
Abstract
Introduction
Cyanobacteria perform oxygenic photosynthesis, which makes them unique among prokaryotes and sets the scene for complex redox biology (22). Cyanobacteria share a common ancestor with chloroplasts of photosynthetic eukaryotes and have a great impact on global photosynthetic productivity (6). Reactive oxygen species (ROS) are generated during aerobic metabolism in most organisms, whereas cyanobacteria and plant chloroplasts should also suffer the consequences of additional ROS production that are associated with the photosynthetic electron transport (PET) and oxygen evolution (9, 22). Adverse environmental conditions, such as extreme temperatures, excess light, or nutrient starvation, lead to elevated ROS levels resulting from molecular oxygen acting as a sink for PET-derived reducing equivalents. The adaptation to such conditions is essential for survival and involves changes of gene expression and enzymatic activities in order to optimize photosynthesis and to avoid toxic ROS levels (9, 22). Therefore, the perception of ROS and subsequent signaling pathways in plants and cyanobacteria remains a topic of active investigation (22, 33).
Innovation
Redox regulation of eukaryotic protein phosphorylation through the reversible cysteine oxidation of kinases and phosphatases is well established. In contrast, such convergence of redox signaling and phosphorylation signaling pathways has not been reported for prokaryotes and kinases, and phosphatases are absent from the prokaryotic disulphide proteomes reported to date. Here, we state that the cyanobacterial Serine/Threonine kinase SpkB is inactivated by oxidation and reactivated through reduction catalyzed by the thioredoxin TrxA. The extreme sensitivity of the SpkB null mutant to oxidative stress demonstrates that this Serine/Threonine kinase is involved in the antioxidant response of this cyanobacterium, thus extending the functions of bacterial eukaryotic-like Serine/Threonine kinases.
Bacterial signaling relies to a large extent on two-component systems involving histidine kinases and their corresponding response regulators and the unicellular cyanobacterium Synechocystis sp. PCC 6803 (hereafter referred to as Synechocystis) harbors 47 histidine kinases (2). DNA microarray experiments have shown that the expression of 32 of the 77 Synechocystis genes strongly induced by H2O2 depends on the histidine kinases Hik33, Hik34, Hik16, Hik41, or the peroxide-sensitive transcriptional regulator PerR (17). However, the ROS-inducible expression of 45 of the 77 genes is not governed by any known factor, which illustrates that there are ROS-inducible signaling pathways which are still to be discovered.
Until recently, phosphorylation in serine, threonine, and tyrosine residues in cellular signaling has been mainly associated with eukaryotes (36). In animal cells, Ser/Thr kinases participate in redox signaling pathways and may also be directly regulated by redox modifications of critical cysteine residues. For example, a member of the mammalian mitogen-activated protein kinase (MAPK) family, MEKK1, is reversibly inhibited by oxidative stress through S-glutathionylation (4). The reduction of oxidized cysteines is frequently catalyzed by thioredoxin, and this has also been observed for eukaryotic protein kinases. For example, the mammalian Ser/Thr protein kinase PKA (48) and the plant Ser/Thr protein kinase STN7 (3, 7, 39) are substrates for thioredoxin. However, the redox regulation of prokaryotic protein kinases has not been reported so far. Previously, we performed in-depth studies of the Synechocystis thioredoxin target proteome and found 77 proteins that interact with thioredoxin (25, 26, 30). Notably, most of these target proteins are metabolic enzymes, and no kinases or phosphatases are identified in these studies. Furthermore, kinases and phosphatases are conspicuously absent from the disulphide proteomes of other prokaryotic organisms reported to date (27 and references therein). This could mean that the thiol-based redox regulation of such enzymes does not occur in prokaryotes or, alternatively, that the kinases and phosphatases are not abundant enough to allow detection in proteomic screens.
Since the first discovery of a eukaryotic-like Ser/Thr protein kinase in the bacterium Myxococcus xanthus (35), progress in large-scale genomic sequencing projects has shown a ubiquitous distribution of such protein kinases in prokaryotes (19). Indeed, approximately two-thirds of all prokaryotes harbor eukaryotic-like Ser/Thr protein kinases (37), indicating the widespread occurrence of signaling involving phosphorylation in serine and threonine residues also in these organisms. The Synechocystis genome encodes seven Ser/Thr protein kinases of the Pkn2-type, named SpkA through G (15, 49). However, SpkE lacks several key amino acids that are necessary for ATP binding and is not a true protein kinase (15). In addition, there are five putative kinases of the ABC1-type, named SpkH through L (50). The functions of a few cyanobacterial Ser/Thr kinases have been addressed through phenotypical characterization of the respective mutants. Thus, the knockout mutants for Synechocystis SpkA and SpkB were found to be deficient in phototactic motility (14, 16), a SpkD null mutant displayed impaired growth at low concentrations of inorganic carbon (23), and a knockout mutant for SpkG was hypersensitive to high salt concentrations (24). However, none of the substrates for any of these enzymes have hitherto been identified, and it remains unknown whether the cyanobacterial Ser/Thr kinases are subject to post-translational modification and regulation.
In this study, we have investigated the influence of the cysteine redox state on Synechocystis protein phosphorylation, given the role of reversible cysteine oxidation in the regulation of Ser/Thr protein kinases in eukaryotes. We also examined the possible roles of the Synechocystis Ser/Thr kinases in tolerance toward ROS. The results obtained extend the roles of Ser/Thr kinases in prokaryotes to include functions in oxidative stress response and affirm that the regulation of these enzymes may be as complex as that of eukaryotes.
