
Abstract
Introduction
Methionine sulfoxide reductases (Msrs) catalyze the reduction of free and protein-based methionine sulfoxide to methionine. They consist of two families according to substrate stereospecificity. MsrA is specific for the S-form of methionine sulfoxide, whereas MsrB reduces the R-form (9, 28, 30). A single MsrA and three MsrBs (MsrB1, B2, and B3) are present in mammalian cells (16, 34). The principal function of Msrs is to repair oxidatively damaged proteins by reversing the oxidation of methionine residues. The cyclic oxidation/reduction of methionine is believed to be an important process in scavenging reactive oxygen species (ROS) (21, 44). A number of studies have demonstrated that MsrA and MsrB function as antioxidant enzymes, protecting cells against oxidative stress (22, 24, 25, 29).
Methionine, an essential amino acid in mammals, supports the methylation cycle involving S-adenosylmethionine and is relevant to the biosynthesis of cysteine and other compounds (19, 45). Homocysteine is generated by methionine metabolism and is converted to cysteine by the consecutive actions of cystathionine-β-synthase (CBS) and cystathionine-γ-lyase (CSE) in the trans-sulfuration pathway. Methionine restriction leads to the downregulation of the trans-sulfuration pathway by reducing levels of CBS (39). Cysteine is a substrate of the two pyridoxal-5′-phosphate-dependent enzymes, CBS and CSE, generating hydrogen sulfide (H2S) (42). H2S, which has been known for decades as a toxic gas, is currently recognized as a gaseous physiological molecule (1, 11, 42, 48). Recent studies have demonstrated that H2S performs a cytoprotective role in a variety of organs, including the heart, brain, intestine, and kidney, against ischemia/reperfusion (I/R) injury (5, 41, 49). Thus, it is possible that Msrs may be involved in the regulation of the trans-sulfuration pathway and the production of H2S.
Innovation
The present study shows that MsrA deletion exacerbates the susceptibility of the kidneys to I/R injury, thereby indicating that MsrA is an important protein in protecting the kidney against I/R injury. Additionally, MsrA knockout reduces the levels of plasma homocysteine and H2S, both of which are relevant to I/R injury. Moreover, MsrA regulates the activity and expression of both CBS and CSE enzymes in the trans-sulfuration pathway during I/R injury. A better understanding of the MsrA-homocysteine-H2S axis will clearly be of benefit in regard to efforts to preserve kidney function under I/R injury conditions.
I/R in the kidney induces acute kidney injury, which is characterized by high mortality and morbidity. No effective therapeutic strategy for the treatment of I/R injury has yet been developed (3). I/R injury is associated with a number of factors, including ROS, cytokines, and chemokines, all of which are generated abundantly during I/R (3, 43). ROS are recognized as a major cause of kidney I/R injury, as they induce lipid peroxidation, protein dysfunction, cellular cytoskeletal disruption, cellular integrity degradation, DNA breakdown, and inflammatory responses (3, 7, 16, 33).
MsrA is known to be abundantly expressed in the kidney (26). Considering that ROS play a causal role in kidney I/R injury, we hypothesized that MsrA might protect the kidney against I/R injury. We also hypothesized that MsrA might affect homocysteine and H2S production in methionine metabolism and the trans-sulfuration pathway. To test these hypotheses, we employed MsrA gene-deleted mice in a kidney I/R injury model (23). We found that MsrA knockout (MsrA−/− ) mice were much more susceptible to kidney I/R injury than wild-type mice (MsrA+/+ ), and that MsrA could indeed regulate the production of homocyteine and H2S under I/R conditions.
Results
MsrA gene deletion accelerates renal functional and morphological impairments after I/R injury
To investigate the functions of MsrA in the susceptibility of the kidney to I/R insult, MsrA+/+ and MsrA−/− mice were subjected to bilateral renal ischemia for 30 min. 30 min of bilateral renal ischemia markedly increased the plasma creatinine (PCr) levels in both MsrA+/+ and MsrA−/− mice at 4 and 24 h after ischemia, indicating that 30 min of renal ischemia resulted in kidney dysfunction (Fig. 1A). The increase of PCr levels in the MsrA−/− mice was more profound than in the MsrA+/+ mice (Fig. 1A). Consistent with renal functional impairment, I/R resulted in severe loss of the brush borders of kidney tubular epithelial cells, luminal congestion, and atrophy and dilation of the tubules in the outer medulla of the kidney (Fig.1B). These post-ischemic histological changes were more severe in the MsrA−/− mice than in the MsrA+/+ mice (Fig. 1B). In accordance with PCr concentrations, histological damage scores were higher in the MsrA−/− mice than in the MsrA+/+ mice (Fig. 1C). These results indicate that the deletion of MsrA gene accelerates renal injury after I/R insult.

