
Abstract
Introduction
Different microbes encounter varying concentrations of environmental oxidants, reductants, and diatomic gases during growth. Not surprisingly, aerobic microbes have evolved sensing mechanisms to monitor and balance intracellular redox fluctuations to maintain cellular integrity (22). Similarly, anaerobic organisms also require “sensors” that respond to lethal concentrations of O2 or reactive oxygen species (ROS), while facultative aerobes have developed systems that allow switching from one metabolic state to another based on the concentration and availability of oxygen (26). These sensors usually respond to varied redox signals by altering the expression of specific genes that allow the organism to survive in vivo.
Environmental sensor proteins that harness the oxidation-reduction property of transition metals, such as heme iron, at their active centers are abundant and well-studied in bacteria. Iron is exquisitely sensitive to the cellular redox environment, and the selective oxidation, reduction, or chemical modification of this element can cause a rapid change in signal transduction in response to redox fluctuations. Notably, in aerobic microorganisms, the production of oxidants (e.g., ROS) is not perfectly balanced by antioxidants that dampen but do not eliminate effects of ROS. As a result, if the balance is severely disturbed, a state of oxidative or reductive stress follows [(29), see (15) for a review on reductive stress].
In this review, we refer to environmental heme-based sensor proteins as proteins that coordinate a ferric or ferrous iron at their active site that reacts specifically with diatomic gases. The heme center in most sensor proteins is a regulatory rather than a catalytic domain. In these proteins, oxidation, reduction, or modification of heme generally results in an alteration in protein activity. Many microbial sensor proteins have been shown to bind O2, NO, and/or CO via their heme group (65), which causes structural changes to the heme that ultimately regulate protein activity (13, 50). NO and CO are particularly important as they are physiologically relevant and generated by human iNOS (inducible nitric oxide synthase) and HO-1 (heme oxygenase-1), respectively, as part of the protective response against microbial invasion (44) or oxidative stress (67). Since Mtb requires O2 for growth, and since reactivation of disease occurs primarily in the oxygen-rich upper lung lobes (54), O2 is regarded as an important pathological factor during Mycobacterium tuberculosis (Mtb) persistence (46).
Notably, from a human pathogenesis point of view, there is little doubt that an exciting, although mostly unexplored area of research, is the identification of heme proteins that specifically sense and respond to host gases to affect virulence. Further, defining the mechanisms by which bacterial heme proteins sense and respond to O2, NO, and CO via their heme irons to avoid the onslaught of the host immune system is a technically challenging and largely overlooked area of research.
We will begin this review by describing the basis of heme iron sensing mechanisms by focusing on several well-characterized heme sensor proteins. This is then followed by a discussion on the current knowledge of bacterial heme proteins capable of sensing O2, NO and CO. We also highlight recent discoveries regarding the Mtb heme sensor kinases DosS and DosT [originally designated Dev by Kinger and Tyagi (43)]. This review is not meant to be a comprehensive summary of all heme sensor proteins; rather, our intention is to focus on heme-based NO, CO, and O2 sensing mechanisms to provide a better foundation for future interpretation of the physiological events associated with bacterial infection. With respect to the role of iron-based redox sensors in biology, and how a family of iron–sulfur cluster proteins modulates redox homeostasis in mycobacteria, we refer the reader to other reviews that discuss these issues in detail (45, 59, 69).
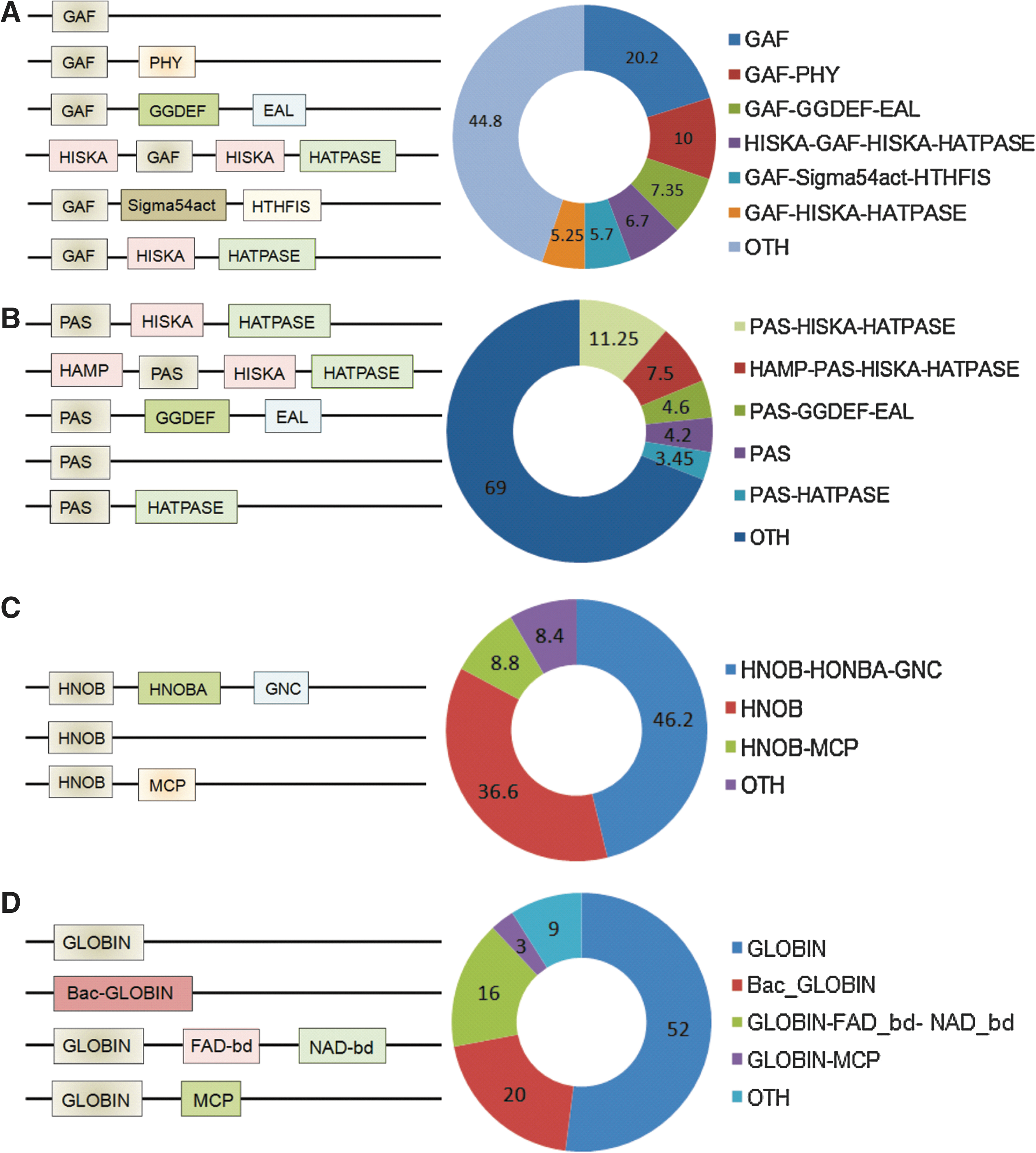
Distribution of Heme-Based Sensors
Heme-based sensor proteins have a uniform distribution across all forms of life. To date, six different types of heme-binding domains have been identified in sensor proteins; namely, (i) heme-nitric oxide binding domain [H-NOX], (ii) the globin fold, (iii) the heme binding Per-Arnt-Sim (PAS) fold, (iv) the GAF domain (present in cGMP-specific phosphodiesterases, adenylyl cyclases, and FhlA proteins), (v) CooA (bacterial CO oxidation transcriptional activator) homologues of the CRP/FNR family, and (vi) the recently discovered SCHIC (Sensor Containing Heme Instead of Cobalamin) domain of PpaA/AerR family members of anoxygenic phototrophic proteobacteria (22, 55). Despite having heme as co-coordinating ligand, these proteins harbor tremendous diversity in terms of their domain architectures (Fig. 1) with the highest diversity being in the PAS fold. These variations in protein tertiary structure may in part explain the diverse role played by heme-containing proteins in living systems. For example, heme can be located within: (i) GAF domains (e.g., Mtb DosS and DosT), (ii) PAS domains (e.g., FixL in Rhizobium and EcDos in Escherichia coli), or (iii) in typical globin domains (e.g., Bacillus subtilis HemAT). The PAS domain usually uses heme cofactors, while GAF domains (containing a six-stranded anti-parallel β-sheet core) are frequently associated with binding of cyclic nucleotides. Our bioinformatic analysis of the PFAM/InterPro protein database revealed that ∼45% of all known proteins with GAF domains act as two-component systems while ∼23% act as phosphodiesterases (Fig. 1).
