
Abstract
Introduction
By mediating electron donation, disulfide formation, hydrolysis, metal binding, and redox catalysis and sensing, the sulfur in cysteines can fulfill a wide variety of molecular functions (33, 46). Bacterial redox sensors of various reactive species utilize cysteines because of their ability to bind metals and adopt various oxidation states. Most of the regulators presented in this review rely on the redox-sensing activity of the free thiolate side chain of cysteines. This review also discusses the mechanisms by which redox active cysteines in zinc fingers help bacteria sense ROS. By analogy to other metalloproteins, the cysteines in zinc fingers provide a scaffold where a structural metal cation is assembled. In addition to serving a structural function, redox active cysteines in zinc fingers transmit signals in response to ROS. Free cysteine side chains and cysteines bound in zinc fingers are common strategies of thiol-dependent bacterial redox sensors of ROS and RNS. To learn how cysteines indirectly sense oxidative and nitrosative stress through ligation of redox active metal prosthetic groups, the reader is directed to the accompanying reviews in this forum.
The contribution of thiol-dependent signaling to the resistance of bacteria to oxidative stress was first hinted at in investigations with the frank pathogen Salmonella enterica serovar Typhimurium (20). Many investigations on thiol switches have been carried out in commensal Escherichia coli and saprophytic environmental bacteria of the species Bacillus subtilis and Streptomyces coelicolor [see (1, 77) for recent reviews]. A growing number of Gram-positive and Gram-negative pathogens are now known to apply the signaling properties of thiol switches in order to increase fitness during their associations with vertebrate hosts. The mechanisms by which thiol-dependent redox sensors respond to ROS and RNS can be broadly categorized within families of transcriptional regulators, irrespective of whether they are expressed by commensal, saprophytic, or pathogenic bacteria. Consequently, the mode of action by which active thiols sense reactive species is presented herein within the context of families of transcriptional regulators, and their contributions to bacterial pathogenesis are highlighted. A section in this review also discusses the structural and chemical determinants that impact the sensing activity of cysteines. The last section presents the dominant chemistry driving the interaction of ROS and RNS with redox active cysteines.
Sensing of Oxidative and Nitrosative Stress by Members of the LysR/OxyR Family of Transcriptional Regulators
The first indication that bacteria adapt to oxidative stress was the observation that the exposure of Salmonella to sublethal concentrations of hydrogen peroxide (H2O2) elicits hyperresistance to a subsequent challenge with high amounts of this ROS (20). OxyR was identified as a member of the LysR family of transcriptional activators after mapping of an interesting transposon mutant named oxyR2 (21). The critical role that the oxyR locus plays in the adaptation of E. coli to oxidative stress was demonstrated by the fact that OxyR-deficient bacteria are readily killed by H2O2 (20, 21). In addition to regulating antioxidant resistance, OxyR helps coordinate the expression of antinitrosative defenses (38). OxyR is less responsive to nitrogen oxides than to H2O2, especially if the bacterial cytoplasm is rich in glutathione (GSH) (38, 111). Nonetheless, E. coli oxyR mutants are hypersusceptible to the bacteriostatic effects of nitric oxide (NO) (38). Why is it that a highly specialized H2O2 sensor such as OxyR can also be modulated by RNS? Nitroxidative stress may explain this conundrum. Most of the biological chemistry of NO is dominated by oxidative processes (61). Taking into consideration this notion, the OxyR-regulated GSH oxidoreductase, glutaredoxin (Grx), thioredoxin (Trx), catalase, and alkyl hydroperoxide reductase re-establish thiol homeostasis and detoxify a variety of peroxides (9, 45, 71), independently of whether the bacterial cells are exposed to oxidative or nitroxidative species.
OxyR is a redox active regulator that maintains thiol homeostasis
The exposure of Salmonella to sublethal concentrations of H2O2 induces the expression of about 30 proteins (20). Hydroperoxidase I, alkyl hydroperoxide reductase, the nonspecific DNA-binding protein Dps, Grx1, GSH oxidoreductase, Trx 2, and the DsbG disulfide isomerase are some of the gene products positively regulated by oxidized OxyR (21, 44, 111). The first two enzymes detoxify H2O2, organic hydroperoxides, and peroxynitrite (ONOO−), while the iron-binding Dps protein protects DNA from Fe++-catalyzed, H2O2-dependent Fenton chemistry (9, 37, 83). GSH oxidoreductase, Grx1, DsbG disulfide isomerase, and Trx 2 re-establish thiol/disulfide homeostasis and protect the cell against disulfide stress. Thiol/disulfide exchange systems exert feedback regulation over oxidized OxyR, as suggested by the fact that OxyR is more easily activated by ROS and RNS in E. coli strains deficient in GSH oxidoreductase or Grx1 (38, 111). Moreover, the intramolecular disulfide bond of OxyR is a substrate of thiol/disulfide exchange by Grx1 (111).
Mechanisms of sensing oxidative and nitrosative stress by OxyR redox active thiols