MsrA gene deletion enhances post-ischemic kidney congestion and inflammatory responses
The severe congestion in the outer medulla was noted in the hemi-sect kidney at 24 h after ischemia (Fig. 2A). The congestion in the MsrA−/− mouse kidney was significantly greater than in the MsrA+/+ mouse kidney (Fig. 2A). The infiltration of leukocytes has been implicated in the cell damage occurring pursuant to I/R insult. Expression levels of lymphocyte antigen 6 complex (Ly6G), an index leukocyte, were significantly higher in MsrA−/− than in MsrA+/+ (Fig. 2B). Microscopic examination revealed much greater accumulation of leukocytes in the interstitium in the MsrA−/− mice than in the MsrA+/+ mice (Fig. 1B). The data suggest that MsrA gene deletion induces enhanced post-ischemic congestion and inflammation in the kidney.
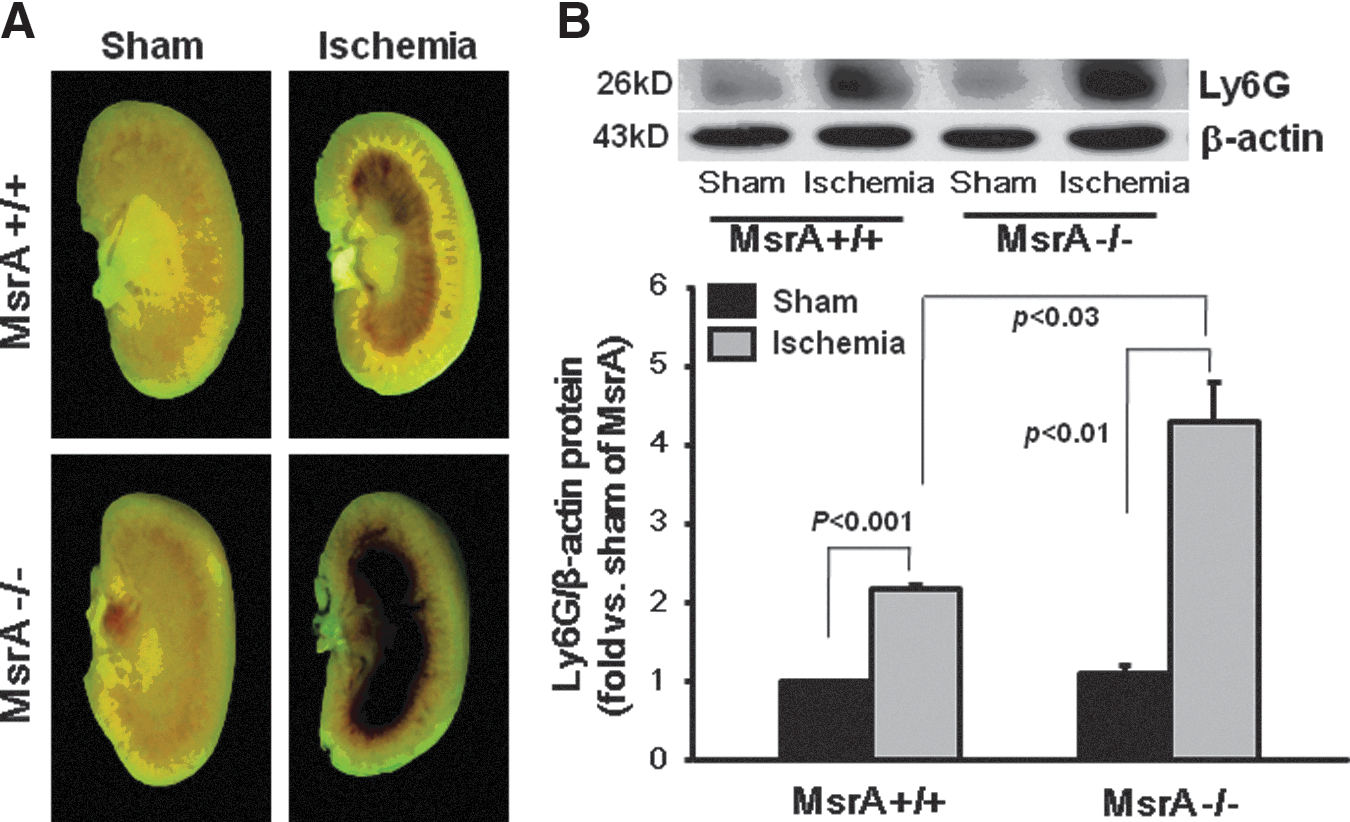
MsrA gene deletion augments kidney oxidative stress after I/R injury
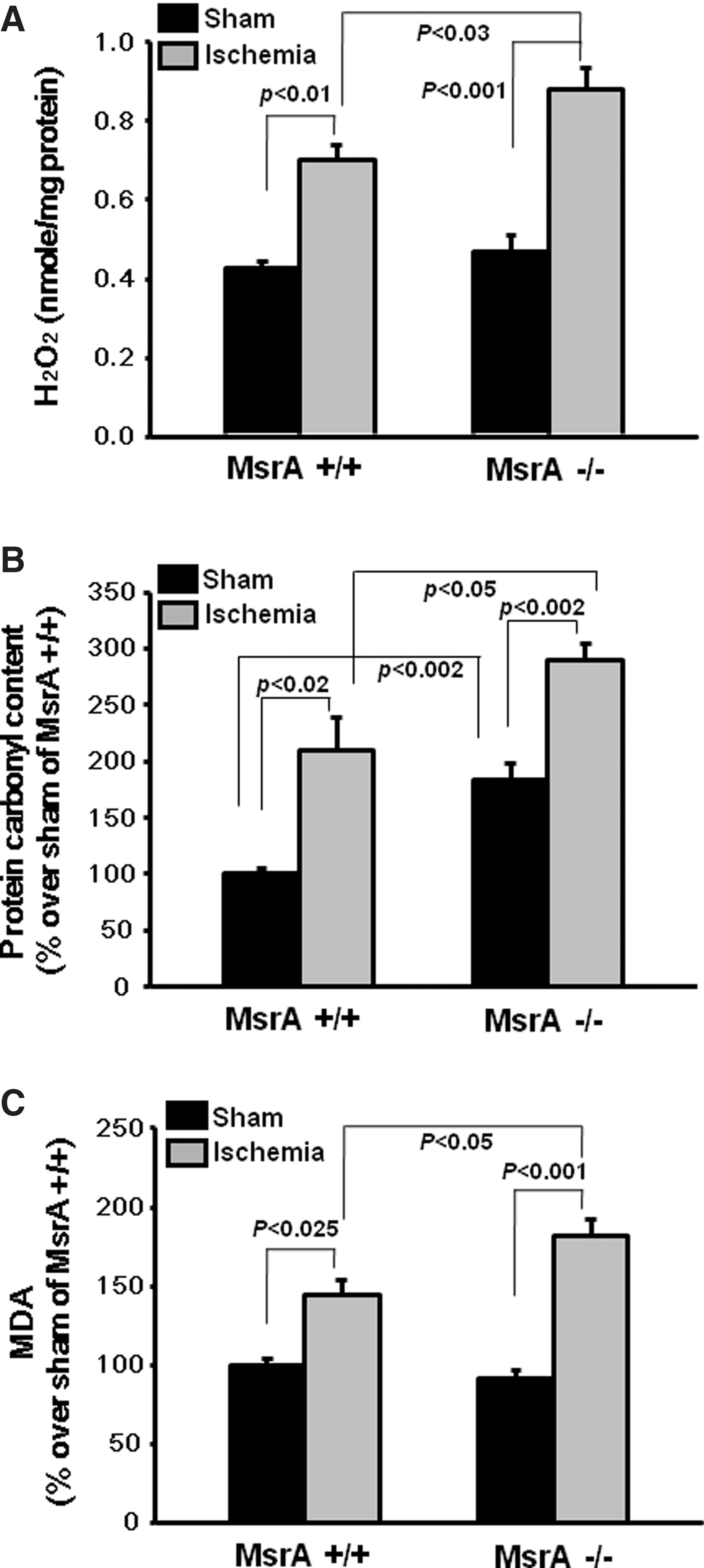
Since MsrA functions as an antioxidant enzyme, we investigated its protective role against I/R-induced oxidative stress. 