
Abstract
Introduction
Indeed, the widespread detection of the products of lipid peroxidation, from both enzymatic and nonenzymatic sources, in a broad range of pathological conditions has led to the concept that RLS may also contribute to disease progression (3, 14, 39, 55, 67). For example, in cardiovascular disease, the underlying inflammatory process, which through regulation of gene expression by nuclear factor kappa B (NFκB) can increase cyclo-oxygenase (COX)-2 as well as PLA2, leads to enhanced oxidation of fatty acids (88, 100). In ischemia-reperfusion injury, hemolysis occurs, leading to an increase in free-radical catalyzed lipid peroxidation due to the release of hemoglobin and free heme (44, 47). In hemolytic anemias, increased levels of RLS have been reported, and lower concentrations of the lipid radical scavenger Vitamin E are also consistent with increased lipid peroxidation (1, 28, 44, 65, 69, 86). Several factors in heart failure, including increases in ROS from xanthine oxidase and the mitochondria, enhance the oxidation of phospholipids (4, 61, 84, 92).
While it is then established that these pathological processes are associated with lipid peroxidation, the mechanisms through which they mediate biological effects are less clear. More recently, these concepts have been further developed to encompass the finding that low levels of RLS activate the signal transduction pathways which control the levels of intracellular antioxidants such as glutathione (GSH) and heme oxygenase-1 (17, 28, 56, 71, 101). A contemporary view of lipid peroxidation is that oxidized lipids can elicit different cellular effects depending on the RLS present (the oxylipidome), their concentrations, and, importantly, their reactivity with biomolecules, including lipids, DNA, and proteins (6, 38, 51, 63). The focus of this review will be to discuss new concepts through which the oxylipidome modulates biological functions through the formation of electrophile-responsive proteomes (ERP) in cells.
Generation of Oxylipidomes in Biological Systems
Lipid peroxidation in cells can occur through controlled enzymatic pathways, including the lipoxygenase (LOX), COX or cytochrome p450 enzymes (43, 57, 74, 83, 94). Some of these enzymes are constitutive and contribute a low level of oxidized lipids with a predominantly autocrine function, and some are inducible and generate high levels of oxidized lipids that exhibit both autocrine and paracrine functions. Nonspecific lipid peroxidation describes any oxidation process that is not mediated by a controlled enzymatic mechanism. Recent advances in mass spectrometry are overcoming difficulties in defining the complete oxylipidome produced by different mechanisms of lipid peroxidation, allowing for the characterization of the regulatory oxylipidomes for COXs and LOXs (27, 97). It has been established that the oxylipidome generated by controlled enzymatic pathways (e.g., LOX and COX) and nonspecific lipid peroxidation are distinct in two important areas: (i) The isomers, for example, F2-isoprostanes, generated from nonspecific lipid peroxidation, show little stereospecificity. In contrast, the enzymatic mechanism of lipid peroxidation generates specific and limited numbers of stereoisomers, for example, PGF2α and, hence, different oxylipidomes (26, 32, 98). (ii) The nonspecific oxylipidome is populated by RLS not formed by enzymatic mechanisms. These include many aldehydes, as well as nitrated or chlorinated lipids formed by RNS or chlorine species, respectively (2, 24, 93, 99). Selected examples of the members of the oxylipidomes produced by nonspecific and specific lipid peroxidation are shown in Figure 1. From the Venn diagram, it is obvious that biological oxylipidomes are composed of varying proportions of different RLS with distinct properties and reactivities as shown in Figure 2. From this, it is then a logical step to propose that the biological effects arising from an oxylipidome will be related to how the members are distributed among these functionally distinct classes of lipid electrophiles.
