
Abstract
Introduction
The macrophage receptor CD163 is a scavenger receptor that is expressed in cells of the monocytic lineage (18, 23, 25, 34). The receptor belongs to the ancient scavenger receptor cysteine-rich (SRCR) class B superfamily, and CD163 orthologs have been reported in most mammalian species, including monkey (48), pig (38), cattle (14), dog (7), and rodents (33, 41). CD163 is a 130 kDa type I transmembrane protein that is composed of nine consecutive extracellular SRCR domains (18). CD163 expression is detected in all circulating monocytes (23) but is strongly up-regulated in tissue-resident and anti-inflammatory macrophages (5, 17, 25, 34, 47).
Innovation
Many observations now link hemoglobin (Hb) metabolism to diseases, such as diabetes and cardiovascular disease (19), and CD163 is currently being evaluated as both a biomarker of inflammatory conditions (10, 24) and a potential target for new therapy (9, 10). Understanding the mouse system for CD163-mediated Hb scavenging and the differences compared with the human system are essential for translating data on Hb pathophysiology from mouse to man and for development of new drugs and diagnostics in this field.
In humans, CD163 functions as a scavenging receptor for the haptoglobin (Hp)-hemoglobin (Hb) complex formed on intravascular hemolysis (17, 26). During physiological conditions, intravascular hemolysis accounts for ∼10% of daily red blood cell degradation (11, 12, 14). Exposure of Hb to the cell-free environment after hemolysis or tissue injury may result in a series of toxic events that are driven by the highly reactive heme molecule. Autoxidation of Hb generates nonoxygen-binding methemoglobin (HbFe3+) and highly reactive superoxide ions, which can be transformed into H2O2 and water. In the presence of H2O2, the cell-free Hb (HbFe2+O2) is oxidized to the oxo-ferryl form (HbFe4+=O), whereas methemoglobin formed by autoxidation generates a Hb protein radical (Hb.Fe4+=O) (6, 39). This leads to oxidative damage of the Hb molecule and, in turn, tissue injury, notably in the kidney where Hb is filtered (20). In addition, Hb-mediated nitrosylation of the vasodilator nitric oxide (NO) (13, 36) results in an excessive NO scavenging, leading to several pathological conditions, including smooth muscle dystonia and gastrointestinal contractions. To neutralize Hb-mediated toxicity (6, 40), cell-free Hb is rapidly bound by the acute-phase protein Hp with a high affinity (Kd, 1 pM) (15) and subsequently removed from the circulation by tissue macrophages in, for example, the spleen and liver via CD163-mediated endocytosis (17, 28). It has also been reported that, in addition to the high-affinity binding of the HpHb complex (Kd, 12 nM), Hb alone, but not Hp, also interacts with CD163, although via a low-affinity binding (Kd, 370 nM) (30, 42). Consequently, it is generally accepted that the main function of HpHb complex formation is to restrain the filtration of Hb in renal glomeruli and to accelerate removal of cell-free Hb by CD163-mediated endocytosis. The function of CD163 in relation to Hb metabolism is largely based on studies on human metabolism (17, 26, 42), which in several ways differs from other mammalian systems; especially with regard to the Hp system, which differs among species in terms of multimerization and the expression level during the normal and acute phase. The overall structure of Hp (2) seems rather conserved based on sequence similarity. However, in contrast to most other species, human Hp exist in variant multimeric forms because of two different hp gene alleles. Moreover, an Hp-related protein with retained Hb binding but no CD163 binding has evolved in old world primates (27, 29). The expression of Hp as an acute phase protein is conserved among mammals, whereas the normal plasma concentration of Hp is much higher in humans compared with most other species (8, 16). In fact, some species of mice have been reported to have very low or virtually absent Hp expression under normal conditions (32). In rodents, the induction of Hp expression during the acute phase is several folds higher than in humans, thus potentially neutralizing some of the species-specific differences during inflammatory conditions (16). The variation in Hp expression levels between human and rodents may reflect species-specific differences in the regulatory elements governing Hp expression. In support of this, a stimulatory effect of glucocorticoids and IL-1 has been reported in rodents, whereas a diminished effect of these cytokines is seen in humans (4, 31).
Despite the many studies of Hp in mice including Hp-deficient mice (20), the CD163-mediated clearance has never been characterized. In the present study, we have, therefore, performed a comparative in vitro analysis of the CD163-mediated uptake of Hb in man and mouse. Moreover, to examine the physiological importance of CD163 in the clearance of Hb in mice, a CD163-deficient mouse line has been established and analyzed.
Results
The binding of mouse Hb (mHb) to mouse CD163 (mCD163) was analyzed by surface plasmon resonance (SPR) and compared with binding to the human CD163 (hCD163) system (Fig. 1).
