
Abstract
Introduction
There is collective evidence that DCs are the main antigen-presenting cells (APCs) that can cross-present antigens in vitro and in vivo (64). High efficiency in cross-presentation is, at least in part, a consequence of fine-tuned regulation of antigen processing, in which ROS production seems to play a critical role (61, 113, 114). Given that DCs represent a heterogeneous cell population, it is still unclear however which DC subtypes are the most efficient cross-presenting cells, in particular, under specific inflammatory situations.
In mice, spleen DCs (also known as resident DCs) can be divided into two main populations according to the expression of CD8αα at their cell surface (141). Therefore, CD8+ DCs were shown to be more efficient than CD8− DCs in terms of cross-presentation at steady state (122), whereas both subtypes presented antigens efficiently in the MHC-II context. However, other DC subsets, such as migratory DCs and CD103+ DCs, have also been shown to cross-present efficiently (32, 35). In humans, the situation of DC subsets is as complex and heterogeneous as in mice. BDCA3+ DCs were recently proposed to be the homolog of murine lymphoid organ-resident CD8+ DCs (5, 63), and therefore to be specialized in cross-presentation. Among the human skin DCs, Langerhans cells and CD1a+ DCs have been suggested to be the most efficient cross-presenting cells (121).
It is most likely that the capacity to cross-present antigens between different DC subtypes is the final result of their intrinsic adaptation of intracellular pathways to this function, to their surrounding environment, and to the quality of the antigen.
In this review, we want to emphasize how phagosomal ROS generation has an impact on antigen processing and presentation.
The NOX Family
The NOX family consists of membrane-bound enzymes that catalyze the reduction of oxygen to superoxide using NADPH as an electron donor. The central component of all members of this family is a heterodimer of the product of the X-chromosomal cybb gene, the glycoprotein gp91phox (also referred to as NOX2), and the cyba gene, p22phox (76). These two membrane components of the inactive NOX form the flavocytochrome b558 and will be referred as such in this review. In addition to the membrane components, the cytosolic subunits p40phox, p47phox, and p67phox, as well as Rac GTPases (Rac1 or Rac2 depending on the cell type), regulate the activity of the NOX. In humans, the entire NOX family consists of NOX2 and six homologs: NOX1, NOX3, NOX4, NOX5, DUOX1, and DUOX2 (101).
NOX1 and NOX2 are the closest homologs of the family. NOX1 was identified in the colon, uterus, and prostate tissues, in epithelial barriers, as well as vascular smooth muscle cells. NOX1 has been implicated in signaling cascades of angiotensin-II, tumor necrosis factor α (TNFα), platelet-derived growth factor, and epidermal growth factor. For full activation in a cell type- and stimulus-dependent manner, NOX1 requires two additional proteins, the organizer NOXO1 and the activator NOXA1 (p47phox and p67phox homologs, respectively) (99).
NOX3 has been described in fetal tissues, the inner ear, hepatocytes, and macrophages and requires only p22phox and NOXO1 to generate ROS (26). However, participation of NOXA1 is able to enhance ROS production, whereas the involvement of Rac1 in NOX3 regulation is still controversial (136). More recent data have shown that the regulation is Rac-dependent at similar levels compared to NOXA1 (65, 129).
NOX4 is constitutively active and does not need the interaction with cytosolic regulatory subunits (65). It has been found in the kidney, osteoclasts, aortic endothelial cells, cardiac, and lung fibroblasts, as well as fetal tissues (101).
NOX5 has been described in human testis, spleen, lymph nodes, and endothelial and smooth muscle cells, but seems to be absent in rodent tissues. Although it is able to bind p22phox, complex formation is not required for ROS production. The recruitment of NOX5 to the plasma membrane requires PI (4,5) P2 and is regulated in Ca2+-dependent pathways without the involvement of p22phox, p47phox, p67phox, and Rac (69, 70).
The dual oxidases DUOX1 and DUOX2 possess an additional transmembrane segment with peroxidase homology and are able to generate hydrogen peroxide while their involvement in superoxide production is not clear (99). They are highly expressed in thyroid tissues and also seem to be involved in innate immunity in the gastrointestinal tract (51).
The Phagocyte Oxidase NOX2
The best-characterized member of the NOX family is NOX2, which has been described not only in neutrophils, macrophages, DCs, and eosinophils but also in nonphagocytic cells such as fibroblasts, endothelial cells, and cardiomyocytes (75). In resting conditions, it is highly glycosylated and resides in intracellular secondary granules and, at lower amounts, in the plasma membrane, whereas activation leads to translocation to the plasma membrane and phagosomal membranes (99). During activation, the whole complex is assembled by the recruitment of the cytosolic components p40phox, p47phox, p67phox, and Rac toward the transmembrane component, the flavocytochrome b558 (Fig. 1).

Rac proteins are members of the family of small GTPase, cellular proteins that cycle between a GDP-bound inactive state (cytosolic) and a GTP-bound active state (membrane-associated). They have many disparate regulatory functions in phagocytes and are also key components of the NOX2 complex that control its assembly (reviewed in 16). Upon cell activation, GDP is exchanged by GTP by mechanisms involving the actions of GDP/GTP-exchange factors (GEFs). Rac binds directly to p67phox (37) and also to the membrane through its C-terminal domain, giving proper orientation to p67phox within the oxidase complex (77). Two Rac members, Rac1 and Rac2, participate in the NOX assembly in phagocytic cells, although their roles appear to be dependent on the cell type and the cell activation status. Rac2 was identified as a key player in the regulation of NOX2 in human neutrophils (72), and soon after, Rac1 was described in macrophages (1). In neutrophils, NOX2 activation is dependent on Rac2, whereas Rac1, even though expressed at similar levels, was proven to be dispensable for the complex activation and ROS production (48). Conversely, in macrophages, Rac1 is by far the predominant isoform, and studies carried out with cells from Rac2-deficient mice show that this isoform is necessary for NOX2 activation and ROS production under some circumstances depending on the activation stimulus (143). Recent work by Robin Yates' group has shown that the control of phagosomal functions, such as proteolysis, exerted by NOX2 was dependent on the activation stimulus in murine macrophages (6, 109).