OxyR activates gene transcription after the intramolecular oxidation of strategically situated cysteines (Fig. 1). It was noted that oxidized OxyR drives katG transcription in vitro (90). Although both oxidized and reduced OxyR bind target genes, they do it with different conformations and outcomes. Oxidized, tetrameric OxyR cooperatively binds four adjacent major DNA grooves, whereas reduced OxyR binds two pairs of adjacent DNA grooves separated by a single helical turn (96). Oxidized OxyR binds the RNA polymerase and drives transcription; the reduced protein does neither.
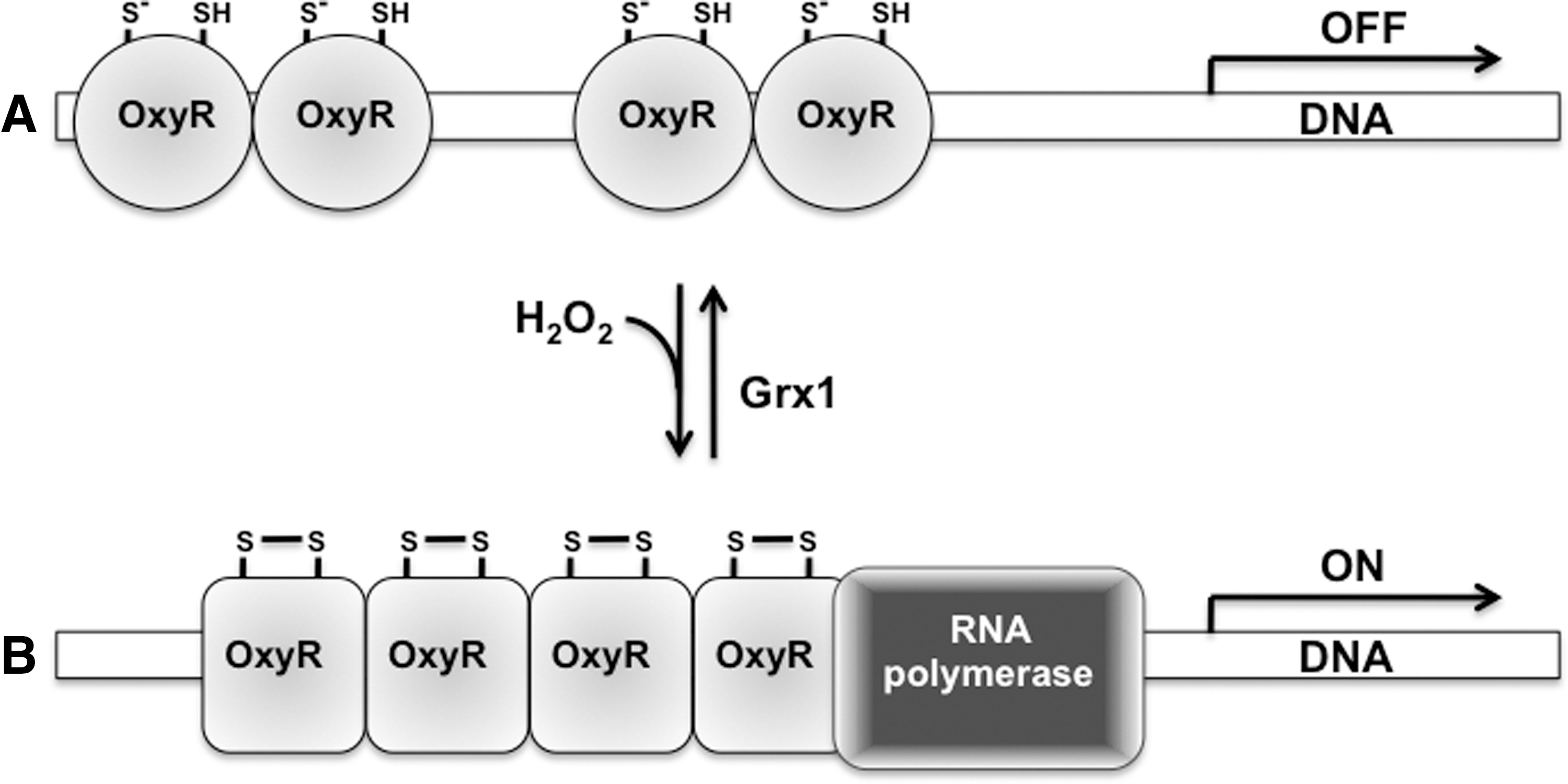
Oxidation of the thiolate side chain of OxyR Cys199 to a −SOH precedes the dramatic structural changes responsible for the activation of OxyR (111). The attack on Cys208 by the Cys199 sulfenic derivative generates an intramolecular disulfide that drives massive conformational changes in the protein (18, 63, 93, 111). Unexpectedly, mutational analysis indicates that Cys199 is more important than Cys208 for the activation of OxyR (111). The fact that a C208S substitution renders a partially constitutive active OxyR variant rationalizes the seemingly greater importance of Cys199 over Cys208 (63). Having a redox potential of −185 mV (111), Cys199 is maintained in the reduced form in the −280 mV potential of the cytoplasm of resting E. coli. However, these thermodynamic considerations cannot explain the rapid oxidation of OxyR in response to small rises in the intracytoplasmic concentration of H2O2. OxyR is oxidized within 30 s after E. coli is treated with 5 μM H2O2, a concentration of peroxide that does not appreciably change the ratio of the oxidized glutathione/GSH redox couple (2, 93). OxyR and H2O2 react with a second-order rate constant of about 105 M −1 s−1 (2). Therefore, kinetics rather than thermodynamics drives the fast oxidation of OxyR by H2O2. On the other hand, re-reduction of OxyR is a slow process that takes about 5–10 min (2, 93). The delay in the re-reduction of OxyR allows for the transient expression of antioxidant defenses encoded in the regulon.
In addition to undergoing oxidation in response to H2O2, OxyR gets S-nitrosylated in E. coli exposed to RNS (38, 55). Similar to the oxidized protein (90), S-nitrosylated OxyR drives katG transcription in vitro (38). In addition to S-nitrosylating OxyR, nitrogen oxides modify the Cys199 thiol side chain to sulfenylated and S-glutathionylated derivatives (55). All these covalent modifications affect the secondary α-helical structure of OxyR. These RNS-modified OxyR products are transcriptionally active; however, they differ in cooperativity, structure, DNA-binding affinity, and promoter activity. One of the most salient conclusions that can be drawn from these investigations lies in the realization that a redox active regulator does not have to be a simple switch that alternates between “ON” and “OFF” positions, but one whose activity can be fine tuned according to the nature of its post-translational modifications.
Contribution of LysR/OxyR regulators to bacterial pathogenesis
OxyR is dispensable for the pathogenesis of Mycobacterium tuberculosis and Sal. enterica. The M. tuberculosis oxyR gene has several inactivating nonsense and frameshift mutations, and oxyR-deficient Salmonella are completely virulent (25, 95). However, members of the OxyR regulon such as Dps are required for Salmonella virulence (37). dps and other members of the regulon are controlled by several regulators, which may explain the dispensability of OxyR in Mycobacterium and Salmonella. For example, the expression of dps in Salmonella is under the control of the ferric uptake regulator Fur (109), a gene product that is not only under OxyR regulation, but also essential for Salmonella pathogenesis (98, 104). One of the advantages of putting dps under the control of Fur is that its expression becomes responsive to iron levels as well as ROS and RNS (23, 94). Curiously, the OxyR-regulated KatG catalase capable of detoxifying high fluxes of H2O2 is dispensable for the survival of Salmonella in macrophages sustaining a respiratory burst (10). Several other catalases in the Salmonella genome may compensate for a lack of KatG (4).
In contrast to Salmonella and M. tuberculosis, OxyR augments the virulence potential of pathogenic microorganisms such as E. coli and Pseudomonas aeruginosa (47, 62). E. coli is an extraordinarily diverse species, encompassing common inhabitants of human and animal intestines as well as enterotoxigenic, enterohemorrhagic, and uropathogenic strains associated with high morbidity and mortality in humans and a variety of domestic animals. OxyR potentiates the ability of E. coli to cause ascending urinary tract infections (47). Curiously, OxyR-dependent antioxidant defenses do not antagonize the ROS produced by the NADPH phagocyte oxidase (47). Instead, OxyR may foster E. coli virulence by enhancing resistance to the reactive species generated during endogenous metabolic processes. According to this idea, alkyl hydroperoxide reductase, a member of the OxyR regulon, is quite effective at detoxifying low micromolar concentrations of H2O2, but it is inefficient against the large fluxes of H2O2 usually associated with inflammation (87). OxyR also augments the virulence of P. aeruginosa in rodent and insect models of infection (62). Future studies may reveal that other members of the growing LysR/OxyR family of transcriptional regulators do, in fact, foster colonization, invasion, and growth of many pathogenic microorganisms during their associations with vertebrate hosts. Recent evidence indicates that the redox-sensing activity of Cys235 in AphB, a member of the LysR/OxyR family of transcriptional regulators, is necessary for toxigenic Vibrio cholerae to colonize the anaerobic environment of the mammalian intestine (67).
