
Abstract
Introduction
A chloroplast from a plant leaf has a complex ultrastructure (Fig. 1A). Inside, there is the thylakoid membrane that performs the primary events of oxygenic photosynthesis. The thylakoid membrane forms cylindrical stacks known as grana that are connected by the circular stroma lamellae (6). It is surrounded by a compartment known as the stroma, where many metabolic processes occur. These include the Calvin-Benson cycle for carbon dioxide fixation (39). Both the stroma and the thylakoids are enclosed by the envelope membrane, which consists of an inner and an outer membrane separated by an intermembrane space. The envelope has a complex system of transporters for transferring proteins and metabolites between the cytoplasm and the chloroplast (30). The thylakoid membrane encloses a compartment called the lumen, which is essential for photosynthetic electron transport and for proton gradient formation during photosynthesis. However, the functions of most of the lumen proteins are currently unknown (48).
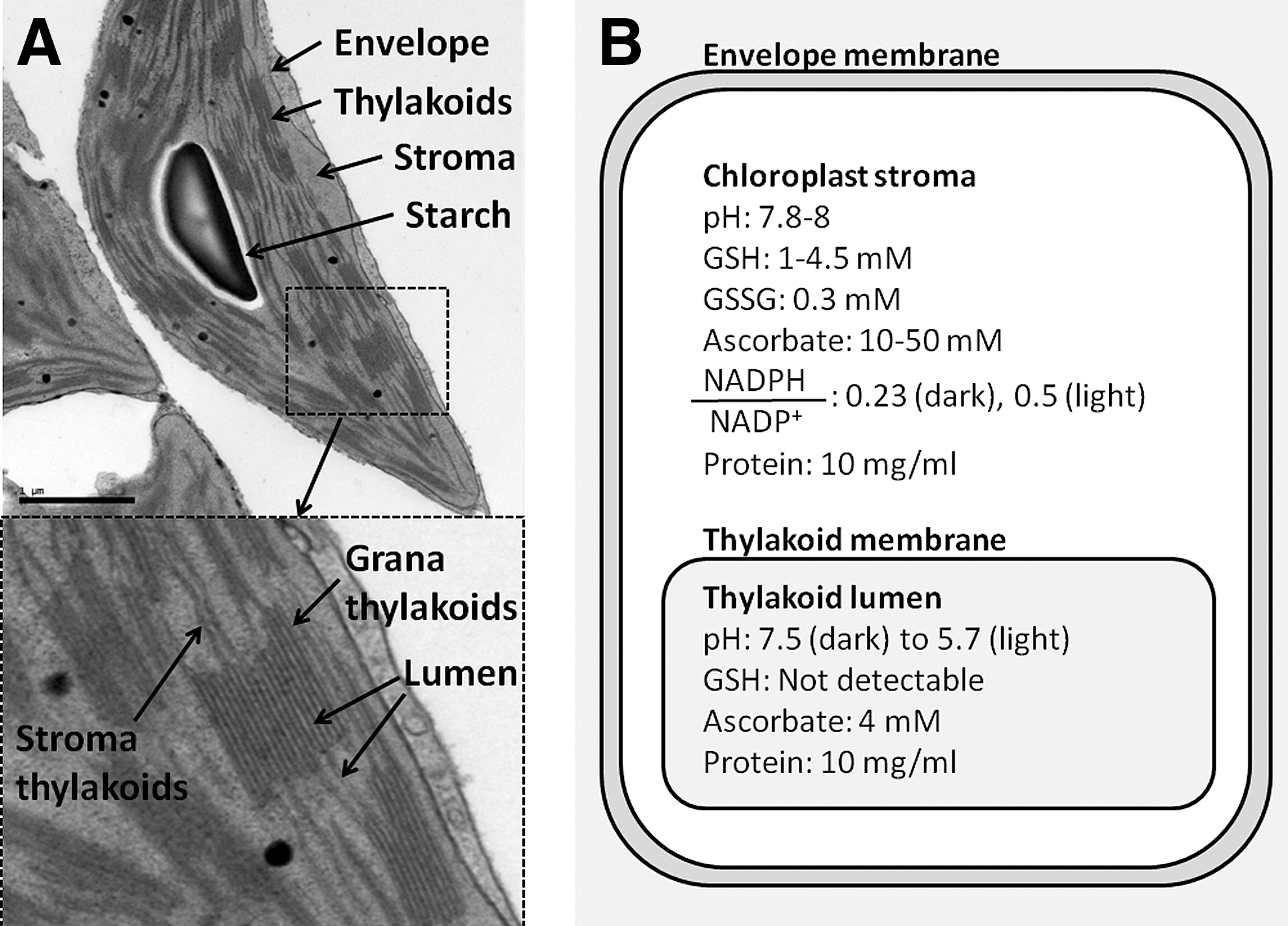
Different lines of research have suggested that chloroplasts can catalyze the formation of disulfide bonds in proteins, a process that is also referred to as oxidative folding. Notably, photosynthetic activity regulates the activity of enzymes in the chloroplast stroma due to the reduction of disulfide bonds via a cascade involving ferredoxin, ferredoxin thioredoxin (Trx) reductase, and Trxs m and f (75). During the last decade, more than 100 new potential Trx targets have been discovered in chloroplasts (57). Although the chloroplast stroma is reducing (Fig. 1B), the disulfides of these proteins are at least partially oxidized. Some of these proteins were also identified during in vivo screens for disulfide-bonded proteins in Arabidopsis thaliana (2, 53). In addition, chloroplasts from transgenic plants can produce functional therapeutic proteins that have correctly formed disulfide bonds (24, 80, 86). Overall, these findings indicate that chloroplasts can catalyze disulfide bond formation in proteins (88).