30 min of bilateral renal ischemia, followed by 24 h reperfusion, significantly increased kidney tissue H2O2 levels (Fig. 3A). The H2O2 levels were significantly higher in the I/R-subjected MsrA−/− mouse kidney than in the I/R-subjected MsrA+/+ mouse kidney (Fig. 3A). The control levels of H2O2, when sham operation was performed, were not significantly different between the MsrA+/+ and MsrA−/− kidneys (Fig. 3A). The levels of protein-carbonyl content were significantly higher in the sham-operated MsrA−/− kidney than in the MsrA+/+ kidney (Fig. 3B), thereby indicating that MsrA deletion resulted in increased basal levels of protein oxidation. I/R increased the protein-carbonyl content in the mouse kidneys (Fig. 3B). A significantly higher increase in the protein-carbonyl content was observed in the kidneys of MsrA−/− mice after I/R injury, compared with the MsrA+/+ mice (Fig. 3B). The levels of lipid peroxidation were also significantly greater in the MsrA−/− mouse kidney after I/R than in the MsrA+/+ mouse kidney (Fig. 3C), while no significant differences between the sham-operated MsrA+/+ and MsrA−/− kidneys were detected. These results indicate that the increased susceptibility of MsrA−/− mice to I/R injury is associated with enhanced oxidative stress.