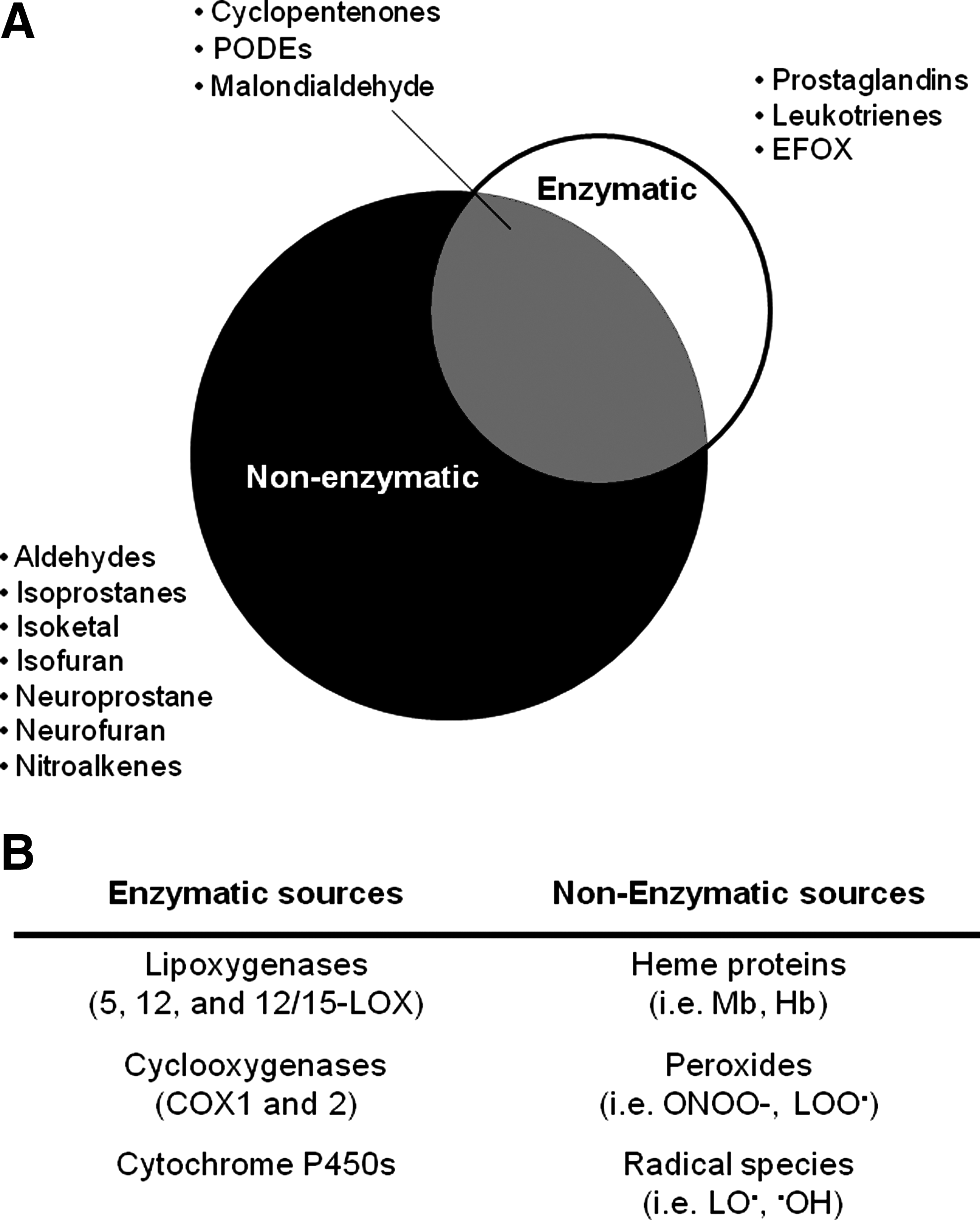
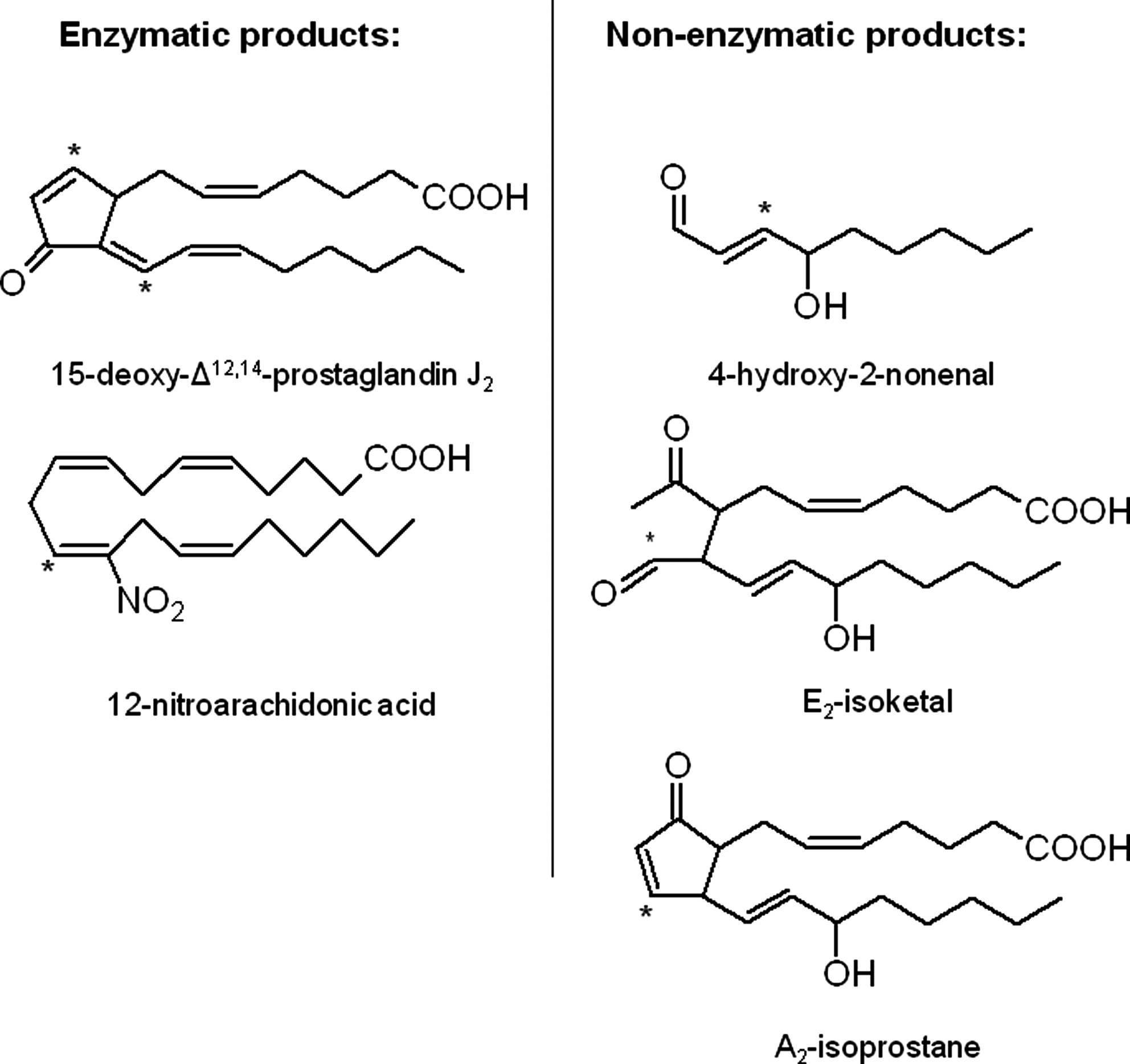
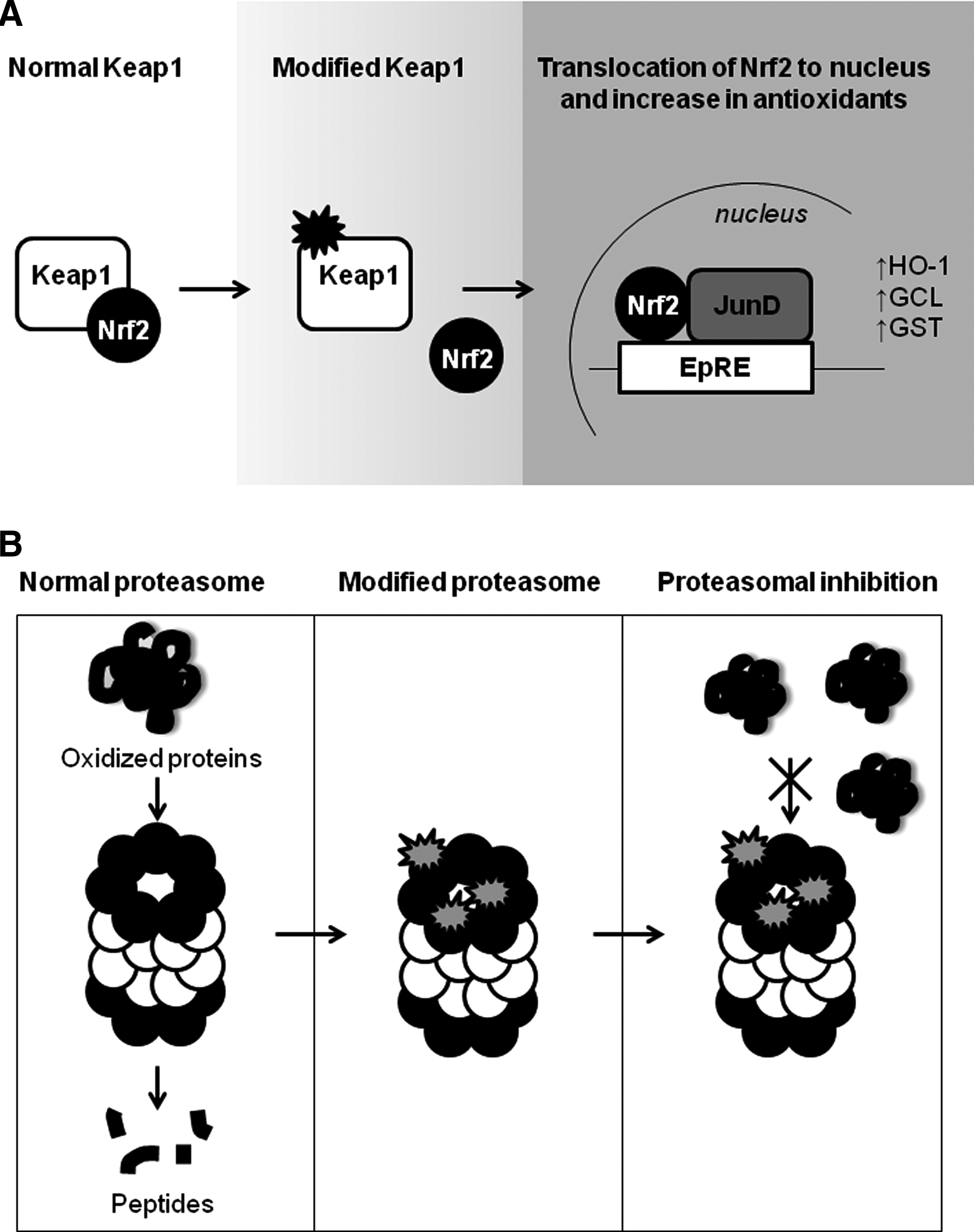
The biological effects of RLS build on a considerable amount of research that has defined the biological effects of a candidate member of an oxylipidome in cells or more complex animal models of a disease. Two typical, but structurally distinct examples (Fig. 2) are the electrophilic lipid aldehydes HNE, a nonspecific lipid peroxidation product and the cyclopentenone prostaglandin, 15-deoxy-Δ12,14-prostaglandin J2 (15d-PGJ2) (12, 72, 80). From such studies, some generalized conclusions of the biological outcomes associated with specific and nonspecific lipid peroxidation have been achieved, and these are outlined in Figure 3. The enzymatic lipid peroxidation products can increase cytoprotective pathways in the cell through modification of cysteine residues in cell signaling proteins, such as the Keap1/nuclear factor-erythroid 2 related factor (Nrf2) system (Fig. 4B) (40, 49, 51, 68, 85). In contrast, the nonenzymatic lipid peroxidation products while also showing some cross-over activation of the same cytoprotective pathways damage proteins, cause bioenergetic dysfunction and depletion of cellular antioxidants, such as GSH (52). This is partly due to the differential reactivity of oxidized lipids that are nonenzymatically produced. Highly RLS such as isoketals, isofurans, and aldehydes (such as 4-oxo-2-nonenal) are formed by free-radical catalyzed lipid peroxidation and unlike softer electrophiles, these react with amino acids other than cysteine and are, therefore, less specific. It is important to note that if the concentrations of the enzymatic lipid peroxidation products are increased beyond the physiological levels, then they induce the same pathological responses as the nonspecific lipid peroxidation products (49).