Previous reports had already shown that, independently of the cell type, Rac1 is preferentially located at the plasma membrane, whereas Rac2 was present on endomembranes (87). This different localization was later explained by the different lipid composition of the membranes that regulates protein charge and thus affinity to membranes, conferring a specific localization (146). These findings may explain how the two Rac isoforms influence the localization site for the assembly of NOX2 in different cell types. This could be the reason why, in neutrophils, where Rac2 is the isoform involved in the complex, NOX2 is active mainly in phagosomes to exert its microbicidal function through the production of intraphagosomal ROS. In contrast, in macrophages, where Rac1 is the isoform involved in the complex, NOX activation is mainly driven to the plasma membrane. As a result, the main phagosomal strategy for pathogen killing in this cell type is the activation of their proteolytic activity through an important acidification rather than the respiratory burst. Interestingly, Allen and colleagues showed that infection of neutrophils with Helicobacter pylori retargets the NOX complex assembly from the phagosome to the plasma membrane as a survival strategy. The consequent release of ROS extracellularly provokes major tissue damage (2). A following study described that H. pylori is also able to specifically activate Rac1 in epithelial cells (67). This observation suggests that a similar situation may occur in H. pylori-infected neutrophils and may provide additional evidence that the activation of a specific Rac isoform would account for the localization of the NOX complex assembly.
Localization of NOX2 and ROS Production in Different Phagocytes
Neutrophils, macrophages, and DCs are all phagocytes that play different functions in the immune response. Neutrophils and macrophages are involved in the local inflammatory response and clearance that take place upon phagocytosis of pathogens or infected cells. Additionally, macrophages, although not efficient in activating naïve T cells, can activate memory T cells (105, 112). DCs are mainly responsible for processing and presenting derived antigenic peptides to naïve T lymphocytes, triggering the adaptive immune response.
In resting neutrophils, the flavocytochrome b558 is found mainly in the membrane of secondary granules (3, 66), whereas a smaller amount is located in the membranes of a subpopulation of peroxidase-negative granules (71). Upon activation, translocation of flavocytochrome b558 occurs when granules fuse with the phagosomal membrane or the plasma membrane (18, 28, 47). As a consequence, extensive phosphorylation of p47phox accompanies this translocation. The oxidase activity is initiated after the phagosome is formed and occurs intracellularly, while superoxide cannot be detected extracellularly (119). The NOX2 complex can be found at different states of activation: resting, primed, active, and inactive (39). The resting, unassembled oxidase is present in circulating cells and becomes primed upon a first stimulus, making the cell more susceptible to activation. Priming stimuli are neutrophil adhesion, bacterial lipopolysaccharides (LPS), and proinflammatory cytokines. Phosphorylation of the p47phox subunit is believed to be the key step to reach the primed state (40), which can be quickly activated by a second signal even at a suboptimal concentration. The active, assembled form is the one found in cells at inflammatory sites and can be induced by the pathogen itself or pathogen-derived products. Inactivation of the complex through dephosphorylation of some of the subunits is a consequence of the action of anti-inflammatory agents that limit inflammation (40).
Derived from circulating monocytes, macrophages are tissue-localized phagocytes that have specific features and functions according to their localization. Nonetheless, all types of macrophages express gp91phox and p22phox as well as the cytosolic subunits p47phox, p40phox, and p67phox. In monocyte-derived macrophages (10) and murine peritoneal macrophages (137), gp91phox was detected at the plasma membrane, but also in intracellular vesicular structures. Human monocyte-derived macrophages were described to recruit gp91phox to their phagosomal membranes after internalization of latex beads (84). Since macrophages lack specific granules present in neutrophils, the acquisition of NOX2 membrane components differs from the process described above. Phagosomes mature into phagolysosomes by sequential fusion and fission events exchanging membranes and soluble materials with endocytic compartments (36, 139). Maturation of phagosomes is driven by small GTPases of the Rab family. Early events are controlled by Rab5, which is replaced afterward by Rab7, to regulate the fusion with late endosomes and lysosomes (103, 140). Rab11 is involved in membrane recycling during this process (79). This model of phagosome maturation by sequential fusion with endosomes/lysosomes is generally accepted in the macrophage models. Nevertheless, this classical picture has been questioned by some studies showing a more complex dynamic of the phagosome maturation. Recently, the subcellular localization of NOX2 membrane components in macrophages has been described. Unassembled monomers localize to the endoplasmic reticulum (ER), and heterodimer formation is a prerequisite for efficient trafficking to target membranes (22). Using murine macrophages, the authors show that the flavocytochrome b558 localizes to the plasma membrane and to intracellular compartments, which are positive for Rab11. These recycle to the plasma membrane and reach nascent phagosomes from there, suggesting that the endocytic recycling compartment could be a reservoir of NOX2 membrane components in the same way that specific granules are thought to be the NOX2 storage site in neutrophils (22).