Sensing of Oxidative Stress by Members of the MarA/OhrR Family of Transcriptional Regulators
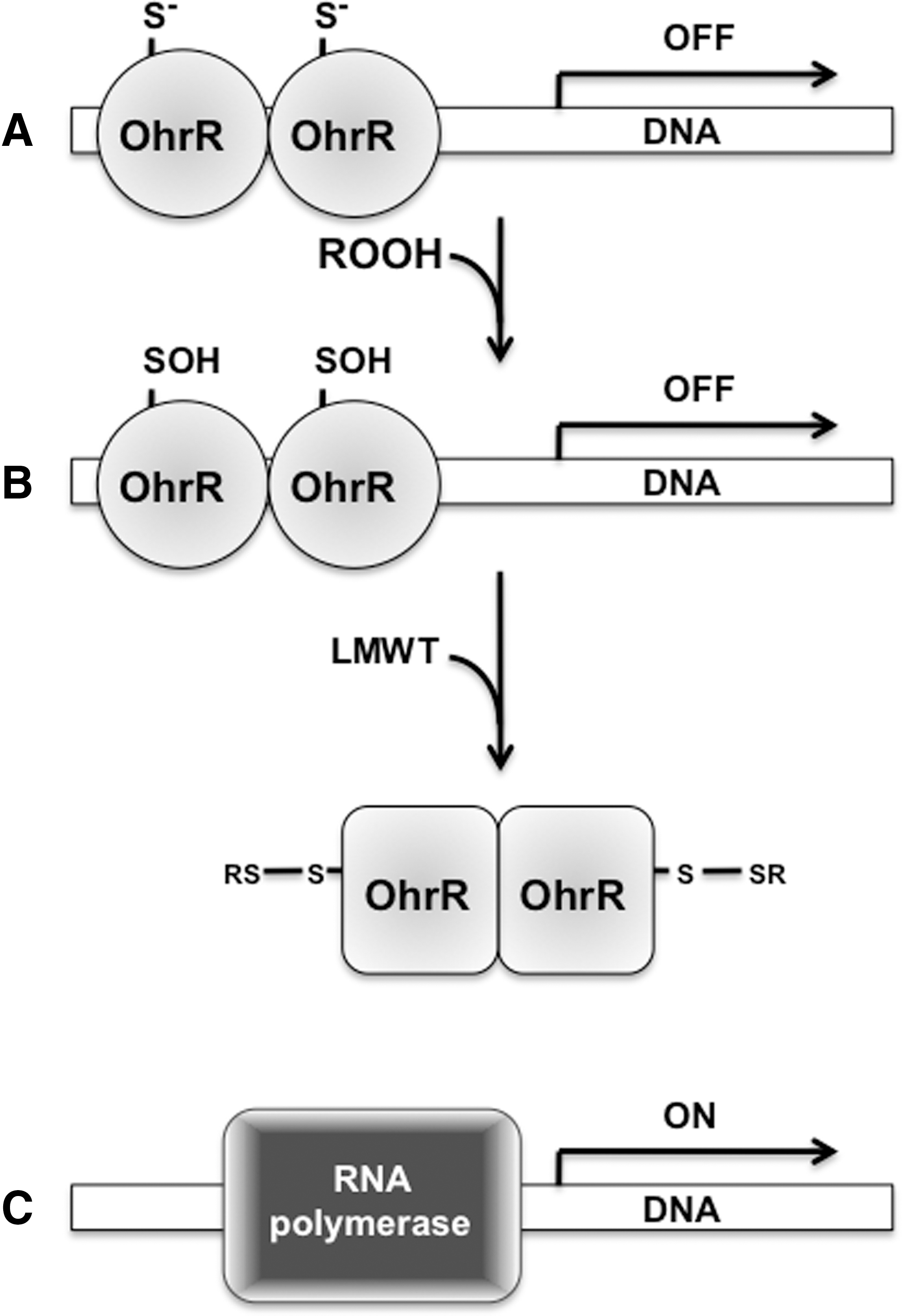
The DNA binding domain in the MarA/OhrR family of transcriptional regulators has a characteristic winged helix-turn-helix. A growing number of regulators in the MarA/OhrR family have been noted as containing one or two redox-active cysteines. OhrR homologs have been described in Staphylococcus aureus, Str. coelicolor A3(2), Xanthomonas campestris, and P. aeruginosa (3, 15, 72, 75, 92). A great deal of what we know about OhrR homologs has been unmasked while studying OhrR from the soil saprophyte B. subtilis. Reduced OhrR binds to the operator regions of target genes via a winged helix-turn-helix (Fig. 2), and, thus, represses the transcription of antioxidant defenses (31). The exposure of B. subtilis to organic hydroperoxides increases antioxidant defenses through the oxidation of OhrR (30).
Mechanisms of sensing oxidative stress by MarA/OhrR thiol switches
In Gram-positive bacteria, members of the MarA/OhrR family of transcriptional regulators sense oxidative stress through a lone cysteine residue, whereas their counterparts in Gram-negative bacteria involve two cysteines. A lone cysteine in B. subtilis OhrR, Sta. aureus SarZ, and Sta. aureus MgrA is oxidized by organic hydroperoxides to a remarkably stable −SOH (15, 31, 84). The formation of a −SOH is necessary but not sufficient to derepress transcription. In fact, reduced and sulfenylated B. subtilis OhrR and Sta. aureus SarZ bind with apparently similar affinities to the operator regions of target genes (64, 84). In B. subtilis, sulfenamide generated from the reaction of sulfenylated Cys15 with the amide group of the adjacent Phe16 may trigger the conformational changes required for the dissociation of OhrR from the operator regions (64). However, the OhrR sulfenamide derivative is far less abundant than an S-thiolated, mixed disulfide-bonded derivative (64). The −SOH in oxidized OhrR is prone to nucleophilic substitution with endogenous, low-molecular-weight thiols (LMWTs) (64, 84). Both mixed disulfides and sulfenamide are reversible modifications and, hence, the DNA binding activity of OhrR-like proteins can be restored on re-reduction. Together, these data unequivocally identify the redox-active thiol in members of the OhrR family of transcriptional regulators as a genuine signaling switch.
As just mentioned, the OhrR homologs in Gram-negative bacteria have two redox-active cysteines (13, 14, 80). In X. campestris, OhrR Cys22 is oxidized by organic hydroperoxides to −SOH, which rapidly reacts with Cys127 to form an intermolecular disulfide in the homodimer (80). As expected, disulfide-linked dimer formation drives the dissociation of X. campestris OhrR from DNA. P. aeruginosa MexR also has two redox-active cysteines (13). Organic hydroperoxides trigger the oxidation of MexR Cys30 and Cys62 to form two intermolecular disulfides with Cys62’ and Cys30’ of the homodimer (14). Disulfide formation is the driving force behind the depression of the mexAB-oprN drug efflux operon, and it increases the resistance of P. aeruginosa to antibiotics (14).
Contribution of the redox-sensing activity of the MarA/OhrR family members to bacterial pathogenesis
The contribution of the MarR/OhrR members to bacterial pathogenesis has been investigated in Sta. aureus and P. aeruginosa. A Sta. aureus sarZ mutant is attenuated in both silkworms and a 24-h mouse model of infection, but regains full virulence at later times of the infection (50). The transitory contribution of SarZ to Sta. aureus virulence probably reflects the complex SarZ regulatory network. SarZ regulates the expression of 87 genes in Sta. aureus (6, 16), including the repression of sarS and the induction of genes encoding exoproteins and cell wall biosynthesis. Decreased expression of these genes could contribute to the initial attenuation of sarZ-deficient Sta. aureus. However, by derepressing antioxidant defenses, the lack of sarZ increases the resistance of Sta. aureus to organic hydroperoxides and H2O2 (16). Moreover, an sarZ mutant forms denser biofilms, another aspect that may also foster Sta. aureus pathogenesis (92). In comparison to SarZ, MgrA plays a much greater and more durable role in Sta. aureus virulence, as measured by bacterial growth in viscera and the degree of sinovitis and arthritis affecting infected animals (48). The predominant role of MgrA over SarZ in the pathogenesis of Sta. aureus is not surprising, as MgrA regulates sarZ as well as more than 300 other genes (48, 69) such as those encoding α-hemolysixnα-toxin, capsular polysaccharide biosynthesis, coagulase, efflux pumps, leukotoxins, serine proteases, and sortase A.
In analogy to Sta. aureus, the opportunistic pathogen P. aeruginosa expresses two members of the MarR/OhrR family of transcriptional regulators, OhrR and OspR. A P. aeruginosa strain lacking ospR is hypervirulent in an acute pneumonia model of infection, because the derepression of GSH peroxidase enhances resistance to H2O2 (60). However, P. aeruginosa OhrR mutants are hyperresistant to organic hydroperoxides, but attenuated in a Caenorhabditis elegans model of infection (3). These findings indicate that the resistance to oxidative stress does not always equate to enhanced virulence. The overproduction of antioxidant defenses may, in turn, elicit reductive stress that leads to detrimental changes in gene expression.