The Relationship Between the Oxylipidome and ERPs
An important and technically challenging problem has been the identification of the relationship between the oxylipidome and the protein targets (the ERP) through which a biological response can be initiated. As noted earlier, the structurally different RLS result in a range of electrophilicities that changes their reactivity with the nucleophilic amino-acid residues in proteins. For example, the aldehyde HNE is a reactive electrophile forming adducts with histidine, cysteine, and lysine residues (34), whereas a cyclopentenone 15d-PGJ2 is more selective for protein cysteine residues (51, 78, 81). From this, we hypothesize that the proteins modified by any complex oxylipidome will be both a property of the mechanism of lipid peroxidation and the nucleophilic proteome to which they are exposed. The implication of this hypothesis is that the Venn diagram in Figure 1 should map onto a corresponding ERP, as shown in Figure 5. In this scheme, we show several enzymatic and nonenzymatic pathways that can lead to lipid peroxidation in biological membranes, in this case, the mitochondrion (Fig. 5A). The oxidation reaction leads to distinct oxylipidomes (a–e) which will be composed of different RLS that are then exposed to a proteome containing reactive nucleophilic amino-acid residues. This results in different ERPs that we have represented as 3 hypothetical different protein spot patterns for RLS-protein adducts a, b, and c. In Figure 5B, the putative relationship between ERP-a, b, and c with regard to biological responses is illustrated. The physiological/cytoprotective ERP-a can be ascribed to cysteine modification, whereas ERP-c is dominated by protein damage and bioenergetic dysfunction. ERP-b is an intermediate state in which the cytoprotective and cytotoxic pathways are in tension and is the arena in which therapeutic intervention could result in maximum benefit.