While many studies have addressed the function of disulfide bonds in the redox regulation of chloroplast enzymes and signal transduction (13, 54), relatively few have examined the mechanisms of disulfide bond formation in chloroplast proteins (42). This review provides an overview of the current knowledge in this field, with particular emphasis on oxidative folding in the chloroplasts of green plants. The mechanisms of disulfide bond formation in chloroplast proteins are not well understood. However, there is evidence that most chloroplast members of the Trx superfamily from the model plant A. thaliana are known, and some of them probably have functions related to oxidative folding. These include the protein disulfide isomerase PDIL1-3 (At3g54960) and the Lumen thiol oxidoreductase 1 (LTO1, At4g35760), which may be a useful starting point for future studies.
Chloroplast Protein Import Pathways Indicate that Disulfides Are Formed at Different Subcellular Sites
The folding of chloroplast proteins is closely linked to their biosynthesis and subsequent transport to their subcellular locations. Although chloroplasts have retained their own small genomes, most chloroplast proteins are nuclear-encoded and synthesized in the cytoplasm. They reach their subcellular destination by importation into the organelle (Fig. 2) (45).
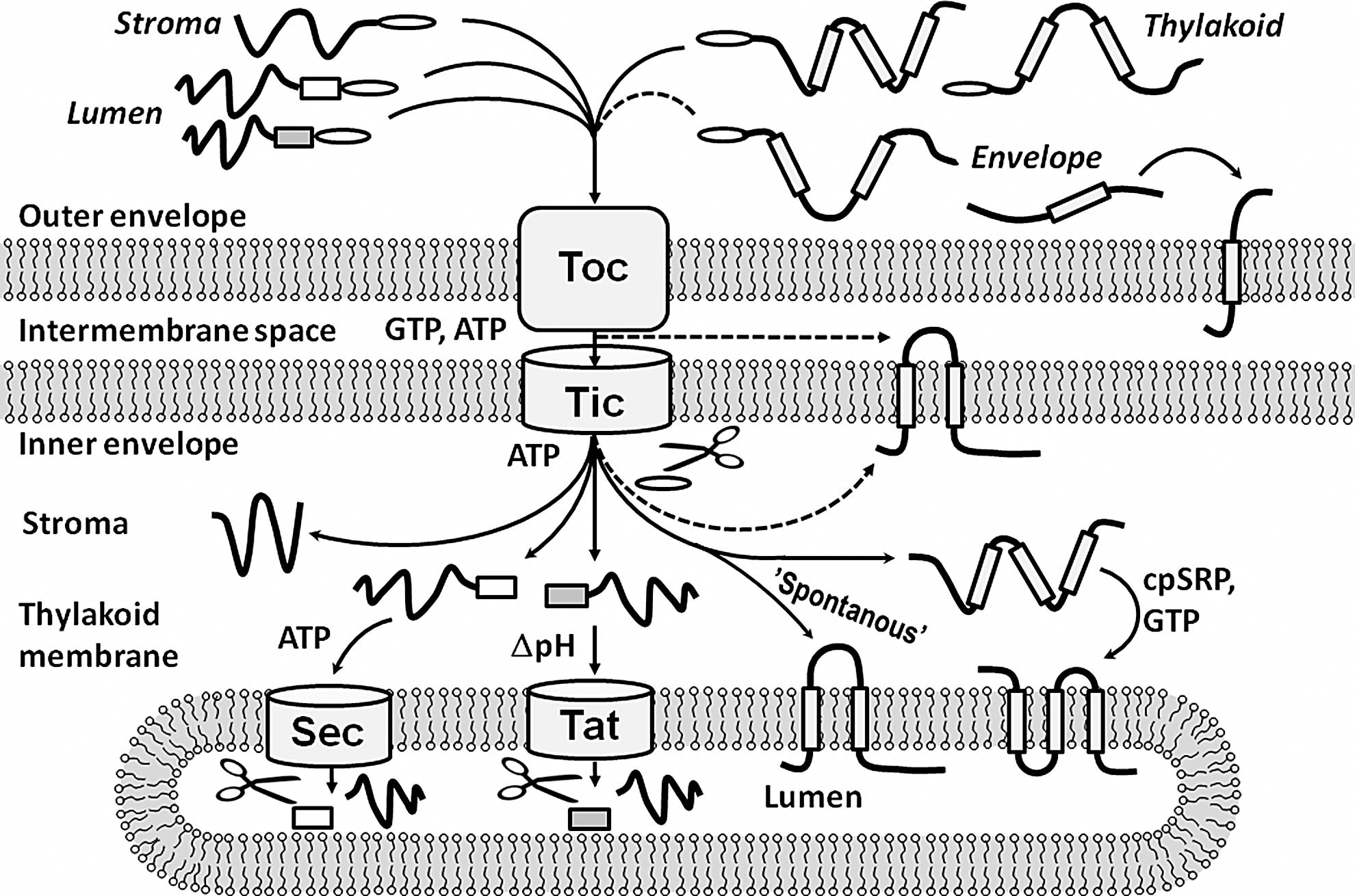
Most outer envelope proteins can be directly inserted into the membrane from the cytoplasm, but proteins with subcellular locations inside the chloroplasts are imported via the translocation systems of the envelope (Fig. 2). Usually, the cytoplasmic protein precursors have cleavable transit peptides with a chloroplast-targeting sequence that binds to receptors in the outer envelope. The protein precursors are then unfolded and enter the pore of the translocon outer complex. Finally, Hsp93 chaperones in the stroma pull the unfolded protein precursors through the translocon inner complex (TIC) (76). On the stromal side of the inner envelope, the stromal processing protease removes the chloroplast transit peptides of the imported precursors. At this point, the imported proteins may fold or be delivered to one of the chloroplast membranes (45).
