
Abstract
In transsulfuration reaction catalyzed by rat mercaptopyruvate sulfurtransferase (MST), a stable persulfide is formed at the catalytic site cysteine Cys247 as a reaction intermediate. The outer sulfur atom is donated by the substrate, thiosulfate, or by mercaptopyruvate. MST serves as a thioredoxin-dependent antioxidant possessing self-regulated enzymatic activity. After oxidation of persulfurated MST by treatment with hydrogen peroxide, mass spectrometric analysis showed that the outer sulfur atom of the persulfide is oxidized to form Cys-thiosulfenate (Cys-Sγ-SO−), Cys-thiosulfinate (Cys-Sγ-SO2 −), and Cys-thiosulfonate (Cys-Sγ-SO3 −). Next, sulfur acceptor substrates including reduced thioredoxin convert all modified cysteines to nonmodified cysteines. Another sulfur acceptor substrate, cyanide, also converted these cysteines via cyanolysis. Thus, sulfur oxides are suggested to release in the redox cycle of persulfide of MST. Antioxid. Redox Signal. 16, 747–753.
Introduction
During the enzyme-catalyzed process of the transsulfuration of rat MST (EC 2.8.1.2), a stable persulfide is formed at the catalytic site Cys247 as a reaction intermediate. Evolutionarily related rhodanese (5, 6) also forms persulfide (2), which is stable by hydrogen bonds donated from side chains of adjacent amino acids to the outer sulfur atom of the persulfide. Recently, biological importance of the sulfane sulfur of persulfide in a thiol enzyme has been recognized as an antioxidant (9) and a hydrogen peroxide (H2S) precursor (8).
We noticed that persulfurated MST (Cys-Sγ-S−) resisted oxidation to a greater extent than nonmodified MST. Therefore, we assumed that the persulfide played a different biological role. Based on the results of preliminary study using a matrix-associated laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS), we hypothesized that the stable sulfane sulfur of the persulfide proposed the biological importance; the outer sulfur atom of the persulfide at Cys247 was oxidized to form Cys-thiosulfenate (Cys-Sγ-SO−), Cys-thiosulfinate (Cys-Sγ-SO2 −), and Cys-thiosulfonate (Cys-Sγ-SO3 −), which were converted to nonmodified MST by sulfur acceptor substrates including dithiothreitol (DTT), reduced thioredoxin, or cyanide. These processes explain why persulfurated MST resists oxidants. Further, in this redox cycle of the persulfide, potential new signal transducer, sulfur oxide (SO x , x=1, 2, or 3), can be produced.
Innovation
We showed an analysis method using matrix-associated laser desorption/ionization time-of-flight-mass spectrometry (MS), in which shoulder peaks were also analyzed to estimate change in mass number. Results of this study using MS suggest that persulfide plays an important biological role. The outer sulfur atom of the persulfide at Cys247 in a stable reaction intermediate is oxidized by hydrogen peroxide to form thiosulfate, thiosulfinate, and thiosulfenate. Then, sulfur acceptor substrates including dithiothreitol, reduced thioredoxin, thioredoxin with the reducing system, or cyanide convert them to nonmodified mercaptopyruvate sulfurtransferase. During the redox cycle of the persulfide, sulfur oxide (SO x ) can be produced and these persulfide and SO x are a potential new signal transducer like NO x , H2S, and CO x . To confirm it, we have developed electrochemical microanalysis method using SO x (or ionic state SO x+1 −)-selective electrodes, which is much more specific and reliable compared with previous methods that use photometry with pararosaniline, conductometry, chemiluminescence detectors, and fluorophotometry.