Results
Synechocystis protein phosphorylation responds to the cysteine redox state
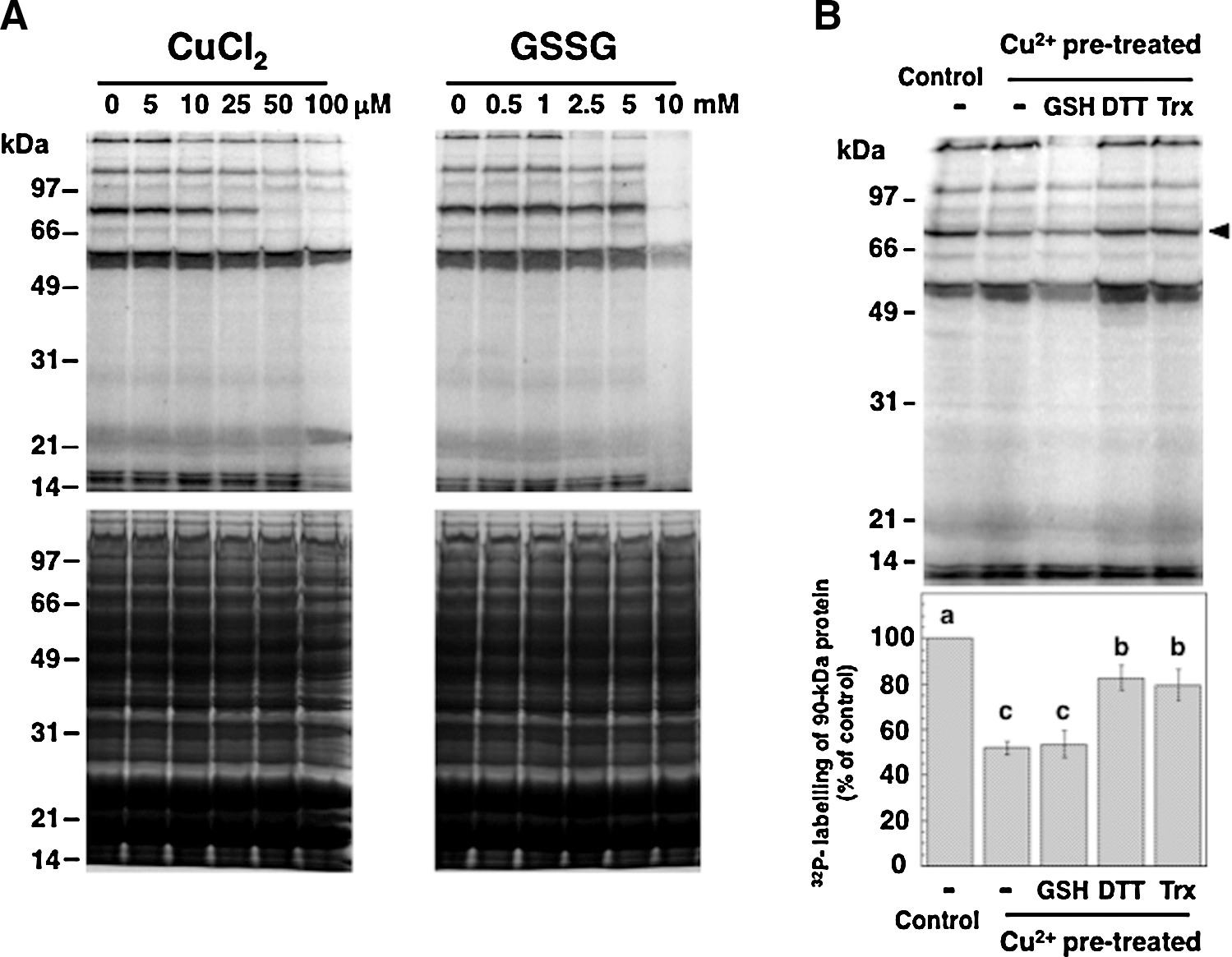
To monitor protein phosphorylation in Synechocystis and to search for a possible redox control of its kinase activities, soluble extracts of Synechocystis cells were pretreated with reagents affecting the cysteine redox state before the radioactive labeling of newly phosphorylated proteins. Initially, the effects of dithiothreitol (DTT), CuCl2, and hydrogen peroxide were tested (Fig. 1). DTT stimulated the phosphorylation of a 53-kDa protein, and hydrogen peroxide had a slight and generalized inhibitory effect. In contrast, CuCl2 strongly inhibited the phosphorylation of two proteins of 150 and 90 kDa (Fig. 1). The ability of Cu2+ to catalyze thiol oxidation, which is superior to that of Fe3+ and Ni2+, has been documented (11), and CuCl2 has been applied as a thiol oxidant in numerous studies on enzymatic redox regulation (13, 34). Oxidized glutathione (GSSG) has also been previously used to monitor the oxidative inactivation of protein kinases (41). Here, CuCl2 and GSSG displayed inhibitory effects on the phosphorylation of some of the heaviest labeled Synechocystis phosphoproteins (Fig. 2A, upper panels). However, GSSG was only efficient at very high concentrations. In order to investigate the nature and possible reversibility of the inhibition induced by Cu2+, samples were briefly pretreated with CuCl2, which was thereafter eliminated. Then, the samples were incubated with reduced glutathione (GSH), DTT, or purified Synechocystis thioredoxin TrxA, before the phosphorylation assay (Fig. 2B). TrxA is the most abundant of the four thioredoxins in Synechocystis and also the only thioredoxin that is essential for the survival of this organism (8). Quantification of the radioactive labeling of a 90-kDa protein showed that the pretreatment with CuCl2 led to about 50% inhibition of its phosphorylation (Fig. 2B). Incubation with DTT or TrxA could restore phosphorylation to about 80% of the control level, whereas GSH did not relieve the inhibition (Fig. 2B).