Proteins of the inner envelope are exported to the inner envelope membrane from the stroma. Alternatively, they can exit the TIC laterally and insert into the inner membrane of the envelope from the intermembrane space (45). Proteins that are targeted to the thylakoid membrane usually insert into the membrane without apparent assistance by other factors in a so-called spontaneous way. However, the light-harvesting complexes are inserted into the membrane by the chloroplast signal recognition particle (45). The thylakoid membrane has two distinct import systems for lumen proteins (1). The Sec pathway, which resembles the bacterial SecYEG channel, imports unfolded precursors and requires ATP for the translocation. The twin-arginine translocation pathway imports folded proteins and is driven by the pH gradient across the thylakoid membrane. Lumenal protein precursors are tagged with a transit peptide that determines which of these import pathways will be used (1).
Both the envelope and the thylakoid membrane have the ability to unfold proteins for translocation. Although the envelope can import small folded proteins (18), protein precursors usually translocate the envelope in an unfolded conformation (15, 74, 87). It is likely that disulfide bonds are reduced when this happens because the precursors enter the chloroplast from the reducing environment of the cytoplasm. That implies that the disulfide bonds of newly imported chloroplast proteins are formed at their final subcellular location.
Proteins containing disulfide bonds have been observed in most of the compartments and membranes of the chloroplast, including the stroma (2, 53, 75), the thylakoid membrane (62, 64), and the lumen (2, 34, 53, 82). There is also evidence that proteins with disulfide bonds are present in the inner (9) and outer envelopes (10). The only chloroplast compartment that is not known to contain proteins with disulfide bonds is the intermembrane space. It is not currently known whether disulfide bonds are formed in the intermembrane space, but it is likely that both the stroma and the thylakoids contain enzymes that catalyze disulfide bond formation. The heat shock protein Hsp93 is a component of the import apparatus of the inner envelope that promotes the correct folding of imported proteins. Because the importation and folding of chloroplast proteins are closely related, Hsp93 may have co-chaperones that assist in the formation of disulfide bonds in newly imported proteins. Such co-chaperones would represent a sort of missing link, but they have not yet been observed in plant chloroplasts.
The folding of proteins that have multiple disulfide bonds requires both disulfide bond-forming activity and also isomerase activity to reshuffle incorrect disulfide bonds and establish the required connectivity. A representative disulfide-bonded protein whose folding requires both of these activities is the basic pancreatic trypsin inhibitor BPTI (aprotinin), which has three disulfide bonds. The chloroplasts of transgenic tobacco lines can synthesize BPTI in the stroma and export it to the lumen, where it is folded with the correct disulfide bonds (86). This suggests that the lumen has an enzymatic activity that can form and isomerize protein disulfide bonds, which raises a question: Do any known plant chloroplast enzymes have such functions?
The Chloroplast Members of the Trx Superfamily in A. thaliana
Many enzymes that catalyze disulfide bond formation in proteins are members of the Trx superfamily. A survey of the Trx superfamily from Arabidopsis shows that it has 34 members with a confirmed location in chloroplasts (Fig. 3). Most of these proteins are located in the stroma and/or peripherally associated with the envelope and the thylakoid membrane. This group includes 10 Trxs (4, 14, 19, 20) and 9 Trx-like proteins (14, 23, 61, 71). In addition, there are four glutaredoxins (8, 16, 21, 22) and peroxiredoxins (7, 11, 43). Finally, there are four protein disulfide isomerases (3, 14, 44) as well as the proteins low quantum yield of Photosystem II 1 (LQY1) and snowy cotyledon 2 (SCO2), which lack Trx active sites but have protein disulfide isomerase activity (58, 78). Only three known members of the chloroplast Trx superfamily are located in the thylakoid membrane or the lumen. These proteins include the Trx-like protein Hcf164 (55), the LTO1 (29, 46), and the lumen-located peroxiredoxin Q (48, 68). No Trx-like proteins have been reported in the envelope.