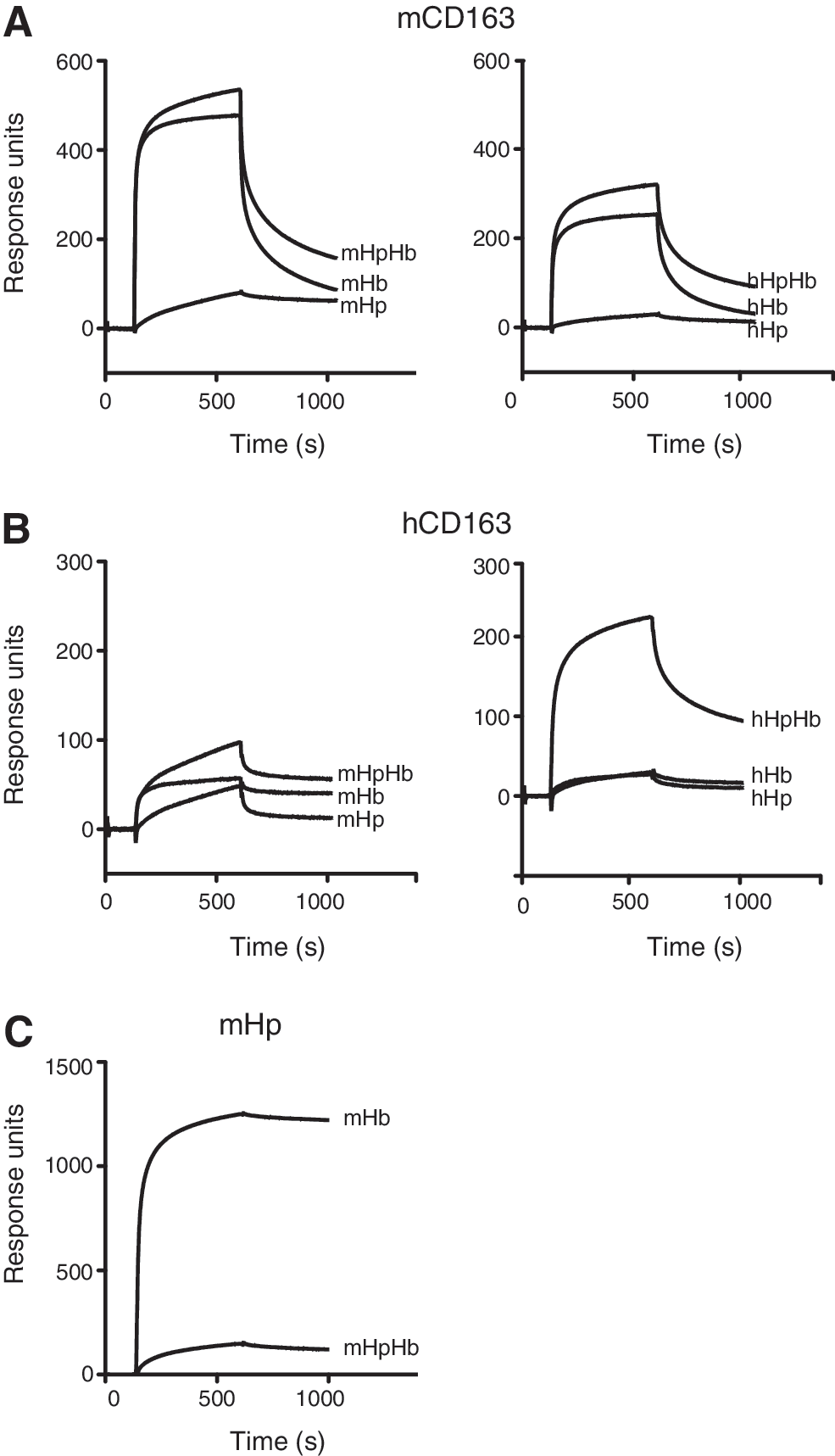
Figure 1A shows that both the noncomplexed mHb and the mHpHb complex bind to immobilized mCD163 with a comparable affinity (mHb; Kd 61 nM and mHpHb; 18 nM). In contrast to this and in agreement with previous observations (17, 30), an SPR analysis of human Hb (hHb) and human HpHb (hHpHb) binding to immobilized hCD163 recorded a high-affinity binding of the hHpHb complex (Kd 19 nM) but only a low-affinity binding of free Hb (Kd 198 nM) (Fig. 1B). Cross-species analysis revealed that the binding of hHb and hHpHb to immobilized mCD163 (hHb; Kd 159; and hHpHb; Kd 30) was similar to that of the mHb and mHpHb counterparts (Fig. 1A). However, only a very weak binding of mHb and mHpHb to immobilized hCD163 was observed (Fig. 1B). This indicates that species differences in both the ligands and the receptors contribute to the observed differences between mouse and man. The differences were not due to absent HpHb complex formation, as an SPR analysis showed a strong and almost irreversible binding of mHb to immobilized mHp (Fig. 1C) as previously demonstrated for the human protein counterparts (29). Likewise, sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) analysis eliminated the possibility that the observed differences were due to a contamination of the mHb sample with preformed mHpHb (Supplementary Fig. S1; Supplementary Data are available online at
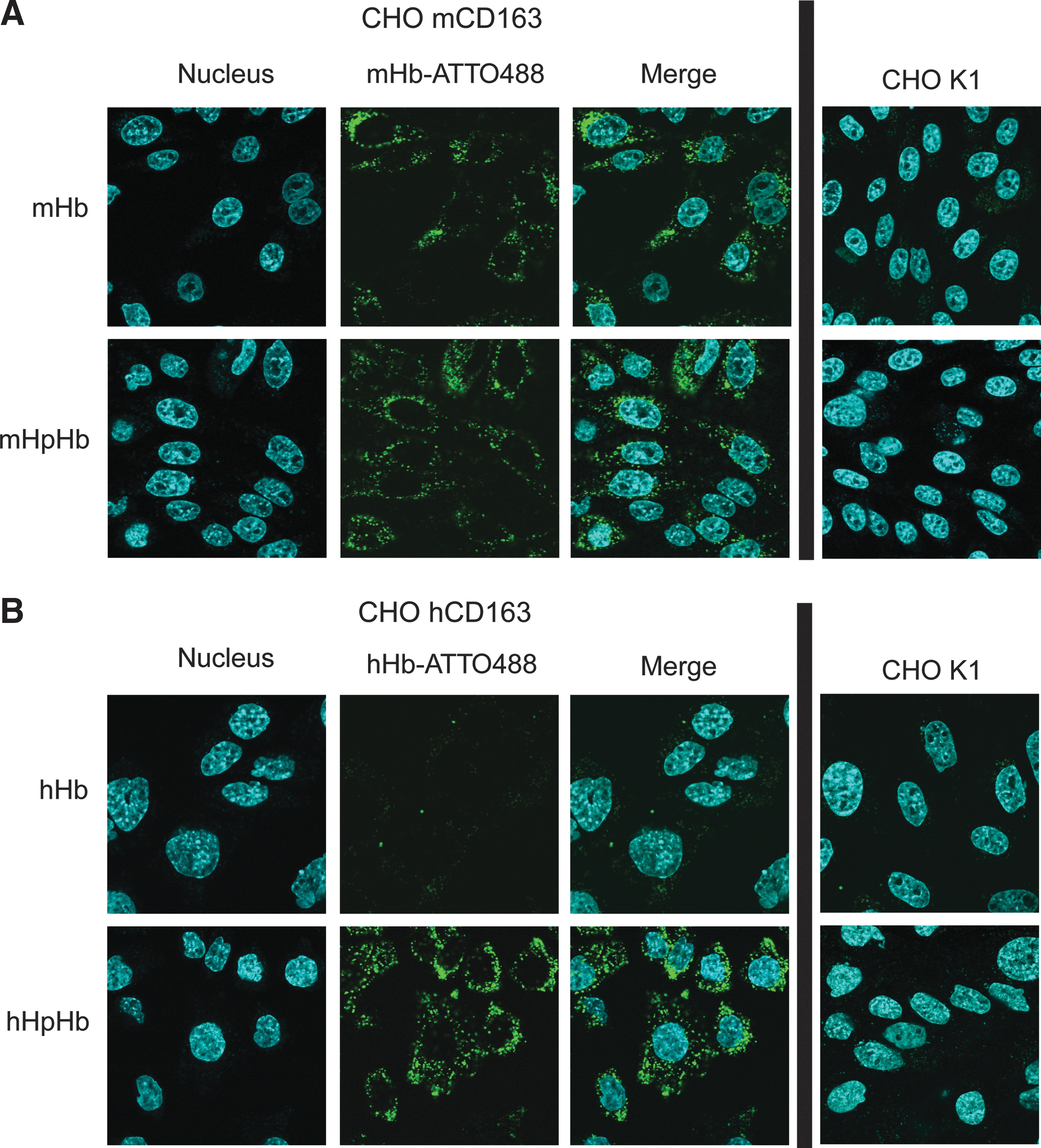