Results and Discussion
In vivo persulfuration of the catalytic site Cys247 of MST
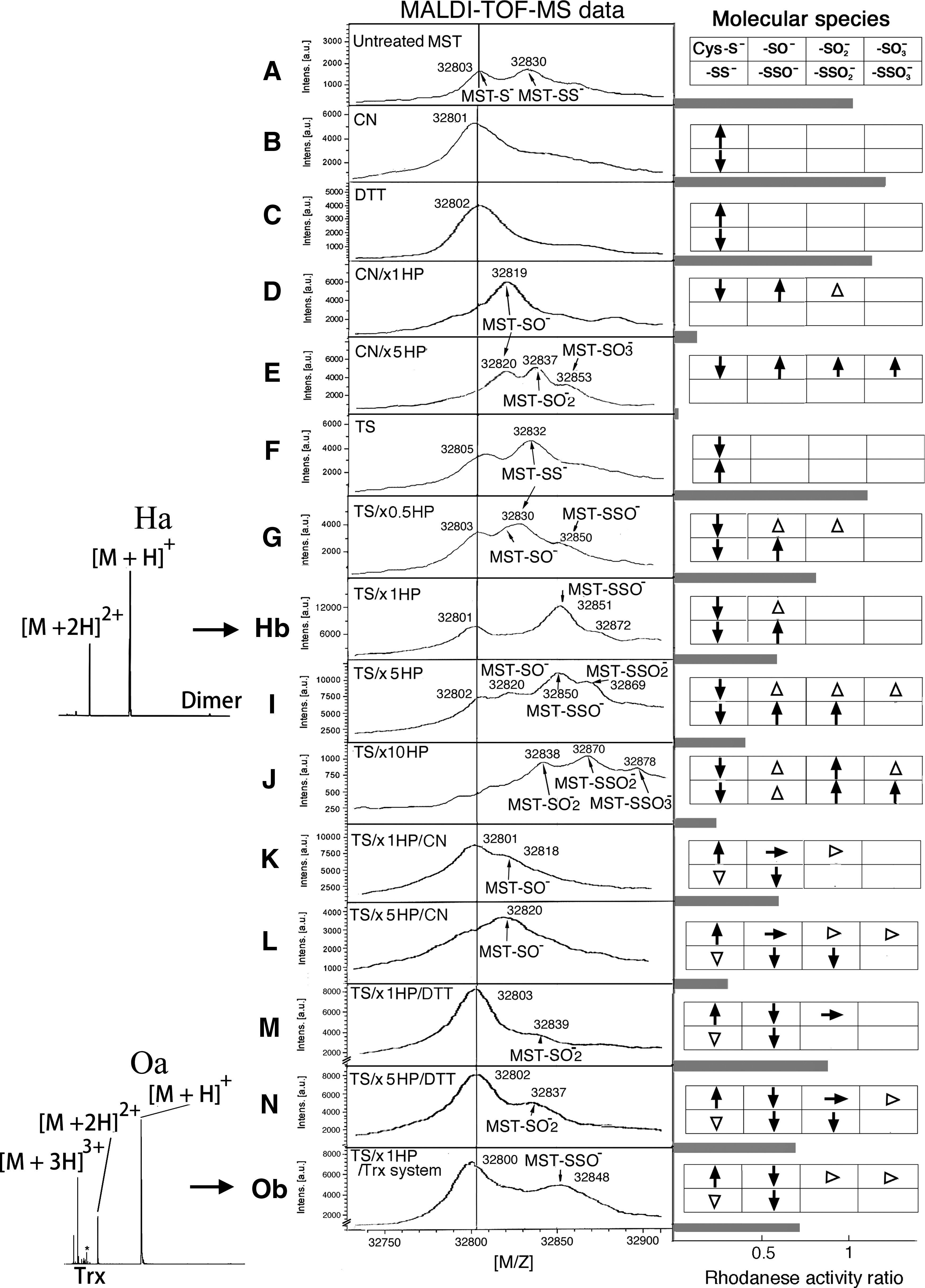
When rat recombinant MST is overexpressed in Escherichia coli, ∼60% of enzymes are converted to persulfurated forms. In a spectrometric data of nontreated MST (sample A), two peaks exhibiting the molecular masses (m/z 32,803; [M+H]+; assigned to as nonmodified MST) and (m/z 32,830; [M+H]+; persulfurated MST) were observed (Fig. 1A). The outer sulfur atom of the persulfide was removed by cyanolysis using cyanide or by reduction using DTT with a shift in the molecular mass to (m/z 32,801; [M+H]+; nonmodified MST) (Fig. 1B) or (m/z 32,802; [M+H]+; nonmodified MST, sample C) (Fig. 1C), respectively. Enzymatic activities did not significantly change (110% and 105% of that of nonmodified MST, respectively) (Fig. 1B and C, respectively).
Oxidation of catalytic site Cys247
Catalytic site Cys247 was confirmed to be a target of oxidants. DTT-treated MST (sample C) was oxidized by a stoichiometric concentration of hydrogen peroxide, resulting in a shift in the molecular mass (m/z 32,802; [M+H]+; DTT-treated MST) to (m/z 32,819; [M+H]+) of sulfenyl MST (4) (Fig. 1D). Oxidation by hydrogen peroxide at a 5-fold molar concentration of MST unevenly shifted the molecular mass (m/z 32,802; [M+H]+; DTT-treated MST) to (m/z 32,820; [M+H]+; sulfenyl MST), (m/z 32,835; [M+H]+; sulfinyl [Cys-SγO2 −] MST), and (m/z 32,853; [M+H]+; sulfonyl [Cys-SγO3 −] MST) (Fig. 1E). Enzymatic activities markedly decreased to 7% of that of nonmodified MST (between samples C and D; p<0.001) (Fig. 1D) and 15% (between samples C and E; p<0.001) (Fig. 1E) of that of nonmodified MST, respectively. These findings confirmed our previous results (4).