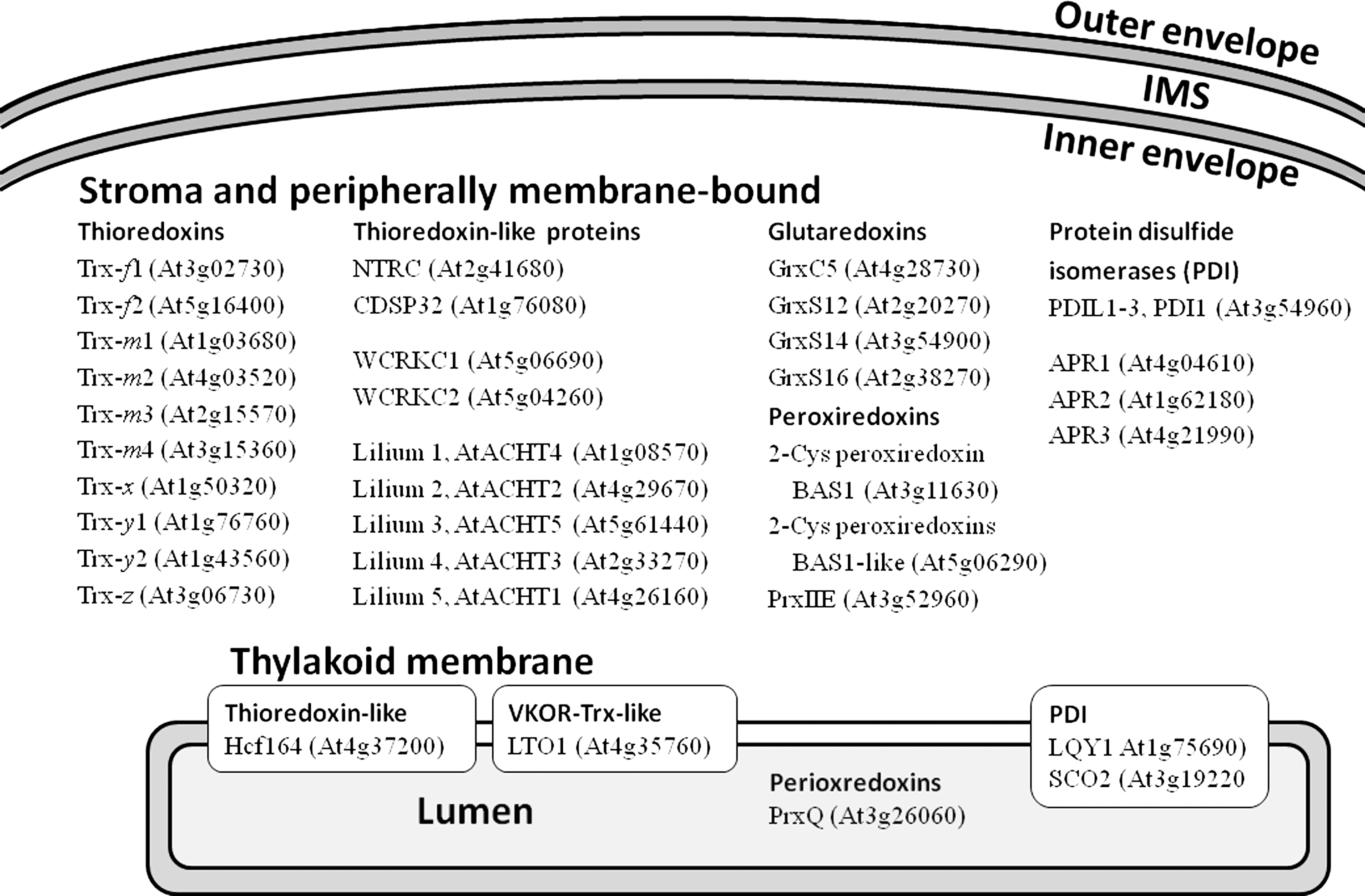
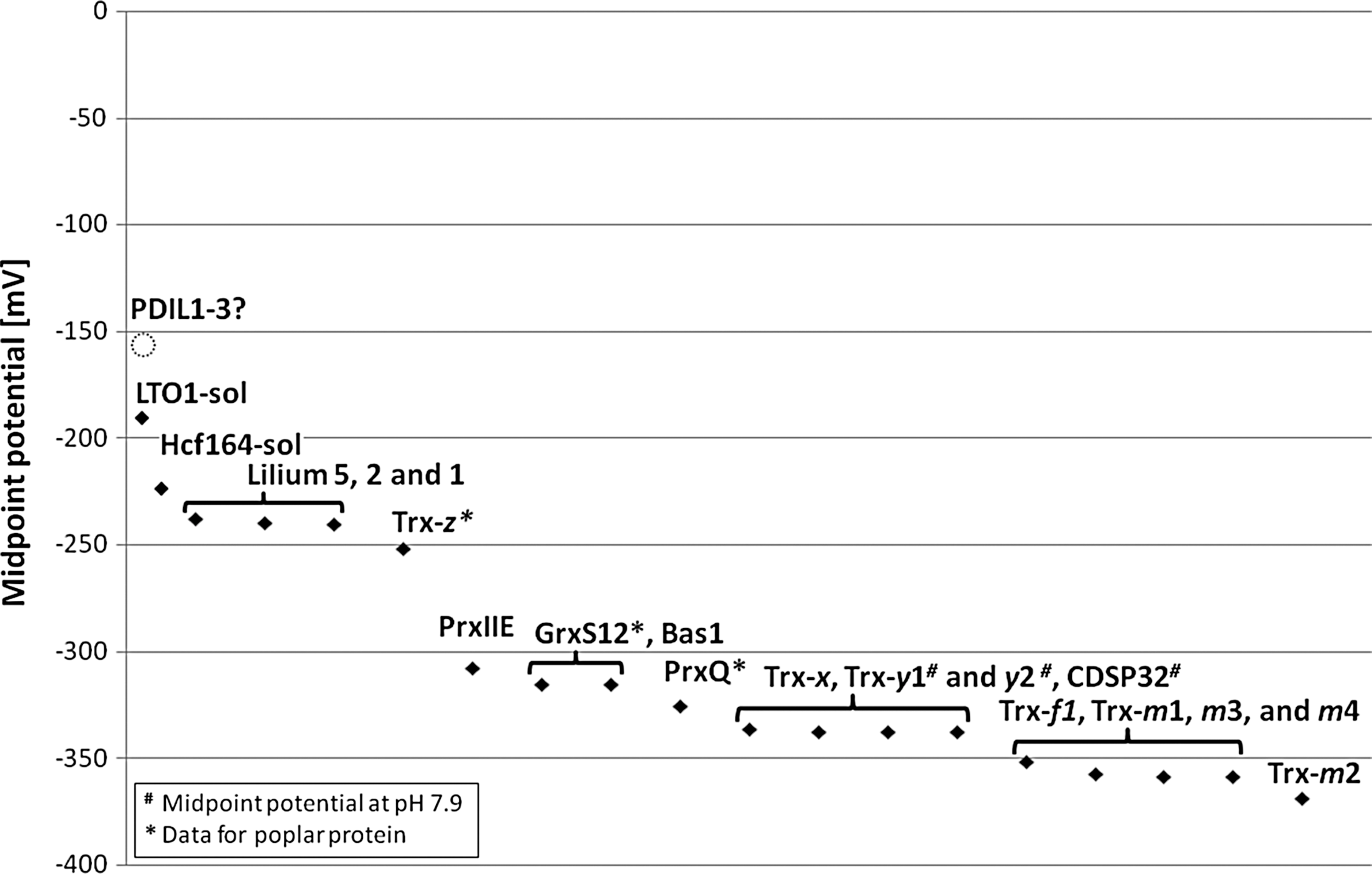
Some of the chloroplast members of the Trx superfamily have been studied in detail. For instance, the impact of Trxs and peroxiredoxins in redox regulation of chloroplast enzymes and in antioxidative defense has been highlighted in many excellent articles and some recent reviews (13, 26, 50, 54, 75). However, no functions have been identified for some other members of the Trx superfamily, including the WCRKC proteins (14), and the lilium proteins (23). The only chloroplast proteins for which disulfide bond-forming activity has been reported are the LTO1 (29, 46), and the thylakoid proteins LQY1 (58) and SCO2 (CYO1) (78). The soluble Trx-like domain of LTO1 (starting at L231) was determined to have a midpoint potential of −180 mV (32), which matches that for typical protein disulfide isomerases (5). Can midpoint potentials help to elucidate which other proteins of the Trx superfamily of Arabidopsis chloroplasts might function in disulfide bond formation? While the biological function of a dithiol-disulfide oxidoreductase can certainly not be inferred from its in vitro midpoint potential alone, it may provide a helpful suggestion concerning a protein's function in vivo.