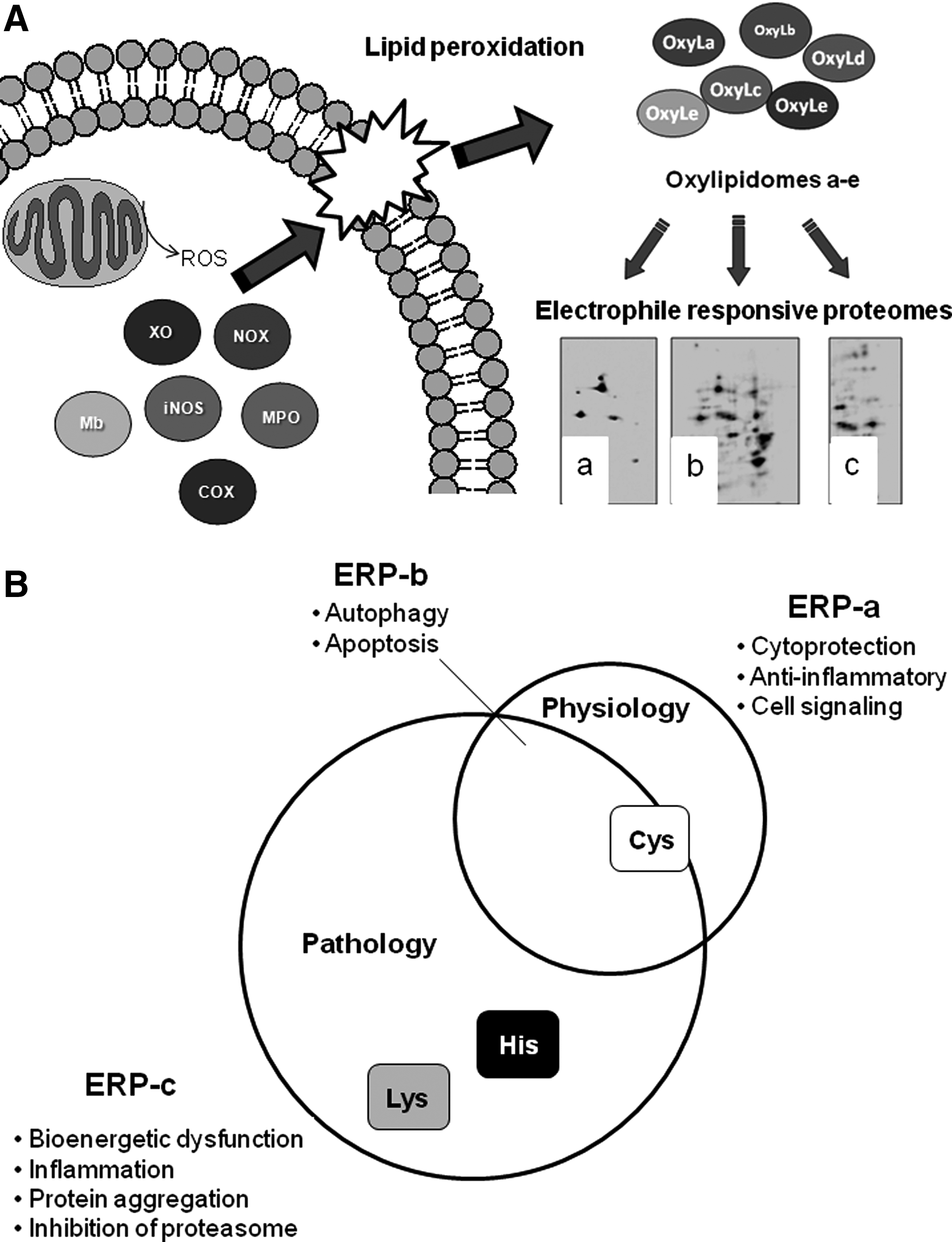
Due to the importance of thiol modification in redox signaling, many studies have focused on the role of cysteine modification in eliciting the effects of various electrophiles. While the modification of a cysteine can lead to pathological effects if, for example, this amino acid is a part of the active site of an enzyme, then it is becoming increasingly appreciated that the modification of thiol residues can lead to physiological cell signaling (Fig. 5B). Interestingly, recent studies have demonstrated that the complete thiol-reactive ERP in a cell can be defined using two different, biotin-tagged electrophiles, PEO-IAB (biotinyl-iodoacetamidyl-3, 6-dioxaoctanediamine) and BMCC (1-biotinamido-4-(4′[maleimidoethylcyclohexane]-carboxamido) butane) with a specificity for reactions with cysteine residues (16). Approximately 500 cytosolic and nuclear proteins were modified, and of these, only 14% were modified by both electrophiles. This suggests that even within the population of electrophiles that react with cysteine residues there are subpopulations of differing selectivity. In a separate experiment, a distinct mitochondrial ERP was also identified. These data support the concept that the modification of proteomes by electrophiles is selective depending on both the electrophile and the local environment in which the reactive protein nucleophiles are located (16). It is important to note that these studies using thiol-specific reagents at high concentrations identify the thiol ERP which will be distinct from the ERP generated by more reactive electrophilic lipids.