I/R reduces the expression and activity of Msrs in the kidney
Next, we attempted to determine whether I/R affected the activity and expression of Msrs. We first determined the activity and expression levels of MsrA and MsrB1 in the kidneys. It should be noted that among the three MsrBs, selenoenzyme MsrB1 is the main MsrB in the kidney (10). The MsrA protein level in the MsrA+/+ kidney declined significantly at 24 h after ischemia (Fig. 4A). Consistent with the reduced MsrA protein levels, its mRNA levels were reduced by 30% (Fig. 4B). In accordance with the reduced MsrA expression in association with I/R injury, the MsrA activity was reduced by 30% after I/R (Fig. 4C). In the case of MsrB1, I/R did not alter both the protein and mRNA levels of MsrB1 in the MsrA+/+ kidney (Fig. 4A and B). However, MsrB activity was slightly but significantly reduced in the I/R-subjected MsrA+/+ mouse kidney (Fig. 4D). The expression levels of MsrB2 and MsrB3 were not altered after I/R in the MsrA+/+ kidney (data not shown). MsrB activity in the I/R-subjected MsrA−/− mouse kidney decreased markedly when compared with that measured in the I/R-subjected MsrA+/+ kidney (Fig. 4D). Consistent with the data of MsrB enzyme activity, Western blot and RT-PCR analyses showed 60% and 20% reductions in MsrB1 protein and mRNA levels, respectively, in the MsrA−/− kidney after I/R (Fig. 4A and B). Taken together, the data show that I/R reduces MsrA expression and activity in the MsrA+/+ kidney, and that it reduces MsrB1 expression and activity more profoundly in the MsrA−/− kidney than in the MsrA+/+ kidney.

MsrA gene deletion accelerates the decrease of hydrogen sulfide after I/R injury
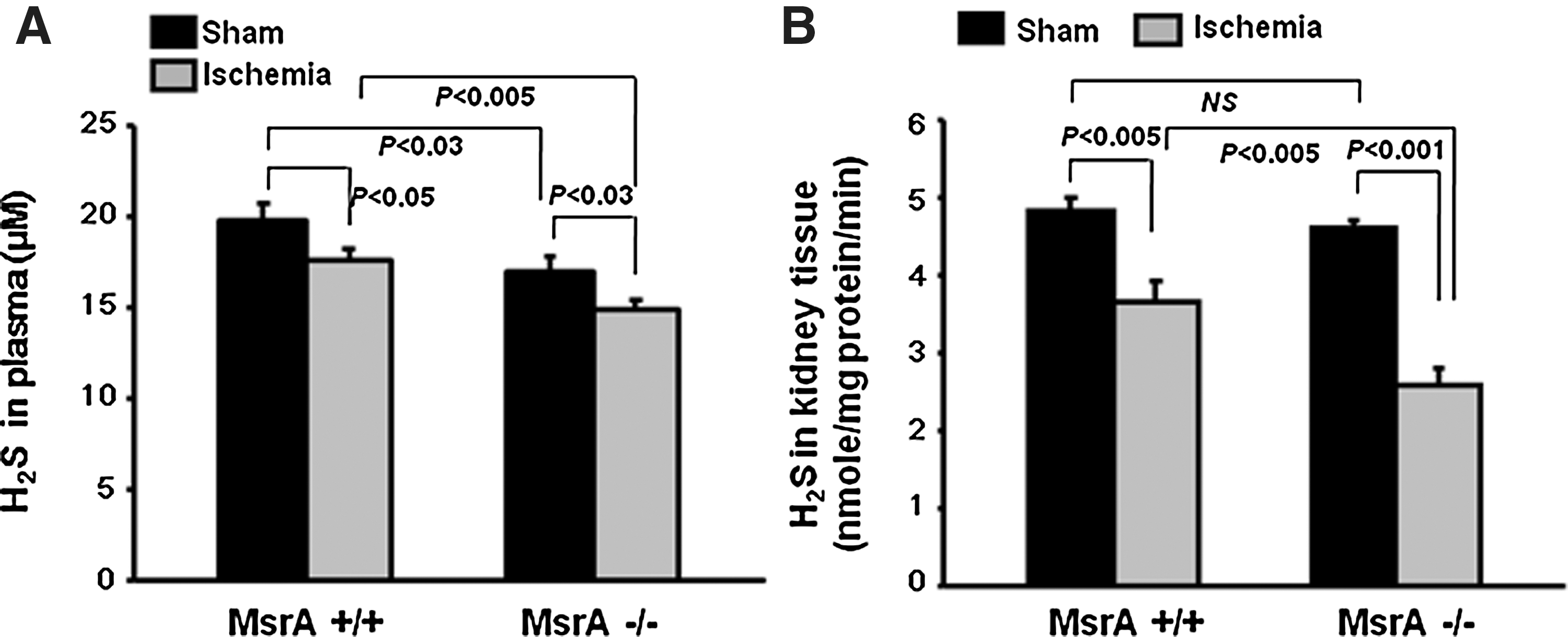
H2S is generated during the metabolism of homocysteine, a metabolite of methionine. Since MsrA may affect methionine metabolism, we investigated H2S production under sham and I/R conditions. Plasma H2S levels were lower in the MsrA−/− mice than in the MsrA+/+ mice at 24 h after either sham operation or I/R (Fig. 5A). Although no significant differences in the H2S levels of kidney tissue were noted between the sham-operated MsrA−/− and MsrA+/+ mice (Fig. 5B), I/R induced reductions in H2S to a significantly greater degree in the kidneys of MsrA−/− mice as compared to MsrA+/+ mice (Fig. 5B).