To study the CD163-mediated uptake of mHb in vitro, a transfected Chinese hamster ovary (CHO) cell line stably expressing mCD163 was generated, and the uptake of mHb covalently labeled with the fluorescent dye ATTO488 (mHb-ATTO488) was analyzed by comparing the mCD163-mediated uptake of mHb-ATTO488 to the hCD163-mediated uptake of hHb-ATTO488 (Fig. 2). The modification of mHb with ATTO488 did not alter its CD163-binding characteristics (Supplementary Fig. S2). When incubating CHO cells expressing mCD163 with mHb-ATTO488 or mHpHb-ATTO488, a high increase in cell-associated mHb was observed after 6 h (Fig. 2A). However, more than 10% of added mHb-ATTO488 was taken up after 6 h, whereas only 6% uptake of mHpHb-ATTO488 was measured (Fig. 2A). When adding hHb-ATTO488 to CHO cells expressing the hCD163 protein, no significant uptake was detected after 6 h (Fig. 2B). In contrast, when incubating hCD163-expressing cells with hHb-ATTO488 in a preformed complex with hHp (hHpHb-ATTO488), nearly 15% uptake was measured after 6 h (Fig. 2B). For both Hb- and HpHb-ATTO488, only low uptake was measured in nontransfected control cells. The CD163-mediated uptake of nonconjugated and Hp-conjugated mHb-ATTO488 was verified by confocal microscopy, showing cytoplasmic location of both mHb- and mHpHb-ATTO488 in mCD163-positive CHO cells (Fig. 3A); whereas cytoplasmic localization of hHb-ATTO488 was identified only when incubating hCD163-positive cells with a preformed hHpHb complex (Fig. 3B).