The Cytoprotective ERP
A low-abundance thiol proteome, which controls redox signaling pathways and leads to cytoprotection in the cell, is activated by electrophiles (11, 13, 41, 77). The pathways have a common target in a low pKa reactive cysteine residue controlling the activity of a regulatory cell signaling molecule (13, 37). Members of this ERP include Keap1, which regulates the transcription of antioxidant enzymes, the heat shock family, which control protein folding, and the NFκB system, which regulates the transcription of pro-inflammatory proteins (37, 77). Interestingly, recent studies also suggest that the Nrf2 transcription factor coregulates the autophagy pathway, and electrophilic signaling may also be important in handling oxidatively damaged or aggregated proteins or organelles (75, 76).
Perhaps the best understood electrophile sensor in the cell is Keap1 (29, 59). This protein expresses an array of thiol sensors that detect electrophiles and other thiol reactive molecules differentially (59). The Keap1/Nrf2 pathway is activated by electrophilic lipid oxidation products, and complex oxylipidomes such as oxidized low-density lipoprotein (19, 50, 51, 56). The molecular mechanism for the transcriptional regulation of antioxidant enzymes is mediated, in part, through the dissociation of the transcription factor Nrf2 from its cytoplasmic binding protein Keap1 and binding to the electrophile response element or antioxidant response element (51, 66) as shown in Figure 4B. The control of cell signaling requires a reversal mechanism that prevents permanent activation. In the case of electrophilic signaling, the covalent modification of protein thiols by electrophiles is reversed by the proteasomal degradation of the modified protein (35, 58). Another interesting feature of electrophile signaling is that the formation of a covalent bond between the RLS and a signaling protein allows the accumulation of a signal over time (68). This finding reconciles the long-standing problem in the electrophile signaling field that the free forms of RLS are invariably found at low concentrations in biological systems and below that required to initiate the same signaling pathways with a pure electrophile in cell cultures. Thus, in the case of the ERP, there is a “covalent advantage” to signaling through the covalent modification of the nucleophilic protein target (17, 68). Since, in the model we propose (Fig. 5) the cell responds to an ERP, the accumulation of a covalent adduct offers an additional level of regulation. This could occur through modulating the composition of the ERP through differential proteasomal-dependent degradation of members of the proteome.
An interesting possibility which is now emerging is that the mitochondrion serves as a generator of low levels of endogenous RLS that contributes to the phenomenon of retrograde signaling in which the mitochondrion communicates with the nucleus. In support of this concept, cell signaling mediated by the promotion of mitochondrial ROS is inhibited by mitochondrially targeted, lipid radical scavengers, suggesting that mitochondrial hydrogen peroxide can be transduced to a lipid-derived electrophilic signal (9, 95). In endothelial cells, it has recently been shown that the Keap1/Nrf2 induction by 15d-PGJ2 of the antioxidant enzyme heme oxygenase-1 requires the participation of the mitochondria and that Keap1 can be tethered to the mitochondrion (53, 73). A critical cell signaling pathway involved in cardioprotection is known as ischemic preconditioning. The finding is that brief periods of mild ischemia result in protection of the heart from a more severe ischemic episode. A recent study has shown that electrophilic nitroalkenes are formed in the mitochondria and are cardioprotective (64). The molecular targets are still unclear but appear to involve mild uncoupling.
The Cytotoxic ERP
Defining the targets of electrophiles in cells that promote cell death has been more challenging, as this typically requires much more reactive electrophiles at high concentrations. Interestingly, more reactive electrophiles, such as HNE, have been shown to modify and inhibit the function of the proteasome, removing one key mechanism of degrading proteins modified by these electrophiles (22, 23). This modification is one key example of a detrimental modification by RLS (Fig. 4B) and demonstrates a means whereby low concentrations of an electrophile could result in prolonged dysfunction due to the inability to remove modified proteins.