Sensing of Oxidative Stress by Redox Active Thiols That Coordinate Zinc
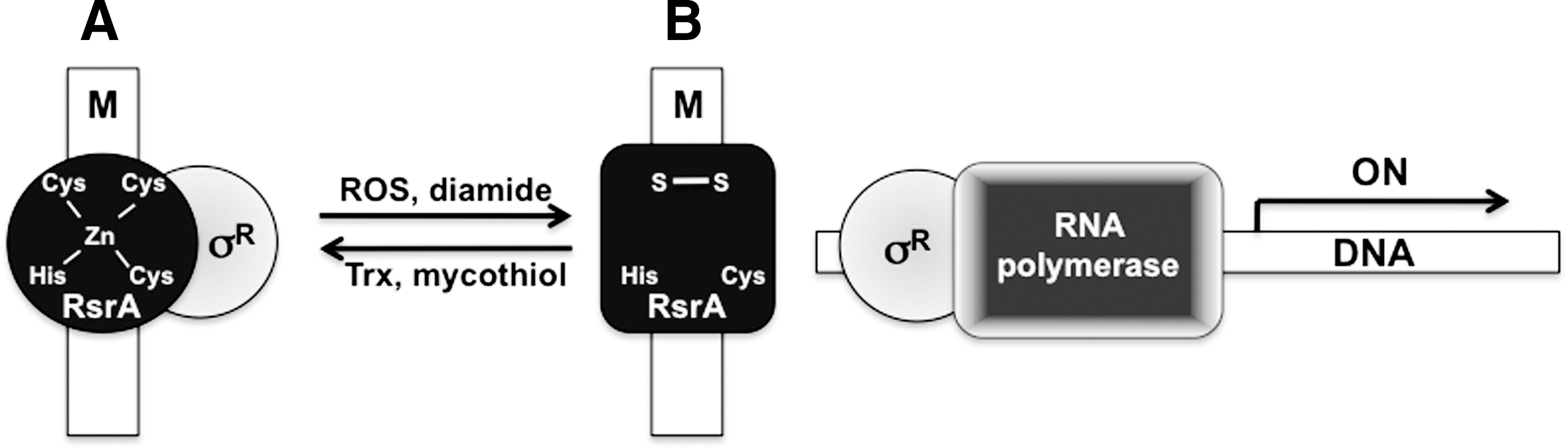
Most of the characterization of RsrA and the σR regulon has been done in the soil saprophyte Str. coelicolor A3(2). The σR regulon coordinates the response of Streptomyces to disulfide and oxidative stress associated with diamide, H2O2, and the superoxide (O2 •−) generator plumbagin (78), suggesting that members in this regulatory pathway are prone to redox control. The σR protein lacks cysteines. In contrast, its cognate antisigma factor RsrA has several cysteines, raising the possibility that redox active thiols in RsrA could mediate the response of the σR regulon to oxidative challenges (Fig. 3).
Regulation of antioxidant defenses by members of the σR family of extracytoplasmic function sigma factors
A functional σR is necessary for the proper expression of disulfide reductase activity in Str. coelicolor (78). Further analysis revealed that σR in fact regulates transcription of the trxAB operon encoding Trx 1 and Trx reductase, and the trxC-encoded Trx 2 (78, 79). A search of the Str. coelicolor genome for conserved σR binding sequences identified a total of 27 σR targets (79). These include the aforementioned trxC, and genes in the cysteine and molybdepterin biosynthetic pathways. σR also regulates the expression of msrA and msrB methionine sulphoxide reductase genes, the mshA-encoded mycothiol biosynthesis gene, and the mca-endoced amidase (81). The expression of these genes is consistent with the fact that the oxidation of RsrA derepresses σR-dependent antioxidant defenses. Oxidation of the cell also induces the accumulation of unfolded proteins in cytoplasmic aggregates (73). It is, therefore, not surprising that σR directly regulates Clp and Lon ATP-dependent AAA(+) proteases in order to eliminate damaged proteins (51). M. tuberculosis expresses σH, a σR homolog. Detailed transcriptional analysis indicates that σH regulates 31 genes in M. tuberculosis, including genes involved in Trx recycling, a Grx-like protein, ferredoxin, chaperones, and universal stress proteins (53, 70, 85). Therefore, it is becoming clear that actinomycetes use σR-like extracytoplasmic function sigma factors which regulate the transcription of classical antioxidant defenses and stress responses to unfolded proteins.
Mechanisms of sensing oxidative stress by the RsrA antisigma factor
The metalloprotein RsrA binds a single zinc cation with an affinity of 1017 M −1 (5, 76). The zinc metal is ligated by His37, Cys41, and Cys44 that form a characteristic HX3CX2C zinc-containing antisigma (ZAS) motif (49, 76, 110). Cys11 completes the tetrahedral coordination of zinc by RsrA (110). Genetic studies indicate that Cys11, Cys41, and Cys44 are essential for the antisigma activity of RsrA (76). Cys11 is the triggering residue that responds to oxidative stress, most likely by forming a −SOH intermediate that attacks the neighboring thiol Cys44 (5, 66, 76). The importance of Cys11 as the triggering residue responsive to oxidative stress is circumstantially supported by the fact that members of the ZAS family of transcriptional regulators that lack a cysteine at this position are indifferent to oxidative stress (66). In addition to Cys11-Cys44, a second disulfide forms between Cys41 and Cys61 (5). Disulfide formation releases zinc from RsrA (66). Nonetheless, disulfide bond formation, rather than the dissociation of zinc from the protein, increases the α-helical structure of RsrA that releases σR and allows it to transcriptionally activate target genes (66).
Similar to its RsrA homolog in Str. coelicolor, the M. tuberculosis RshA antisigma factor is responsive to oxidative stress (89). The binding of RshA to σH can also be modulated in response to metabolic changes in the cell. Thr94 is phosphorylated by the eukaryotic-like serine/threonine protein kinase PknB (36, 82). By analogy to the oxidized protein, phosphorylated RshA binds less efficiently to σH (82). The oxidation of thiols in the RshA zinc finger and phosphorylation of RshA Thr94 may facilitate the graded regulation of M. tuberculosis gene transcription in response to shifting redox and metabolic challenges.
RsrA is redox active
The oxidative modifications that mediate redox signaling are often reversible. Strains of Str. coelicolor deficient in mycothiol biosynthesis exhibit increased expression of σR, suggesting that decreases in intracellular mycothiol content could be the natural signal that drives the oxidation of RsrA and the consequent activation of the σR regulon (81). Both Trx and mycothiol limit RsrA disulfide bond formation, thereby stimulating the binding of this antisigma factor to σR (52, 81). On the other hand, σR activates the transcription of Trxs and the mycothiol biosynthetic pathway. These findings demonstrate that RsrA is a bona fide redox switch subjected to feedback regulation by members of the regulon.
Role of redox-active antisigma factors in bacterial pathogenesis
σH defends the human pathogen M. tuberculosis against both RNS generated from acidified nitrite, and ROS such as organic hydroperoxides, H2O2, and O2 •− (24, 28, 70, 85). σH is expressed by M. tuberculosis inside macrophages (34). However, this extracytoplasmic function sigma factor does not increase the intracellular fitness of mycobacteria (70), and M. tuberculosis σH mutants grow and persist with apparent normality in murine models of infection (53). Nonetheless, σH contributes to M. tuberculosis pathogenesis, and its expression affords M. tuberculosis the ability to recruit CD4+ and CD8+ T lymphocytes to well-formed granulomas in the lungs of infected mice (53). It remains to be investigated whether the immune response triggered by M. tuberculosis is related to the expression of σH-dependent antioxidant defenses.