SpkB is a Ser/Thr kinase responsible for redox-sensitive protein phosphorylation
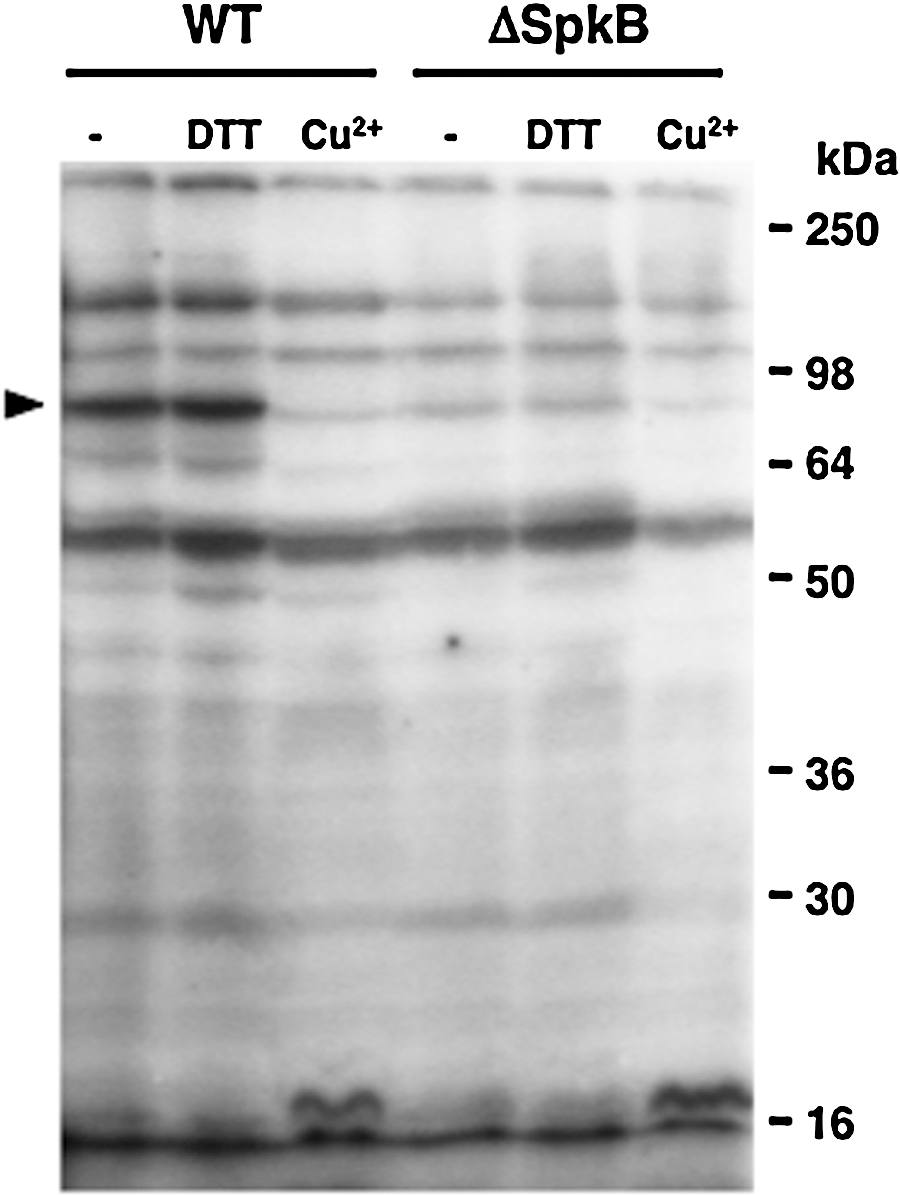
Aiming at identifying the kinase responsible for phosphorylation of the 90-kDa protein, we examined the phosphorylation patterns of a collection of Synechocystis mutants carrying insertions in the genes coding for the six Pkn2-family Ser/Thr kinases (Supplementary Fig. S1B, C; Supplementary Data are available online at
SpkB is inhibited by oxidation and reactivated by thioredoxin-catalyzed reduction
To examine whether the redox sensitivity of this phosphorylation involves modifications of the SpkB kinase, we decided to first clone and express Synechocystis SpkB in Escherichia coli. In an earlier study, the recombinant Synechocystis SpkB was found to be able to undergo autophosphorylation and also phosphorylate casein, myelin basic protein, and histone (14). However, no endogenous substrate or regulation of its activity was reported. In this study, we used casein to monitor the activity of SpkB expressed in E. coli (Fig. 4A). Extracts from E. coli cells transformed with an empty vector did not show casein kinase activity as compared with samples containing only casein without cellular extracts (Fig. 4A). However, the extracts from cells expressing SpkB displayed kinase activity towards casein and, in addition, several other proteins were heterologously phosphorylated by SpkB. The observed activity was substantially inhibited by the incubation of the cell extracts with CuCl2 before the assays (Fig. 4A), suggesting that SpkB might undergo inactivation as a result of thiol oxidation.

In order to test whether thioredoxin could reactivate the oxidized kinase, we intended to purify the recombinant histidine-tagged SpkB. The predominant Coomassie-stained band, sometimes observed as a doublet, was confirmed by MALDI-TOF analysis to be identical to SpkB (Supplementary Table S1). The pretreatment with CuCl2 led to nearly a complete inhibition of SpkB kinase activity (Fig. 4B). Only a partial recovery of the activity was achieved by incubating the SpkB preparation with 0.2 mM DTT, whereas the addition of increasing concentrations of the thioredoxin TrxA significantly enhanced activation of the SpkB kinase (Fig. 4B). Thus, TrxA is able to catalyze the reactivation of oxidized SpkB.