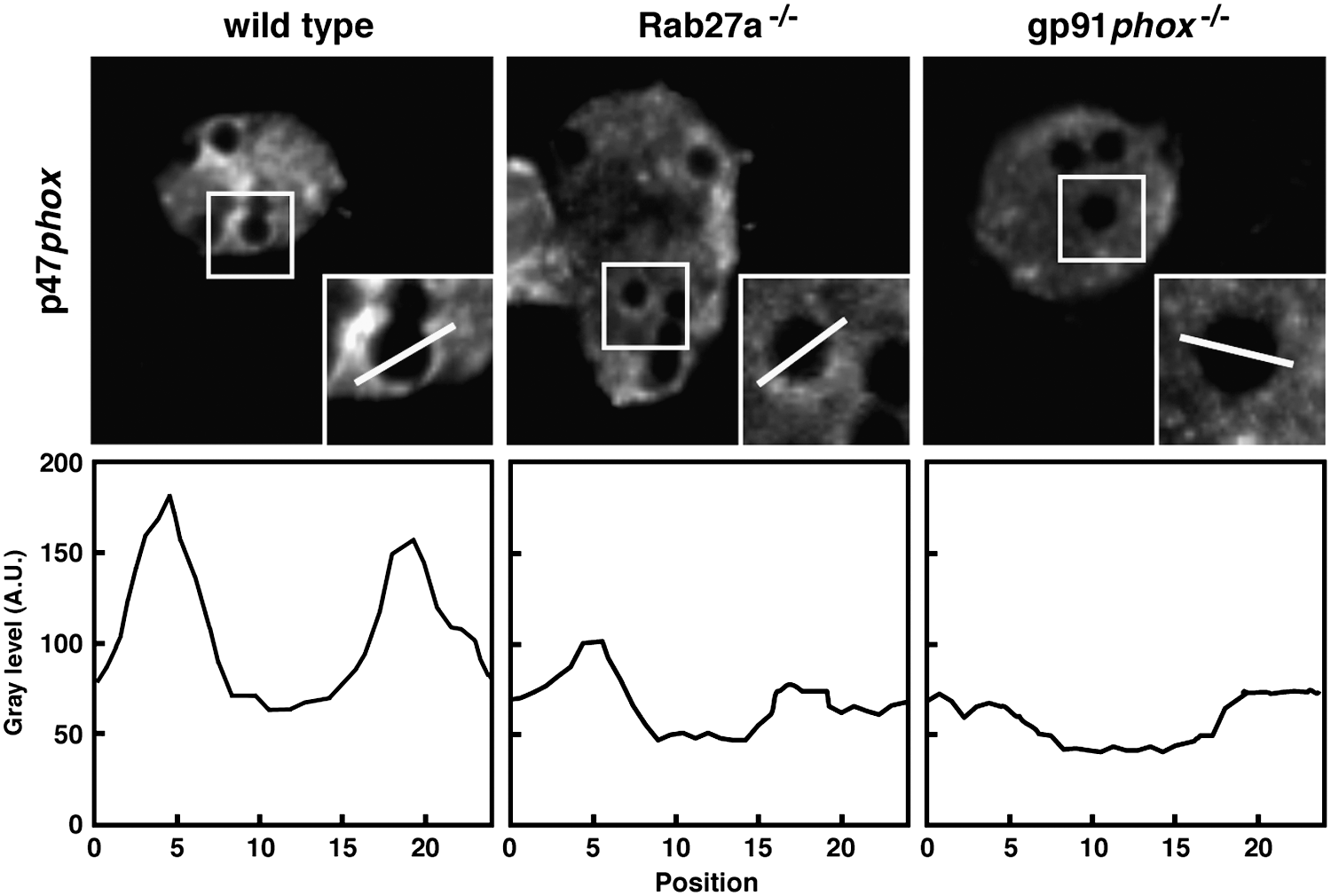
The generation of ROS in DCs was proposed by Matsue et al. (85), but Elsen and coworkers were the first to characterize the expression of the NOX2 complex in mouse splenic DCs (41). The very low amount of generated ROS led them to term this activity as cryptic, since the amount of O2 − and H2O2 secreted was <3% of that of neutrophils. They detected ROS production intracellularly only after concomitant treatment with the NOX activator phorbol-12-myristate-13-acetate (PMA) and toll-like receptor (TLR) agonists. Interestingly, the protein detected with the gp91phox antibody, which has a size of 90–100 kDa in human and bovine neutrophils, migrated with an apparent molecular mass of 60 kDa in murine DCs. The correct identity was confirmed, and the lower molecular mass explained by a difference in glycosylation (13). By immunoprecipitation and confocal microscopy, the authors showed the association of p40phox, p47phox, and p67phox in the cytosol and the localization of gp91phox and p22phox at the plasma membrane in resting DCs (113). A more detailed study of the subcellular distribution of NOX2 components in murine DCs was done later. In resting DCs, gp91phox was detected by confocal and immunoelectron microscopy at the membrane of vesicles throughout the cell (113). Furthermore, we also found gp91phox in Rab27a-positive vesicles that were also positive for the lysosomal marker LAMP1. These vesicles were recruited to and fused with phagosomes through a Rab27a-dependent mechanism (61). By using Rab27a-deficient DCs (isolated from ashen mice), it was shown that acquisition of the membrane components of the NOX complex by phagosomes was strongly delayed in absence of Rab27a. Recruitment of the flavocytochrome b558 to phagosomes was concomitant with the recruitment of p47phox to the phagosome membrane, suggesting a functional assembly of the complex. As a consequence and as shown in Figure 2, the activation of the complex measured by the phagosomal recruitment of the cytosolic p47phox was significantly affected in Rab27a-deficient DCs compared to wt DCs and totally absent in DCs from gp91−/− mice (Fig. 2). Similar results were obtained by functional assays, where the actual oxidative capacity of the phagosomal environment was measured by using FACS-based assays applying a ROS sensor bound to beads (113). NOX activity and ROS production were also described in human DCs. We have shown that after antigen internalization, NOX2 complex activation occurs at the membrane of antigen-containing compartments (84).
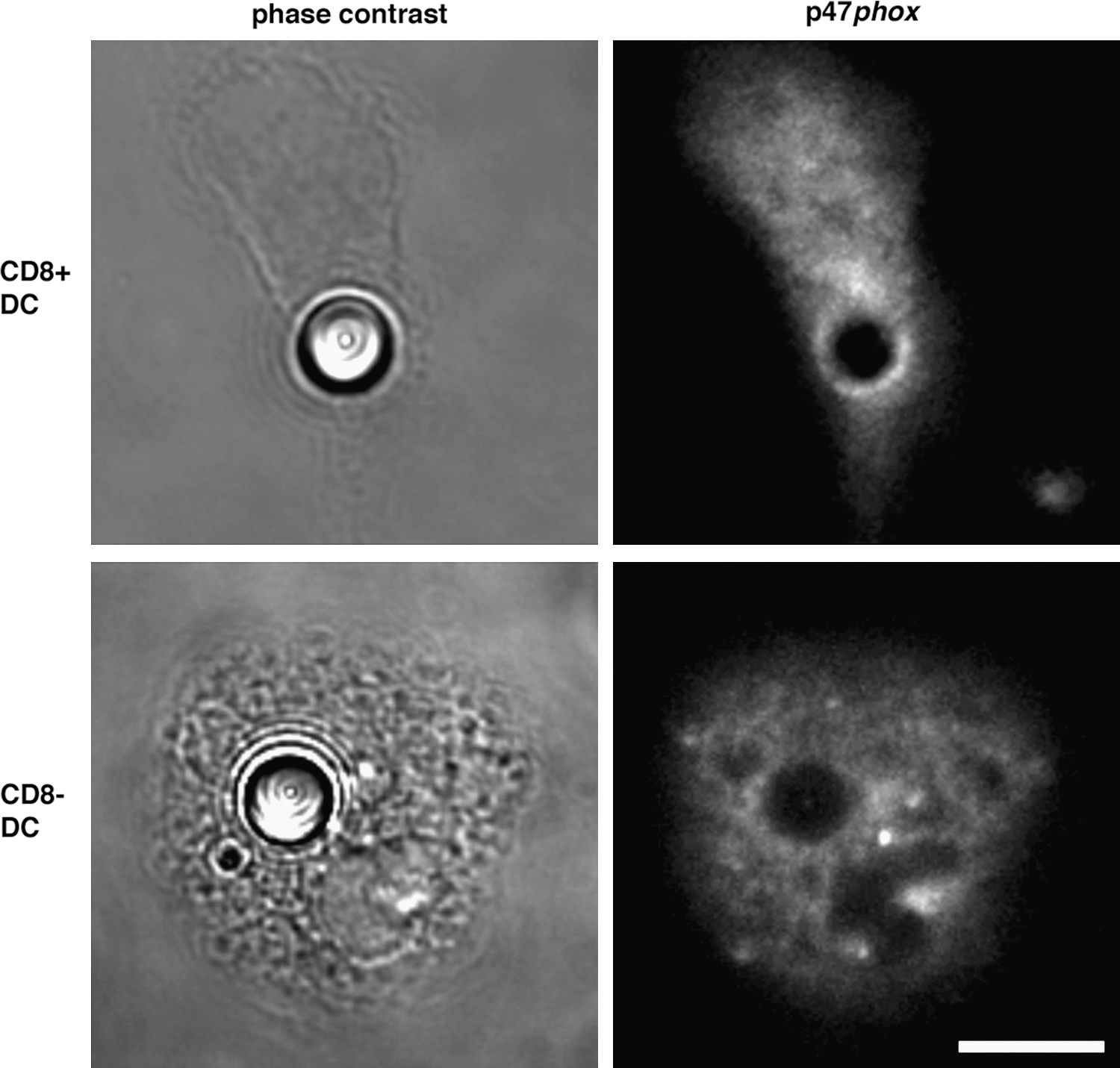
As explained in the Introduction, different DC subsets have been identified in mouse lymphoid organs according to the expression of specific surface markers (141). Splenic DCs can be divided into CD8+ and CD8− subsets, which have distinctive oxidative capacities caused by a different localization and assembly of NOX2 (114). CD8− spleen DCs produce more total ROS than CD8+ DCs, and both subsets from cybb−/− mice fail to show any activity, confirming that NOX2 is responsible for ROS production in both cases. Consistent with this, CD8− DCs express higher amounts of gp91phox. However, only CD8+ DCs were able to produce ROS at the phagosomal level, whereas phagosomes from CD8− DCs failed to produce ROS even after stimulation with PMA. CD8+ DCs recruit gp91phox and p47phox at the phagosomal membrane, whereas CD8− DCs do not recruit p47phox; thus, the NOX2 complex cannot assemble (Fig. 3). These differences can be explained by the role of Rac GTPases in the localization of the NOX complex (as described above). Although both cell types express the two isoforms, CD8+ DCs displayed higher levels of Rac2, whereas the expression of Rac1 was similar. After phagocytosis of latex beads, Rac2 accumulated strongly around the phagosomes of CD8+, but not CD8− cells, and Rac1 was detected at the plasma membrane, around phagosomes and in vesicles in both CD8+ and CD8− DCs. The presence of Rac2 at the phagosomal membranes exclusively in CD8+ DCs may account for the localization of the active NOX complex at the membrane of these intracellular compartments in this DC subset. In agreement with these findings, CD8+ DCs from Rac2-deficient mice (where only Rac1 is present) showed a defect in recruiting NOX2 at the phagosomal membrane, and the complex was relocalized to the plasma membrane (probably through a Rac1-dependent mechanism). These observations suggest that Rac2 drives the assembly of the NOX2 complex to phagosomes of CD8+ DCs, whereas Rac1 directs its activation to the plasma membrane in CD8− DCs. Given that both Rac isoforms are present in the different subsets, the differential expression may not be the only explanation for the different localization of the complex in the different cell types. It is most likely that the specific DC subset expression of Rac effectors, GEFs, and GTPase-activating proteins may account for favoring the activation of one Rac isoform or the other one. As in macrophages and neutrophils, these differences in the Rac-induced assembly of NOX2 among DC subsets would also reflect and contribute to their different functions during immune responses, as we will discuss later in this review.