Sensing of Nitrosative Stress by Members of the NarL/SsrB Family of Transcriptional Regulators
The response regulator SsrB is the only member of the NarL/FixL family of transcriptional regulators that has so far been shown as possessing a redox-active cysteine. In contrast to the thiol switches just described, SsrB is a regulator of genes dedicated to bacterial pathogenesis. SsrB and its cognate SsrA sensor kinase form a two-component regulatory system essential for the intracellular replication of Salmonella (22). By analogy to other two-component regulatory systems, the SsrA sensor kinase phosphorylates Asp56 in the receiver domain of SsrB, unfolding the DNA binding domain for activation of gene transcription. SsrB binds to a flexible AT-rich palindrome sequence in ancestral and horizontally acquired genes (11). The phosphorylated SsrB protein relieves the gene silencing imposed by polymerized H-NS nucleoid protein on target DNA (105). SsrB partially binds to the predicted σ70 −35 region and activates cyclic AMP receptor protein-type class II promoters independently of the αCTD subunit of the RNA polymerase (105).
More than 100 loci are regulated by SsrB, including the 25 kb Salmonella pathogenicity island 2 (SPI2) that encodes a secretion apparatus, chaperones, translocon, the SsrA/SsrB two-component regulatory system, and effector proteins (39, 97, 108). SPI2 effectors translocated into the host cell cytoplasm help remodel Salmonella phagosomes and Salmonella-containing vacuoles. SsrB increases antioxidant and antinitrosative defenses (7, 12, 32, 91, 100, 101, 103); however, it does it in dramatically different ways from other regulators discussed in this review (Fig. 4). OxyR, OhrR, and RshA regulate the expression of classical enzymatic detoxification systems, whereas SsrB lessens oxidative and nitrosative stress of intracellular Salmonella by activating the expression of the SPI2 type III secretion system that decreases contact of phagosomes with incoming NADPH phagocyte oxidase- and inducible NO synthase-containing vesicles. In addition, the SPI2 type III secretion system prevents phagolysosome formation, while promoting the interactions of Salmonella-containing vesicles with nutrient-rich vacuoles in the exocytic pathway (58, 86, 99). As a consequence of the vacuolar remodeling, the SPI2 type III secretion system allows Salmonella to survive and grow in a variety of epithelial cells and mononuclear phagocytes, thereby playing a vital role in the course of systemic infection (74, 88). SsrB and the SPI2 type III secretion system are essential for Salmonella pathogenesis, as Salmonella strains lacking a functional SPI2 type III secretion system are attenuated at least five orders of magnitude (88).
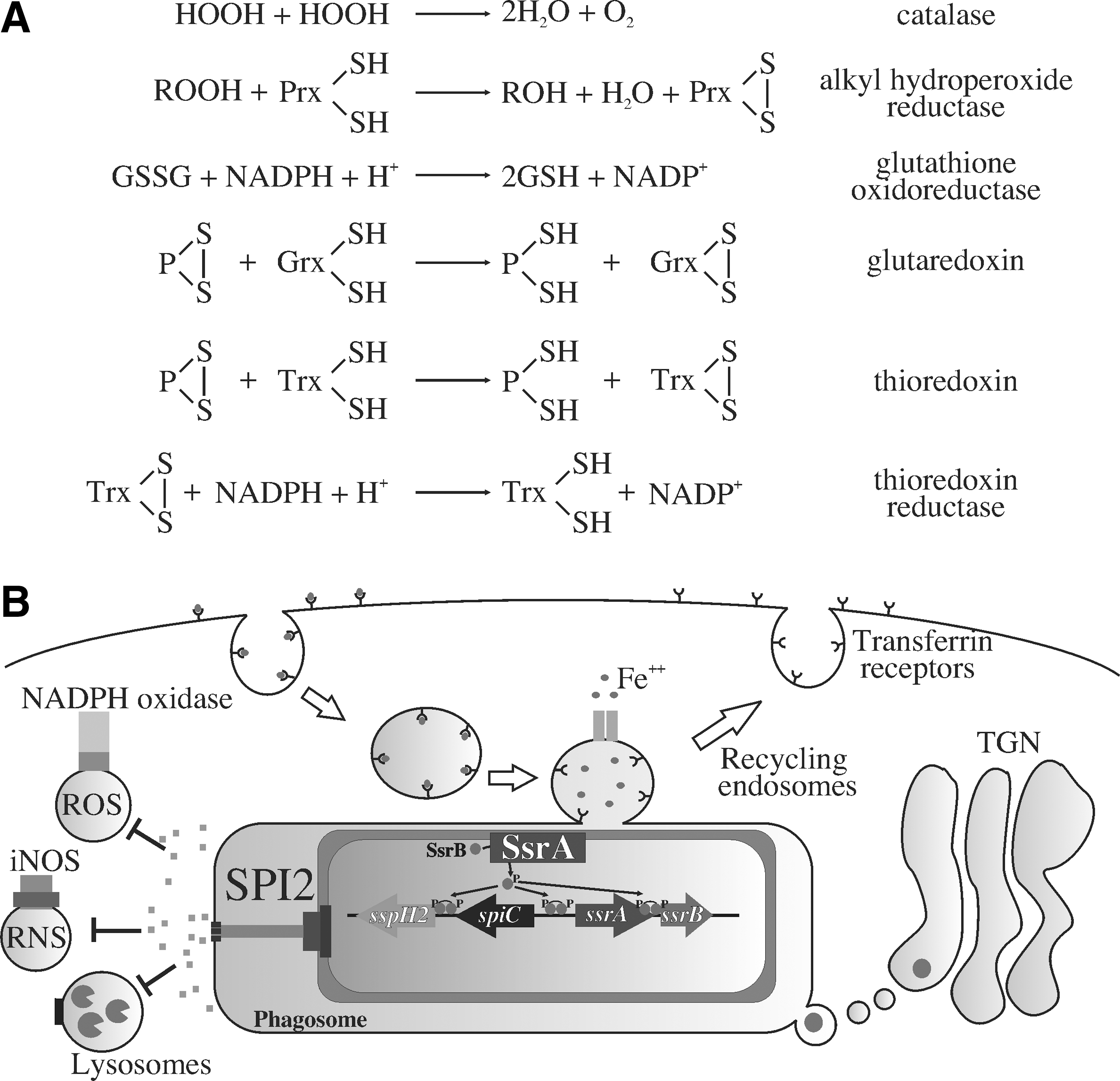
As it will become clear next, SsrB Cys203 is modified by RNS (43). A Salmonella strain expressing the SsrB C203S variant exhibits wild-type virulence in an acute model of infection, which is consistent with the fact that this allele is transcriptionally active (11, 43). Nonetheless, a strain of Salmonella bearing the SsrB C203S variant is attenuated in the context of nitrogen oxides produced in an NrampR murine model of oral infection (43). These findings suggest that Cys203 in the C-terminal dimerization domain of the SsrB response regulator is a sensor of RNS produced in the course of a gastrointestinal infection.
Regulation of SsrB function by the redox-active Cys203
As is the case for other response regulators, phosphorylation of a key aspartate in the receiver domain triggers the binding of SsrB to cognate promoters (11, 27). Recent investigations indicate that in addition to classical regulation by phosphorylation, two other signals influence SsrB's modus operandi. The EIIANtr component of the nitrogen-metabolic phosphotransferase system prevents the detrimental hyperactivation of SPI2 transcription by post-translationally precluding the association of SsrB to target promoters (19). The DNA-binding activity of SsrB is also subject to regulation by RNS (43). Cys203 in the α4 helix of the dimerization domain is susceptible to S-nitrosylation and oxidation by RNS such as ONOO−, and products of the reaction of NO and oxygen (O2). ONOO− oxidizes the thiol group of Cys203 to a mixture of sulfenic, sulfinic, and sulfonic oxidative products (43). C203D or C203E substitutions mimicking the negative charge of high-order oxidative products of ONOO−-treated SsrB block dimerization and the transcriptional activity of SsrB (11, 43). The side chain of Cys203 interacts with the hydrophobic residue Leu192 located in the loop between the DNA-binding H3 helix and the dimerization H4 helix (11). Sulfinylation or sulfonylation of Cys203 would disrupt this interaction, and, thus, alter the secondary structure required for homodimerization of SsrB, DNA binding, and transcription. Accordingly, the electronegative charge introduced by C203D or C203E substitutions inhibits transcriptional activity (11, 43). S-nitrosylation would not be expected to alter the interactions between Cys203 and Leu192. However, S-nitrosylation is not likely to be a terminal modification, but a precursor of further oxidations and/or S-thiolation. The extent to which these modifications prevent the formation of SsrB homodimers and affect gene transcription awaits further investigation.
