
Abstract
Prohibitin (PHB) is a highly conserved major sperm mitochondrial membrane protein whose absence in somatic cells is associated with mitochondrial membrane depolarization and increased generation of reactive oxygen species (ROS). Our recent findings suggest that high levels of oxidants in human semen may contribute to male infertility and that sperm motility could be the earliest and most sensitive indicator of oxidative damage. Based on PHB's roles in mitochondrial sub-compartmentalization and respiratory chain assembly, we examine sperm PHB expression and mitochondrial membrane potential (MITO) in infertile men with poor sperm motility (asthenospermia, A) and/or low sperm concentrations (oligoasthenospermia, OA). Here, we demonstrate that MITO is significantly lower in sperm from A and OA subjects than in normospermic (N) subjects; the decrease is more severe for OA than for A subjects. PHB expression is also significantly lower in sperm from A and OA subjects. Significantly positive correlations are found among PHB expression, MITO, and sperm motility in normospermic, asthenospermic, and oligoasthenospermic subjects. Collectively, our observations lead to the hypothesis that PHB expression is an indicator of sperm quality in infertile men, and that it regulates sperm motility via an alteration in MITO and increased ROS levels. Antioxid. Redox Signal. 17, 513–519.
Introduction
ROS, such as superoxide anions, hydroxyl radicals, and hydrogen peroxide, are free radicals and have been shown to influence the structural integrity of spermatozoa and their functions, including motility, capacitation, and sperm–oocyte fusion (1). These functions of the sperm are particularly vulnerable to ROS, because the sperm plasma membrane is rich in polyunsaturated fatty acids and membrane-bound NADPH oxidase. When the fine balance needed to maintain the optimal level of ROS controlled by antioxidant factors in secretions of the male reproductive system is lost, then the sperm may experience oxidative stress with resulting sperm dysfunction, which leads to male infertility. Our previous studies showed that the male accessory sex gland secretions are the major source of antioxidant enzymes in the ejaculate and have the important function of protecting the integrity of sperm DNA from the oxidative stress experienced in the uterus. Moreover, our recent findings suggest that high levels of oxidants in human semen may contribute to male factor infertility (2). Thus, sperm motility could be the earliest and most sensitive indicators of sperm oxidative damage (8).
Innovation
Prohibitin (PHB) is a highly conserved major sperm mitochondrial membrane protein whose absence in somatic cells is associated with membrane depolarization and increased generation of reactive oxygen species (ROS). Our recent findings suggest that high levels of oxidants in human semen may contribute to male factor infertility and that sperm motility could be the earliest and most sensitive indicators of the oxidative damage of the sperm. In the present study, we further demonstrate that there are significant positive correlations between the mitochondrial PHB expression, mitochondrial membrane potential (MITO), and sperm motility, indicating that mitochondrial PHB could be used as an indicator of sperm quality in infertile asthenospermic and oligoasthenospermic subjects. We hypothesize that PHB's deficiency exerts its effect on sperm motility via its alteration of MITO and, consequently, elevated levels of ROS.
Mitochondria are the major source of ROS, and they also play key roles in ATP production and the maintenance of sperm motility (3). Approximately 22–28 mitochondria are helically arranged around the mid-piece axoneme of mature sperm, and it is notable that the expression of PHB mRNA and PHB protein has been identified in spermatogenic cells of the rat during germ cell development. The protein has also been found to be expressed as an unusual high-molecular-mass isoform in spermatids and spermatozoa of bulls and rhesus monkeys and was shown to be one of the ubiquitinated substrates that mediate the rapid degradation of the paternal sperm mitochondria after fertilization (9).
The human PHB gene has been localized to 17q21, a region that contains a suspected human breast cancer tumor suppressor gene and is associated with various types of cancer. However, little is known regarding the functional significance of mitochondrial PHB in human male germ cells. Based on the roles of PHB in the functional compartmentalization within the mitochondria and in the assembly of the respiratory chain, we speculated that the expression of sperm PHB could be altered in a subset of men with infertility resulting from poor sperm motility (asthenospermia, A) and/or low sperm concentration (oligoasthenospermia, OA).