A survey of midpoint potentials reported in the literature for the chloroplast-located members of the Trx super family from Arabidopsis (Fig. 4) revealed values ranging from −368 mV for stromal Trx-m2 (19) to −180 mV for the soluble Trx-like domain of LTO1 (32). After LTO1, the highest midpoint potentials in this group are those of the soluble part of the thylakoid protein Hcf164 (residues 116 to 261; −224 mV) and the stroma-located lilium 5 (AtACHT1; −237 mV) (23, 62). These values are lower than those of protein disulfide isomerase (−180 mV) and DsbA (−124 mV), which are both known to be involved in disulfide formation (5). This suggests that the chloroplast Trxs, the Trx-like lilium proteins, CDSP32, and the peroxiredoxins have no direct roles in the formation of disulfide bonds in proteins. The midpoint potential of NADPH-dependent thioredoxin reductase (NTRC) has not been reported, but NTRC can efficiently reduce the peroxiredoxin Bas1 (67), and must therefore have a lower midpoint potential. Some of the midpoint potentials for the glutaredoxins GrxC5, GrxS12, GrxS14, and GrxS16 have also not yet been reported. However, glutaredoxins are reduced by glutathione, which has a midpoint potential of −240 mV (5). This suggests that they do not have any direct function in the formation of protein disulfide bonds.