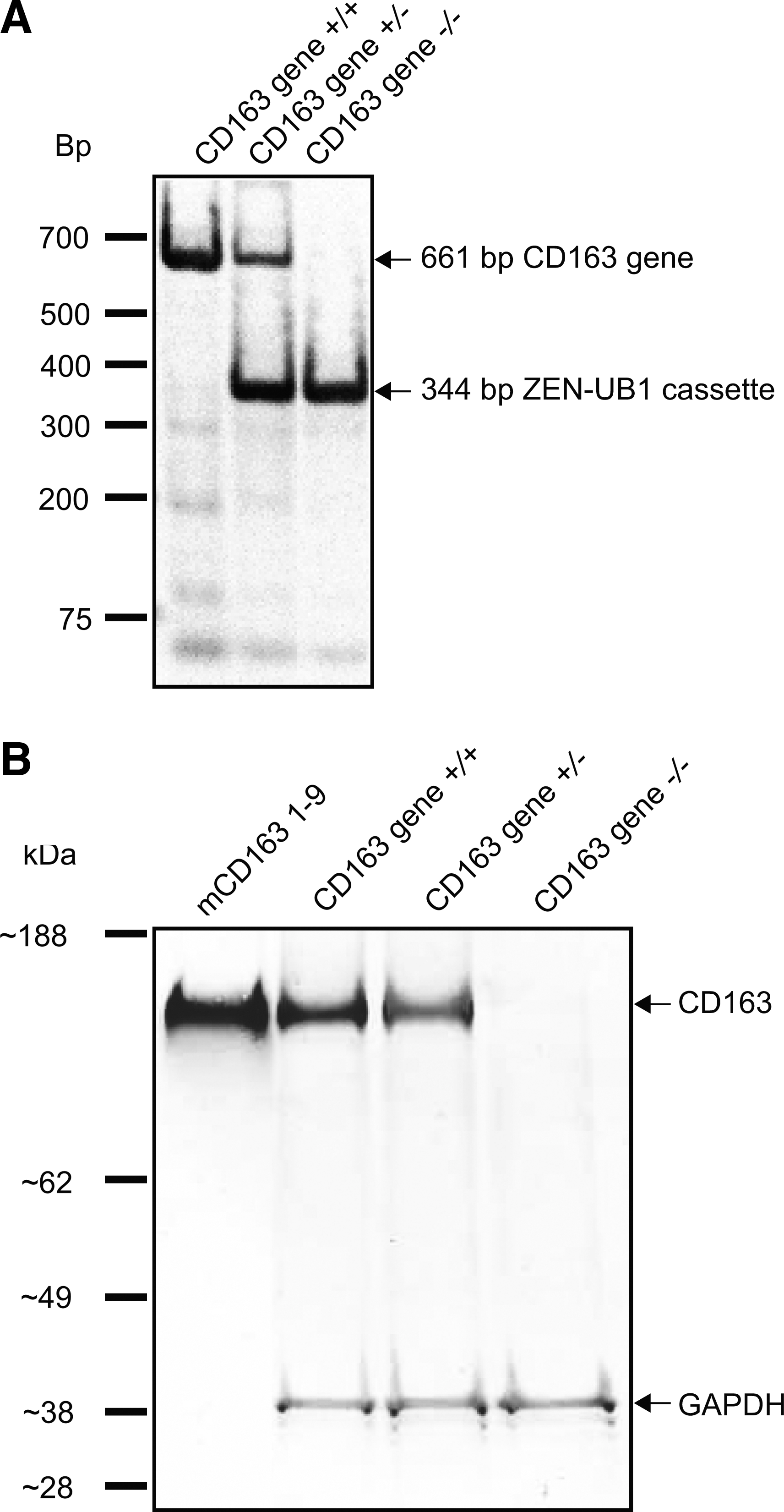
To analyze the CD163-mediated clearance of mHb in vivo, we compared the plasma clearance of mHb-ATTO488 in CD163-deficient mice (CD163 gene−/−) and normal littermates (CD163 gene+/+). The mCD163 gene was deleted in the VGB6 ES cell line by homologous recombination of the CD163 gene with the ZEN-UB1 cassette using a BAC-based targeting vector. Positive recombinant clones were used to generate chimeras in C57BL/6NTac mice, and resulting heterozygote mice (+/−) were intercrossed to produce homozygote (−/−) CD163 null progeny and control mice (+/− and +/+). Genotyping of the resulting offspring revealed that ∼50% of the offspring were heterozygotes, ∼25% were (+/+) mice, and the remaining ∼25% harbored the desired deletion of both CD163 gene alleles (Fig. 4A). Western blotting analysis identified the CD163 protein in the liver of the CD163 gene+/+ and CD163 gene+/− mice (reduced expression), whereas no CD163 was detected in CD163 gene−/− mice (Fig. 4B). Immunostaining of the liver in normal mice (CD163 gene+/+) demonstrated macrophage staining that was similar to what has been reported in the human and rat liver (Supplementary Fig. S3) No apparent phenotype changes in the CD163 gene−/− mice compared with the CD163 gene+/+mice were observed, and offspring were viable and fertile.
