×106 cells/ml. About 5 μl of Annexin V FITC was added and cells were gently vortexed and incubated for 10 min at room temperature in the dark. After the cells were centrifuged at 1000 g for 5 min, 190 μl binding buffer and 10 μl PI were added. Cells that have bound Annexin V-FITC will show green staining in the plasma membrane. Cells were placed on a glass slide and covered with a glass coverslip, and then observed under a laser scanning confocal microscope (Olympus). Cells that have lost membrane integrity will show red staining (PI) throughout the nucleus and a halo of green staining (FITC) on the cell surface (plasma membrane).
Statistical analysis
All experiments were performed at least three times, and results were expressed as the means±SD. The results were analyzed by one-way ANOVA followed by a SNK-q test for multiple comparisons. All analyses were performed using the Statistical Package for the Social Sciences (SPSS) software. Data were considered statistically significant for p<0.05.
Footnotes
Acknowledgment
This study was supported by the National Natural Science Foundation No. 30872135.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.