In vitro persulfuration of catalytic site Cys247 of MST
DTT-treated MST (sample C) was transsulfurated at Cys247. A new peak exhibiting the molecular mass (m/z 32,832; [M+H]+; persulfurated MST, sample F) appeared (Fig. 1F), which was converted to nonmodified MST during incubation with DTT or cyanide (data not shown). Enzymatic activity did not significantly change (104% of that of nonmodified MST) (Fig. 1F). Notably, in vitro persulfuration of MST was limited to a maximum of ∼70%, probably because oxidized and dimeric (an inactive form) MSTs were remained (4, 7).
Oxidation of persulfurated MST
Persulfurated MST (sample F) was oxidized using hydrogen peroxide at 0.5-, 1-, 10-, and 50-fold molar concentrations of MST to form three species of oxidized persulfides (Fig. 1G–J, respectively). In Figure 1G, new peaks exhibiting the molecular masses (m/z 32,830; [M+H]+; persulfurated MST) and (m/z 32,850; [M+H]+; thiosulfenyl [Cys-Sγ-SO−] MST) appeared. Further, the shoulder peak appeared just before peak (m/z 32,830; [M+H]+; sulfenyl MST). Enzymatic activity decreased to 72% of that of nonmodified MST (between samples A and G; p<0.001) (between samples F and G; p<0.001), primarily due to conversion of nonmodified MST to sulfenyl MST.
In Figure 1H, two new peaks (m/z 32,851; [M+H]+; thiosulfenyl MST) and (m/z 32,872; [M+H]+; thiosulfinyl [Cys-Sγ-SO2 −] MST) appeared. Sample H contains sulfenyl, sulfinyl, thiosulfenyl, and thiosulfinyl MSTs. Enzymatic activity decreased to 56% of that of nonmodified MST (p<0.001) (between samples F and H; p<0.001), probably due to the conversion of nonmodified MST into sulfinyl and/or sulfonyl MST.
In Figure 1I, new peaks (m/z 32,820; [M+H]+; sulfenyl MST), (m/z 32,850; [M+H]+; thiosulfenyl MST), and (m/z 32,869; [M+H]+; thiosulfinyl MST) appeared. Sample I contains sulfenyl, thiosulfenyl, and thiosulfinyl, and may contain sulfinyl and sulfonyl MSTs. Enzymatic activity decreased to 32% of that of nonmodified MST (p<0.001) (between samples F and I; p<0.001), probably due to the conversion of nonmodified MST into sulfinyl and/or sulfonyl MST.
In Figure 1J, new peaks (m/z 32,832; [M+H]+; sulfinyl MST), (m/z 32,870; [M+H]+; thiosulfinyl MST), and (m/z 32,878; [M+H]+; thiosulfonyl MST) appeared. Enzymatic activity decreased to 21% of that of nonmodified MST (p<0.001) (between samples F and J; p<0.001), probably due to the conversion of nonmodified MST into sulfinyl and/or sulfonyl MST.
Cyanolysis of oxidized persulfide–containing MSTs
After incubation of sample H with cyanide, two main peaks were observed (Fig. 1K); the main peak exhibiting the molecular mass (m/z 32,801; [M+H]+; nonmodified MST) was converted from thiosulfenyl and thiosulfinyl MSTs. The second peak (m/z 32,818; [M+H]+; sulfenyl MST) disappeared following incubation with DTT (spectrometric data not shown). Enzymatic activity did not significantly change (57% of that of nonmodified MST [p<0.001]) (between K and H; p=0.43) (Fig. 1K) due to cyanolysis of sample H by cyanide contained in the assay mixture.
After incubation of sample I with cyanide, a peak (m/z 32,820; [M+H]+; sulfenyl MST) (Fig. 1L) appeared and then disappeared following incubation with DTT (spectrometric data not shown). These findings indicated that Sγ of Cys247 is oxidized more easily than the outer sulfur of the persulfide (Fig. 2) and that SO x is released via cyanolysis. Because the assay mixture contained cyanide, it is reasonable to assume that the enzymatic activity was not significantly different between groups treated (Fig. 1L) or not treated (Fig. 1I) with cyanide (32% [p<0.001] and 32% of that of nonmodified MST, respectively).