Reactivity of Heme Iron for NO, CO, and O2
The extraordinary redox chemistry of heme is exploited by several bacteria to sense and respond to variable levels of O2, NO, and CO. These heme-based sensors usually harbor heme-b (protoporphyrin IX) in its iron-bound form, as a prosthetic group in their active site. Heme b, is a tetradentate chelating ligand that coordinates a central iron ion in four histidine-iron (His-N-Fe) coordinate bonds (17). The typical structure of heme-b outside of a protein matrix is in the form of a planar ligation of the iron ion by protoporphyrin IX. However, heme may acquire nonplanar structures when present within a protein. The heme group is usually embedded within the protein's hydrophobic core and is not solvent accessible. The interaction of heme with the protein is through axial binding between the heme iron and amino acid residues, usually by one or two bonds. In the majority of heme proteins, this interaction occurs via His residues although heme may interact via thiolate ligation through Cys, Met, and Tyr residues (86). The Fe present at the center of the heme porphyrin is capable of coordination with six ligands: (i) four coordinate with the heme porphyrin ring; (ii) one with a protein His or Cys side chain residue, and (iii) one with an exogenous ligand or with another amino acid side-chain residue. This sixth binding site in some heme proteins is capable of binding diatomic gases O2, NO, or CO (5) (Fig. 2). It is the capacity of heme to switch between coordination states by virtue of its ligation with Fe that provides the redox sensing capability of various heme-containing regulatory proteins (59). Additionally, the amino acids surrounding the sensor protein's heme group are responsible for the specific reactivity of each protein towards its ligand (38). In bacteria, such proteins usually contain any one of the three known sensor domains that bind heme; namely, the PAS, GAF, or a typical globin domain. These sensor domains are usually linked to an effector domain such as histidine kinase, phosphodiesterase, or diguanylate cyclase (22, 60).

Representative proteins from four known families of bacterial heme-based sensor proteins have been characterized, which has provided substantial insight into their structure and function: the bacterial O2 sensors FixL and HemAT, the CO sensor CooA from Rhodospirillum rubrum, and the heme-NO/O2 binding H-NOX sensors found in facultative aerobes, obligate anaerobes, and thermophiles (24, 75). Though these proteins use the same protoporphyrin IX heme cofactor, they harbor different heme-binding protein scaffolds. HemAT and FixL have a globin fold and a PAS domain fold, respectively, whereas CooA belongs to the cAMP receptor protein family of transcriptional regulators (65). The mechanism of how these sensors function will be discussed later in this review.
Heme-Based Sensing Mechanisms
Heme-based sensor proteins usually function via a two-step process that includes ligand binding to the heme within the heme pocket and its subsequent release or exchange at the active site of the sensor protein. This process occurs at a femtosecond or picosecond scale and may induce a conformational change in the vicinity of heme group. While these ultrafast dynamics cannot be exactly reproduced in experimental settings, they can be studied by mimicking the actual effects.
Heme-based sensors consist of a sensory domain (i.e., a PAS domain) and an activator/transmitter domain. The binding of diatomic gas ligands (NO, CO, O2) to heme induces conformational changes in protein structure that either activate or inhibit the activator/transmitter domain (13). Heme sensors are highly sophisticated nano-sensors that respond to a specific signal with or without differentiating among its ligands for binding with heme. Several sensors have evolved to modulate their selectivity for certain gases, which is critical for their physiological activity and integral to downstream signal transduction (38). The major gas ligands, NO, CO, and O2, differ by only one valence electron. When His is the proximal binding site, the ratio of binding of the three gases remains nearly the same, which is 1:∼103:∼106 for NO, CO, and O2, respectively (83). A recent study demonstrated that selectivity for the ligand is directed via the sliding scale rule, which indicates parallel lines for plots of log K D for NO to CO to O2 and allows a prediction of the affinity of a specific ligand if the KD for the other ligand or ligands are known (83). The hydrogen bond donors present at the active site of the sensor deviate from this rule and thus increase O2 affinity with minimal effect on CO and NO binding. Thiolate, tyrosinate, and imidazolate ligands, on the other hand, induce a leveling effect on the binding affinity of the sensors. In the case of soluble guanylate cyclase (sGC) and other major NO sensors, NO binding drastically differs from the sliding scale rule due to the multistep mechanism involved in binding that leads to a change in coordination state of the protein from 6-coordinate to 5-coordinate, wherein it excludes O2 binding from the active site in order to capture lower levels of NO and avoid its oxidation by dioxygenation (83).