The activation of key signaling pathways involving cell death has been reported for electrophilic lipids, including p53 (42). Not surprisingly, the mitochondria have emerged as a potential mediator of these effects. The release of cytochrome c from the organelle is one of the key steps in apoptosis and is regulated by the interaction of cytochrome c with cardiolipin. Oxidation and the formation of RLS have been shown to be a key step in the apoptotic cascade (39). Experiments with candidate lipid electrophiles have identified some of the key steps in the cytotoxic responses to electrophiles. Not surprisingly, HNE has featured prominently in these studies and has been shown to induce bioenergetic dysfunction, mitochondrial ROS in both isolated mitochondrial and cells (10, 31). There are multiple targets in the mitochondrion that could be contributing to bioenergetic function, and we would propose that it is the composite ERP which determines the overall bioenergetic dysfunction. For example, in a cellular setting, we have observed an increased proton leak that indicates damage to the ion transport or integrity of the inner mitochondrial membrane and decreased maximal respiration which is consistent with modification of the respiratory chain (31, 82). An interesting example of RLS-induced mitochondrial dysfunction occurs on exposure of cells to free heme. We have recently shown that the oxidant hemin increases lipid peroxidation and inhibits mitochondrial function in endothelial cells (30). The modification of proteins by RLS and alterations of mitochondrial function are attenuated by α-tocopherol, suggesting an important contribution by lipid peroxidation and inducing mitophagy (30). Indeed, the lipid peroxidation product HNE has been shown as modifying both complexes III and IV, and promoting uncoupling in isolated mitochondria (7, 8).
The permeability transition pore in the mitochondria is a complex multi-component channel intimately involved in both necrotic and apoptotic cell death (48). The regulation of the pore is highly dependent on thiol modification and can be modulated by RLS (60). For example, the exposure of mitochondria to cyclopentenone, 15d-PGJ2, results in a complex ERP associated with increased sensitivity to the mitochondrially induced permeability transition.
The mitochondrion may be a key integrating center for electrophile-dependent redox cell signaling because of the high pH in the mitochondrial matrix. This results in mitochondrial protein thiols being present in the thiolate form, which is more reactive with electrophiles. As discussed in the previous section, the localized production of mitochondrial ROS could control the production of endogenous electrophiles for cell signaling. Exogenous electrophilic lipids have also been shown to localize with the mitochondrion and modify proteins in this subcellular compartment.
Taken together, these data strongly support the concept that the mitochondrial ERP may predominantly promote pro-death signaling, whereas the cytosolic signaling pathways are cytoprotective, albeit with some cross-talk with the mitochondrion. To test this, we have taken the electrophile 15d-PGJ2 and synthesized a mitochondrial derivative (mito-15d-PGJ2). When compared with the native 15d-PGJ2, we found mito-15d-PGJ2 that was essentially incapable of inducing the Keap1/Nrf2 pathway and promoted bioenergetic dysfunction and apoptotic cell death (20). These data suggest that the ERPs are susceptible to pharmacological manipulation, and this will allow the differential control of cell signaling.
Summary
An important concept that has emerged in our understanding of oxidative stress in disease is that RLS modify the function of proteomes, not just single targets (70, 87, 93) (Fig. 5). Despite the success generated by the newer technologies capable of defining the oxylipidome, the resulting rich datasets are overwhelming the capacity of investigators to interpret this information. Further understanding of how oxylipidomics can explain biological effects will require the development of new bioinformatics platforms that integrate the data from lipidomics, protein modification, cell signaling, and cellular responses. This will be essential to address a key basic mechanistic question in redox biology: How does lipid peroxidation contribute to the pathophysiology of disease? In this review, we proposed a new paradigm which encompasses the fact that while lipid peroxidation is a consistent feature of multiple disease processes, the abundant literature in this area of redox biology reveals multiple protein targets (ERPs) that could elicit the biological response in question. The new concept is that it is the integrated behavior of the whole proteome which is responsible, not any given protein target in isolation, and this can be revealed through emerging technologies capable of integrating the bioinformatics datasets that relate oxylipidome with the ERPs and biological effects.
Footnotes
Acknowledgments
This work was supported by NIH R01 Grants ES10167, AA13395, DK075867 (to V.D.U.), P50 AT00477, R21 AT004661, U54 CA100949 (to S.B.), and HL096638 (to A.L.). The authors have no financial conflicts of interest related to this work.