In the present study, we investigated sperm motility in semen samples from subjects attempting intra-cytoplasmic sperm injection (ICSI)/in vitro fertilization (IVF). Semen parameters were analyzed according to the 2010 World Health Organization (WHO) standards as shown in Table 1, and the mitochondrial membrane potential (MITO), the localization, and expression of the PHB in the sperm were studied. Then, correlations between PHB expression, MITO, and sperm motility were analyzed. Positive correlations reveal that PHB could be used as an indicator of sperm quality in infertile asthenospermic and oligoasthenospermic subjects.
N, normospermic; A, asthenospermic; OA, oligoasthenospermic subjects.
MITO Significantly Positively Correlated with Total Motility and Progressive Motility in Human Sperm
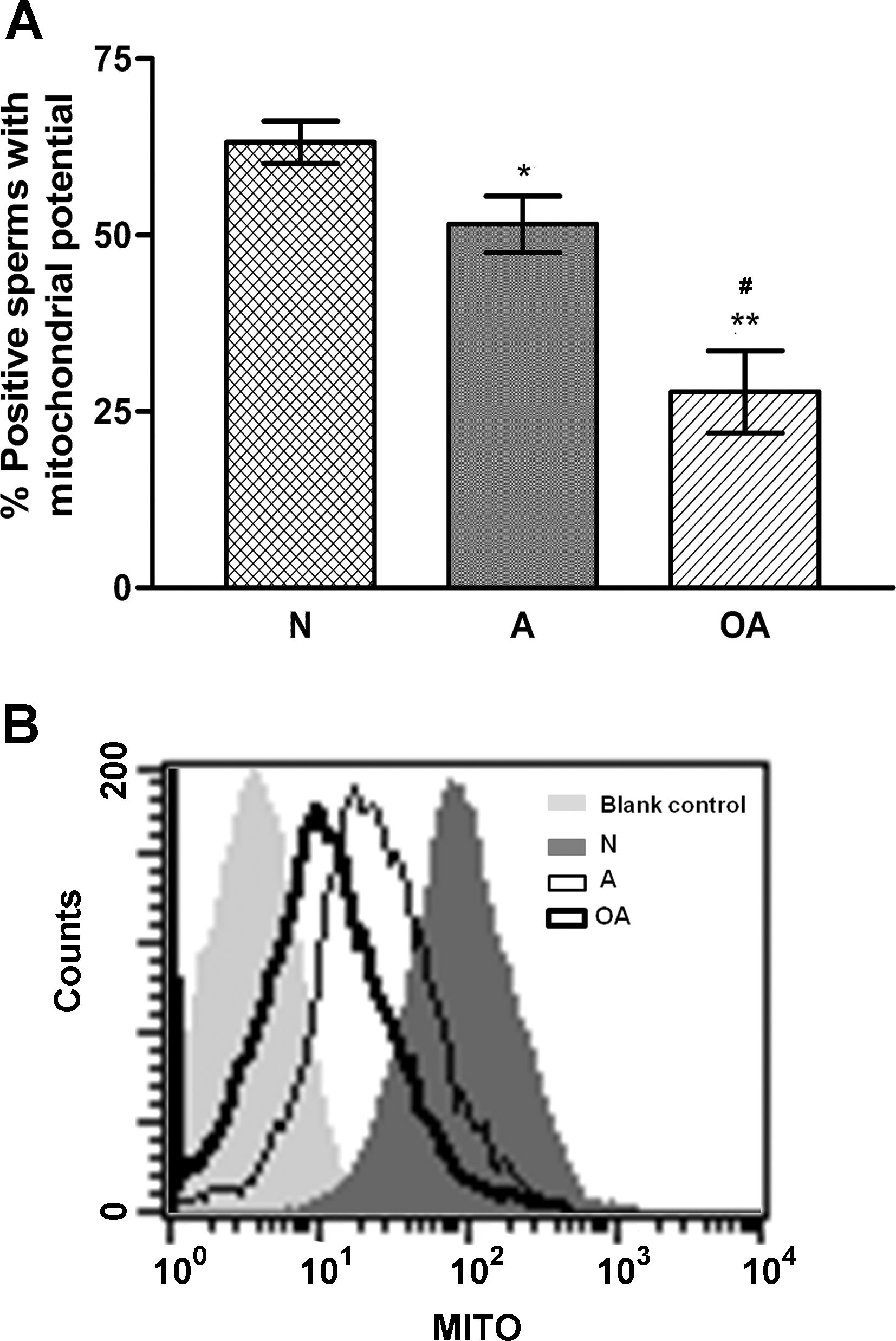
To assess the MITO of the human sperm, the percentage of positive sperm with various fluorescence intensities was scored using flow cytometry and correlated with the sperm motility using linear