Heme-based sensors are either able to differentially bind and be activated in response to specific gas ligands, or does not differentiate among ligands for binding. For example, sGC can bind and be activated in response to NO and CO, but does not bind or gets activated in response to O2 (79). On the other hand, FixL does not differentiate among ligands for binding and can bind to NO, O2, and CO (26), but its activity is inhibited only upon binding with O2 (21, 23). Thus, in the case of heme-based sensors, the reversible binding of O2 to heme iron to generate a Fe-O2 bond or the oxidation of iron by O2 serves as a sensing mechanism that leads to the switch between the active and inactive forms of the protein.
Bacterial Heme-Based Sensors
A number of bacterial heme-based sensor proteins have been characterized including FixL of Rhizobium, direct oxygen sensor of Escherichia coli (EcDos), AxPDEA1 of Acetobacter, HemAT of Halobacterium, DosS and DosT of Mtb, and AvReg of Azobacter. This section will describe these classic heme-based sensors of O2, NO and CO and their general role in the biology of the microbe.
The oxygen-sensing heme protein FixL
FixL is a classic example of a bacterial heme-based O2 sensor. FixL and FixJ constitute a two-component system (TCS) in Rhizobium that regulates the expression of genes involved in nitrogen fixation and denitrification. FixL is the sensor kinase while FixJ is the response regulator in this TCS. FixJ directly regulates expression of nifA and fixK, which are transcriptional regulators of 21 genes involved in nitrogen fixation and denitrification (64). Since these processes are performed by enzymatic systems that are oxygen sensitive, the expression of genes involved in these pathways is negatively regulated by FixL-FixJ in response to O2. FixL senses the presence of O2 through the heme bound to its PAS domain. The heme iron of FixL can bind O2, NO, and CO (the affinities for NO and CO being higher) (26), but the kinase activity of FixL is inhibited only when bound to O2 (21, 23).
Recent studies have suggested that the dimer of FixL associates with the dimer of FixJ (84) to form a tetramer complex. This association allosterically enhances the auto-phosphorylation of FixL in the absence of O2. Evidence emerging from the crystal structure of different derivatives (ferric, ferric-cyanide, ferrous with O2, and NO) of the FixL heme-binding domain forms the basis of our current understanding of the mechanism of sensing by FixL. The oxy form of the protein complex bears heme iron in the low spin state, while the deoxy form has a high spin state. The transition in the spin state is believed to lead to alteration in the structure and activity of the protein (25). Due to this transition from high to low spin, the iron moves into the porphyrin plane, thus leading to the conformational change that drives the modulation of kinase activity. In the absence of structural data from full length proteins, it has not been determined how these conformational changes translate into modulation of kinase activity. Such studies would tremendously increase our understanding of the oxygen sensing mechanism of heme proteins.
Nitric oxide sensing proteins sGC and SONO
Soluble guanylate cyclase (sGC) is a classic example of a protein capable of sensing NO in the presence of up to a 1000-fold molar access of O2. sGC has a H-NOX domain that contains heme capable of detecting low levels of NO. A search for the H-NOX domain in prokaryotes resulted in the identification of a large number of heme-based sensor proteins (37). Some of these proteins sense NO in the presence of abundant O2, while others sense O2 itself. The ligand discrimination properties of H-NOX proteins are primarily dependent on the presence of a distal Tyr in the heme pocket. Examples of H-NOX-dependent NO sensors include VF_A0071 of Vibrio fischeri that modulates symbiotic colonization of Vibrio in the squid Euprymna scolopes (91), and SO2144 of Schewanella oneidensis that controls respiratory denitrification (39).
Another important protein that contains an H-NOX domain and plays a role in bacterial pathogenesis is the Clostridium botulinum sensor of nitric oxide (SONO) (56). SONO contains a methyl accepting chemotaxis domain in addition to an H-NOX domain, suggesting that it may help Clostridium to avoid NO. SONO is an extremely sensitive NO sensor and can detect femtomolar levels of NO in vivo. The crystal structure of SONO shows an unusual proximal αββαββ motif and a distal α-helical motif. The heme is sandwiched between the distal and proximal domain and is ligated to His. The heme pocket is highly nonpolar, thereby providing selectivity for NO over O2. SONO is also unusually resistant to oxidation in the nitrosyl-heme form with a half-life of ∼70 hours; however, NO-stripped SONO is sensitive to air oxidation. The crystal structure of SONO with substitution of Tyr139 with Phe generated a protein incapable of forming a 5c- heme nitrosyl, but forms a stable 6c Fe(II)-NO complex. This further demonstrates that Tyr139 in the distal pocket may uniquely modulate the bond strength of a proximal Fe-histidine linkage (56). The precise roles of newly discovered H-NOX-based sensor proteins in the pathogenesis of communicable human disease still remain to be discovered.
The carbon monoxide sensing protein, CooA
Rhodospirillum rubrum CooA is a model CO sensor and transcription regulator of the CRP/FNR family that regulates the expression of genes encoding both CO dehydrogenase and hydrogenase in Rhodospirillum (2, 75). These genes help Rhodospirillum to oxidize CO to CO2 coupled with the reduction of protons to H2, thereby allowing growth in the presence of CO as the sole source of energy. Since CO dehydrogenase is inhibited by O2, and because hydrogenase can function only in the presence of CO, CooA must function as a dual sensor of CO and O2 to ensure transcription in presence of CO under hypoxic conditions (30, 40). In contrast to FixL and sGC that sense the presence of O2 and NO, respectively, through the coordination state of their heme iron, CooA remains in a hexacoordinated low spin state in its inactive (ferric and ferrous) and active (CO-bound) forms. This suggests that binding of CO through replacement of an axial bond is sufficient to induce the transcriptional activity of CooA (3, 49, 76, 82). The most interesting property of CooA is its coordination chemistry. It is the only known heme protein that uses Pro as a ligand for heme (49). Several independent lines of evidence suggest that Pro2 and Cys75, Pro2 and His77, and His77 and CO are the axial ligands for the ferric form, ferrous form, and CO-bound forms, respectively (3, 49, 76, 82). During aerobic conditions, the inactive ferric form exist where heme is ligated to Pro2 and Cys75, while under anaerobic environment the reducing cytoplasm leads to the displacement of Cys75 by His77 and heme is bound to Pro2 and His77 (3, 49, 76, 82). Significant levels of CO in the anaerobic phase could displace Pro2 as an axial ligand. The specificity of CooA for CO is achieved by the ligand's ability to only displace Pro2. Subsequently, these changes in the coordination structure convert the inactive CooA into active CooA. Binding of O2 to CooA heme is not stable and leads to oxidation of the heme iron (3). The other important ligand that CooA binds is NO, which leads to dissociation of axial ligands (Pro2 and His77), thereby attaining a pentacoordinated inactive state (63). CooA is a unique sensor that senses the redox state of the cell and presence of CO for transcriptional regulation.