Reduction of oxidized persulfide–containing MST by DTT
In Figure 1M, after incubation of sample H with DTT, two main peaks appeared; the first peak exhibiting the molecular mass (m/z 32,803; [M+H]+; nonmodified MST) was converted from sulfenyl, thiosulfenyl, and thiosulfinyl MSTs. The second peak (m/z 32,839; [M+H]+) was assigned to be sulfinyl MST. Enzymatic activity increased to 83% that of nonmodified MST (p=0.03) (between samples M and H; p<0.001) because sulfenyl MST was reduced to nonmodified MST in the assay mixture.
In Figure 1N, after incubation of sample I with DTT, two main peaks appeared; the peak (m/z 32,802; [M+H]+; nonmodified MST) was converted from sulfenyl, thiosulfenyl, and thiosulfinyl MSTs. The second peak (m/z 32,837; [M+H]+) was assigned to be sulfinyl MST. Enzymatic activity increased to 73% that of nonmodified MST (p<0.001) (between samples N and I; p<0.001) because sulfenyl MST was reduced to nonmodified MST in the assay mixture.
These oxidized persulfide–containing MSTs were reduced via two possible pathways (A): Cys-Sγ-SO x −→Cys-Sγ−+SO x or (B): Cys-Sγ-SO x −→Cys-Sγ-S−→Cys-Sγ− (Fig. 2). Reaction (A) is reliable in view of redox chemistry: (i) sulfate is released during the reaction of thiosulfonate-containing sulfonucleotide reductase to nonmodified enzyme via reduction by thioredoxin (1) and (ii) during reduction of Cys-thiosulfinate intermediate in peroxiredoxin, glutathione reduces the disulfide bond (3). Cyanide attacks Cys-Sγ-SO x to release SO x via cyanolysis also according to reaction (A).
Reduction of oxidized persulfide–containing MST by thioredoxin and the reducing system
As shown in Figure 1Oa and Ob, after incubation of sample H with rat thioredoxin and the reducing system, which can also proceed in vivo (7), two main peaks were observed; the first peak exhibiting the molecular mass (m/z 32,800; [M+H]+; nonmodified MST) was converted from sulfenyl, thiosulfenyl, and thiosulfinyl MSTs. The second peak (m/z 32,848; [M+H]+) was the remaining thiosulfenyl MST. Enzymatic activity increased to 75% of that of nonmodified MST (p<0.001) (between samples O and H; p<0.001), primarily because sulfenyl MST was partially reduced to nonmodified MST. This study confirms our preliminary results (Fig. 3).
It is generally concluded that SO x can be produced during the redox cycle of the persulfurated MST. The toxicological aspects of SO x , particularly SO2, have been emphasized as air pollutants, but the biological importance has not been examined. They are reactive species that may function in the ionic state as nucleophilic signal transducers. However, a specific microanalysis method to detect these species at the physiological level has not been developed.
Notes
Preparation, overexpression, and purification of rat wild-type MST
Rat recombinant wild-type MST was overexpressed and purified according to a previously described procedure (4).
Removal of an outer sulfur of a persulfide at Cys247 of overexpressed MST using DTT and cyanide
A total of 50 μl of 1.2 mM MST (sample A) was incubated with 5 μl of 1 M potassium cyanide (Wako Pure Chemicals, Osaka, Japan) in 20 mM potassium phosphate buffer, pH 7.4 (buffer A), on ice for 30 min. After the excess of cyanide was removed by gel-filtration using a G-25 column (1×24 cm; GE Healthcare BioSciences, Uppsala, Sweden), the enzyme-containing fraction was concentrated using a VIVASPIN (10,000MWCO, PES; Sartorius, Goettingen, Germany) (sample B). Next, 50 μl of 1.2 mM MST (sample A) was incubated with 5 μl of 1 M DTT on ice for 30 min. After the excess of DTT was removed by gel-filtration using a G-25 column (1×24 cm), each enzyme-containing fraction was concentrated using a VIVASPIN (10,000MWCO) (sample C). A 50 μl aliquot of sample C (1.2 mM) was treated with 5 μl of 1 M potassium cyanide on ice for 30 min. After the excess of cyanide and DTT was removed by gel-filtration using a G-25 column (1×24 cm), each enzyme-containing fraction was concentrated using a VIVASPIN (10,000MWCO) (sample B).