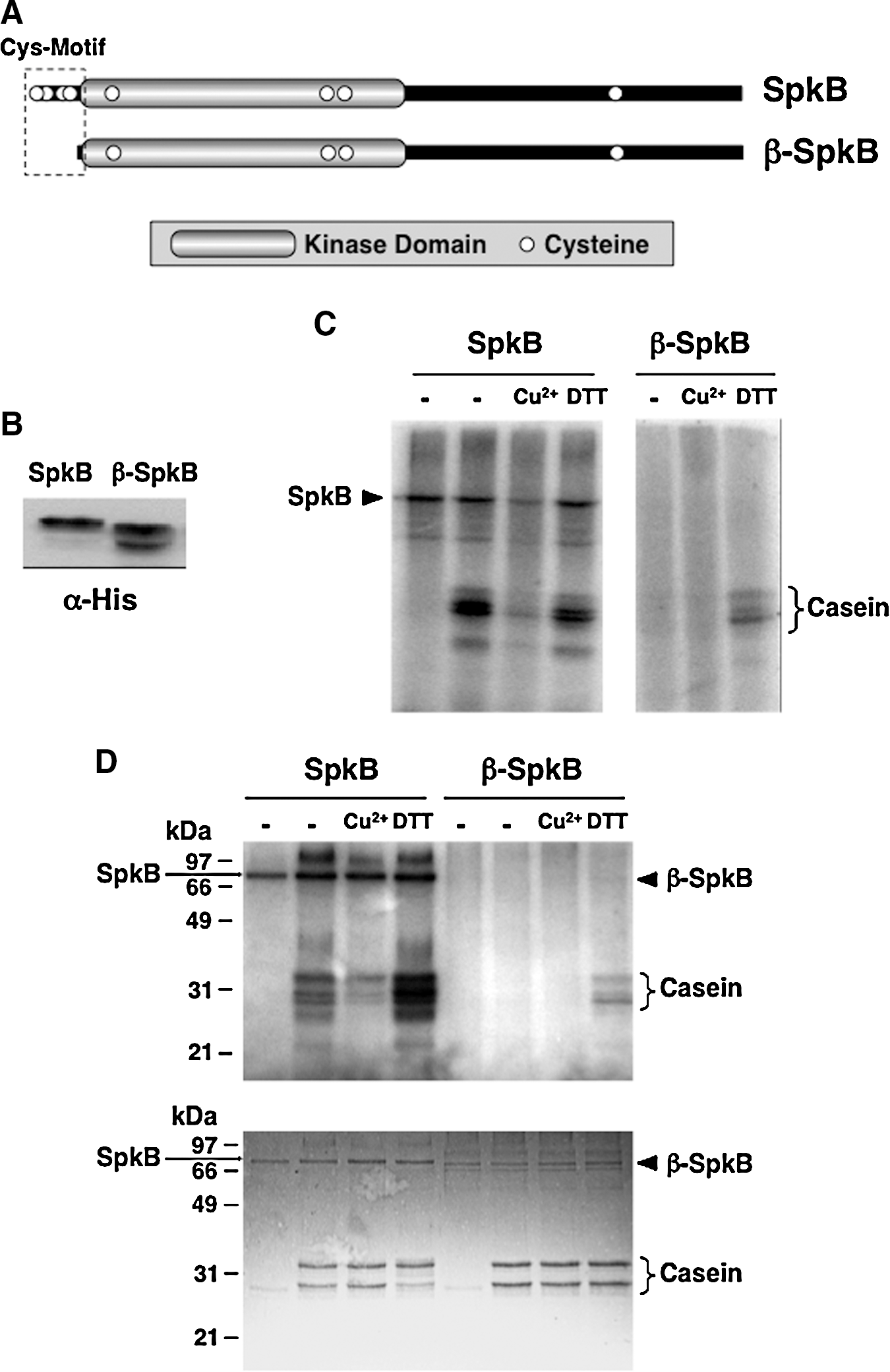
Deletion of the N-terminal Cys-motif of SpkB leads to requirement for exogenous thiols
Since thioredoxin mediates the activation of oxidized SpkB, at least one of its eight cysteines should be involved in such redox-dependent modulation of the kinase activity. The presence of a short N-terminal stretch containing two pairs of cysteines (Fig. 5A) has previously been observed for SpkB (14) and was denoted the “Cys-motif” (15). To investigate the potential role of this motif in the regulation of kinase activity, we constructed a truncated version of the enzyme, which was denoted β-SpkB (Fig. 5A). SpkB and β-SpkB were expressed in E. coli (Fig. 5B), and their activities were compared in cellular extracts using casein as a substrate (Fig. 5C). Unlike SpkB, β-SpkB displayed hardly any casein kinase activity in the absence of additions. However, the addition of DTT induced casein phosphorylation catalyzed by β-SpkB (Fig. 5C), showing that its catalytic activity remained intact. Similar results were obtained when comparing affinity-purified recombinant SpkB and β-SpkB (Fig. 5D).
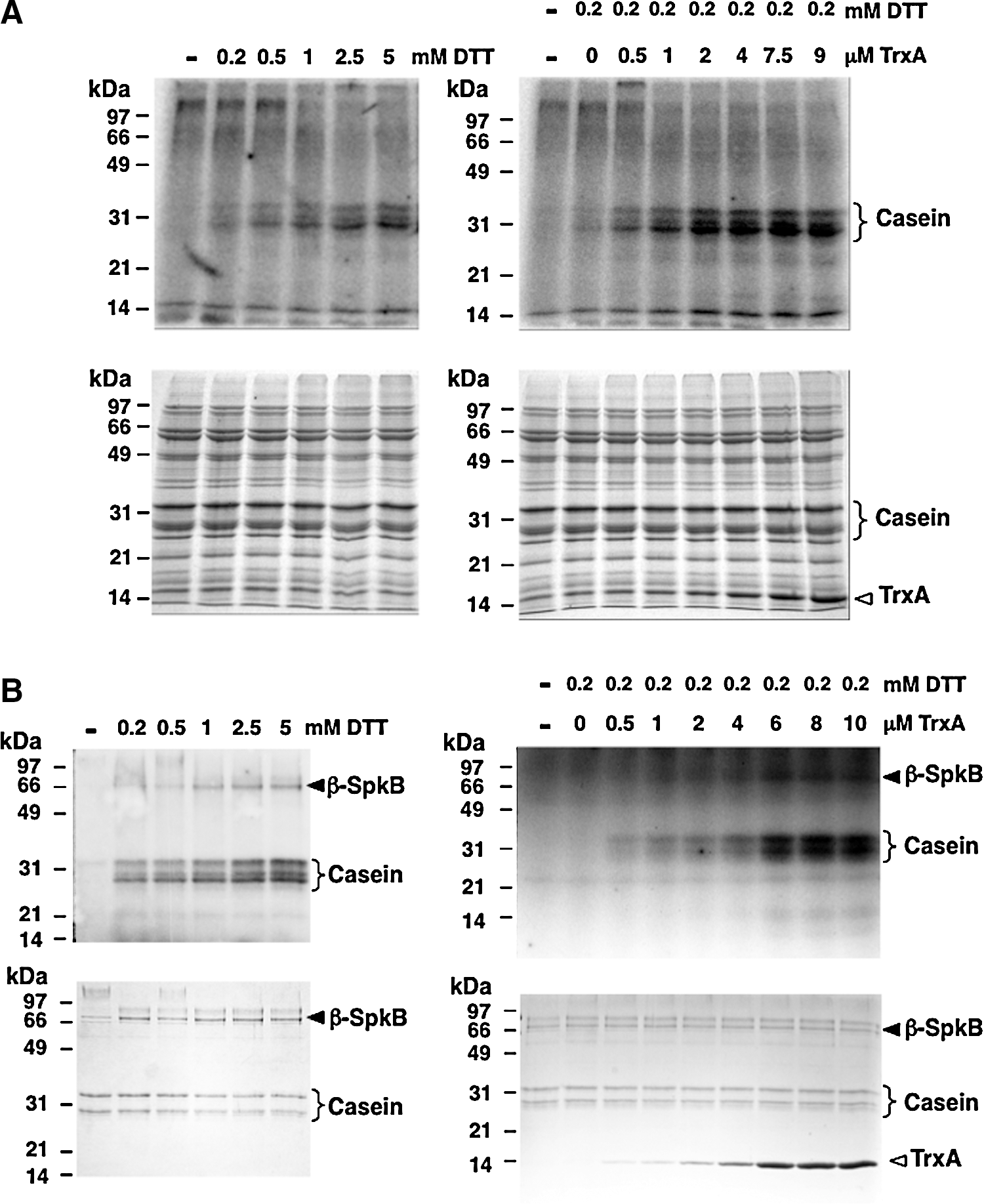
In order to explore the requirement for a reductant of β-SpkB, we performed titrations with increasing concentrations of DTT using extracts from cells expressing β-SpkB (Fig. 6A) and purified β-SpkB (Fig. 6B), respectively. Hence, we found that the addition of DTT at concentrations above 1 mM increased the casein phosphorylation more than 10-fold above the low basal level (graphs shown in Supplementary Fig. S4A, B). The addition of increasing concentrations of TrxA in the presence of 0.2 mM DTT (Fig. 6B, D) led to more than a 50-fold increase in casein phosphorylation as compared with the basal level (Supplementary Fig. S4A, B). Hence, β-SpkB proved to be as active as the full-length enzyme provided that exogenous thiols, such as DTT or thioredoxin, are present in the assay.
The glycyl-tRNA synthetase β-subunit GlyS is a substrate for SpkB