Although these proteins are not able to form protein disulfide bonds on their own, they might participate in alternative pathways. It has been suggested that chloroplast peroxiredoxins might catalyze peroxide-mediated disulfide bond formation in target proteins (50). This reaction is believed to proceed in a similar fashion to the Orp1-Yap1 redox relay in yeast (37). In that reaction, the peroxiredoxin Orp1 oxidizes the transcription factor Yap1 by forming a disulfide bond. The proposed mechanism for this reaction involves the oxidation of a thiol group in the catalytic site of Orp1 to a sulfonic acid (Cys-OH) by hydrogen peroxide. The sulfenic acid then forms an intermediate mixed disulfide bond with a target thiol (Cys598) of YAP1, after which the mixed disulfide bond between Orp1 and Yap1 is reduced by a nucleophilic attack from a second thiol group (Cys303) of Yap1. The final reaction products are oxidized Yap1, which migrates to the nucleus to induce a transcriptional response, and reduced Orp1 (37).
Finally, there is a group of protein disulfide isomerases and related proteins that includes the protein PDIL1-3, and the 5′-adenylylsulfate reductases APR1, APR2, and APR3. The presence of three 5′-adenylylsulfate reductases in this group was unexpected. However, these proteins were identified by a phylogenetic analysis, as members of the protein disulfide isomerase family of Arabidopsis (44) and in vitro import assays indicated that they are located in the chloroplast stroma (14). The enzyme 5′-adenylylsulfate reductase has an important function in the biosynthesis of cysteine (59), but it is not known whether it also catalyzes disulfide bond formation in proteins. The protein disulfide isomerase PDIL1-3 (3) has domains that are very similar to protein disulfide isomerase from yeast and mammalian protein disulfide isomerases (Fig. 5) (36). PDIL1-3 has not yet been reported to have a role in the formation of disulfide bonds of chloroplast proteins, but it is very similar to the protein disulfide isomerase OaPDI from Oldenlandia affinis (35). OaPDI catalyzes the formation of disulfide bonds in cyclotides—cyclic peptides that have three disulfide bonds and are involved in defense against insects. The protein disulfide isomerase and chaperone activity of OaPDI are about 70% and 140% as strong as those of human protein disulfide isomerase (PDI), respectively. The midpoint potential of OaPDI is −159 mV, which is similar to that of mammalian protein disulfide isomerases (77). The amino acid sequences of OaPDI and PDIL1-3 have 58% identity, which suggests that PDIL1-3 from Arabidopsis has functions similar to those of OaPDI. It is thus likely that PDIL1-3 can catalyze disulfide bond formation and shuffle non-native disulfide bonds into their correct connectivity.

Finally, the proteins LQY1 (58) and SCO2 (78) have been suggested to assist in the repair and assembly of photodamaged Photosystem II (PSII) (58) and the integration of light-harvesting complexes into the thylakoid membranes in the chloroplasts of cotyledons (83). LQY1 and SCO2 have protein disulfide isomerase activity (58, 63, 78) but lack the CXXC sequence motif that is found in the active site of Trx and is characteristic of the Trx superfamily. Instead, these proteins resemble the chaperone DnaJ, although they do not have its N-terminal domain for binding to DnaK. It was shown that DnaJ of Escherichia coli has protein disulfide isomerase activity in vitro, and it was proposed that this activity is based on its four CXXCXGXG Zn-binding motifs in the C-terminal region (25). This Zn-binding domain is also present in the C-terminal regions of the proteins LQY1 and SCO2, and may be important in their function as protein disulfide isomerases.
Disulfide Formation in Different Chloroplast Compartments
The above survey of the chloroplast-located members of the Trx superfamily in Arabidopsis identified the protein disulfide isomerase PDIL1-3 in the stroma and the LTO1 in the thylakoid membrane as potentially being involved in catalyzing disulfide bond formation in other proteins. These two proteins can be studied to investigate the roles of disulfide bond formation in the folding, regulation, and signal transduction of chloroplast proteins. In addition, the proteins LQY1 and SCO2 demonstrate that the ability to catalyze disulfide formation in proteins is not limited to members of the Trx superfamily. At present, relatively little is known about the biochemical reactions involved in the formation of disulfide bonds in chloroplast proteins. However, the chloroplasts of green plants contain numerous potential Trx targets that have functions in transcription and translation, and many metabolic pathways (57). This suggests that the network of biochemical reactions involving disulfide bond formation in chloroplast proteins is probably very complex.