The redox sensing proteins CooA, EcDos, and RcoM
Heme-based sensors also exploit the redox chemistry of heme to sense the intracellular redox state of the bacterium. As described in the previous section, CooA is a classic example wherein the binding of O2 leads to oxidation of heme iron to generate Fe3+ and inactivation of protein function under normoxic conditions (3). This oxidation incorporates the redox state of cell into sensing capability of CooA. Oxidized CooA must be reduced (by as yet unidentified reductants) upon anoxia in order to bind CO. Other examples of proteins that may utilize redox active heme to sense the redox state of the cell is EcDos of E. coli and RcoM of Burkholderia (36, 52).
EcDos has two PAS domains, a diguanylate-cyclase (DGC) or GGDEF domain, and a diguanylate phosphodiesterase EAL domain. The GGDEF domain that enables synthesis of cyclic diguanylate is inactive due to the absence of key residues. The EAL domain is active and is required for specific phosphodiesterase activity for cAMP and c-diGMP. The first PAS domain binds a hexa-coordinated low spin heme at His77. EcDos is a tetrameric heme protein that can bind to O2, NO, and CO, or can exist in the ferric form (72). A redox-dependent ligand switch has been suggested for EcDos (47). The ferric form of EcDos is inactive whereas the ferrous form is active and can bind O2, NO, and CO (47). Interestingly, the cAMP phosphodiesterase activity of EcDos is activated by oxygen binding and inhibited by binding of NO, CO, or oxidation of its heme iron to Fe(III) (71). However, EcDos c-diGMP phosphodiesterase activity is activated by binding to O2, NO, and CO, and inactivated by conversion to the ferric form (80). This observation suggests that EcDos could potentially act as a redox sensor to modulate cGMP phosphodiesterase activity. However, it is important to note that the redox potential of EcDos is +67 mV (71), making it relatively resistant towards oxidation in vivo. Importantly, an E. coli EcDos gene knockout strain was defective in cell division and accumulated cAMP under aerobic conditions (93), suggesting that oxygen sensing either through the oxygen ligation state or redox state of EcDos plays an important role in the physiology of E. coli.
Another redox-mediated switch is RcoM of Burkholderia xenovorans (52). RcoM is a putative regulator of aerobic and anaerobic CO metabolism wherein the heme moiety is ligated to RcoM via Cys (thiolate) and His74. In the oxidized Fe(III) state, the protein is inactive. The reduction to the ferrous Fe(II) form leads to replacement of Cys (thiolate) with the neutral thioether ligand, Met104. The binding of the gas ligands CO and NO to the heme iron occurs via the displacement of Met104 (52). CO-bound RcoM gets converted to the Fe(III) form upon CO dissociation by incandescent illumination. The exposure of Fe(II) RcoM to oxygen leads to the formation of Fe(III) RcoM (41), suggesting that this protein may act as a redox sensor. B. xenovorans contains three Cys residues (Cys94, Cys127, and Cys130) and to date it is unknown which of these is the axial ligand to the met heme.
Biogenesis of Heme-Containing Proteins in Mtb
Initial comparative genomic studies using Hidden Markov Model-based profiling and COG (cluster of orthologous genes) analysis to identify heme uptake systems indicated that Mtb lacks a heme uptake system (7). However, a recent study has identified a mycobacterial heme acquisition system that may be important for meeting the iron requirements of Mtb. The Mtb heme acquisition machinery spans from (Rv0201–Rv0207), which encodes for a hemophore (Rv0203), and MmpL11 (Rv0202c) and MmpL3 (Rv0206), both of which belong to a family of mycobacterial transmembrane proteins that is involved in the transport of molecules such as heme across the mycobacterial membranes (85). See Figure 3 for a diagrammatic depiction of Mtb genes that participate in heme biosynthesis. The newly established Mtb hemophore (Rv0203) possess a unique fold with an unusual self-association, but structurally it contains a heme binding motif similar to that of S. marcescens HasA.