Role of NOX2 in Immunity
The role of ROS produced by the NOX2 oxidase and those produced thereafter by other enzymes, such as myeloperoxidase, in bacterial killing has been extensively studied in neutrophils (reviewed in 75, 101). First evidence came from studies investigating immune function of patients suffering from chronic granulomatous disease (CGD). This disease results in chronic and repeating intervals of bacterial and fungal infections and has been characterized by the absence of establishing effective immune responses (9, 19). Previously, different studies have reported that CGD patients display mutations in genes encoding gp91phox, p22phox, p47phox, and p67phox (reviewed in 56). The recognition of bacterial products on the cell surface triggers engulfment and bacterial destruction through the generation of a respiratory burst, which is characterized by high oxygen consumption initiating a rapid increase of ROS inside the phagocytic vacuole. However, the direct effect of ROS in bacterial killing has been questioned (119). The modulation of the environment of the phagosome was proposed to be the key function of ROS in microbe killing. The ion flux that occurs to compensate the charges produced by the electrogenic NOX2 complex causes an increase in the ionic strength and together with the changes induced in pH, produce the release of granule proteins and ensure their correct function in microbe killing (102, 119). In addition, chemotaxis of neutrophils toward the site of infection has been also proposed to depend on NOX2-induced ROS generation, since neutrophils silenced for NOX2 or isolated from CGD patients have impaired migration (55, 74). The involvement of ROS in the expression of chemokine receptors could also contribute to its effect on chemotaxis (78).
Although ROS and the oxidative burst have been linked to the promotion of inflammation, increasing evidence points toward the idea that ROS act as a modulator of immune responses (reviewed in 111). Data showing that individuals with CGD are more prone to autoimmune diseases and that defective ROS production in neutrophils is linked to an increased inflammatory response (46) support the regulatory function of ROS in immunity using different autoimmune disease models, such as arthritis and multiple sclerosis (94).
ROS production by NOXs has been linked to several signaling mechanisms that have immunological consequences and has been extensively reviewed (44, 92). Signaling functions have been proposed to be the main role of the oxidative burst in macrophages, where most of the superoxide produced at the plasma membrane is dismutated outside the cells (45). A portion of the resulting peroxide could enter cells surrounding the site to exert immunological functions. Although the permeability of membranes to peroxide is limited, there are other mechanisms through which peroxide could enter cells in the small amounts that are necessary for signaling purposes (reviewed by 8, 11, 45, 111). Moreover, intracellular ROS have been shown to modify key factors that control the inducible expression of genes whose products are part of the inflammatory response (50, 131).
In conclusion, the killing of intraphagosomal bacteria within phagosomes, which is the main function of neutrophils and a key component of innate immunity, is dependent on the oxidative burst to provide the adequate environment for protease function. This is in agreement with the fact that most neutrophil NOX2 is present in granules of resting cells that access phagosomes soon after the internalization of microorganisms. The phagosomes of macrophages have a lower pH, which is considered necessary to exert its degradative functions. While their oxidative burst is lower than in other phagocytes, the control exerted by phagosomal ROS in antigen proteolysis has been studied in detail (107, 109, 145). Considering that NOX2 assembles also at the plasma membrane in macrophages, it is not surprising that ROS function as intra- and extracellular signaling molecules in macrophages. In contrast, the main function of DCs is to process and present antigens and initiate and shape adaptive immune responses (112). In this sense, it is reasonable to expect that NOX2 oxidase localization and production of ROS are regulated in a manner to serve this purpose. In the next sections, we will focus on the roles of NOX2 activity in antigen processing and presentation in DCs.