Determinants That Define the Redox Activity of Cysteines
Cysteines are abundant in proteins, but only a few of them are modified by ROS and RNS. What makes a cysteine become redox active? A comprehensive review by Stamler and collaborators, and elegant recent work by the Ischiropoulos' group coincide to identify electrostatic interactions that affect pKa, hydrophobicity, and allosteric constraints as important determinants for the S-nitrosylation of selected thiols (26, 35, 40). These principles also dictate the reactivity of thiols toward ROS. Thiol-based bacterial redox sensors discussed in this review nicely illustrate the physical and chemical determinants that affect the reaction of redox-active cysteines with peroxides and nitrogen oxides.
Redox mechanism for the formation of thiolate in OxyR
Cys199 is located in a hydrophobic pocket between two α/β domains surrounded by acidic and basic residues (18). His130, Glu151, His198, Arg201, Asp202, and Arg266 may influence the reactivity of OxyR with H2O2 (18, 59). Arg266 has been shown to lower the pKa of Cys199 (18). The positive effect of Arg266 on Cys199 redox capacity is demonstrated by the fact that a strain of E. coli expressing the oxyR R266A variant is both hypersusceptible to oxidative stress and defective in oxyS expression in response to H2O2 (18). Dipoles from neighboring α-helices may also influence the ionization of Cys199 thiol (18). The Cys199 thiolate is likely stabilized by hydrophobic interactions with Leu200, Leu224, and Pro241 (18). The reaction of the thiolate with H2O2 forms a −SOH that forces Cys199 out of the hydrophobic pocket. This, in turn, induces flexibility of the region between 205 and 216 residues where Cys203 resides, thereby increasing the probability of collision between sulfenylated Cys199 and reduced Cys208 to form a disulfide bond (63).
Sensing by MarR/OhrR family members
The proximity of charged residues within 6 Å of a thiol group increases its potential to be ionized (26). No basic residues are found within a 7-Å distance from the OhrR Cys15 side chain (42). Instead, a positively charged macrodipole in helix 1 decreases the B. subtilis OhrR Cys15 pKa value to 5.2 (42). Therefore, this cysteine is kept in the ionized, thiolated form under physiological conditions, ready to react with incoming organic hydroperoxides. The negative charge of the thiolate in helix α1 is stabilized by hydrogen bonding with the side chains of Tyr29 and Tyr40 in helix α2 (42). The mutations of Tyr29 and Tyr40 have shown that hydrogen bonding to the Cys15 thiolate is important for B. subtilis OhrR sensing of organic hydroperoxides (42). The residues that form hydrogen bonding with the redox active thiolate and a hydrophobic pocket near it are conserved between B. subtilis OhrR and Sta. aureus SarZ and MgrA repressors (15, 42, 84). Hydrogen bonding is also important for the stabilization of the sulfenic modifications of SarZ Cys13 and MgrA Cys12. S-thiolation disrupts hydrogen bonding, thereby triggering a conformational change that lowers the affinity of these repressors for DNA.
Activation of thiols in the zinc-binding RsrA antisigma factor
Zinc can increase the reactivity of cysteine by lowering the pKa of the coordinating thiol side chains. However, similar to the situation described for the eukaryotic protein Sp1 (56), zinc protects the redox-active cysteines of RsrA from oxidative stress (66). Since zinc actually lowers the reactivity of the triggering thiolate in the finger, other determinants should activate the critical cysteine(s). The electronegative charge provided by glutamate residues flanking the HX3CX2C ZAS motif serves as an important redox determinant for the reactivity of RsrA (49). It remains to be investigated to what extent these glutamate residues influence the pKa and reactivity of the Cys11 triggering switch.
Activation of SsrB Cys203
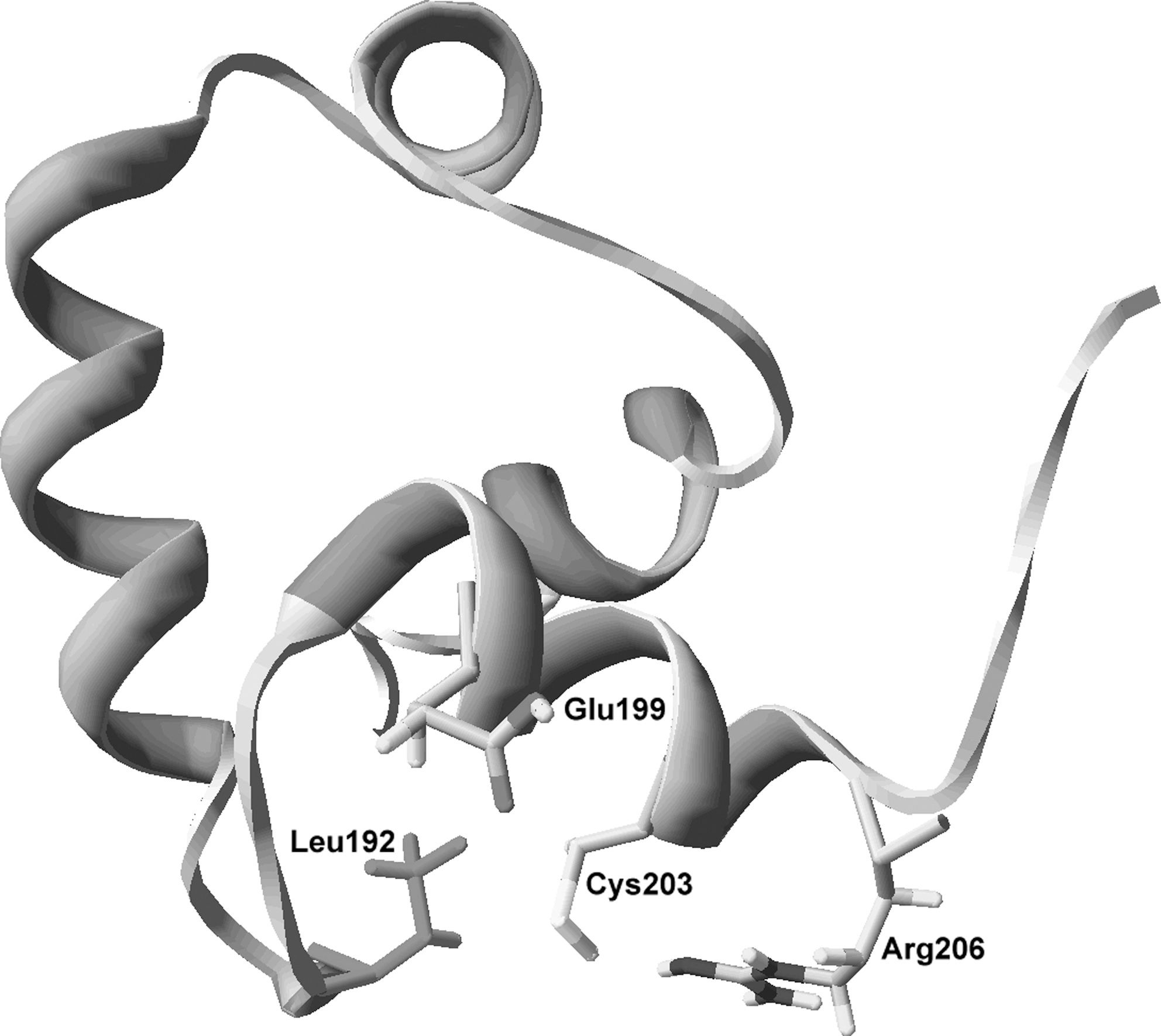
The fact that Cys203 reacts with nitrogen oxides and forms a disulfide bond with Cys203’ of the homodimer (11, 43) indicates that the thiol group of this cysteine is redox active. The determinants that make SsrB Cys203 redox active have yet to be identified. However, lessons learned with the bacterial regulators just mentioned and well-characterized redox active cysteines in a variety of proteins help make some informed predictions on how the environment surrounding SsrB Cys203 may dictate the redox activity of this response regulator (Fig. 5). As predicted by the fact that most reactive cysteines are overrepresented in α-helices (26), Cys203 is located in the SsrB's H4 α-helix. Basic residues at position 2 and acid residues at positions −3 and −4 influence the pKa of thiol groups (35). The analysis of the flanking residues around Cys203 indicate the presence of both positive and negative residues, raising the possibility that Cys203 is ionized in the cytoplasm of Salmonella. Of note, the guanidine group of Arg206 is 4.1 Å from the Cys203 thiol side chain. Moreover, the carboxyl group of Glu199 is 6.1 Å from Cys203. This configuration raises the possibility that the Cys203 side chain is, in fact, in the thiolate form. The hydrophobicity of the H4 helix and Leu192 located only 4.2 Å away could stabilize the Cys203 thiolate. These interesting scenarios await further investigation.