In order to facilitate identification of the prominent 90-kDa phosphoprotein, radioactively labeled samples from WT and ΔSpkB strains were subjected to isoelectric focusing/SDS-PAGE two-dimensional electrophoresis (Fig. 7). Autoradiographs (Fig. 7, right panels) clearly showed the intense labeling of the 90-kDa phosphoprotein in the WT, but not in ΔSpkB. This label corresponded to a protein spot visualized by Coomassie stain in both WT and ΔSpkB (Fig. 7, left panels). The identification by peptide mass fingerprinting revealed that the protein giving rise to this spot is the glycyl-tRNA synthetase β-subunit GlyS (Supplementary Table S1).
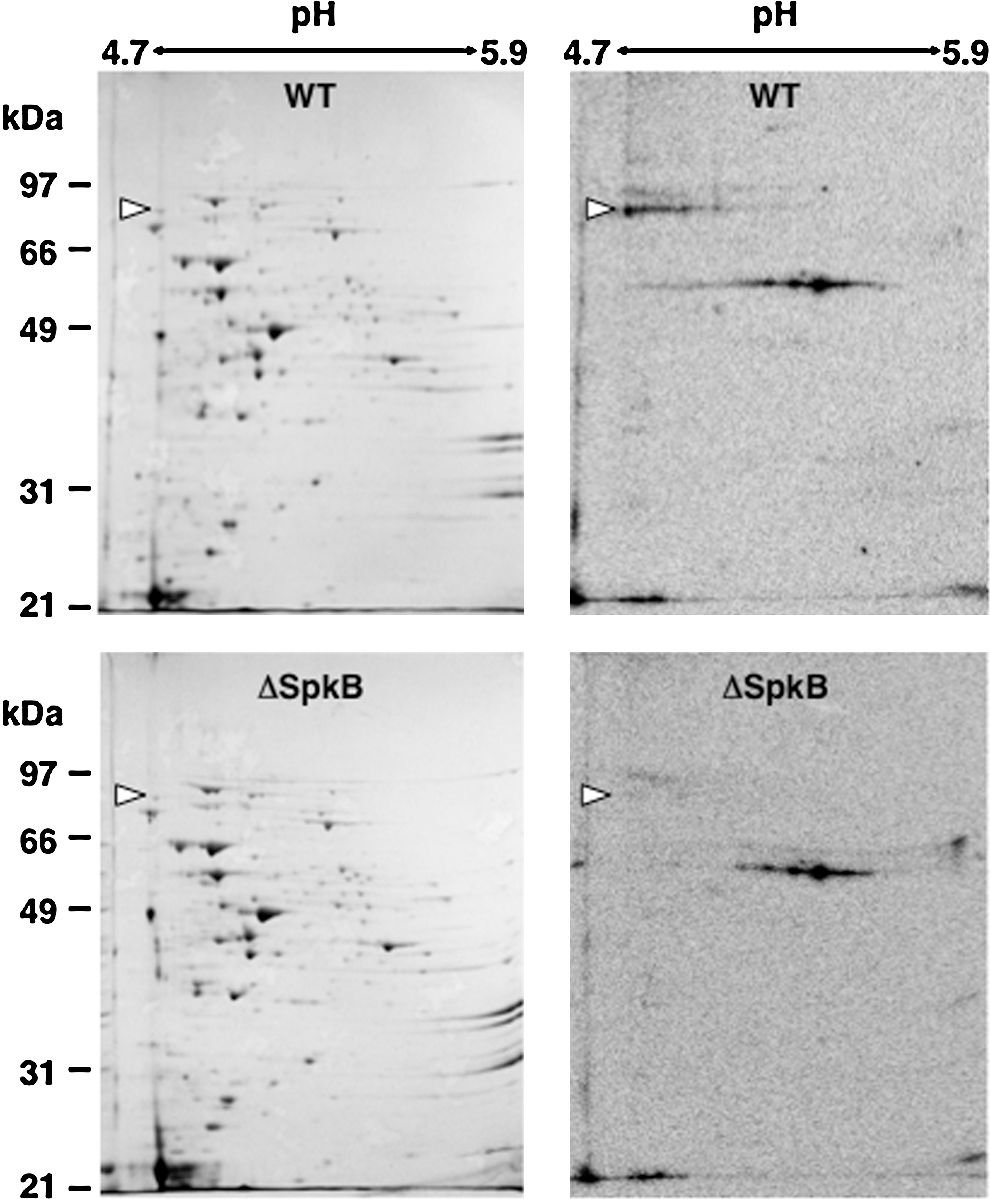
After establishing that GlyS does not become phosphorylated in the SpkB mutant strain, it remained to be determined whether SpkB could directly phosphorylate the GlyS protein. This issue was addressed using two different approaches. First, we added recombinant SpkB to cystosolic extracts from the ΔSpkB mutant (Fig. 8A). These experiments showed that the addition of 0.5 to 1 μg of recombinant SpkB to ΔSpkB samples resulted in the complete restoration of GlyS phosphorylation (Fig. 8A). Higher amounts of SpkB yielded the additional phosphorylation of a number of proteins previously undetected in our assays.

A heterotetrameric structure of glycyl-tRNA synthetases of the type α2β2 has been suggested to exist only in bacteria, and a dimeric structure of the type α2 can be observed mainly in eukaryotes (31). The α-subunit and parts of the β-subunit are required for aminoacylation of tRNA, and the α-chain contributes to amino acid- and ATP binding (10). Synechocystis possesses two genes that code for the α- (GlyQ) and β- (GlyS) subunits of glycyl-tRNA synthetase, respectively. Therefore, we decided to co-express histidine-tagged GlyS and GlyQ for purification. The phosphorylation assay based on radioactive labeling was performed using co-purified GlyS and GlyQ in the presence and absence of SpkB (Fig. 4B). The α-subunit of the glycyl-tRNA synthetase, GlyQ, was labeled in both the presence and absence of the kinase, probably as a result of its ATP-binding function. However, the β-subunit, GlyS, was labeled only in the presence of SpkB, thus confirming that it is able to undergo phosphorylation catalyzed by SpkB (Fig. 8B).
The SpkB null mutant is hypersensitive to oxidative stress