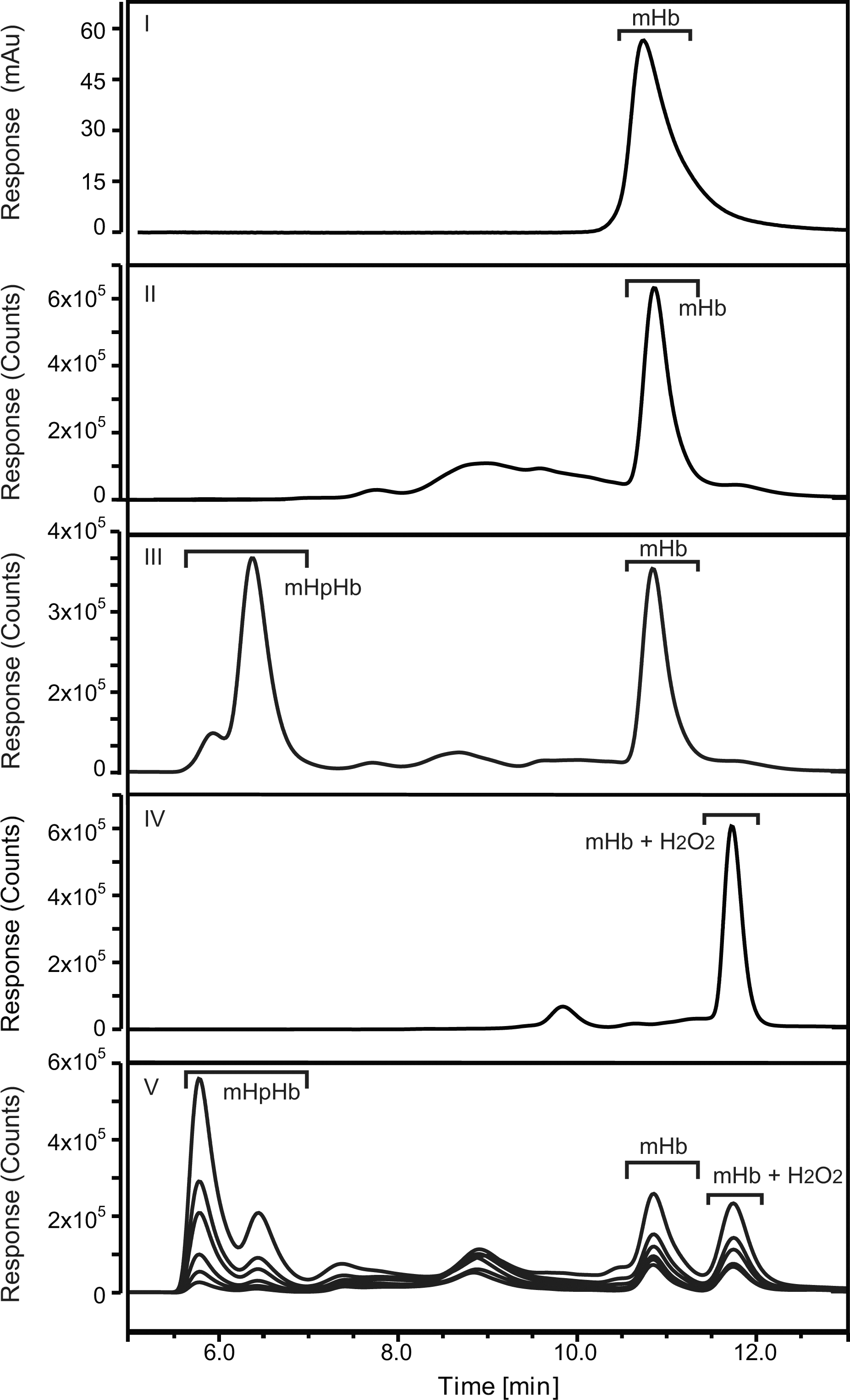
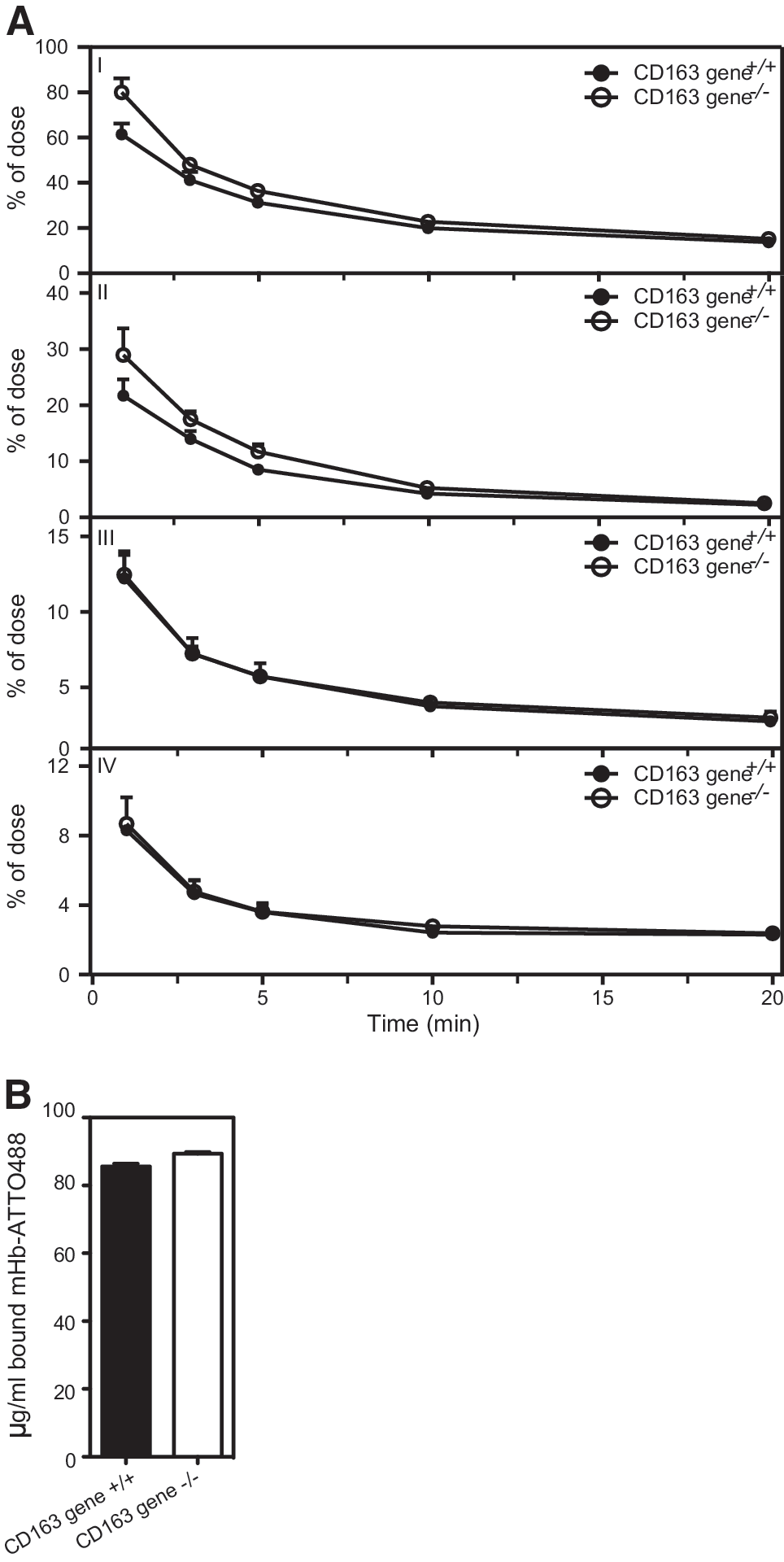
CD163 gene+/+ and CD163 gene−/− littermates were injected intravenously in the tail vein with mHb-ATTO488 (0.1 mg/10 g bodyweight). The concentration of injected mHb-ATTO488 was measured at different time points (1, 3, 5, 10, and 20 min) by high-pressure size-exclusion chromatography (HPSEC)-coupled fluorescent analysis of collected serum (Fig. 5). Clearance of injected mHb-ATTO488 fluorescence in CD163 gene+/+ mice followed a two-phase decay (Akaike's Informative Criteria [AICc]: 99.71%, R2: 0.9769) with a fast half life of 0.4 min and a slow half life of 3.6 min (Fig. 6A, panel I and Supplementary Fig. S4). In contrast, serum analysis of injected mHb-ATTO488 in CD163 gene−/−mice showed that the clearance of mHb was described by a one-phase decay (AICc: 88.60%, R2: 0.9686) with a half life of 2.3 min (Fig. 6A, panel I and Supplementary Fig. S4). Interestingly, the clearance of injected mHb-ATTO488 in complex with Hp (mHpHb-ATTO488) was also genotype dependent (Fig. 6A, panel II); whereas the clearance of free Hb in plasma was similar for both CD163 gene+/+ and CD163 gene−/−mice (Fig. 6A, panel III). In addition to free mHb and mHpHb, the HPSEC-coupled fluorescent analysis also identified a serum-specific fluorescent peak (Fig. 5, panel V) with a longer retention time compared with both mHpHb and free mHb (Fig. 5, panel III). This peak was similar to that of mHb-ATTO488 that was preincubated with a molar excess of H2O2 (Fig. 5, panel IV) and may, therefore, represent oxidized mHb-ATTO488. The level of the presumed oxidized mHb-ATTO488 that was formed and its clearance rate were not influenced by CD163 expression (Fig. 6A, panel IV). The