Chemistry Underlying the Modification of Redox-Active Thiols by ROS and RNS
Redox active thiol groups involved in signaling react selectively with ROS and RNS. The resulting oxidative or nitrosative modifications are often reversible. The mechanism by which thiol-dependent bacterial redox sensors react with peroxides is well understood (Fig. 6); however, several models could explain the reactivity of thiol switches with RNS for the formation of S-nitrosothiols.

Selectivity of the reaction of thiols and ROS
The rate constants of hydroxyl radical, O2 •−, and H2O2 with LMWTs are 1010, 30–1000, and 20 M −1 s−1, respectively (106, 107). Although it has high affinity for thiols, the hydroxyl radical reacts nonspecifically with nearly all biomolecules, and, therefore, it is not endowed with the discriminatory power to serve as a signaling molecule. Superoxide dismutases detoxify O2 •− with a rate constant of 2×109 M −1 s−1 (29), making the O2 •−-dependent oxidation of thiols of little physiological importance. On the other hand, H2O2 can react with fast kinetics with certain thiols. For example, the second-order rate constant for the reaction of OxyR Cys199 and H2O2 is about 105 M −1 s−1 (2), making this regulator an excellent H2O2 sensor. As discussed elsewhere in this review, OxyR Cys199 reacts with RNS, but with apparently lower affinity than ROS. It is currently unknown why OxyR Cys199 in particular and redox active cysteines in general exhibit preferential affinity toward selected reactive species. To define the determinants that provide selectivity of redox-active thiolates for some reactive species but not others is an important goal for the future.
Oxidation of redox active thiols by H2O2
Although the reactivity of thiols with ROS is inversely related to their pKa (107), there are examples where this does not seem to be the case (106). In addition to a low pKa that ionizes the thiol to a thiolate, the environment of the protein is critical for the stability and reactivity of the thiolate. Reactive thiols in MarR, LysR, and zinc-finger family members just discussed have low pKa values and are generally located in hydrophobic pockets. As described in other systems (83), the two-electron oxidation of the redox-active thiolate in the regulators just presented by H2O2 or organic hydroperoxides gives rise to a sulfenic group. The oxidation of a thiol to a −SOH is not enough to transmit the signal in most regulatory proteins. The −SOH is frequently oxidized to a disulfide in order to drive the structural changes that modulate protein function. In the case of E. coli OxyR and Str. coelicolor RsrA, intramolecular disulfides are responsible for major structural changes, whereas mixed disulfides are necessary in the case of B. subtilis OhrR. In addition to mediating dramatic changes in secondary structures that transduce the signal, disulfide bonds prevent further oxidation of cysteine −SOHs to high-order oxidative species such as sulfinic and sulfonic groups. Disulfides are reversible modifications that on reduction turn off the signal elicited by H2O2, whereas the oxidation of thiols to sulfinic and sulfonic are, for the most part, irreversible and less amenable for signaling. Although the reaction of cysteines with peroxides is facilitated when the sulfur in the side chain is a thiolate, the modification of cysteines by other reactive species does not have to be limited to the ionic form of cysteines. For example, as will be discussed next, radicals can promote modifications of cysteines through a sulfenyl (i.e., thiyl) radical intermediate.
Selective reaction of thiols with peroxides
Not all redox active cysteines react equally with ROS. Cys15 of B. subtilis OhrR is an excellent case in point. Antioxidant defenses and intrinsic allosteric determinants explain the selective reaction of B. subtilis OhrR with organic hydroperoxides. H2O2 can oxidize the thiol of OhrR; however, this reactive species does not appear to be a relevant signal in vivo. The activity of catalases in B. subtilis diminishes the effective concentration of H2O2 to levels that are insufficient to initiate the oxidation of the critical cysteine residue in OhrR (31). The reaction of OhrR with organic hydroperoxides is fast. A patch of hydrophobic residues in the vicinity of the redox active cysteine increases the affinity of OhrR for organic hydroperoxides (42). Most eukaryotic organic hydroperoxides are derived from membrane lipids. A radical abstracts an allylic hydrogen in the polyunsaturated hydrocarbon, giving rise to a conjugated diene that is susceptible to the O2 attack for the generation of lipid hydroperoxides (41). The lack of polyunsaturated hydrocarbons in prokaryotic lipids greatly limits the formation of lipid hydroperoxides in their membranes. Although extremely active when supplied in pure form, it remains to be seen whether organic hydroperoxides are the actual signal for members of the OhrR family of regulators in vivo. The thiol oxidizer diamide also derepresses the OhrR regulon (31), raising the possibility that the cysteines triggered in OhrR may respond to thiol/disulfide exchange reactions. OhrR also appears to be a sensor of hypochlorite stress (17).
Reaction of thiols with RNS
Thiol groups in regulators such as OxyR and SsrB can be modified by RNS. S-nitrosothiols can be generated by several independent mechanisms (Fig. 7). Transnitrosation and radical reactions may account for the nitrosylation of redox-active thiols in the regulators discussed in this review. In the case of OxyR, RNS modify the Cys199 thiolate to form an S-nitrosothiol. The nitrosonium cation is the most likely candidate that covalently modifies the nucleophilic thiolate of OxyR Cys199. However, nitrosonium does not exist free in solution. It has consistently been reported that OxyR is more efficiently activated by nitrogen oxides in GSH-deficient E. coli strains, suggesting that transnitrosation reactions from species other than S-nitrosoglutathione are the most likely nitrosating species. Lancaster has proposed that dinitrosyl iron complexes (DNICs) are the most important nitrating agents in the cell (8). DNIC arising from the complexation of NO, iron, and cysteine, or other LMWTs, could be a source of the nitrosonium-like species that S-nitrosylates the OxyR Cys199 thiolate group.