Oxidation of a catalytic site cysteine of MST by hydrogen peroxide and reduction using DTT as a control study
DTT-treated MST was prepared as described previously. A 50 μl aliquot of the MST (2.6 mM) was incubated with 5 μl of 26 mM or 130 mM hydrogen peroxide in buffer A on ice for 20 min. After the excess of hydrogen peroxide was removed using a NAP5 gel filtration column (GE Healthcare BioSciences), each enzyme-containing fraction was collected (samples D and E, respectively).
Determination of sodium thiosulfate concentration for persulfide formation at Cys247
About 10 μl of DTT-treated MST (sample C, 0.5 mM) was incubated with 1 μl of 50, 100, and 500 mM sodium thiosulfate (Wako Pure Chemicals) in buffer A on ice for 20, 40, 60, and 120 min. In our preliminary study, a maximum percentage value of persulfuration of MST was ∼70% (data not shown). In this study, MST was then incubated with sodium thiosulfate at 100-fold molar concentration of MST on ice for 1 h. After the excess of thiosulfate was removed by gel-filtration using a NAP5 column, each enzyme-containing fraction was collected and concentrated using a VIVASPIN (10,000MWCO) (sample F).
Oxidation of the outer sulfur of a persulfide at the catalytic site Cys247 and reduction of the oxidized forms or cyanolysis of the disulfide bonds
After a 30 μl aliquot of the persulfurated MST (sample H) was incubated with 0.5, 1, 5, and 10 μl of 50 mM hydrogen peroxide in buffer A on ice for 20 min, the excess of hydrogen peroxide was removed by gel-filtration using a NAP5 column, and each enzyme-containing fraction was collected and concentrated to 100 μM using a VIVASPIN (10,000MWCO). These samples were referred to as G, H, I, and J, respectively.
For cyanolysis using cyanide, a 50 μl aliquot of each oxidized sample (H and I) was incubated with 10 μl of 500 mM potassium cyanide on ice for 30 min (samples K and L, respectively). For reduction using DTT, a 50 μl aliquot of each sample (H and I) was incubated with 10 μl of 500 mM DTT on ice for 30 min (samples M and N, respectively). For reduction using thioredoxin, 10 μl of 10 μM sample H was incubated with 10 μl of a thioredoxin combined with the reducing system (1 μM rat thioredoxin, 0.05 μM rat thioredoxin reductase [Sigma-Aldrich], and 12.5 μM NADPH [Sigma-Aldrich]), and was incubated on ice for 30 min (sample O).
Mass spectrometric analysis
Each sample was diluted to 1 μM with distilled water (salt concentration should be <2 mM). About 0.5 ml aliquot of each sample in 70% acetonitrile (Wako Pure Chemicals) containing 0.1% trifluoroacetic acid (Wako Pure Chemicals) was mixed with 0.5 μl of sinapinic acid (Bruker Daltonics, Brumen, Germany) saturated in 50% acetonitrile containing 0.1% trifluoroacetic acid. The mixture was dried at room temperature on the target plate.
MALDI-TOF/TOF-MS was performed using an Ultraflex III (Bruker Daltonics) equipped with a SCOUT 384 ion source laser operating in the linear positive mode with a 25 kV acceleration voltage. Mass spectra were obtained by averaging 300 individual laser shots. External mass calibration was performed using the protein mixture of Protein Calibration Standard II (Bruker Daltonics) containing protein A and trypsinogen (m/z 44,613; [M+H]+) and (m/z 23,482; [M+H]+), respectively. Peak determination over the entire mass range and/or on its shoulder in an extended spectrum is reliable as discussed previously (4). The mass measurement error was 0.03% for the mass spectrometric analysis of a chemical modification of 30–40 kDa protein.
Assay for rhodanese activity of MST
A procedure to measure the catalytic activity of transsulfuration from mercaptopyruvate to 2-mercaptoethanol is not appropriate for this experiment because 2-mercaptoethanol can cleave a disulfide bond in the persulfide and/or reduce SO x involving the outer sulfur of a persulfide formed at a catalytic site cysteine during incubation. The rhodanese activity of MST was measured in this experiment although cyanide can cleave a disulfide bond in a persulfide formed at the catalytic site cysteine (4).
Protein determination
Protein concentrations were determined using a Coomassie protein assay kit (Pierce Biotechnology, Inc., Rockford, IL) with crystalline bovine serum albumin (MP Biochemicals, Irvine, CA) as the standard.
Statistical analysis
The significance of differences between values was estimated with Student's t-test. p-Value <0.05 was significant.