observed difference in Hb clearance was not caused by a genotype-specific change in the binding capacity of cell-free Hb (Fig. 6B), as the Hb-binding capacity in the serum of both CD163 gene+/+ and CD163 gene−/− mice was ∼90 μg/ml. When analyzing the tissue distribution of injected mHb-ATTO488 ex vivo (Fig. 7A), a genotype-specific difference was measured in the liver after 2 min (Fig. 7B, C) and in the kidney after 10 min (Fig. 7B, C). The highest specific fluorescence activity after 10 min was measured in the kidney (Fig. 7C).

Discussion
The present data reveal distinct and important differences between CD163-mediated scavenging of hHb and mHb. Interestingly, we found that the CD163-mediated scavenging of Hb in mice is largely independent of HpHb complex formation. This actually concurs with a study on Hp knockout mice, which shows that the clearance rate of Hb from the circulation was independent on Hp (20). Instead, it was observed that cell-free Hb accumulated in the kidneys to a high extent on Hp knock down. Based on this, it was suggested that the function of Hp in mice in relation to Hb elimination is not to accelerate the clearance of cell-free Hb but to avoid its excessive filtration in the kidney (20). The present data support this. Furthermore, the present data in mice also indicated that the oxidized form of hemoglobin (methemoglobin) has an extended clearance rate. If this applies to a human setting, the extended clearance may have major implications in diseases with high levels of methemoglobin (such as malaria and inherited methemoglobin). In addition, the data also suggest an indirect positive role in Hb elimination of other endogenous anti-oxidative agents such as ascorbate and gluthathion.