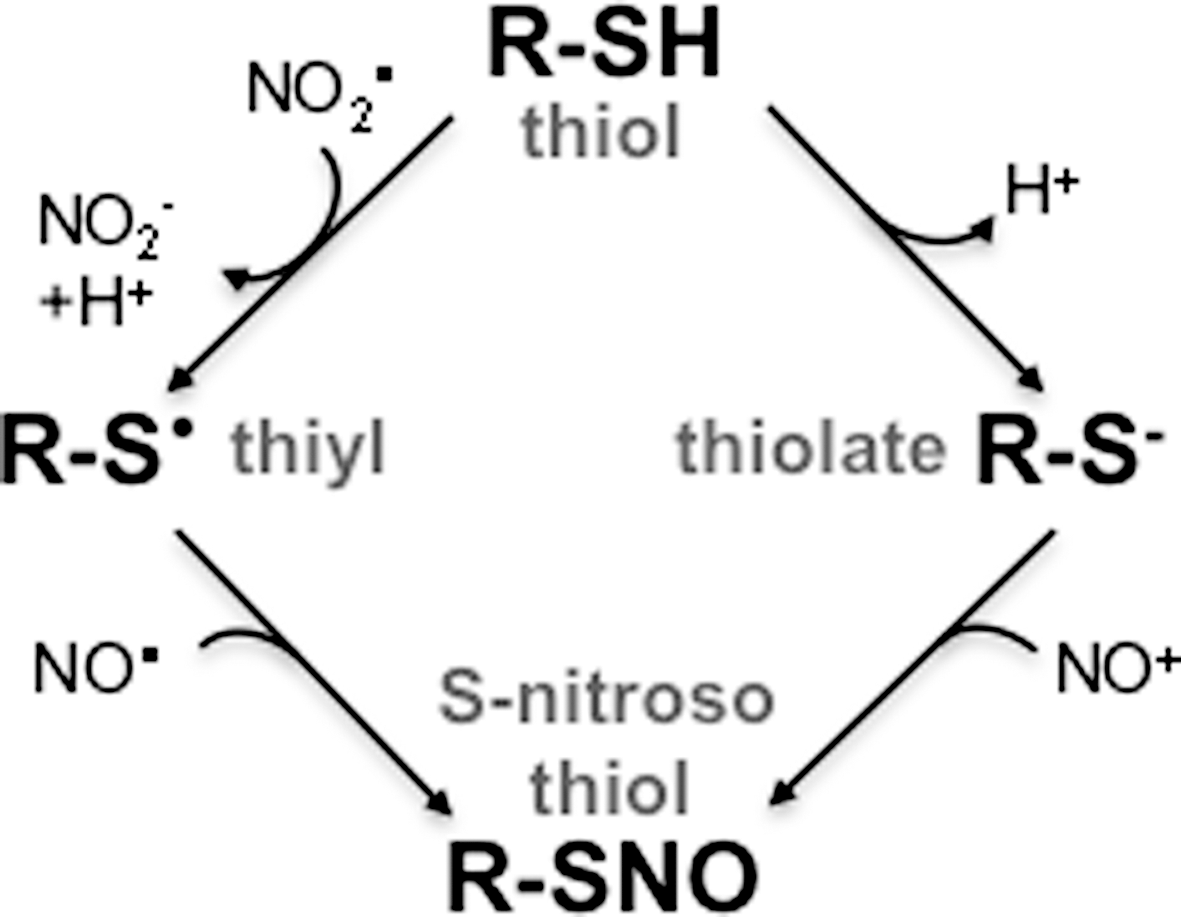
The Cys203 of the response regulator SsrB has also been shown to react with nitrogen oxides (43). The DNA binding activity of SsrB is inhibited by high fluxes of NO in the presence of O2. This indicates that products of the autooxidation of NO mediate the inhibition of binding of SsrB with cognate promoters. Together with DNIC and S-nitrosothiols, dinitrogen trioxide (N2O3) is a potent nitrosating agent that serves as a source of nitrosonium groups (54). The production of N2O3 is second order for NO, and some investigators have argued that the generation of N2O3 is not favored under physiological conditions. Alternatively, nitrogen dioxide produced in the autooxidation of NO could abstract an electron from the thiol group of SsrB Cys203, generating a thiyl radical that could condensate with the NO radical to form S-nitroso SsrB. As is the case for most biomolecules, the chemical species that mediates S-nitrosylation of SsrB is currently unknown.
Concluding Remarks
Cysteine and methionine have a sulfur atom in their structures. Sulfur in methionine is frequently oxidized to a sulfoxide group. This oxidative modification can be repaired through the enzymatic activity of the methionine sulfoxide reductases. The reactivity of methionines with ROS, the reversibility of the oxidation, and the enrichment of oxidizable methionines in protein surface areas and enzyme active sites have led to the proposal that the sulfur in methionine acts as an endogenous antioxidant which protects biomolecules from the attack of reactive species (65, 68). Despite being redox active, methionine has not yet been found to be a redox-sensing moiety of bacterial transcriptional regulators. Several reasons may explain the predominance of cysteines rather than methionines as common signaling redox determinants. Cysteines and methionines can be found in 10 and 3 different oxidation states in the cell, respectively (46). Thus, cysteines provide a larger repertoire in oxidation states, making it possible to sense diverse ROS and RNS. However, of all possible oxidation states, thiol-dependent bacterial sensors rely quite frequently on thiolates and disulfide bonds as effective ways to transduce a signal. Why would disulfide bonds be a preferred mode of signal transduction? Disulfides are reversible modifications that can be in balance with thiolates by the action of Trx, GSH, and Grx thiol/disulfide exchange systems. Equally important, the dramatic structural rearrangements associated with disulfide bond formation are accompanied by changes in function. A case in point is the fact that the newly formed disulfide between Cys199 and Cys208 in oxidized OxyR brings together regions separated by 17 Å in the reduced protein (18). This massive structural reorganization allows binding of OxyR to the RNA polymerase with the consequent initiation of transcription.
The regulators discussed in this review reveal variability in the chemistry by which disulfide bonds are generated, and the diverse structural means by which they modulate molecular function (Fig. 8). In addition to the aforementioned intramolecular disulfide bond that drives OxyR transcriptional activity, intermolecular and mixed disulfide bonds affect the function of bacterial regulatory proteins. Intermolecular disulfide bonds in X. campestris OhrR and mixed disulfides in redox active thiolates in lone cysteine OhrR-type regulators are additional ways by which redox-active cysteines transduce signals in response to oxidative and nitrosative stress. The mechanisms underlying the S-thiolation of redox-active cysteines are equally varied in nature. Mixed disulfides in the OhrR-family members have been shown to occur through a −SOH oxidative intermediate. S-thiolation can independently arise through an S-nitrosylated intermediate as described for OxyR (55). In either case, mixed disulfides make possible the structural and functional changes of the oxidized or S-nitrosylated regulatory protein.

Thiol-dependent redox sensing coordinates bacterial responses to changes in physiology and pathogenesis. OxyR, OhrR, RshA, and SsrB add to the antioxidant and antinitrosative defenses of bacteria. In addition, the oxidized thiols in some sensors such as OxyR and RsrA have been shown to be substrates of the antioxidant defenses activated in their regulons. This feedback regulation ensures that redox homeostasis is re-established after the burst of reactive species has subsided. This is an important consideration, as bacterial cells experience fluctuations in oxidative stress during different growth phases and during preferential utilization of certain metabolic pathways (57). In addition, pathogenic microorganisms are exposed to a burst of intense oxidative stress (102). The redox activity of thiol-based sensors increases fitness of saprophytic, commensal, and pathogenic bacteria to fluctuations in oxidative and nitrosative stress in their ever-changing environment.
Since investigations with E. coli and Salmonella discovered the thiol-based H2O2 sensor OxyR (20), an increasing number of members within the LysR, MarR, RsrA, and NarL families of transcriptional regulators have been shown as using thiol-based redox sensors that coordinate responses to endogenous and exogenous ROS and RNS. As the range of thiol-based prokaryotic redox sensors broadens, it will be interesting to see the extent that the concepts discussed in this review apply.
Footnotes
Acknowledgments
The author thanks Drs. M. Crawford, J. Helmann, H. Ischiropoulos, J. Jones-Carson, and M. Song for their excellent suggestions. He is also grateful to Dr. C. Henard for discussions and help with the in silico analysis of the SsrB NMR structure. This work was supported by the National Institutes of Health grant AI54959 and the Burroughs Wellcome Fund.