Role of Phagosomal ROS Production in Antigen Presentation in DCs
The phagocytic pathway of DCs, highly adapted to antigen processing, has been shown to display remarkable differences compared to other phagocytes. Contrary to macrophages and neutrophils, the main function of DC phagosomes is antigen presentation rather than pathogen killing or clearance of cell debris. Accordingly, their phagosomes are the source of peptides derived from extracellular antigens that will be processed and presented to T lymphocytes. As we mentioned earlier, DCs have the ability to present exogenous antigens to both CD8+ and CD4+ T cells. Activation of CD8+ T cells via cross-presentation leads to cytotoxic immune responses against a wide variety of exogenous antigens (including tumor cells, viruses, and bacteria). Similarly, cross-presentation is believed to play an important role in maintaining the equilibrium between tolerance and immunity through the presentation of self-antigens.
As explained in the Introduction, this pathway is almost exclusive to DCs, and the intracellular mechanisms are still not clear. After antigen uptake and probably a gentle early degradation, most proteins or polypeptides are transferred to the cytosol, where they are processed by the proteasome and then transported either back to the phagosome or to the ER to be associated to MHC class I molecules (reviewed in 4, 30). These MHC class I/peptide complexes are then finally transported and exposed at the cell surface to activate CD8+ T cells. In contrast to this so-called cytosolic pathway of antigen cross-presentation, a vacuolar pathway has also been described in which the antigen processing and loading onto MHC class I occur without any cytosolic steps, within the phagosome (25).
On the other hand, MHC class II-restricted antigen presentation is the process by which APCs activate specific CD4+ T cells. In this pathway, the phagocytosed antigen is degraded within the phagosome that matures and fuses with lysosomes that provide the necessary proteolytic enzymes. At the same time, MHC class II molecules assembled in the ER are targeted to the endosomal–lysosomal pathway by the invariant chain (Ii), a chaperone that is associated to the MHC class II dimer, which prevents premature peptide loading. Once MHC class II molecules reach phagolysosomes, Ii is cleaved, and peptides are loaded, and the MHC–peptide complex is transported to the plasma membrane (reviewed in 91). Interestingly, recent results also suggest a role of Ii in the regulation of cross-presentation itself (7). Therefore, antigen processing is of central importance in DCs. The loading of antigenic peptides on both MHC class I and II molecules requires tightly controlled proteolysis to preserve antigens from degradation. Several mechanisms have been identified to explain the limited phagocytic proteolysis in DCs, and a link between proteolysis, antigen processing, and antigen presentation has been proposed.
Early evidence of the regulation of the proteolytic activity among different phagocytes came from the work of Lennon-Duménil and coworkers, which assessed the protease activity of phagosomes in DCs and macrophages. They described that phagolysosomal fusion is selectively regulated among these different phagocytes, observing that in DCs, there is lower expression and recruitment of proteases, and that the kinetics of phagosome maturation are faster in macrophages as compared to DCs (80). Later, a study of lysosomal functions described that the inefficient lysosome acidification in immature DCs was due to limited recruitment of the V-ATPase to lysosomes, as compared to macrophages or mature DCs (135). Lower amounts of lysosomal proteases were also detected in DCs in vivo within mouse secondary lymphoid organs, as well as in bone marrow-derived DCs (34). Lysosomal proteolytic activity was decreased in murine DCs, as compared to macrophages, and lower proteolysis was the cause for enhanced antigen presentation. Further work showed that immunogenicity was enhanced by limiting the susceptibility to lysosomal proteolysis in vivo (33). These results suggested that antigen persists in a less-degradative phagosomal environment in DCs, which would favor their efficient presentation on MHC class I and class II molecules.
Further evidence of this specialization of endocytic and phagocytic compartments in DCs came from the observation that ROS production inside DC phagosomes regulates phagosomal properties. We showed that DC phagosomes containing latex beads actively alkalinize their lumen at pH values higher than 7 during the first hours after internalization (pH 7.5 after 60–120 min; 113). A clear alkalinization of phagosomes reaching values of pH above 7.0 was also registered using a DC cell line, DC2.4 (133). As originally proposed in neutrophils, ROS generation in DC phagosomes consumes an important amount of protons that have been provided by the V-ATPase. This H+ consumption provokes an increase in the intraphagosomal pH. Indeed, compared to primary macrophages and macrophage-like cell lines, DC phagosomes had higher pH values, and this increase was dependent on NOX2 activity, because gp91phox-deficient DCs had more acidic phagosomes. In fact, p47phox assembly at the membrane was more sustained over time in DCs than in macrophages. As a functional consequence of this particular phagosomal pH regulation, we showed that antigen cross-presentation of two different model antigens was decreased in DCs isolated from gp91phox-deficient mice. This was due to an increase in antigen degradation as a result of the altered pH values that induced higher activities of proteases that otherwise would be less active (113) (Fig. 4). The above-mentioned studies used polystyrene beads as phagocytic cargo, but similar observations were made in murine bone marrow-derived DCs and macrophages containing Staphylococcus aureus. While in macrophages containing S. aureus only low ROS levels were generated, and their phagosomes acidified very rapidly, the situation in DCs was different. DC phagosomes containing S. aureus exhibited much slower acidification profiles over time, whereas significantly more ROS were generated (Anna Sokolovska and Lynda M. Stuart, personal communication).