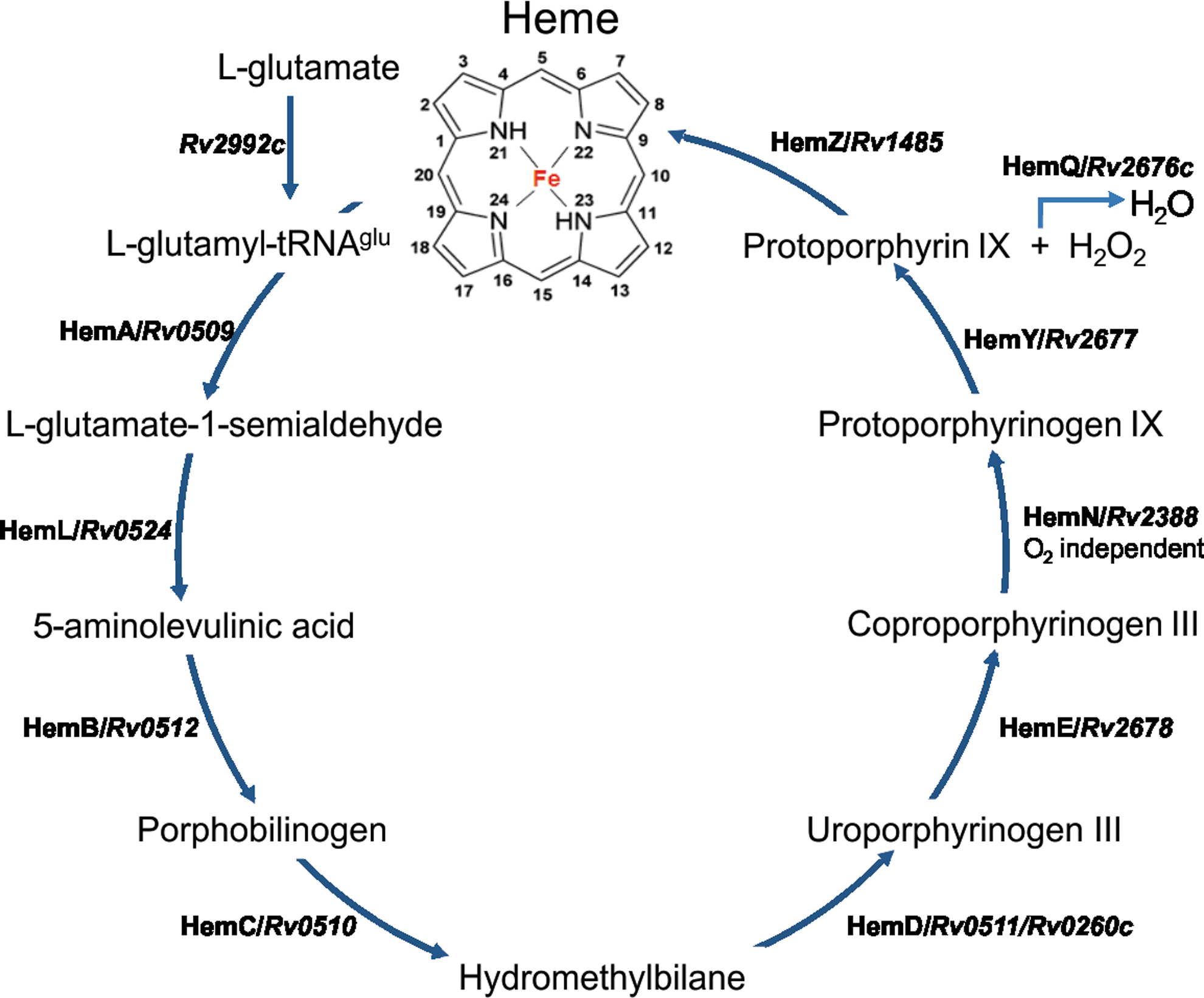
It is reasonable to assume that heme plays an important role in mycobacterial physiology; however, the genetics of the mycobacterial heme biosynthesis machinery is poorly characterized. Nonetheless, even though HemZ (a predicted ferrochelatase) is the only known gene biologically implicated in heme biosynthesis, our bioinformatic analyses have revealed that all orthologs required for heme biosynthesis are present in Mtb (Fig. 3).
Truncated Globins of Mtb
Globins are heme-binding proteins that are widely distributed among prokaryotes and can be classified into at least four main groups namely, flavohemoglobins, globin-coupled sensors, truncated hemoglobin, and protoglobins (18). Of these groups identified in prokaryotes, Mycobacteria possess truncated hemoglobins (trHbs) and flavohemoglobin that contain a globin domain coupled to a ferredoxin reductase such as FAD- or NADH-binding modules. Together, these proteins constitute some of the most widely studied globins among intracellular pathogens and prokaryotes. The trHbs are monomeric low-molecular weight oxygen-binding proteins that share overall topology with vertebrate hemoglobins but are typically 20–40 amino acids shorter and share less sequence identity. These proteins are characterized by a remarkable variability in the heme pocket residues among the various species, which may contribute to their diverse functions. We observed that over 95% of all known trHb proteins in protein databases harbor a single globin domain, even though some ‘‘chimeric’’ proteins have also been identified where the trHb domain is fused to an antibiotic biosynthesis monooxygenase or peroxiredoxin OsmC-like domain.
In Mycobacteria, the distribution of trHbs shares a regressive correlation with pathogenicity as their number decreases from the opportunistic mycobacterial species M. avium (three) to Mtb (two) to pathogenic M. leprae (one). The presence of trHb homologues in the extremely reduced genome of M. leprae suggests their critical role towards adaptation to a pathogenic lifestyle. Mtb truncated hemoglobins GlbN (trHbN, Rv1542) and GlbO (trHbO, Rv2470) are differentially expressed, as glbN is expressed only during the stationary phase while glbO is expressed throughout growth. The low sequence homology between the two trHbs in Mtb and their temporal expression indicate that they may perform different functions in Mtb (4) (Fig. 4). The trHbN protein demonstrates a potent nitric oxide dioxygenase (NOD)-like activity necessary for aerobic respiration as shown by inhibition of aerobic respiration by NO in M. bovis ΔglbN BCG cells (58). NO molecules preferentially act at the entrance of tunnel-like cavities present in trHbN (which contain an abundance of hydrophobic residues) that connect the active site to different protein surface sites. Since ligands such as NO have a higher solubility in hydrophobic environments, a buildup of NO at the entrance of the tunnel takes place, resulting in efficient capture of NO (73). This facilitates access of NO to the bound O2 without any structural changes and allows trHbN to catalyze (NOD)-like reactions at diffusion controlled rates (73). The trHbN of the saprophyte M. smegmatis lacks the N-terminal pre-A helix. In Mtb trHbN, the deletion of the N-terminal pre-A helix results in reduced NOD-like activity and decreased protection from NO toxicity compared to wild type (48), even though the role of the pre-A helix conformation in altering trHbN function remains controversial (73). While trHbN is involved in defense against nitrosative stress imposed by the host immune system, the function of trHbO remains debatable. Based upon the observation that expression of trHbO significantly enhanced cell growth, oxygen uptake rate and ATP levels in E. coli and M. smegmatis, Pathania et al. (61) showed that trHbO is involved in oxygen transfer. However, these findings were later contested by Outler et al. (58).

The Mtb trHbO promoter showed maximum induction under hypoxic conditions (62). Likewise in Frankia, which harbor truncated hemoglobins that are phylogenetically closest to their respective Mycobacterium orthologs, cultures grown in 1% O2 produce about 4 times more trHbO than cultures grown in 20% O2, suggesting a role for trHbO in adaptation to low oxygen concentrations (10). There are critical differences in the trHbN and trHbO amino acid sequences in different mycobacterial species that may contribute toward their distinct functions. Minor strain-dependent sequence length variation was also observed for Mtb trHbO (62). However, the precise mechanisms of action of mycobacterial trHb proteins remain unclear.