In the present article, an Hb concentration of 0.1 mg/10 g bodyweight was used to study the clearance rate of cell-free Hb. The injected dose of Hb constituted ∼1‰ of the total Hb content in mice (37). Earlier reports have suggested that the Hb-binding capacity at a steady state in mice is close to 0.05 mg/10 g bodyweight (16). Consequently, the dose of Hb used in the present study was in excess of the available Hp, allowing for the clearance rate analysis of both HpHb and free Hb.
The deletion of CD163 expression in mice resulted in a significant although minor delay in the clearance of HpHb complexes from the circulation. The highest uptake was seen in the liver and kidneys. Despite a relatively high uptake in the liver, significantly less mHb was accumulated in the liver of CD163−/− mice after 2 min. In line with this, significantly more mHb was accumulated in the kidneys of CD163-null mice after 10 min. The observed differences in mHb clearance were not due to the changes in available mHp, as the ratio of mHb/mHpHb in serum was similar in CD163 gene−/− and CD163 gene+/+ mice. However, the high uptake in the kidney and a similar clearance rate in both CD163-null and wild-type mice suggest a high clearance of free Hb by renal filtration. The almost similar uptake in the CD163-deficient liver tissue suggests alternative routes for HpHb uptake in mice. In line with this, previous studies in rats have indicated the existence of a CD163-independent mechanism for the uptake of Hb and HpHb complexes in hepatocytes (45). Whether an alternative pathway for hepatic uptake of Hb and HpHb may also exist in humans, and if so, has any physiological role, is not known. However, previous attempts to purify HpHb receptors by HpHb-meditated affinity chromatography of different human tissues, including liver, did only identify CD163 as a high-affinity receptor (17).
The present and previous data underscore substantial differences in the human and rodent Hp-related biology. In addition to the differences just mentioned related to CD163-mediated clearance, it should also be noted that the Hp concentration at a steady state and therefore the Hb-binding capacity is ∼10-fold lower in mice compared with humans (8, 16). In contrast, the up-regulation of Hp expression during the acute-phase response is much higher in mice and rodents in general (16), as compared with humans, possibly neutralizing some of the species-specific differences during inflammatory conditions. In addition, gene duplications in the human Hp allele have resulted in two Hp variants (Hp1 and Hp2) (46). Earlier reports have associated the different Hp variants with increased susceptibility in various diseases such as diabetes, malaria, tuberculosis, and others (19, 35). Moreover, old world primates including homo sapiens, have an additional Hp protein designated the Hp-related protein (22). This protein only differs by 23–30 amino acids compared with Hp1. Hp-related protein, which has retained the Hb- but not the CD163-binding properties (29), plays a substantial role in the immune defense against certain forms of trypanosome parasites, causing sleeping sickness in nonprimate animals (44)
Taken together, primate evolution, including the late homo sapiens evolution, seems to have caused a particular pressure on the Hp system, resulting in new and species-specific functions, although all known Hp variants still relate to hemolysis and Hb binding. It is tempting to speculate that the serious infectious hemolytic diseases, such as malaria and sleeping sickness in the central part of Africa, might have caused a strong evolutionary pressure on the Hp system in primates and perhaps other mammals. In addition, a high constitutive intravascular hemolysis as seen in humans (10%–20% of erythrocytes/erytroblasts undergo lysis in circulation) may have pushed evolution into the development of a high-affinity clearance system such as the CD163-mediated HpHb system in humans. However, from a phylogenetic point of view, CD163 was most likely initially evolved as an Hb-binding protein and not as an HpHb receptor.
The differences in Hb clearance between human and mouse may be taken into consideration in future translational studies, including the development of novel drugs and therapies inflicting with Hb metabolism (10). For instance, the mouse seems not to be an obvious animal model for preclinical proof-of-concept studies of Hb-based oxygen carriers to be used as alternatives to blood transfusion.
Materials and Methods
Cell culture, expression, and purification of mouse CD163 and Hb
Stably transfected CHO cells expressing either full-length or the extracellular domain (SRCR domain 1–9) of mCD163 were established by the transfection of a CHO K1 cell line with a pEF4/V5-His vector (Invitrogen, Taastrup, Denmark) containing mCD163 cDNA obtained from Open Biosystems (VWR—Bie & Berntsen A/S, Herlev, Denmark). Single clonal cell lines stably expressing mCD163 were generated by antibiotic selection using 300 μg/ml Zeocin (Invitrogen). Serum-free CHO SFM II medium (Invitrogen) was collected from cells expressing mCD163 SRCR domain 1–9. Protein was purified using a His-trap column (Qiagen Denmark, Copenhagen, Denmark) and eluted in 250 mM imidazole. Hb was purified from washed mouse erythrocytes by adding 1 eq. H2O, collected by centrifugation at 13,000 rpm at 4°C, and finally dialyzed against 1×PBS pH 7.4. mHp and hHp (1–1) were obtained from Cell Sciences (Canton, MA) and Sigma, respectively. HpHb complexes were formed by mixing Hp and Hb 1:1 (w/w) followed by incubation for 30 min at 4°C. Flp-In CHO cells stably transfected with hCD163 were previously described (26).