No significant difference in CBS expression was observed between the MsrA−/− and MsrA+/+ mice with sham operation; however, interestingly, the post-I/R decrease in expression was significantly more profound in the MsrA−/− mice than in the MsrA+/+ mice (Fig. 6A). Consistent with its similar expression levels after sham operation, the CBS activity of the MsrA+/+ and MsrA−/− kidneys did not differ (Fig. 6B). However, 24 h after I/R, CBS activity declined significantly in the kidneys of both MsrA+/+ and MsrA−/− mice. As expected, the decline of CBS activity in the MsrA−/− mouse was much greater than that in the MsrA+/+ mouse (Fig. 6B).
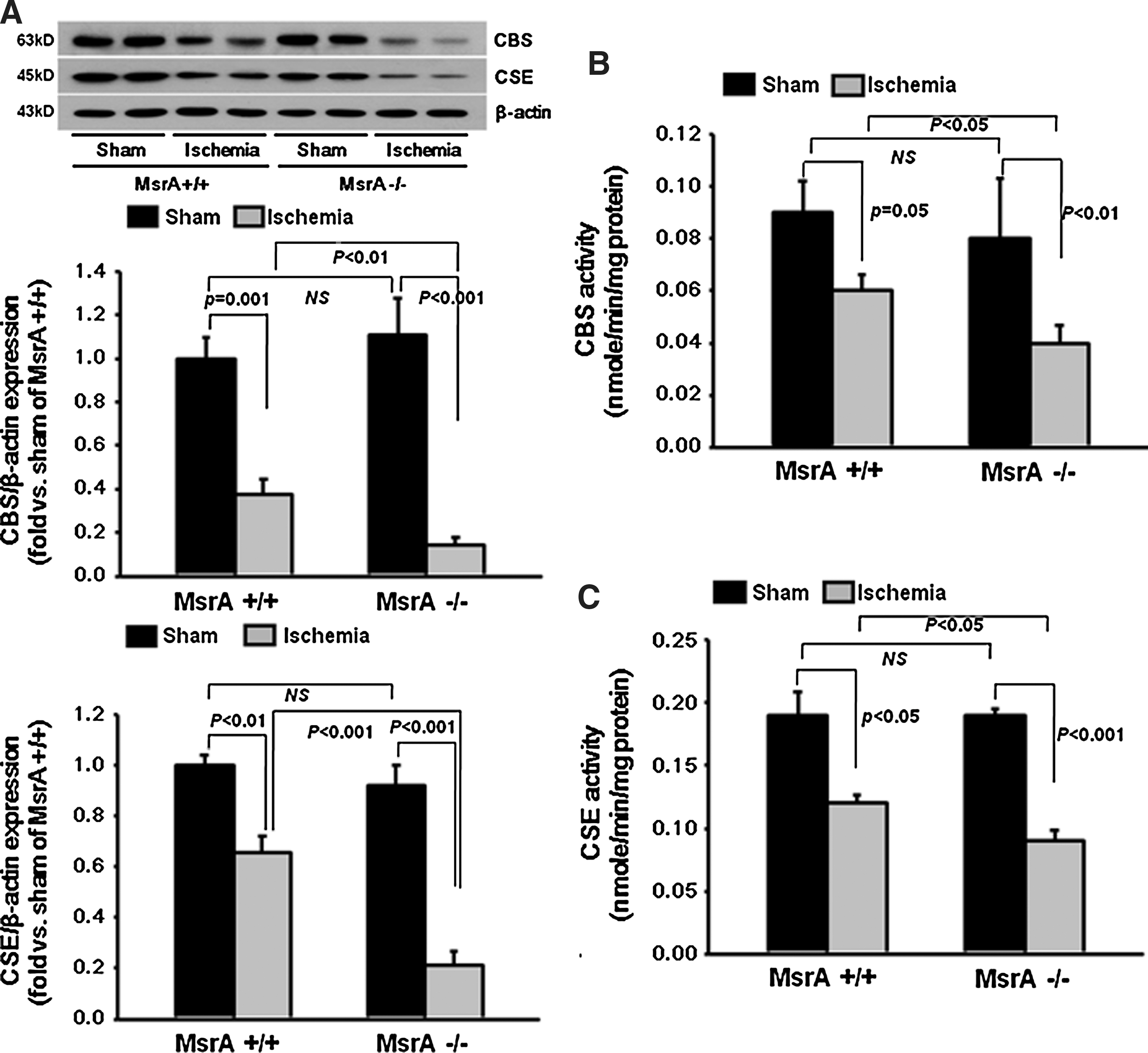
The levels of CSE expression did not differ between the kidneys of the MsrA−/− and MsrA+/+ mice after sham operation (Fig. 6A). In contrast, 24 h after ischemia, the CSE expression levels were greatly reduced in both the MsrA−/− and MsrA+/+ kidneys. Furthermore, the CSE expression after I/R was significantly lower in the MsrA−/− mice than in the MsrA+/+ mice (Fig. 6A). Consistent with the reduced CSE expression, activity was also reduced in the kidneys of both the MsrA−/− and MsrA+/+ mice after ischemia (Fig. 6C). As expected, the reduction in CSE activity was greater in the MsrA−/− mice than in the MsrA+/+ mice (Fig. 6C). Taken together, the data indicate that MsrA regulates H2S production during I/R by modulating the expression and activity of the CBS and CSE enzymes.