Bacterial flavohemoglobins (flavoHbs) share a common globin fold with trHbs, but differ in structural and functional aspects. Comparative studies of trHb and flavoHbs demonstrate a greater tendency of trHb to autoxidize compared to flavoHbs (16). It has been shown that flavoHbs are 1000-fold more efficient in converting NO and O2 to nitrate than in reducing NO to N2O, leading to their classification as NO dioxygenases (20). In Mycobacteria, two types of flavoHb (type I and type II) have been identified; one of them (type I) is restricted to saprophytic mycobacterial species, suggesting that type I and type II flavoHbs proteins may perform different functions (28). Even though several putative functions have been proposed for flavoHbs in other microbial species, their function in Mtb is not understood.
The Mtb Heme Sensor Kinases DosS and DosT
Approximately one-third of the world's population is latently infected with Mtb (12). Not surprisingly, a major question is to identify mechanisms that allow Mtb to persist in humans for decades without replication, to then abruptly resume growth and cause disease.
Studying the Mtb DosR/S/T (Dos) dormancy system may provide mechanistic insight into how Mtb enter, maintain, and emerge from a persistent state. The Dos dormancy system is a three-component system consisting of two histidine kinase sensors, DosS and DosT and a single response regulator DosR (68) (Fig. 5). DosS and DosR constitute a two-component system, whereas DosT is an orphan histidine kinase. A unique property of the Dos regulon is that both DosS and DosT are heme histidine kinases. The Dos regulon has been implicated in human TB, and the importance of this regulon in persistence was demonstrated in the rabbit and guinea pig models for TB [for a review, see (88)]. The Dos regulon was shown to be essential for anaerobic recovery and survival, and recovery from exposure to NO (51). The Dos regulon can be regarded as a genetic response induced by hypoxia, NO, and CO, wherein the gases inhibit aerobic respiration and prevent replication (51).
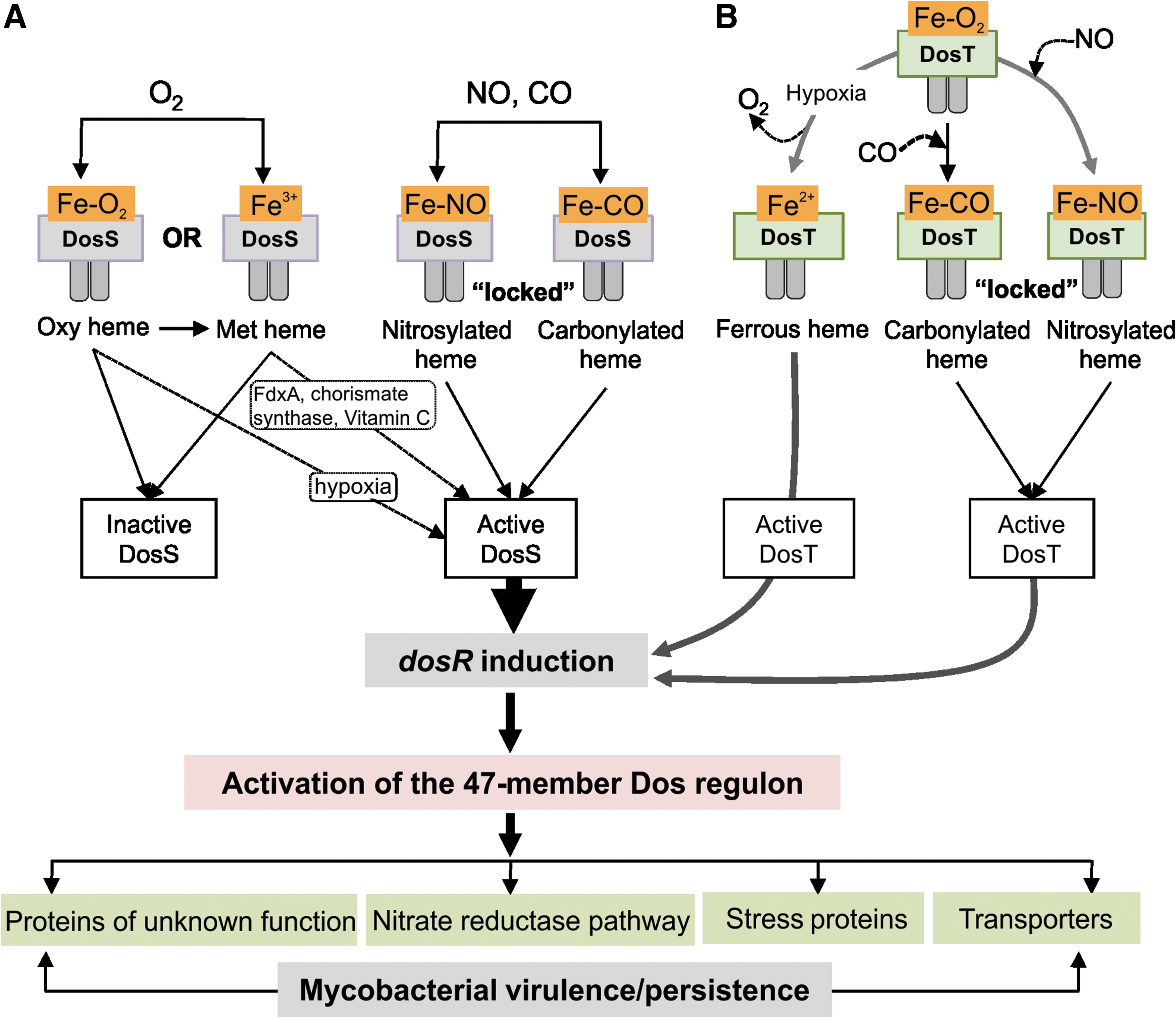
A fundamentally important discovery several years ago was that Mtb DosS and DosT responds to NO, CO, and low O2 levels (hypoxia) in a unique way to induce expression of the 47-member Dos regulon (44, 57, 77, 90). As described (for a review see (45), all three gases are physiologically relevant and implicated in Mtb pathogenesis. The induction of the exact same transcriptional pattern in response to three diatomic gases is unique in the bacteriology field and may represent a new paradigm. A few of many yet unanswered questions include: Why and how do three different gases trigger the exact same transcriptional response in Mtb? How can this knowledge provide mechanistic insight into the physiological role of DosS and DosT, what is the precise role of the Dos regulon in persistence, and does the Dos regulon plays a role in enabling the bacillus to enter, maintain and emerge from a persistent state?
Sophisticated biochemical and crystallography studies of DosS and DosT have made significant contributions towards understanding how these proteins sense NO, CO, and O2. Unfortunately, most of these in vitro studies frequently lacked biological context, and attempts to explain Mtb pathogenesis have to be interpreted with caution. In the section below, we will highlight several new findings that may provide a biological framework for future in vitro biochemical studies.