Two well-known cases of redox control of chloroplast enzymes by Trxs are the Calvin-Benson cycle enzyme fructose 1,6 bisphosphatase and the pentose phosphate pathway enzyme glucose 6-phosphate dehydrogenase. Fructose 1,6 bisphosphatase is light-dependently activated by the reduction of a disulfide bond by Trx f, whereas glucose 6-phosphate dehydrogenase is light-dependently inactivated by reduction mediated by Trxs m and f (75). It has been suggested that reactive oxygen species (ROS) and hydrogen peroxide might be involved in the inactivation of fructose 1,6 bisphosphatase (75) and the activation of glucose 6-phosphate dehydrogenase (12), respectively. While ROS and hydrogen peroxide may indeed be involved, enzymatic catalysis of disulfide bond formation might also be required to ensure the rapid inactivation of fructose 1,6 bisphosphatase and activation of glucose 6-phosphate dehydrogenase during the transition from light to darkness.
The biosynthesis and repair of the photosynthetic complexes of the thylakoid membrane is another area where disulfide bond formation may play an important role. The cotyledons of Arabidopsis seedlings in mutants lacking the protein SCO2 have small chloroplasts with a deviant shape (78) and partially aberrant ultrastructures (83). SCO2 can bind to LHCB1 and has been proposed to have a role in the assembly of PSII in germinating seedlings (83). Arabidopsis mutants that lack the LQY1 protein are more sensitive to high light stress than wild-type plants and exhibit reduced levels of the PSII-LHCII supercomplex when grown under high light conditions. LQY1 binds to different PSII complexes, and has been suggested to aid in the repair and reassembly of photodamage to PSII (58). These examples demonstrate that enzymes that can catalyze disulfide bond formation in other proteins probably play important roles in the biogenesis and function of the thylakoid membrane.
The ability of the LTO1 to catalyze the formation of a disulfide bond in the extrinsic PSII subunit PsbO (46) highlights the importance of disulfide bond formation in the thylakoid lumen, and provides a link between oxidative folding and the import of lumen-located proteins. The PsbO protein binds to the lumen side of PSII and stabilizes the water-oxidizing complex. As discussed in a recent review, reductions in the abundance of PsbO have severe effects on photosynthesis (69). Knockdown plants that express reduced levels of LTO1 show retarded growth and an impaired function of PSII (46). In addition, the levels of PsbO and other PSII subunits are lower than in wild-type plants (46). How is the ability of LTO1 to form a disulfide bond in PsbO linked to the assembly of PSII and the import of lumenal proteins? Shortly after the import of PsbO into the thylakoids, it adopts a transient form that is sensitive to proteolysis (40). Mature PsbO with a correctly folded disulfide bridge is stable, but it is destabilized when the disulfide bond is reduced (65) and becomes sensitive to proteolysis (38). Taken together, these observations suggest that the disulfide bond of newly imported PsbO proteins is reduced. Fast folding and disulfide bond formation would be needed to ensure that PsbO assumes a stable fold and can bind to PSII. The disulfide bond might form spontaneously due to the oxidizing environment in the lumen during photosynthesis, but spontaneous disulfide formation is relatively slow. Therefore, enzymatic assistance is probably required to minimize losses of newly imported unstable PsbO via proteolysis. LTO1 would thus be very important for the correct folding of PsbO and the assembly of PSII.