MsrA gene deletion reduces homocysteine levels
Finally, to determine whether MsrA gene deletion affects methionine metabolism, we measured the levels of homocysteine, a product of methionine metabolism and a substrate of CBS for H2S generation. The basal level of homocysteine in the plasma of MsrA−/− mice was significantly lower than in that of MsrA+/+ mice (Fig. 7A). After ischemia, plasma homocysteine level declined significantly in both types of mice. Much lower levels of plasma homocysteine were observed in the I/R-subjected MsrA−/− mice than in the I/R-subjected MsrA+/+ mice (Fig. 7A). In the kidneys, a reduction in basal homocysteine level was observed in the MsrA−/− mice relative to the MsrA+/+ mice, but this reduction was not significant (Fig. 7B). I/R reduced kidney homocysteine levels in both the MsrA+/+ and MsrA−/− mice, but a significant reduction was observed only in the MsrA−/− mice. Collectively, these data indicate that MsrA modulates homocysteine production under normal conditions, and this effect can be augmented by kidney I/R.

Discussion
Herein, we report the link between MsrA and kidney I/R injury for the first time. Additionally, this study provides the first evidence that MsrA regulates the production of H2S, which is highly implicated in I/R injury, under in vivo conditions. The deletion of the MsrA gene exacerbated kidney I/R injury and inhibited H2S production under I/R conditions by downregulating the expression and activity of CBS and CSE and the substrate homocysteine.
Considering that ROS play a causal role in I/R injury, it is expected that the antioxidant enzyme MsrA may protect against such injury. A recent study by Zhao et al. reported that MsrA overexpression confers resistance to I/R injury in the heart (51). Prentice et al. reported that MsrA overexpression protects cardiac myocytes against hypoxia/reoxygenation (38). The data reported herein clearly demonstrate the protective role of MsrA against kidney I/R injury.
Under normal conditions, H2O2 and lipid peroxidation levels in the MsrA−/− kidney did not differ significantly compared to those observed in MsrA+/+ mice. Additionally, no significant functional and histological impairments were noted in sham-operated MsrA−/− mice. In contrast, under pathological I/R conditions, the MsrA−/− mouse kidney, with more severe functional and histological impairments, evidenced greater levels of H2O2 and lipid peroxidation than were seen in the MsrA+/+ mice kidney. Protein-carbonyl content is generally employed as a marker for protein oxidation and oxidative stress. The accumulation of protein-carbonyl is positively correlated with the methionine oxidation content (27). In this study, the basal content of carbonylated proteins of the normal kidneys was significantly greater in the MsrA−/− mice than in the MsrA+/+ mice, which indicates that MsrA carries out an antioxidant defense function even under normal conditions. Moreover, after I/R, like H2O2 and lipid peroxidation, the content of protein-carbonyl in the MsrA−/− mice kidneys was also significantly higher than in the MsrA+/+ mice. Taken together, the data indicate that MsrA is highly important for defense against pathologically induced oxidative stresses such as I/R injury. Notably, the I/R-induced increase in protein carbonylation was similar between the MsrA+/+ and MsrA−/− mice. Since proteins in the MsrA−/− mice have already been oxidatively damaged to a significant degree in the resting state, we speculate that additional oxidative stress induced by I/R may be somewhat less compelling carbonylation of protein in the MsrA−/− mice.
In wild-type mice, I/R attenuated the expression and activity of MsrA, but did not reduce the expression of MsrB1, which is a main enzyme of the MsrB family. Although the expression level of MsrB1 was not altered by I/R in the kidneys of MsrA+/+ mice, a significant reduction in MsrB activity was noted. The reduced MsrB activity may be due to the inactivation of the MsrB1 protein resulting from the increased oxidation status induced by I/R injury. In contrast, in the MsrA−/− mice, I/R markedly reduced MsrB1 expression and MsrB activity. It remains unclear as to why the effect of I/R on the expression of MsrB1 is so dependent on the MsrA protein. Since MsrB1 functions as an antioxidant, the substantial I/R-induced reductions in its expression and activity in the MsrA−/− mice can exacerbate the oxidative damage.
H2S has been recognized as a potent antioxidant, anti-inflammatory, and physiological gaseous molecule (1, 4, 42, 48). Recent evidence from human and animal studies has demonstrated that H2S is directly involved in various diseases, including hypertension, atherosclerosis, and end-stage renal disease (42). The cytoprotective effects of H2S have been reported in a variety of organs, including the heart, brain, liver, and kidney, against I/R injury (5, 37, 47). In this study, we determined that the deletion of MsrA induced a reduction in the levels of plasma H2S and accelerated the post-I/R decrease of H2S in plasma and kidney tissue. Consistent with the reduced H2S levels, homocysteine levels were lower in MsrA−/− than in MsrA+/+ mice. Recently, Bearden et al. reported that extracellular trans-sulfuration from homocysteine to H2S protects the endothelium against redox stress (2). Our data suggest that MsrA deficiency alters methionine metabolism and results in reduced homocysteine levels, leading to a sequential reduction in H2S production. Therefore, the reduced production of H2S in MsrA−/− mice may increase the sensitivity of the kidney to I/R.