First, expression of the orphan sensor kinase dosT was shown to be constitutive during aerobic growth and during anaerobic dormancy (32) and contrasts with the highly hypoxia-, NO-, and CO-induced dosS. This is important as it appears that DosT plays an early role in response to hypoxia, which then induces the Dos regulon including DosS. As oxygen gets depleted, DosT becomes inactive, probably because of its weak affinity for O2 compared to DosS (78), and DosS maintains induction of the Dos regulon from that point forward (32). Second, a fascinating discovery was that vitamin C (ascorbate) induces the Dos regulon in the presence of oxygen and arrests bacterial growth (81). Further studies have shown that ascorbate induces the Dos regulon via DosS, but not DosT (31). These findings are important because ascorbate is a specific reductant of cytochrome C and therefore establishes a physiological link between DosS redox signaling and the electron transport chain (ETC). Accordingly, it was proposed that DosS responds to the redox status of the ETC. Microarray analysis of ascorbate-treated aerobic cultures of Mtb ΔdosS cells have shown that 35 of 49 differentially regulated genes were part of the Dos regulon (31). Third, exogenous addition of menaquinone to Mtb cells cultured under hypoxic conditions increased Dos regulon expression. The authors proposed that a reduced menaquinone pool induces the Dos regulon via DosS (31). These findings are important as the menaquinone pool is central to the ETC and again establish a mechanistic link between DosS signaling and the respiratory state of the cell.
The above findings highlight the role of a reduced ETC to induce the Dos regulon, and directly implicate DosS in sensing the redox state of the ETC. It is clear that we lack knowledge of the mechanisms Mtb use to maintain redox homeostasis during disease and persistence. This is particularly important as Mtb has the ability to survive extended periods (years) of anaerobiosis (88) and encounters multiple host gases (e.g., NO and CO) that inhibit respiration.
Is Mtb DosS an O2 or Redox Sensor?
It is generally agreed that the sensing mechanism of Mtb DosT involves binding of ligands (O2, NO, CO) to the heme; the controversy comes from conflicting results with DosS, specifically, the mechanism of its interactions with O2. Sardiwal et al. (70) first described the presence of heme in the DosS GAF A domain and the spectrum displayed properties consistent with oxyferrous coordination. In 2007, three groups independently reported the results of investigations into the mechanisms of interaction of the DosS heme group with ligands, most importantly, with O2. Yukl et al. (94) and Ioanaviciu et al. (35) found stable and reversible oxyferrous complexes with three different constructs of DosS. On the other hand, Kumar et al. (46) reported that isolated DosS is fully oxidized to the ferric state and addition of O2 to the reduced protein results in rapid oxidation to the ferriheme (as opposed to binding). On the basis of this result, Kumar et al. (46) proposed that DosS acts as a “redox sensor” rather than a direct sensor of O2 (Fig. 5). Sousa et al. (78) reported a slow but measurable rate of DosS autoxidation (t1/2 approx. 4 h). Consistent with Kumar et al. (46), Cho et al. (9) reported in 2009 that O2 rapidly oxidizes, rather than binds to, the ferroheme of the DosS GAF-A domain, while Ioanoviciu et al. (34) reported that full-length DosS is one of the most stable oxyheme proteins, in terms of its very slow autoxidation.
The origins of these divergent results are unclear; however, substantial insight has been obtained regarding mechanisms that contribute to these two divergent properties. In their studies with stable oxyferro complexes of DosS and various constructs, Yukl et al. (94) and Ioanoviciu et al. (34, 35) provided spectroscopic evidence that stabilization of the oxyferro complex is imparted by both interdomain interactions (perhaps helping to explain why some truncated proteins exhibit enhanced autoxidation rates) and also hydrogen bond interactions at the bound oxygen. In addition, studies with wild-type DosS and a mutant DosS with replacement of a distal Tyr residue with Phe suggest that phenolic hydrogen bonding from Tyr (along with interdomain interactions) imparts ligand discrimination properties (inactivation of histidine kinase activity by O2 but increased activation by CO and •NO). Recent studies by this group have also provided evidence that the oxy complex of DosS is capable of catalyzing NO dioxygenation, which results in conversion of ferrous heme to the inactive ferric state and concomitant formation of nitrate (95). In the presence of •NO, further transformations of the ferriheme occur, including relatively rapid reversible binding of •NO to the ferriheme (maintaining kinase inactivation) and also, upon reduction of the heme, formation of the very stable ferronitrosyl complex (resulting in kinase activation). The biological significance of these transformations is not clear, but they could result in a complex level of regulation depending on the balance between levels of O2 and •NO as well as rates of reduction of the ferriprotein.
On the other hand, Cho et al. reported oxidation rather than binding of O2 to the heme-containing GAF-A domain of DosS (8, 9). Examination of the crystal structures of the oxidized, reduced, reduced then air-oxidized, and cyano-met complexes suggest that the heme group is isolated with restricted ligand access. In addition, an extensive hydrogen bonding network at the heme site involving multiple water molecules and amino acid side chains suggests a potential route for electron flow to O2 from the heme (9). More recently, crystal structural determination coupled with spectral kinetic studies suggest that, while DosT has an open channel for relatively unrestricted small ligand access to the heme, access in DosS is restricted by the carboxylate group of Glu87 acting as a “gate”; directed mutation of Glu87 to Gly transforms the heme interaction of ferrous DosS with O2 from reduction to binding (8). Also, biological functional complementation measurements show that single changes in several amino acids in proximity to the heme in DosS (including Glu87) to the corresponding residue in DosT is sufficient to convert the response to hypoxia in M. smegmatis from that characterized by DosS to that characterized by DosT (42). This result is especially important because it may provide in vivo insights into the evolution of microbial signal transduction.