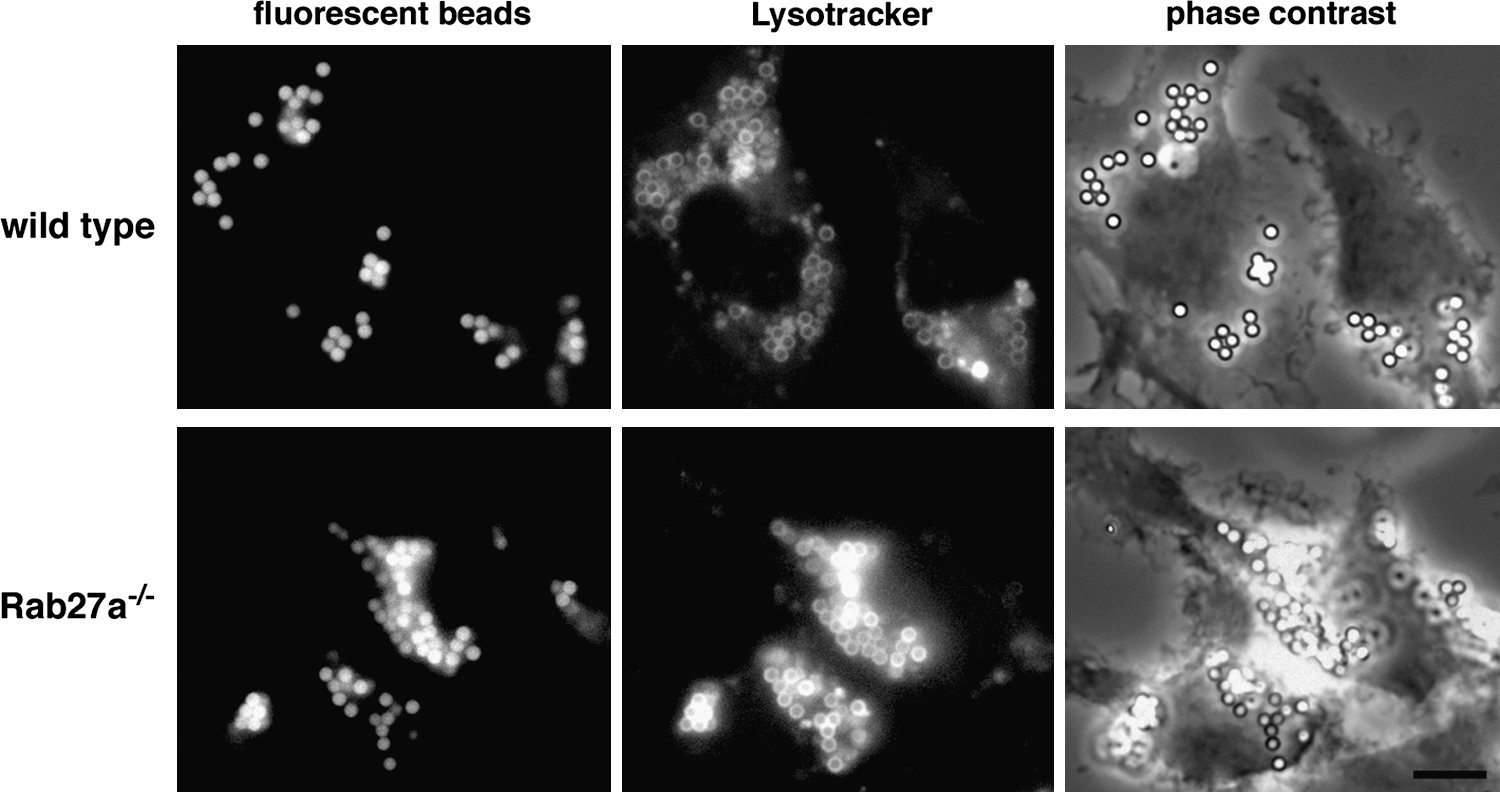
As mentioned above, the recruitment of vesicles containing NOX2 membrane components to phagosomes was dependent on Rab27a. Consequently, DCs from Rab27a-deficient ashen mice displayed a defect in antigen cross-presentation due to increased antigen degradation within more acidic phagosomes (61) (Fig. 5). Similar results were then described for human DCs. Monocyte-derived DCs from CGD patients displayed an internalization pathway with higher acidification, increased proteolytic activity, and impaired cross-presentation of antigens, as compared to DCs from healthy donors (84).
These studies supported the idea of a relationship between antigen presentation and the level of antigen degradation, the latter one being regulated by the pH and ROS within phagosomes. Further evidence for the involvement of NOX2 and ROS production on antigen presentation came from work in Vav-deficient DCs. Vav is a member of the GEF family that catalyzes the exchange of bound GDP to GTP on Rac proteins (29). After stimulation through a variety of immune receptors, Vav undergoes tyrosine phosphorylation that is required for its exchange activity on Rac (17). Their role in ROS production and the oxidative burst in macrophages had already been established (88, 89). Vav-null DCs exhibited defects in ROS production comparable to gp91phox-deficient DCs, but in contrast responded to stimulation by PMA, suggesting that NOX2 oxidase was functional (49). Analysis of phagosomal properties in these cells showed a decrease in pH values and an increase in antigen degradation. DCs failed to cross-present the antigen bound to latex beads efficiently (49), thus providing additional evidence of the link between NOX2 function and antigen cross-presentation.
In line with these findings, further research was carried out with different subsets of murine splenic DCs, which are characterized by different antigen presentation abilities; for example, CD8+ DCs are known to cross-present more efficiently compared to CD8−. This was shown to be partially due to the array of phagocytic receptors expressed (60, 118) and differences in their antigen-processing capacities (38, 117). Our group found that the pH was higher in CD8+ phagosomes compared to the ones of CD8− DCs. Consequently, the proteolytic activity in CD8− DCs was more dominant than in the CD8+ population. As described above, the differences observed in the regulation of phagoendosomal pH correlate with a different localization of the active NOX2 complex. High phagosomal pH in CD8+ DCs correlates with a specific assembly of NOX on phagosomal membranes, whereas an acidic pH in CD8− phagosomes corresponds to the absence of the NOX assembly in these compartments (Fig. 6) (114). As mentioned above, Vav1 regulates ROS production and cross-presentation in DCs (49). Vav1 was previously shown to associate with p67phox and Rac2, but not Rac 1, in human neutrophils (89). The absence of Rac2 in phagosomes from CD8− DCs could provide an explanation for the differences observed between DC subsets according to the differential expression of Rac isoforms. Surprisingly, CD8− DCs produce high levels of ROS at the plasma membrane that could serve other immune-related functions, such as extracellular signaling, chemotaxis, or modulation of the inflammatory environment.

Interestingly, our studies in DCs showed that the regulation of intralumenal pH for efficient antigen processing is not restricted to the phagocytic pathway. The endocytic pathway in DCs is also highly adapted to antigen processing, and a mechanism similar to that described for phagosomes probably accounts for the regulation of the proteolytic activity in endosomes. We have observed that the pH in endosomes is maintained around 7 and acidifies very slowly compared to those from gp91phox-deficient DCs (114, Fig. 4B). Indeed, the cross-presentation of soluble proteins is also impaired in DCs lacking gp91phox or Rab27a (61, 113). This was also confirmed in human DCs that displayed high co-localization of dextran (internalized by endocytosis) and p47phox, suggesting assembly and activation of the complex in endosomes (84).
Very recently, the group of R. Yates suggested a redox-based control instead of a pH-based regulation of phagosomal degradation in DCs (108). These authors observed strong phagosome acidification in mouse bone marrow-derived DCs (pH 5.5 after 10 min of internalization). These results resemble their previous data obtained in macrophages (109). The discrepancy between the results of Yates and coworkers and the result from others (49, 81, 133) and us (84, 113, 114) could be due to multiple technical differences in the protocols used for the generation of DCs and the measurement of the pH. Strikingly, Yates et al. used opsonized particles (IgG-coated beads) that undoubtedly trigger distinctive intracellular events that may impact differently on the phagosomal pH regulation. Indeed, these authors also do not find any difference in phagosome acidification between NOX2-defective and control DCs using IgG-coated beads, which is not unexpected since IgG-containing phagosomes acidified strongly even in wild-type cells (which is not the case for phagosomes containing non-opsonized particles). FcγR engagement is well known to enhance phagosomal acidification and fusion to lysosomes (62, 132, 134). In any case, it is noteworthy that in their assays for dynamic pH measurements (108, 144), the authors employed techniques not usually associated with DC manipulation, but with macrophages (DCs usually adhere very poorly to glass and do not resist washes).