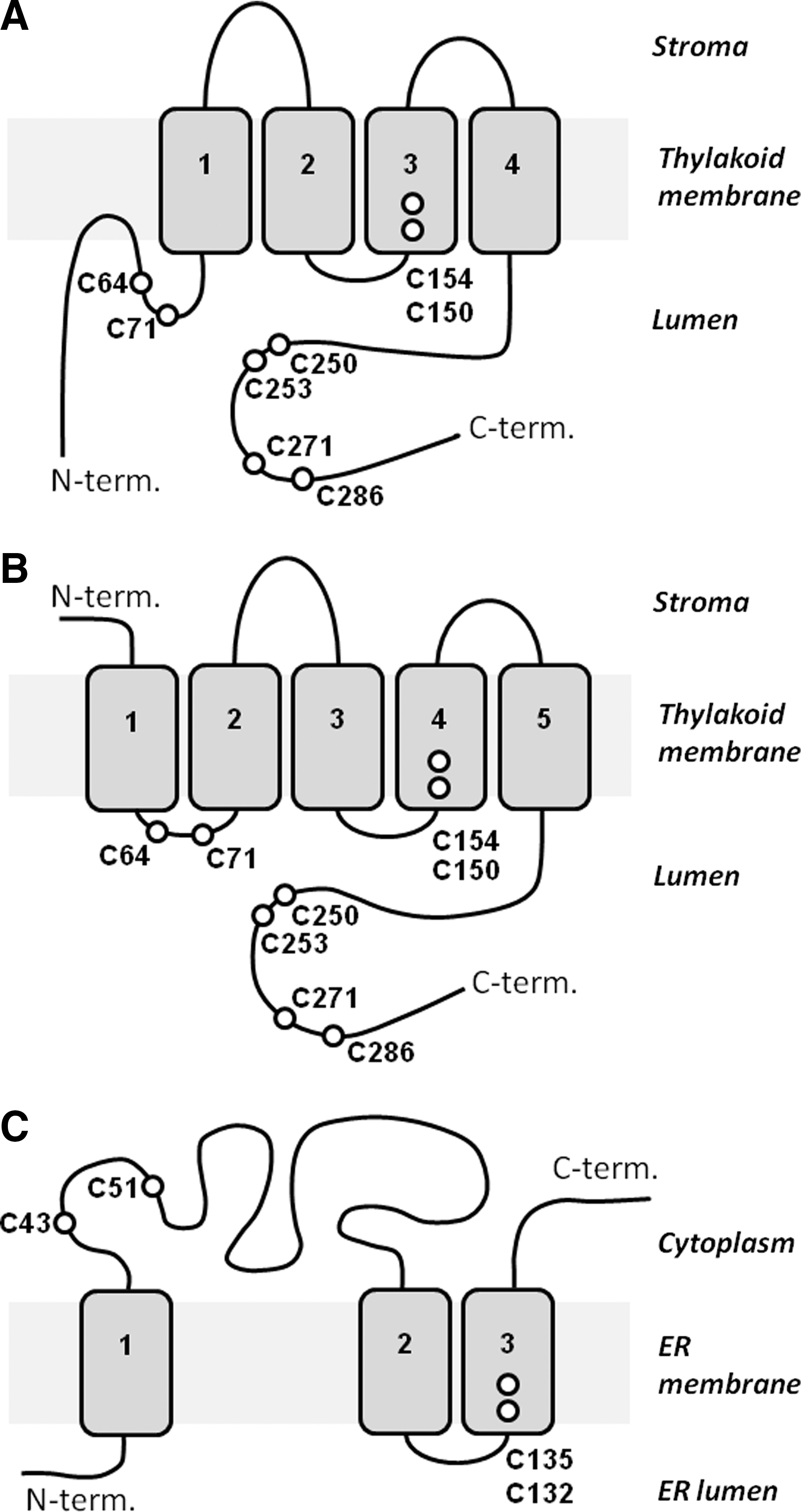
The sequence of LTO1 from Arabidopsis contains an N-terminal vitamin K1 epoxide reductase (VKOR) domain that is followed by a C-terminal Trx-like domain (Fig. 6). The VKOR domain is related to mammalian vitamin K1 epoxide reductase complex subunit 1 (VKORC1) of the vitamin K cycle (79). VKORC1 regenerates active vitamin K1 by reducing the inactive epoxide of vitamin K1 that is produced during the maturation of blood-clotting proteins (79). To understand the function of LTO1, it is important to know its membrane topology and the subcellular location of its C-terminal Trx-like domain. Two studies (29, 46) showed that the C-terminal Trx-like domain is located in the thylakoid lumen (Fig. 7A, B), which is consistent with a role in the formation of disulfide bonds in lumenal proteins. It is also important to determine the mechanism by which the reduced Trx-like domain becomes oxidized. It has been proposed that the electrons used to reduce the vitamin K1 epoxide are provided by a Trx that reduces the cysteine residues C43 and C51 in the cytoplasmic loop of VKORC1, as occurs in human VKORC1 (Fig. 7C). These cysteine residues in turn reduce C132 and C135 in the third transmembrane helix, where vitamin K1 epoxide is reduced (72). In agreement with this model, the recombinant VKOR domain of LTO1 can oxidize the reduced Trx-like domain in vitro, which suggests that it also can serve as an acceptor for electrons from the reduced Trx-like domain in vivo (32). However, unlike VKCORC1, LTO1 cannot reduce vitamin K1 epoxide. The VKOR domain of LTO1 needs vitamin K1 (phylloquinone) as an electron acceptor (32). The mechanism for the re-oxidation of reduced vitamin K1 in the thylakoid membrane has yet to be determined, and doing so will be an important objective for future studies in this area.

Conclusions
Four chloroplast proteins from A. thaliana that are likely to have important functions in the formation of disulfide bonds in target proteins have been identified. The thylakoid proteins LQY1 and SCO2 exhibit protein disulfide isomerase activity and function in the assembly and repair of PSII, and the biogenesis of thylakoids in cotyledons, respectively. The disulfide oxidoreductase LTO1 is important for the disulfide bond formation and folding of the extrinsic PsbO subunit of PSII. It is probably also involved in disulfide bond formation in other lumenal disulfide proteins and might control the transfer of thiol-mediated signals. Finally, the protein disulfide isomerase PDIL1-3 is a likely candidate for the catalysis of oxidative folding in the chloroplast stroma. At this point it is premature to discuss the evolution of the protein folding system of chloroplasts from green plants. However, many cyanobacteria have proteins that are related to LTO1 from Arabidopsis, which suggests a bacterial origin for this protein. Conversely, PDIL1-3 appears to have a eukaryotic origin. Many details of the mechanisms of oxidative folding in green plant chloroplasts are currently unknown, but foundations for future studies in this area have been established. Given the close link between import and folding of chloroplast proteins, it is reasonable to suggest that there may be a set of as-yet unidentified components of the import apparatus of the envelope that function as co-chaperones for Hsp93 and that catalyze disulfide bond formation in newly imported chloroplast proteins.
Footnotes
Acknowledgment
T.K. thanks Wolfgang P. Schröder for critical reading of the manuscript.