Homocysteine is an intermediate amino acid formed during the metabolism of methionine to cysteine. It can be converted to methionine, but is largely metabolized to cysteine (about 78% of homocysteine) in the proximal tubule of the kidney by the sequential reactions of CBS and CSE in the trans-sulfuration pathway (10). CBS and CSE generate H2S via a multitude of reactions and substrate combinations. Recent studies have demonstrated that deficiencies in the CBS and CSE enzymes increase the susceptibility of organs to I/R injury, whereas the overexpression of those enzymes reduces it (4, 18). Wu et al. reported that renal I/R inhibits CBS gene expression mediated by a transcription factor Sp1, leading to the accumulation of homocysteine and the reduction of H2S in the rat kidney (46). Previous studies reported that I/R reduces CBS and CSE activities and H2S production, leading to increased kidney damage and oxidative stress (37, 47). In the heart, Calvert et al. reported that CSE gene-transfer and exogenous H2S treatment protected the heart against I/R injury by attenuating oxidative stress (5).
In this study, I/R reduced homocysteine levels in the plasma and, furthermore, MsrA gene deletion exacerbated these reductions. The homocysteine level in the kidney did not change in the MsrA+/+ mouse after I/R injury, but was significantly reduced in the MsrA−/− mice. Additionally, both CBS and CSE activities were significantly reduced after I/R, and the degree of reduction was greater in the MsrA−/− mice than in the MsrA+/+ mice. Consistent with the reduced CBS and CSE activities, the expression levels of those enzymes were significantly reduced after I/R. Also, the degree of reduction was greater in the I/R-subjected MsrA−/− mice than in the I/R-subjected MsrA+/+ mice. From our results, we speculate that MsrA gene deletion inhibits methionine metabolism to homocysteine by restricting methionine availability, as judged from the reduced homocysteine levels, and this reduction in homocysteine, a substrate of CBS, results in an additional reduction in H2S production. Furthermore, more profound reductions in the activity and expression of both CBS and CSE in the I/R-subjected MsrA−/− mouse kidney may induce a further reduction of H2S, leading to greater oxidative stress, inflammatory responses, and kidney injury.
Kidney oxidative stress induces tissue damage and also accelerates inflammatory responses, including neutrophil accumulation into the kidney tissue, which triggers ROS production, thereby leading to further tissue and cell damage (15, 17, 36). Inflammatory responses perform an important role in kidney I/R injury. During I/R injury, leukocytes adhere to endothelial cells and infiltrate into the tissues. Accumulating evidence has demonstrated that the inhibition of leukocytic adhesion to endothelial cells protects the kidneys against I/R injury (12, 35, 36, 40). Many studies have demonstrated that H2S inhibits the production of inflammatory cytokines as well as leukocyte-endothelial interactions (20, 31, 49, 50). Yang et al. reported that the reduction of H2S induced by CSE deletion diminishes endothelium-dependent vasodilation (48). In this study, we determined that I/R increases kidney congestion and leukocyte infiltration, and those changes were enhanced by MsrA deletion. Therefore, it can be speculated that the higher kidney congestion and leukocyte infiltration noted in MsrA−/− I/R mice may be associated with the attenuation of vasodilation in the renal vessels and with increased inflammatory responses, both of which result from the reduction of H2S.
Materials and Methods
Animal preparation
All experiments were conducted using 8- to 10-week-old MsrA gene-deleted (MsrA−/− ) or wild-type (MsrA+/+ ) male mice weighing 20 to 25 g. MsrA−/− and MsrA+/+ mice used in this study were generated by breeding heterozygous MsrA knockout mice (23) that were of mixed background of 129/SvJ and C57BL/6. This study was conducted wholly in accordance with the guidelines of the Institutional Animal Care and Use Committee of Kyungpook National University and of Yeungnam University. Kidney ischemia was induced as described previously (35).
Renal functional parameter
PCr concentrations were determined using a Beckman Analyzer II, as described previously (34).
Histology
Paraffin sections of kidneys were stained with PAS to evaluate histological changes. Kidney tubular damage level was scored described previously (35).
MsrA and MsrB activity
The reaction mixture (100 μl) contained 50 mM sodium phosphate (pH 7.5), 50 mM NaCl, 20 mM dithiothreitol, 200 μM dabsylated methionine-S-sulfoxide (for MsrA) or methionine-R-sulfoxide (for MsrB), and 200 μg crude protein. The reaction was conducted at 37°C for 30 min, and the reaction product, dabsyl-Met, was analyzed via HPLC.