In conclusion, there are a number of reports that point toward a regulation of antigen presentation by the phagosomal environment, specifically due to the effect of NOX2-generated ROS on phagosomal pH (49, 81, 84, 113, 114, 133). This is particularly clear in the case of antigen cross-presentation. The role of NOX2 and ROS in MHC class II-restricted antigen presentation by DCs has not been analyzed in detail so far. Ebselen, a selenium-containing antioxidant compound that inhibits NOXs, reduced the presentation of ovalbumin (OVA) to CD4+ T cells by DCs (85). Maemura et al. used Kupffer cells, antigen-presenting macrophages that reside in the hepatic sinusoids, to assess the role of ROS in antigen presentation. They found that OVA presentation to antigen-specific CD4+ T cells was inhibited when blocking ROS production with diphenylene iodonium (DPI), an inhibitor of NOXs, although the role of ROS on antigen degradation and the phagosomal environment in this process was not investigated. They concluded that the effect of ROS on antigen presentation was mainly due to its role as a second messenger (83). The work done by our group has shown that presentation of OVA and HY antigens to CD4+ T cells was also defective in gp91phox-deficient DCs and, to a lesser extent, in Rac2-deficient DCs (114). In addition, Vav-null DCs were also impaired in the presentation of bead-bound OVA to antigen-specific CD4+ T cells (49).
There is still a controversy about the compartments dedicated to MHC class I presentation. It is not clear whether they are different from those dedicated to MHC class II-restricted presentation, or if they represent two different maturation states of the same compartment (20, 120). In any case, the requirements for pH, ROS, and protease activities seem to be different. The results showing that ROS production is required for MHC class II presentation did not assess the role of ROS in antigen processing. Since ROS is also necessary for cross-presentation, it would be possible to speculate that ROS serve different functions in these two pathways, neutralizing the pH to avoid excessive proteolysis in the case of cross-presentation and regulating other aspects of the MHC class II presentation pathway (Fig. 7).

Effects of ROS on Antigen Presentation Independent of the pH
Until now, the study of the role of ROS production by NOX2 in antigen presentation by DCs has been focused on its impact on phagosomal pH and consequently on antigen fate. Most of these data support a regulatory function for ROS production to moderate the proteolytic environment by neutralizing the pH, thereby favoring antigen cross-presentation.
The fact that DCs assemble and activate NOX2 and produce ROS constitutively at low levels suggests that ROS could have various biological functions in DCs. Besides the regulation of the pH in the internalization pathway, ROS may also modify, through direct oxidation, the molecular structure of the antigen, as well as proteins implicated in phagosomal functions. ROS are a group of highly reactive molecules with superoxide being the main component produced by the NOX2 oxidase. Superoxide is very rapidly converted into hydrogen peroxide, either spontaneously or by the enzymatic action of the superoxide dismutase, which can be found in phagosomes. Moreover, H2O2 also interacts with superoxide directly to produce singlet oxygen (52). Proteins containing cysteine residues are susceptible to oxidation by H2O2, since the thiol group at the side chain of cysteine can be oxidized to form a disulfide bond. If these cysteine groups are within active sites of enzymes, oxidation can change conformation and function of certain enzymes in a reversible fashion (12, 110). In this sense, it has been shown previously that the V-ATPase could be oxidized to form a disulfide bond between two cysteine residues within the nucleotide-binding subunits of the enzyme, resulting in the reversible inactivation of the complex (42, 43). Likewise, singlet oxygen can inhibit serine and cysteine proteases (cathepsins B, L, and S, as well as different caspases), but not aspartate proteases (cathepsins D and E) and metalloproteases (90, 130). The proposed mechanism includes the action of singlet oxygen modifying an essential residue in the catalytic center that is common to cysteine proteases and prone to oxidative changes. Cysteine proteases play a key role in antigen degradation and are subject to a complex regulation within the phagosome (57).
In addition to these possibilities, ROS could also have a regulatory impact on antigen presentation by direct oxidation of the antigen. This may induce molecular changes in its structure that might have an impact on the susceptibility to proteolytic degradation. One of the examples of regulation through oxidation of substrates is the production of the oxidized form of low-density lipoprotein by Toll-like Receptor (TLR)4-activated NOX1 in macrophages, a process that accelerates their conversion into foam cells, resulting in plaque formation and atherosclerosis (23, 96). Cancer progression is another example of how ROS can regulate proteolysis. In many cancer types, altered ROS production has an impact on protease function and substrate susceptibility to their activity, which leads to an increase in extracellular matrix degradation and tumor invasion (reviewed in 125). In a recent study, presentation of OVA to T cells by DCs was enhanced by hypochlorous acid, a byproduct of ROS and myeloperoxidase, found in primary azurophil granules of neutrophils. Chlorination of OVA promoted its binding to cell membranes, thus augmenting antigen uptake, as well as increased susceptibility to proteolysis due to conformational changes in the protein structure (100). Modifications of antigen conformation due to ROS and other byproducts could therefore influence protein susceptibility to proteolysis, affecting their transport to the cytosol or other steps of antigen processing. In line with this, the group of P. Cresswell has shown that the gamma-interferon (IFN)-inducible lysosomal thiol reductase (GILT), an antioxidant enzyme present in the endocytic pathway, favors presentation of antigens containing disulfide bonds that upon reduction are processed to expose epitopes for MHC class II binding and are transported more efficiently to the cytosol for cross-presentation (54;124).
Additionally, to the direct effect of ROS on proteins due to their reactive nature, there are different indirect pathways by which ROS can regulate cell functions (reviewed in 73). As mentioned before, ROS can inhibit phosphatases through redox-sensitive cysteine residues, and tyrosine phosphatases control the phosphorylation state of numerous signal-transducing proteins (128). The group of tyrosine phosphatases is inactivated by H2O2, and this results in the activation of several protein kinase pathways, which are normally inhibited by tyrosine phosphatases. These are mechanisms where ROS act as second messengers in signaling pathways, since they do not have the chemical structure to allow classical receptor ligand recognition.
Autophagosomes are a source of intracellular antigens for MHC class II-restricted presentation (95). Autophagy plays an important role in antigen presentation (reviewed in 31). In a starvation-induced autophagy model, superoxide and H2O2 lead to autophagy by inhibiting the Atg4 cysteine protease oxidizing reversibly the cysteine residue within the enzymatic active site (116). Atg4 is involved in the regulation of LC3 lipidation, a post-translational modification critical for autophagy. Even though the source of ROS during starvation-induced autophagy is not from the phagosome, but mostly the mitochondria, it provides another example of how ROS can modulate diverse cellular events through chemical modifications of enzymes. Recent work published by Huang et al. showed that in the absence of NOX2 or after treatment of cells with DPI, LC3 recruitment to the phagosome is impaired after microbe-induced autophagy by bone marrow-derived neutrophils (59). The authors hypothesized that ROS are mediating the assembly of the Atg5-Atg12-Atg16 complex, which allows the localization of LC3 to the phagosomal membrane.