regression. As shown in Figure 1, the sperm MITO, as indicated by the percentage of positive cells with various fluorescence intensities, was significantly lower in the sperm from A (49.58±4.0; p<0.05) and OA (26.91±5.8; p<0.001) subjects than in the sperm from N (63.23±3.0) subjects. The decrease in the potential was more severe for the OA subjects than for the A subjects (p<0.01).
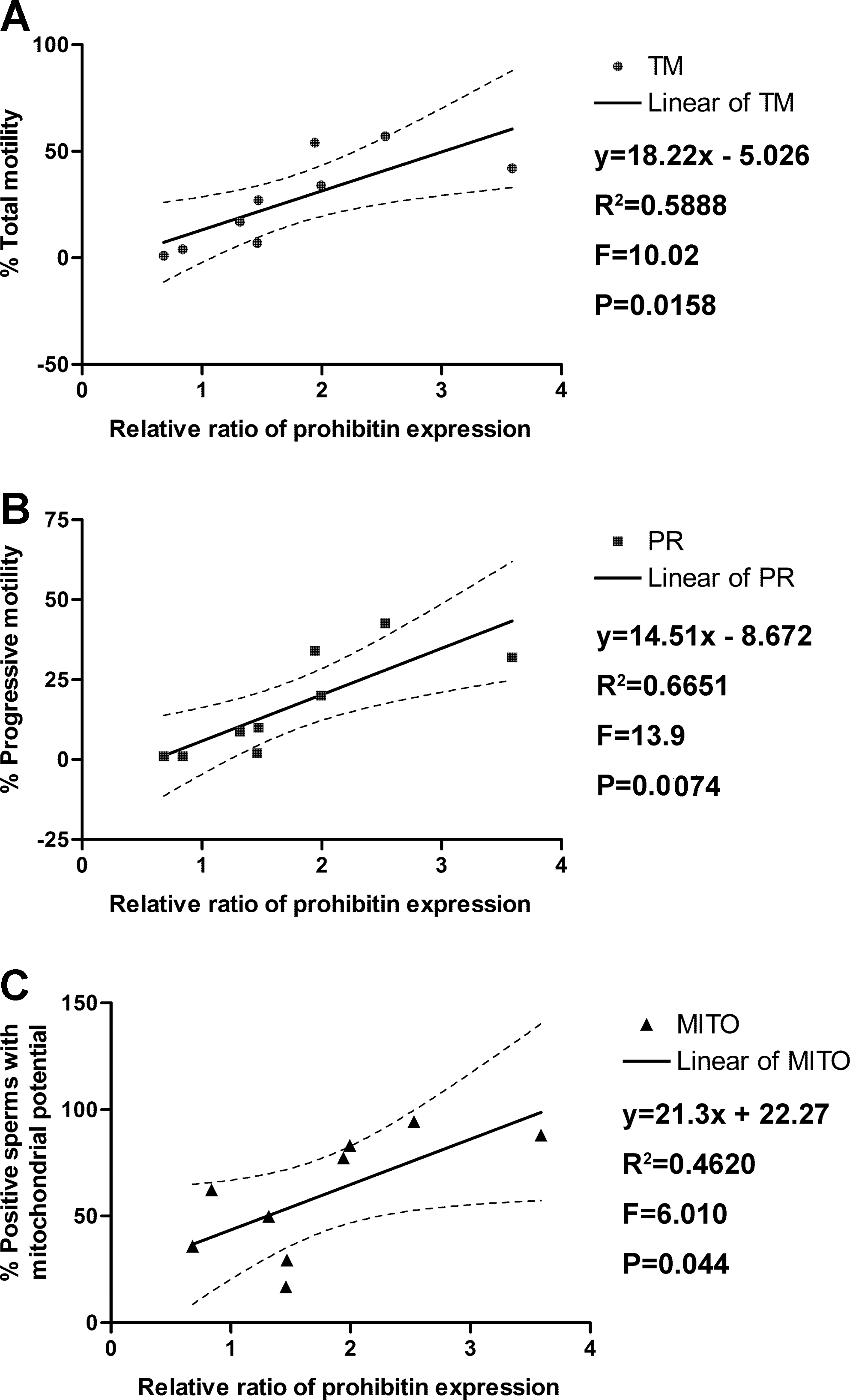
The correlation analysis showed that the MITO was positively correlated with the total sperm motility (Spearman r=0.3160, p=0.0087; see Fig. 2A) and the progressive motility (PR) (Spearman r=0.3219, p=0.0074; see Fig. 2B). Moreover, there was a significant linear relationship between the MITO and both the total motility (TM) and the PR of the sperm (see Fig. 2A, B; p<0.01).

PHB Co-Localized with MITO Probe in the Mitochondrial Sheath of Human Sperm
Figure 3 shows that the MitoTracker® Red probe (in red) was localized to the mid-piece of the sperm tail, which is the site of the mitochondrial sheath. The sperm of the N subjects exhibited an MITO staining in the typical rod-shaped mid-piece structure (Fig. 3, top panel); whereas the sperm of the A subjects, in the shorter or coiled mid-piece structure; while the sperm of the OA subjects, in the distorted mid-piece structure.

PHB, indicated by the green fluorescence (Fig. 3, middle panel), co-localized with the MitoTracker MITO staining of the mitochondrial sheath (red probe), and the yellow fluorescence indicates the regions in which both of the probes overlapped (Fig. 3, lower panel). The sperm nuclei were counterstained with 4′, 6-diamidino-2-phenylindole (DAPI). As shown in Figure 3, PHB (green) co-localized in the mitochondrial sheath (red) with the typical rod-shaped, shorter, or coiled mid-piece (yellow). However, in the distorted mid-piece of the sperm from the A and OA subjects, PHB expression was reduced or missing. Consequently, when images with its green staining were merged with those with the red staining of the MITO, the color remained red rather than yellow.