ΔSpkB grows slower on solid media than the WT Synechocystis strain under normal light conditions, whereas growth rates are comparable under low light conditions (Fig. 9A). The addition of 0.1 mM glucose exacerbated the ΔSpkB growth retardation under control light, while no effect of glucose was observed under low light (Fig. 9A). Given that strong illumination leads to high rates of PET and the concomitant production of ROS (see Ref. 9), these results suggest that ΔSpkB might be deficient in its response to oxidative stress. Furthermore, reducing equivalents derived from glucose enter the Synechocystis respiratory electron transport chain, which shares components with the PET chain (32), thus contributing to high overall electron transport rates. In order to analyze whether the observed effects could be the result of decreased oxidative stress tolerance, we examined the growth of ΔSpkB and WT strains on solid media, including menadione or methyl viologen. Menadione is a diquinone that stimulates the production of ROS (45). Methyl viologen enhances the production of superoxide anion radicals during oxygenic photosynthesis (18), which, in turn, leads to the formation of hydrogen peroxide and hydroxyl radicals. Strikingly, both compounds completely abolished the growth of ΔSpkB (Fig. 9A), confirming the hypersensitivity of ΔSpkB to oxidative stress.

In liquid cultures grown at different light intensities (Fig. 9B), no significant difference was observed at a low light intensity, whereas at a normal light intensity, ΔSpkB grew slower than WT Synechocystis. A further elevation of the light intensity caused an initial 2-day growth arrest of ΔSpkB (Fig. 9B). Iron deficiency leads to the accumulation of ROS in plants (44) and cyanobacteria (20). The ΔSpkB mutant proved to be even more sensitive to iron starvation than the WT (Fig. 9C).
Finally, ROS levels were directly monitored through CM-H2DCF-DA fluorescence labeling after treatments of WT and ΔSpkB cultures with methyl viologen (Supplementary Fig. S5). Interestingly, low concentrations of methyl viologen result in enhanced ROS levels only in the mutant, whereas higher concentrations of methyl viologen lead to higher ROS levels only in the WT (Supplementary Fig. S5). A possible explanation would be that the higher concentrations of methyl viologen are toxic to the ΔSpkB mutant even during the brief treatments applied.