It is also tempting to speculate that ROS can influence other events of the intracellular membrane-trafficking machinery that have been shown to be critical for antigen processing, notably for cross-presentation. Very recently, our group has shown that ER proteins are recruited to the endophagocytic pathway through the intermediate compartment (ERGIC), allowing the molecular machinery of MHC class I-restricted presentation to access the phagosome (24). This recruitment also delays the fusion to lysosomes, favoring the controlled degradation of antigens and resulting in efficient cross-presentation. It would be very interesting to investigate whether the recruitment of ER proteins to DC phagosomes is somehow influenced by ROS, which diffuse or leak out of phagosomes and could act on cytoplasmic proteins that could function as effectors or activators of Rab proteins or SNAREs that constitute the essential molecular machinery for intracellular membrane fusion (reviewed in 127).
TLR Signaling and ROS Production in DCs
In addition to sensing and clearing pathogens, the innate immune system has developed different strategies for presentation of antigenic peptides to induce adaptive immune responses. TLRs are a type of pattern recognition receptors (PRRs) that identify pathogen-associated molecular patterns (PAMPs). Innate immunity and the functions of NOXs are closely coupled. Important evidence came from observations in CGD patients lacking the establishment of effective immune responses against invading microorganisms (reviewed in 93). The impact of TLR signaling and proinflammatory pathways on the activation of NOXs has been reported (86, 98), suggesting a critical role of NOX isoforms and ROS production.
Cell activation can prime the NOX2 complex, which is accompanied by protein phosphorylation and translocation. This complex assembly initiates a weak oxidative response, but the priming stimulus is not sufficient to induce the oxidative burst itself (111). Priming stimuli include TLR agonists (LPS and flagellin), proinflammatory cytokines (TNF-α and Il-1β), and proteases (39). The relationship between host immunity, TLR signaling, and ROS generation has also been found for other members of the NOX family. Kawahara et al. have shown that NOX1 induced ROS production in a TLR4-dependent manner during H. pylori infection (67). Furthermore, detection of Salmonella enteritides flagellin by TLR5 also exhibited higher ROS generation by NOX1 in intestinal epithelial cells (68). NOX3 is very abundant in lung tissues, and one report has shown that deficiency in TLR4 induced upregulation of NOX3, resulting in increased ROS generation and pulmonary emphysema (147). NOX4 is an important effector downstream of TLR4 signaling, which includes direct interaction between the two molecules and NFκB-dependent proinflammatory responses upon LPS engagement (97, 98). Additionally, this oxidase is implicated in the Toll/IL-1R domain-containing adaptor-inducing IFN-ß (TRIF)-mediated activation of the transcription factor IRF-3 (27), which is essential in the induction of interferons and IFN-stimulated genes.
As mentioned above, several pieces of evidence support a relationship between TLR signaling and different NOX family members in innate immunity. Microbial molecules trigger TLRs, favoring ROS production that results in pathogen killing and inflammation. However, the selective regulation and subcellular localization of NOXs in phagocytes pinpoint to an essential link between innate ROS generation and adaptive immune responses and put DCs in the focus. The work of others and us have shown that these cells control the phagosomal environment toward one of their main functions, the presentation of antigenic peptides on MHC I and MHC II molecules (112). Could TLR engagement in DCs contribute to a particular NOX2 regulation in phagosomes with different consequences in antigen processing through pH modulation (Fig. 7)? Little is known about the impact of TLRs on ROS generation in DCs, but different scenarios can be hypothesized. TLR stimulation causes DC maturation, which includes long-lasting changes in their efficacy to process and present antigens. Trombetta and colleagues have shown that the V-ATPase was more efficiently assembled on lysosomes, and its activity was enhanced after stimulation with LPS (33, 34, 135). These results suggest that TLR4 stimulation has an impact on phagosome functions during DC maturation. The modulation of phagosomal functions at early time points by TLR4 stimulation has been a matter of debate (15, 106), and the eventual role of NOX2 in this system has not been evaluated. Compared to neutrophils, DCs display lower NOX activity (41). However, it has been shown that NOX activity in DCs is increased either by the interaction with TLR ligands (142) or by the interaction with antigen-specific T lymphocytes (85). It is therefore likely that TLR engagement upon phagocytic uptake of microorganisms as well as phagosomal NOX activity need to be tightly controlled to allow effective antigen presentation.
The existence of different pathogens that inhibit NOX activity preventing ROS production could be used to gain insight into the relationship between TLRs and NOXs in terms of antigen presentation. Some pathogens such as Legionella pneumophila, Coxiella burnetii, and Yersinia pseudotuberculosis are able to prevent the NOX complex assembly by inhibiting accumulation of its cytosolic components (53, 123, 126). Others, such as Salmonella enterica (138), cause a mislocalization of the NOX, whereas Leishmania donovani abrogates the recruitment of specific cytosolic components of the NOX complex (82). It is not clear and will undoubtedly require exhaustive study whether pathogens can influence cross-presentation through the manipulation of NOX2 activity in DCs.
Differentiation of self from non-self is of essential importance to mount immune responses. An interesting concept in this field adds more complexity to this general picture. The hypothesis of phagosomal autonomy proposes the autonomous control of antigen processing and presentation within microbe-containing phagosomes, but not in other phagosomes within the same cell (14). The involvement of TLRs in phagosomal autonomy and the detection of microbial nonself-, but not self-antigens, by TLRs remain a matter of debate (15, 106). However, it becomes evident that innate sensors do modulate the kinetics and efficacy of phagosome maturation and therefore antigen processing and loading in a phagosome-autonomous manner. This would represent a fine discrimination mechanism at the cellular level that would allow the preferential presentation of PRR-associated antigens. Considering the impact of ROS production in antigen processing, it will be of interest to investigate whether phagosomal NOX2 and the different Rac isoforms are regulated autonomously, and if this is modulated by TLR signaling (Fig. 7).
Closing Remarks
NOX2 is known to play a major role in innate immunity for several decades. In the last few years, however, it has become clear that NOX2 is also involved in the establishment of adaptive immunity. Its role in the control of phagosomal pH, and thereby of the efficacy of antigen cross-presentation in DCs, represents a clear illustration of how NOX2 can influence CD8+ T cell responses. Interestingly, CGD patients do occasionally develop autoimmune pathologies, suggesting again a role for NOX2 in adaptive responses. In the next years, it will be interesting to unravel possible mechanisms involved and to find other possible connections between NOX family members and adaptive immune responses.