PHB Expression Significantly Positively Correlated with TM, PR, and MITO in Human Sperm
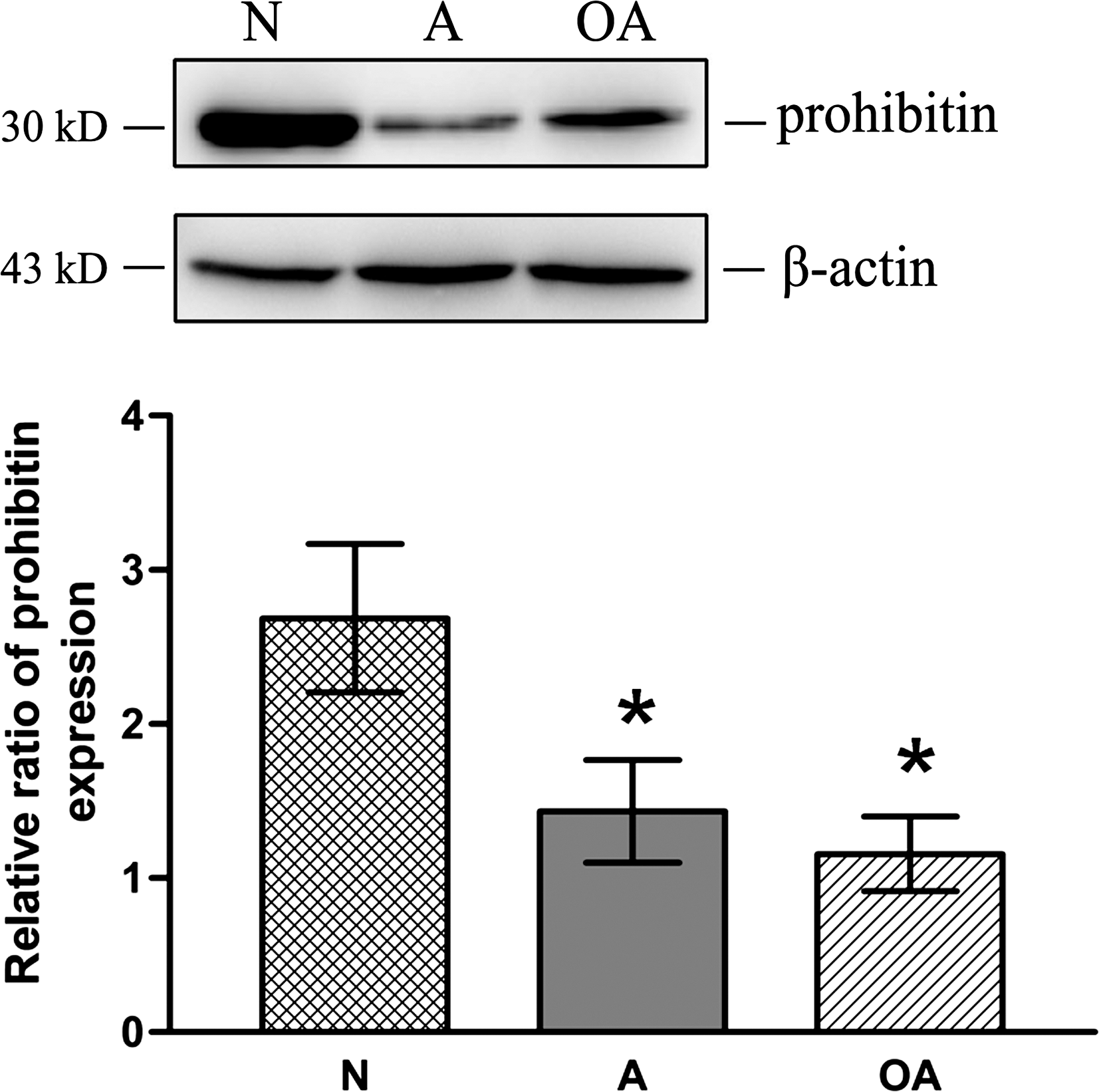
The level of PHB expression was significantly lower in the sperm from the A (0.5-fold of N) and OA (0.45-fold of N) subjects in comparison to the sperm from the N subjects (see Fig. 4; p<0.05). This lower expression of PHB in the A and OA subjects, detected by western blots analysis, confirms the lower intensity of PHB staining revealed by immunofluorescence assays in Figure 3.
Correlation analysis indicated that PHB expression was positively correlated with the total sperm motility (Spearman r=0.9, p=0.0020), the progressive sperm motility (Spearman r=0.8954, p=0.0020), and the MITO (Spearman r=0.7167, p=0.0369). Moreover, there were significant linear relationships between PHB expression and the total sperm motility, progressive sperm motility, and MITO (see Fig. 5, p<0.05).