Discussion
The knowledge on redox-active cysteines in phosphorylation-mediated bacterial signaling is just beginning to emerge. Thus, it was recently reported that the Salmonella enterica response regulator SsrB, substrate for the histidine kinase SsrA, is regulated through the S-nitrosylation of a critical cysteine residue (12). In this study, we found that protein phosphorylation in the cyanobacterium Synechocystis is sensitive to changes in the cysteine redox state through the redox modulation of a eukaryotic-like protein kinase. However, GSSG is not efficient as an inhibitor of protein phosphorylation, and GSH is not capable of restoring the activity of oxidatively inactivated protein kinases in Synechocystis. Therefore, it seems unlikely that the glutathione system would play a role in the regulation of protein phosphorylation in this organism. In contrast, TrxA-catalyzed reduction completely restores the activity of the oxidized inactive SpkB kinase in vitro, which makes this Trx a plausible candidate as a physiological reductant of SpkB. The fact that GlyS phosphorylation in a freshly prepared WT cell extract is not stimulated by the addition of DTT indicates that SpkB is fully reduced in the cell under normal conditions. Considering that the residual phosphorylation of GlyS in the ΔSpkB mutant is still sensitive to Cu2+, redox-sensitive protein kinases that are unidentified as yet may also participate in this process. The truncated mutant β-SpkB, devoid of the N-terminal Cys-motif was found to be inactive unless exogenous thiols, such as DTT or TrxA, were present in the assay. This suggests that the Cys-motif, which includes the 4 first cysteines, functions as a redox buffer which maintains another critical cysteine in the kinase reduced. This other cysteine residue, or cysteine residues, should be essential for the activity of SpkB, as the complete activation of β-SpkB is achieved on reduction. Which of the other four cysteines is involved in this redox regulation is presently unknown; however, sequence alignments of SpkB with homologs from other cyanobacterial species reveal that the 5th and 6th cysteines are conserved, whereas the 7th and 8th cysteines are not (Supplementary Fig. S3).
SpkB shares its domain architecture with homologs from many other cyanobacterial species (49). The C-terminal half of the protein presents numerous pentapeptide repeats (Supplementary Figs. S3 and S6), which consist of several repetitions of a 5-amino-acid motif, usually A(D/N)LXX. The function of this domain in proteins is not clear, though a role in protein-protein interaction has been suggested (46). Eukaryotic-like Ser/Thr protein kinases with putative redox regulatory N-terminal domains and potentially protein-binding C-terminal domains appear to be a recurrent theme among prokaryotes. Thus, a Ser/Thr protein kinase, pkn22, in the cyanobacterium Anabaena sp. PCC 7120 (21, 47), contains an N-terminal Cys-motif, a protein kinase domain, and a C-terminal β-helix repeat domain, a so-called PbH1 domain. The Mycobacterium tuberculosis Ser/Thr protein kinase PknG is a crucial virulence factor that is essential for survival within host macrophages (42). The crystal structure of PknG reveals an N-terminal rubredoxin-like domain, which contains two pairs of cysteines, the kinase domain and a C-terminal domain consisting of tetratricopeptide repeats (42). It remains to be explored whether these kinases might be redox regulated in a manner similar to Synechocystis SpkB. An encouraging observation is that a mutated version of PknG, with the cysteines of the rubredoxin domain exchanged for serines, is completely inactive (42). However, no attempts were made to reactivate this kinase and, thus, a redox regulation of PknG has not yet been confirmed.
A few phosphoproteins from cyanobacteria have been identified to date, and GlyS has not been previously reported as a cyanobacterial phosphoprotein. Nevertheless, an exhaustive analysis of the phosphoproteome of the gram-negative model bacterium E. coli led to the identification of 79 proteins phosphorylated on serine, threonine, or tyrosine residues and, notably, GlyS was one of the phosphoproteins identified in this study (28). The physiological role of GlyS phosphorylation in Synechocystis and other bacteria remains to be established. It should be kept in mind that some eukaryotic aminoacyl-tRNA synthetases exert functions other than the aminoacylation of tRNA, so-called “moonlighting,” and the phosphorylation-mediated release of these enzymes from multi-protein complexes has been suggested to favor their noncanonical functions (1, 38).