H2S concentration in the plasma and kidney
Fifty μl of plasma was mixed with 187.5 μl of 1% zinc acetate and 12.5 μl of 12% NaOH solution and then incubated for 10 min at 37°C. 1.75 ml of distilled water, 0.2 ml of 20 mM N,N-dimethyl-p-phenylenediamine dihydrochloride in 7.2 M HCl, and 0.2 ml of 30 mM FeCl3 in 1.2 M HCl were added and incubated for 10 min in a dark box at room temperature. Optical density at 670 nm was measured using a spectrophotometer (Ultraspec2100, Tecan). The H2S concentration was calculated against the standard curve of NaHS solution.
For the kidney, kidney tissues were homogenized in ice-cold 100 mM potassium phosphate buffer (pH 7.4) with protease inhibitors. 100 μl of homogenate was mixed with 100 μl of 1% zinc acetate, 100 μl of borate buffer (pH 10.0), 200 μl of 20 mM N,N-dimethyl-p-phenylenediamine dihydrochloride in 7.2 M HCl, and 200 μl of 30 mM FeCl3 in 1.2 M HCl and incubated for 15 min at 37°C. The sample was centrifuged for 5 min at 10,000 rpm at 4°C. The supernatant was taken and optical density was measured at 670 nm.
CBS and CSE enzyme activity
The kidney homogenate was prepared as described above. CBS and CSE activities were determined according to the methods described previously (6, 47). 120 μl of homogenate was mixed with 20 μl of 10 mM L-cysteine, 20 μl of 10 mM DL-homocysteine, 20 μl of 0.05 mM S-adenosylmethionine, and 20 μl of 2 mM pyridoxal 5′-phosphate and incubated for 90 min at 37°C. Then, 100 μl of 1% zinc acetate, 100 μl of 10% trichloroacetic acid, 350 μl of 40 mM N,N-dimethyl-p-phenylenediamine dihydrochloride and 350 μl of 60 mM FeCl3 were added and incubated for an additional 15 min at 37°C. The reaction mixture was centrifuged for 5 min at 10,000 rpm at 4°C and the optical density at 670 nm was measured. For the CSE activity assay, 20 μl of 10 mM L-cysteine and 20 μl of 2 mM pyridoxal 5′-phosphate were used for H2S production.
Homocysteine measurements
Homocysteine levels in the plasma were measured using a homocysteine ELISA kit (MyBiosource) according to the manufacturer's protocols. For the kidney, kidney was smashed in 1 ml PBS, spun down at 5000 rpm for 1 min to wash, and then the pellets were resuspended in 1 ml of distilled water. The resuspended kidney was sonicated with an ultrasonic processor (Sonic and Materials Inc., Newton, CT). After centrifugation, 100 μl of the supernatant was used for homocysteine measurement using the homocysteine ELISA kit.
Lipid peroxidation, hydrogen peroxide, and protein-carbonyl content
Thiobarbituric acid-reactive substances (TBARS; Sigma-Aldrich) were used to measure lipid peroxidation (17). H2O2 levels were measured using the ferric sensitive dye, xylenol orange, as previously described (17). Protein-carbonyl content was measured using an OxyBlot protein oxidation detection kit (Chemicon International) according to the manufacturer's recommendations.
Western blot analysis
Western bolt analysis was conducted as described previously (35). Polyclonal rabbit anti-mouse MsrA antibodies were generated. Polyclonal anti-mouse MsrB1 were described elsewhere (8). Other antibodies used were: Ly6G (Sigma-Aldrich), β-actin (Sigma-Aldrich), CBS (Santa Cruz), and CBE (Santa Cruz).
Analysis of MsrA and MsrB1 mRNA levels by reverse transcription-polymerase chain reaction
Total RNA from chopped kidneys was extracted with TRI-Solution (Bioscience). The isolated RNA was reverse-transcribed, followed by a PCR reaction to amplify MsrA or MsrB1 genes. β-actin was used to normalize the amount of RNA in each sample. The primers used were: 5′-GAACCTTTCCCAGAGGGAACAC-3′ and 5′-CTCAGGTACTGCTGGTGGTAGTC-3′ for MsrA, 5′-GCCAGGTGTCTACGTGTGTG-3′ and 5′-TTCTTTGCCTTTAGGGACGA-3′ for MsrB1, and 5′-GAGACCTTCAACACCCCAGC-3′ and 5′-CACATCTGCTGGAAGGTGGAC-3′ for β-actin.
Statistical analysis
The results were expressed as means±S.E. Statistical differences among groups were calculated using analysis of variance (ANOVA) followed by the least significant difference (LSD) comparisons using SPSS 12.0. A p value of<0.05 was considered significant.
Footnotes
Acknowledgments
We thank Drs. Rodney Levine (NIH, USA) and Vadim Gladyshev (Harvard Medical School, USA) for kindly providing the MsrA knockout mice. This research was supported by Basic Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science, and Technology (2012002592 to KMP and 2011-0006178 to HYK).
Author Disclosure Statement
No competing financial interests exist