Conclusions and Open Questions
In somatic cells, the absence of PHB, a major sperm mitochondrial membrane protein, is associated with depolarization of the mitochondrial membrane and increased generation of ROS (7). Recently, we reported that high levels of oxidants in human semen may contribute to male factor infertility (2) and that sperm motility could be the earliest and most sensitive indicator of the oxidative damage of the sperm (8). In the present study, we further demonstrate significantly positive correlations between mitochondrial PHB expression, MITO, and sperm motility, indicating that mitochondrial PHB could be used as an indicator of sperm quality in infertile asthenospermic and oligoasthenospermic subjects.
A primary cause of male infertility is decreased sperm motility which is a major indicator of semen quality. In the present study, sperm of normospermic (N), asthenospermic (A), and oligoasthenospermic (OA) subjects differed significantly with regard to their TM, PR, and concentration (Table 1). Significantly lower MITOs were found in the sperm from the A (p<0.05) and OA (p<0.001) subjects when compared with the normospermic (N) subjects (Fig. 1). Further, MITO was significantly positively correlated with the sperm motility (Fig. 2A, B). These results agree with the recent report of Marchetti et al. (4), who showed that the MITO is positively correlated with standard semen parameters such as motility.
It should be noted that in the present study the presence of a shorter or coiled mid-piece with an MITO similar to the rod-shaped mid-piece of the sperm from the N subjects was observed in that from the A and OA subjects (Fig. 3). These findings suggest that the sperm with the shorter or coiled mid-piece could have an MITO similar to that with the rod-shaped mid-piece. In this regard, Piasecka and Kawiak (6) have also reported that sperm mitochondria can be functionally active with a high membrane potential in some cases of asthenospermia. Piasecka and Kawiak (6) suggested that the low sperm motility did not necessarily result from energetic disturbances in the sperm mitochondria, but may instead be associated with deformations of the mitochondrial sheath.
A possible link between the structural integrity and the functionality of sperm mitochondria in infertile men may be the mitochondrial PHB complex. PHB is localized to the inner mitochondrial membrane, and plays important roles in the maintenance of mitochondrial biogenesis and metabolism (5). Our finding, which in Figure 3 reveals that sperm PHB is localized in the mitochondrial sheath, is consistent with its role in mitochondrial metabolism and ATP production (3). Notably, we also detected that mitochondrial PHB co-localized with the MITO probe in the rod-shaped and short mid-piece of the sperm but not in the ones with the curved mid-piece as shown in Figure 3. Our study, therefore, suggests, as reported by Piasecka and Kawiak (6), that the MITO in the sperm is directly related to the expression of mitochondrial PHB, which renders the mitochondria functional (5).
Significantly, in Figure 4, we demonstrate using western analysis and densitometry that the expression of mitochondrial PHB is significantly lower in the sperm from the A and OA subjects than in the sperm from the N subjects (p<0.05), confirming the immunofluorescence data. This alteration in PHB expression in A and OA subjects could, therefore, impact both the structure of the mitochondrial sheath as well as ATP production and the level of ROS, as mitochondria are the major source of ROS. Moreover, significantly positive correlations were found among PHB expression levels (p<0.05), MITO, (p<0.01) and sperm motility (p<0.01), as shown in Figure 5A–C.
Thus, we propose that the decrease in PHB in the sperm may alter the mitochondrial structure, ultimately resulting in a decrease in the MITO and sperm motility. Moreover, our recent findings (8) that sperm motility is the most sensitive indicator of oxidative damage of the human sperm exposed to oxidative stress induced by the increased ROS levels due to endogenous inflammation may also be explained by the significantly lower level of PHB expression. The latter then leads to the depolarization of the mitochondrial membrane and the increased generation of ROS (7).