The Synechocystis SpkB null mutant showed drastically reduced oxidative stress tolerance. Therefore, we conclude that the most prominent role of SpkB is to promote the survival of this bacterium under conditions, such as elevated light intensities, which lead to enhanced ROS production. However, we also show that SpkB, paradoxically, is inactivated by cysteine oxidation, which would imply that SpkB might be inactive under the conditions of greatest necessity. The explanation probably lies in the SpkB Cys-motif, acting as redox buffer, and, if this would fail, an operative thioredoxin system that allows SpkB to withstand moderate oxidative stress (Fig. 10). Thus, in WT cells, SpkB would be active and functional, except under severe oxidative stress conditions leading to cell death.
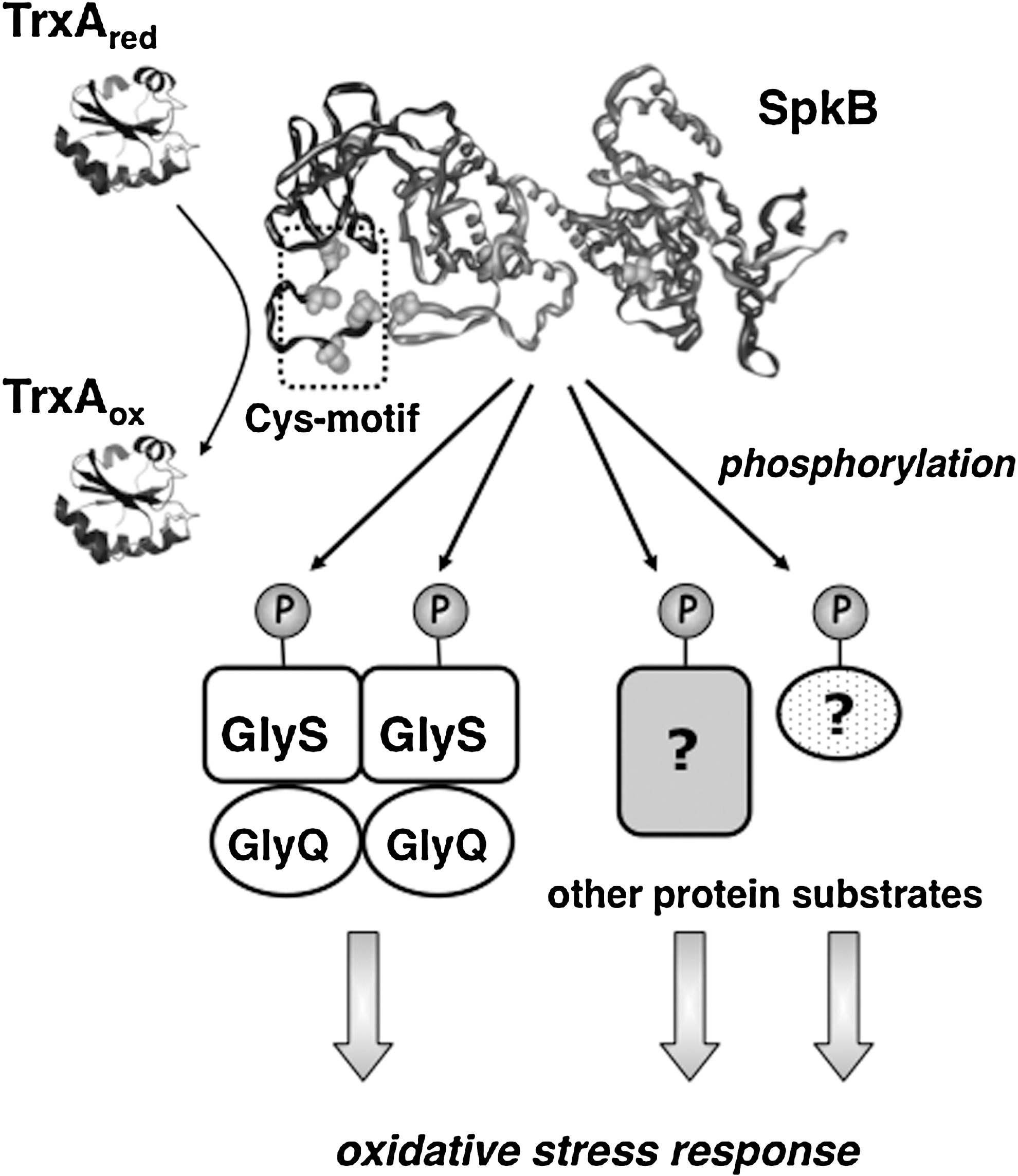
These studies show that the redox control of protein kinases is not unique to eukaryotes and raises the possibility that redox homeostasis might influence signaling through the modulation of protein phosphorylation also in other bacterial phyla.
Materials and Methods
Materials
The chemicals used in this study, such as DTT, GSH, GSSG, CuCl2, 2,2′-dipyridyl, menadione, and methyl viologen, were all obtained from Sigma-Aldrich (St. Louis, Missouri). The [γ-32P]ATP was from Perkin–Elmer (Walthman, Massachusets).
Cyanobacterial strains and growth conditions
Synechocystis sp. PCC 6803 cultures were photoautotrophically grown at 30°C under continuous illumination at an intensity of 50 μE m−2 s−1 (normal light), 10 μE m−2 s−1 (low light), or 120 μE m−2 s−1 (high light). The growth medium was BG11 (40) supplemented with 1 g/l NaHCO3 and bubbled with 1% (v/v) CO2 in air.
Cloning, expression, and purification
For the expression of proteins, the genes encoding SpkB (slr1697), GlyS (slr0220), and GlyQ (slr0638) were amplified from genomic DNA by PCR using gene-specific oligonucleotides, which included NdeI and XhoI sites for cloning into the pET28 vector (SpkB, β-SpkB, and GlyS) and NdeI and HindII sites for pET22 (GlyQ). The sequences of these oligonucleotides were as follows: SpkB, forward (5′-CTGGTGAACCCATATGAGTTTTTGCG-3′) and reverse (5′-GCTTGGTTTGGTCAGAAACACTCGAGATT-3′); β-SpkB, forward (5′-GTGGGCATATGCTGCGCCTC-3′); GlyS, forward (5′-GCTCCCTTGCCATATGCCCCTGC-3′) and reverse (5′-GGAGGCCTCGAGGACATTTAAAAC-3′); GlyQ, forward (5′-CTTGTTGGCTTCATATGACCATTACTTTCC-3′) and reverse (5′-GTTTTTAAATCAAGCTTTACGAGGGAAA-3′). The proteins were expressed in E. coli BL21 (DE3)-pLysS (Promega, Fitchburg, Wisconsin). The recombinant proteins were purified by nickel-affinity chromatography using the His-Bind resin (Novagen, Darmstadt, Germany).
Assays for kinase activity
The cells were harvested and broken with glass beads in a buffer containing 25 mM Hepes-NaOH (pH 7.6), 15% glycerol, 10 mM MgCl2, and 1 mM PMSF. Lysates were centrifuged at 16 000 g for 20 min. Protein concentrations of the cytosolic extracts were determined as described in (29). Cell extracts were incubated for 30 min at 25°C with a mixture composed of 0.5 mM ATP, 10 mM NaF, and 5 μCi [γ-32P]ATP. Kinase activities of recombinant SpkB and β-SpkB were assayed in vitro with [γ-32P]ATP by mixing 2 μg of purified kinase or 20 μg of E. coli extracts expressing SpkB or β-SpkB with 2.5 μg of casein or 10 μg of GlySQ as substrates. The radioactively labeled proteins were resolved in acrylamide protein gels, and the radioactivity was visualized using the Cyclone Plus Storage Phosphor System (Perkin-Elmer).
Two-dimensional electrophoresis
Protein samples were precipitated, washed by means of the commercial system 2-D Clean Up Kit (GE Healthcare, Little Chalfont, UK), and resuspended in DeStreak Rehydratation Solution (GE Healthcare) supplemented with ampholytes with a pH range of 3–10 (BioRad, Hercules, California) at 0.5%, following the manufacturer's instructions. The samples were separated in the first dimension using “DryStrips Immobiline Gel” (GE Healtcare) with a pH range of 4.7–5.9. The second dimension was performed using SDS-PAGE gels at 10% acrylamide concentration.
Footnotes
Acknowledgments
This work was financed by the Spanish Ministry of Science and Innovation (MICINN) through grants no. BFU2007-6300 and BFU2010-15708, co-funded to 70% by FEDER. A.M.-C. was the recipient of an FPU fellowship from the Spanish Ministry of Education and Science (MEC).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.