SPR binding studies
SPR analysis was carried out as described (21) using a Biacore 3000 instrument (Biacore, Uppsala, Sweden). mCD163 was immobilized in 10 mM sodium acetate, pH 4.0, and remaining binding sites were blocked with 1M ethanolamine, pH 8.5. The resulting densities were close to 70 fmol protein/mm2. Sensorgrams were generated using protein concentrations in the range 5–25 μg/ml and CaHBS with 2 mM free Ca2Cl2 (10 mM Hepes, 150 mM NaCl, 3.0 mM CaCl2, 1.0 mM EGTA, +0.005% P20, and pH 7.4) as running buffer. Kinectic parameters were estimated using the BiaEvaluation ver. 4.1 software (Biacore) and the Langmuir binding model for the fitting of experimental data.
Confocal imaging and fluorometric assays
Before fluorescent assays, Hb was conjugated with ATTO488 (Sigma-Aldrich, Copenhagen, Denmark) according to the Sigma-Aldrich protocol. For quantification of Hb uptake in CHO cells expressing CD163, cells were seeded in a black clear-bottom 96-well plate (Sigma-Aldrich) at 10,000 cells/well and incubated with Hb-ATTO488 or preformed HpHb-ATTO488 for varying time points (0, 30, 60, 120, 180, 240, or 300 min). Cells were then washed thrice in a complete medium, after which fluorescence was measured using the Enspire multifluorescent plate reader at ex490/em515 (PerkinElmer, Skovlunde, Denmark). Confocal microscopy was done by seeding cells at 10,000 cells/well in LabTek II chambered coverglasses (Nunc A/s, Roskilde, Denmark) and incubated with either Hb-ATTO488 or HpHb-ATTO488 for 30 min at 37°C. Finally, cells were incubated with Hoechst 33248 (Sigma) and visualized using an Olympus FV10i confocal microscope (Olympus Denmark A/S, Ballerup, Denmark).
SDS-PAGE and immunoblotting
Proteins were separated by SDS-PAGE using a 12% NuPAGE Bis-Tris gel and MES running buffer (Invitrogen) and subsequently visualized by SimplyBlue SafeStain (Invitrogen). Immunoblotting was performed on 50 μg protein isolated from the mouse spleen using a polyclonal rabbit anti-mCD163 antibody directed against mCD163 SRCR1-9 (Dako Denmark A/S, Glostrup, Denmark) and a secondary alkaline phosphatase-conjugated goat anti-rabbit IgG antibody (Sigma Aldrich). Subsequent to stripping of the membrane by 30 min of incubation in 25 mM glycine-HCl (pH 2) and 1% SDS, followed by standard washing and blocking procedures, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was detected using a polyclonal anti-GAPDH antibody (Sigma).
Generation of CD163 gene KO mice
The CD163 null mouse strain was generated from targeted ES cells for CD163 (MGI:2135946) obtained from the Knock-Out Mouse Project (KOMP) Repository (
In vivo clearance and tissue distribution
Four wild-type mice and four CD163-null C57BL/6NTac mice received a single injection of ATTO488-labeled Hb (0.1 mg/Kg bodyweight) in the tail vein. Blood sampling was done under isoflurane anesthesia by orbital bleeding at 1, 3, 5, 10 and 20 min post injection. Serum was collected after the blood had been allowed to coagulate for 1 h at room temperature by centrifugation (3000 g, 15 min). 30 min post injection, the animals were sacrificed for an ex vivo examination of organs. Serum samples were analyzed using a biocompatible Dionex HPLC Ultimate 3000 system equipped with a fluorescent detector (Dionex Denmark, Hvidovre, Denmark) and a TSKgel SuperSW3000 column (Tosoh Bioscience GmBH, Stuttgart, Germany). 300 mM KCl, 20 mM Tris pH 7.4 was used as running buffer. An amount of mHb-ATTO488 at 0 min was calculated by measuring the concentration of mHb-ATTO488 in the injection solution by HPSEC divided by the distribution volume (8% of total bodyweight). For the detection of oxidized mHb, mHb-ATTO488 was treated with H2O2 (30% w/w; Sigma) before HPSEC analysis. The heme:peroxide ratio was 1:10 as previously described (43). An ex vivo examination of fluorochrome distribution in organs was performed in an IVIS® Spectrum imager (Caliper Life Science N.V., Teralfene, Belgium) equipped with filters for ex480/em540 imaging. The study was approved by the Danish Animal Welfare committee (jr. number 2010/561-1715).
Statistics
In vivo data were fitted using nonlinear regression of one or two-phase decays, and the best model was chosen based on corrected AICc method (1, 3). Differences among groups were analyzed using the Mann–Whitney U-test (p≤0.05). All statistical analysis were performed using GraphPad Prism version 5.0d for Mac Os X (GraphPad Software, San Diego, CA).
Footnotes
Acknowledgments
The authors wish to thank Anne Marie Bundsgaard and Gitte Petersen Ratz, Department of Biomedicine, Aarhus University, for excellent technical assistance. The present study was supported by grants from the Danish Medical Research Council, the Novo Nordisk Foundation, the European Research Council (Advanced Grant for the TROJA Program), and the Lundbeck Foundation.
Author Disclosure Statement
S.K.M. is a minority shareholder of the small biotech company Cytoguide Aps. Other authors have no competing interest.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.