In conclusion, the present study demonstrates that there are significant positive correlations between mitochondrial PHB expression, MITO, and sperm motility, leading to the hypothesis that mitochondrial PHB expression could be used as an indicator of sperm quality in infertile asthenospermic and oligoasthenospermic subjects, and that it regulates sperm motility via an alteration in MITO and increased ROS levels.
Notes
Materials and Methods
Chemicals
All chemicals were purchased from Sigma Chemical Co., unless otherwise stated. Phosphate-buffered saline (PBS) solution was purchased from Ambion Inc. MitoTracker mitochondria Red CMXRos probe and Alexa Fluor® 488 goat anti-rabbit IgG antibody were from Molecular Probes Inc. Normal goat serum (NGS) and VECTASHIELD® Mounting Medium with DAPI were from Vector Laboratories Inc. The rabbit polyclonal anti-human PHB antibody was from Abcam Inc. The horseradish peroxidase (HRP)-conjugated monoclonal mouse anti-β actin antibody and the secondary goat anti-rabbit IgG antibody were from Santa Cruz Biotechnology, Inc. 3-([3-Cholamidopropyl] dimethylammonio)-1-propane sulfonate (CHAPS), urea, thiourea, dithiothreitol (DTT), acrylamide, and phenylmethanesulfonyl fluoride (PMSF) were purchased from Amersham Biosciences. All the buffers were prepared using Milli-Q water (Millipore).
Sample collection and preparation
The project was approved by the ethics committee of the Fudan University for Investigations in Humans. After informed consent had been obtained, the semen samples were collected according to the WHO criteria from 68 male subjects between 30 and 40 years of age who were attempting ICSI/IVF at the Shanghai Ji'Ai Genetics and IVF Institute of the Fudan University. Semen samples were collected from 37 normospermic, 23 asthenospermic, and 8 oligoasthenospermic subjects (Table 1). After liquefaction at 37°C for 30 min, the collected samples were subjected to sperm concentration, motility, and morphology analyses by optical microscopy according to the 2010 WHO criteria as shown in Table 1.
The experiments, including flow cytometry and immunofluorescence, were performed after the semen samples had been washed twice with PBS, followed by centrifugation at 300 g for 10 min at room temperature.
Flow cytometry
For the detection of the MITO, the samples were processed using flow cytometry after MitoTracker Red probe staining of live sperm according to the manufacturer's instructions, with some modifications. Briefly, after washing twice with PBS and centrifuging at 300 g for 10 min at room temperature, the sperm suspensions (1.0×106 sperms/ml) were incubated with a pre-warmed staining solution of 250 nM MitoTracker mitochondria Red CMXRos probe at 37°C for 30 min. The blank control did not include the MitoTracker probe. Then, the sperm pellets were washed twice with fresh, pre-warmed PBS, followed by centrifugation at 300 g for 10 min at room temperature. The pellets were then resuspended in fresh, pre-warmed PBS for the flow cytometry analysis or the subsequent immunofluorescence staining.
For flow cytometry analysis, 4.0×104 events for each sample, including the blank control, were recorded using an FACScan® flow cytometer set at 568 nm. The data were analyzed using CellQuest™ software (BD Biosciences), and dot plots and scatter diagrams were generated to analyze the results. The percentage of the cells that were positive for the MitoTracker mitochondria Red CMXRos probe was determined relative to the staining of the blank control.
Immunofluorescence
To detect the co-localization of PHB with the MitoTracker mitochondria Red CMXRos probe in the sperm from the N, A, and OA subjects, the sperm suspensions (1.0×106 sperms/ml) remaining after flow cytometry analysis were fixed in 4% paraformaldehyde (pH 7.4) for 15 min at room temperature. After washing twice with PBS for 15 min, the sperm were permeabilized with 2% Triton in PBS for 15 min and then washed thrice for 15 min with a washing buffer that contained 0.2% Tween 20 and 1% NGS in PBS. Then, the sperm were blocked for 40 min in PBS with 5% NGS and washed again. The samples were then split into two aliquots. One aliquot was incubated at 4°C overnight with a rabbit polyclonal antibody raised against human PHB, diluted 1:50 in the antibody diluent that consisted of PBS containing 0.5% Tween 20 and 1% NGS. The other aliquot was used to prepare negative controls without the primary antibody or with an isotype for rabbit IgG. After washing twice with the buffer, the sperm were incubated in the dark for 2 h with Alexa Fluor 488 goat anti-rabbit IgG, diluted 1:2000 in the antibody diluent. Thereafter, the samples were washed twice, resuspended in 50 μl PBS, smeared on slides, and mounted using VECTASHIELD Mounting Medium with DAPI. After sealing the slides with nail polish, the green fluorescence was detected at an excitation wavelength of 488 nm, and the red fluorescence was detected at 561 nm using a confocal Leica TCS SP5 scanning microscope equipped with argon and He/Ne Green laser sources for the green and red fluorescence measurements, respectively. The emitted fluorescence was detected using a Leica Plan Apo×63 oil immersion objective, and the images were captured and processed using Leica Application Suite Advanced Fluorescence software version: 1.8.1 build 1390 (LAS AF Lite, Leica Microsystems CMS GmbH).
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis and western blotting
The level of PHB expression in the sperm from the N, A, and OA subjects was measured by western blot analysis. Briefly, after washing twice with PBS, 10×106 sperm were lysed in 200 μl cold lysis buffer (7 M urea, 2 M thiourea, 4% CHAPS, 65 mM DTT, 1 mM PMSF, and 1× proteinase cocktail) using sonication on ice four times for 10 s, with each sonication period separated by 15 s; the lysates were homogenized on ice for 30 min before centrifugation (16,000 g, 30 min, 4°C). The total protein concentrations of the lysates were then determined using the Bradford method (Pierce Biotechnology) before the one-dimensional gel electrophoresis (Bio-Rad Life Science). After being mixed with 5× sample loading buffer and boiling for 5 min, aliquots of the cell lysates (equivalent to 50 μg of total protein) were analyzed using 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis. The separated proteins were transferred to a polyvinylidene fluoride membrane (Millipore), which was then blocked with 5% nonfat milk in TBST buffer (20 mM Tris, pH 8.0, 150 mM NaCl, and 0.1% Tween-20) for 2 h at room temperature. The membranes were incubated with the primary antibody against PHB at a 1:1000 dilution overnight at 4°C and then with an HRP-conjugated secondary antibody at a 1:5000 dilution for 1 h at room temperature. The proteins were detected using an enhanced chemiluminescence kit (Amersham Biosciences) according to the manufacturer's instructions, and the images were captured using the LAS-3000 image analyzer (Fujifilm). The protein molecular weights were estimated using the Prestained Protein Ladder (Invitrogen). β-Actin was used as a loading control. The relative ratio of PHB protein versus β-actin was analyzed using densitometry.
Statistical analysis
All the results are presented as the mean±standard error of the mean. The distributions of all of the variables were confirmed to be normal before the statistical analysis (Prism software version 3.0; GraphPad). All the results, including the percentages of the sperm with different fluorescence intensities after MitoTracker Red probe staining and the relative density ratios of PHB expression in the N, A, and OA subjects, were analyzed by one-way analysis of variance, followed by the Newman–Keuls post-test. The relationships among the percentage of sperm stained with the MitoTracker Red probe, the relative density ratio of PHB expression, and the sperm motility (total or progressive) of the samples from these subjects were analyzed by Spearman's correlation and linear regression. p-values of <0.05 (two-sided probability) were considered statistically significant. Each experiment was repeated at least five times.
Footnotes
Acknowledgments
The authors gratefully acknowledge Drs. Liming Wei and Shuhui Sun for their help and advice regarding the western blotting and flow cytometry experiments. This project was funded by grants from the National Natural Science Foundation of China (No. J0730860 and No. 31071053).
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.