Mtb Redox Sensing and Its Possible Significance
At this point it is difficult to determine the relative validity of the Mtb DosS O2 binding vs. oxidation claims, but perhaps the crucial mechanistic question in this and related controversies is what exactly is the meaning of “redox sensing”? In fact, both proposed mechanisms will sense O2, the binding mechanism being determined by the binding affinity of O2 to ferroheme, and the redox mechanism being determined by the availability of O2 for heme oxidation; oxygen concentration will thus be a major determining factor for both. So how might the “redox sensing” mechanism respond differently to O2 concentration than the binding mechanism, and how might such a mechanism provide additional useful information to the organism regarding changes in host environment? Two crucial considerations for this (and indeed all biological response systems) are the relative and biologically relevant time scales for changes in input and resultant changes in response. With regard to input, the time scale for signal input for the binding mechanism is on the order of subseconds to seconds, determined by the rate constants for heme/O2 association and dissociation (34). It is likely that the response timescale is likewise on the order of subseconds, assuming the response involves modifications in protein structure which would presumably be immediately conveyed to the kinase domain. This information transmission can reasonably be assumed to be proportional to the relative amount of time the protein spends in the deoxy (active) compared to the oxy (inactive) state, which can be expected to change relatively rapidly in response to environmental changes in O2 concentration. For the “redox sensing” mechanism, the input signal is considerably more complex, both mechanistically and temporally. In this case, it is reasonable to assume that the major parameter determining signal input is proportional to the relative amount of time the protein spends in the reduced (active) compared to the oxidized (inactive) state. Conversion from the active to the inactive state is determined by both the rate of heme oxidation by O2 and by the rate of reduction of the ferriheme. The nature of this latter step, heme reduction, is most poorly defined and quite possibly the single most important parameter, as described below.
How might reduction of heme be accomplished and how might this process convey important information (in addition to O2 concentration)? The extent to which DosS will respond to a diversity of oxidative inputs will be determined by the extent to which the specific redox couple(s) that reduces oxidized DosS responds to this diversity of inputs. The two most obvious mechanisms to accomplish this are to have multiple reductants for DosS or to have the single reductant be a central intermediate in redox communication with multiple cellular redox couples. The latter mechanism would seem more specific and also more amenable to cellular regulation, and in fact there is evidence for such a mechanism. In vitro, met DosS has been shown to be reduced by an endogenous Mtb ferredoxin reductase/ferredoxin system (34). Based upon this data, it is tempting to speculate that during hypoxia, upregulation of fdxA (a member of the Dos regulon) participate in the reduction of met DosS to active ferrous DosS. Since DosS exists in the oxy state and extremely low autooxidation rates have been reported (34), the biochemical data is in agreement with a genetic study (32) demonstrating a role for DosS during the late stage (and DosT during the early stage) of in vitro dormancy where the O2 concentration is very low, leading to deoxygenation of DosS. It will be important to determine the Mtb growth phase and environmental O2 concentration where oxy DosS are oxidized to deoxy DosS and eventually met DosS using the Wayne mode for in vitro dormancy.
Another in vitro study has shown that met DosS can be reduced by reduced flavin (FMNH2) to active ferrous DosS (9). The biological significance of this latter result is not clear since flavin normally functions in the cell as a bound prosthetic group in oxidoreductases and does not function as a free electron-carrying cofactor. However, the authors pointed out that Mtb Rv2540c is a good candidate as it encodes a bifunctional chorismate synthase capable of accelerating NADH-dependent FMN reduction (14).
As described in the previous section, more relevant in vivo cellular results have been reported by Honaker et al. (31) where cellular DosS induces the Dos regulon in response to increased electron transfer in the presence of O2; results with inhibitors suggested that the cellular menaquinone pool plays a central role. Indeed, a potential connection between these results may be provided by the findings that ferredoxin in Mtb (fdxA) is a member of the DosR regulon core (44, 57, 77, 89), and although not previously described for mycobacteria, electron transfer between menaquinone and ferredoxin is documented in other anaerobic bacteria (33). Thus, it may be that the redox state of ferredoxin, a central mediator (and thus detector) of the ambient intracellular redox status, may transmit signals to DosS via reduction of the oxidized heme. Furthermore, DosS sensing of the electron transport chain may allow the pathogen to sense a wide spectrum of environmental cues such as alternative electron acceptors (e.g., nitrate), pH, or different carbon sources that would affect respiration. Under this scenario, such enzymatically controlled oxidation/reduction is a more integrative response than the immediate response of rapid O2 association/dissociation of DosT, providing desired relevant time scale responses to the organism's environmental challenges. These challenges are potentially more finely tuned over a variety of timescales (and to a diversity of oxidative challenges) than simply second-to-subsecond changes in the heme “gas” sensor DosT. Indeed, examination of the relative time scales of induction of DosT vs. DosS activation suggest that the former response is rapid, while the latter response is delayed and more sustained (6).
In sum, it is clear that, despite several thoughts, we lack detailed experimental evidence of how inactive met DosS is converted into the active ferrous DosS state to activate the Dos regulon. Since two studies have shown that ascorbate can induce the Dos regulon (31, 81), the provocative question is raised whether ascorbate can directly reduce met DosS to ferrous DosS (Fig. 5)? Although this has not yet been investigated for DosS or DosT, earlier studies have shown that ascorbate is capable of reducing ferryl and met myoglobin (19), and can function as an antioxidant and prooxidant (27). Future studies are eagerly awaited.
Conclusions and Innovation
Virtually all bacterial pathogens require iron and heme, and will either synthesize their own heme and/or acquire it from the host. Not surprisingly, disruption of genes involved in heme acquisition typically results in attenuation. A fundamentally important challenge is to understand how NO, CO, or O2 gases bind to or compete with heme iron to generate Fe-NO, Fe-CO, and a Fe-O2 bonds, and how the oxidation of heme iron by O2 serves as a sensing mechanism for bacteria. The elucidation of these mechanisms will almost certainly depend on the effective integration of biochemical data with physiologically relevant experimental settings.
Mtb, the etiological agent that causes TB, is a particularly good example of a global pathogen that has evolved a sophisticated sensory system capable of sensing or reacting with mixtures of host gases such as NO, CO, and O2 via its heme sensor kinases, DosS and DosT. NO, O2, and CO are physiologically relevant gases associated with the establishment and maintenance of latent tuberculosis (44, 46, 77, 89). The ability of Mtb to induce the exact same transcriptional pattern in response to the three diatomic gases NO, CO, and O2 is unparalleled in the bacteriology field and will generate many new research opportunities. This, together with the significant interest that O2 tension has received in generating persistent Mtb in vitro, strongly points toward these gases as highly relevant host ligands that may shape disease progression. However, simple in vitro characterization of the interactions of DosS and DosT with NO, CO, and O2 may not always be physiologically relevant and must be interpreted with caution. For instance, on one hand, “optimal levels” of NO, CO, and O2 could either maintain Mtb in a dormant state (or inhibit respiration). On the other hand, because of strong anti-apoptotic and anti-inflammatory properties, CO together with excessive amounts of NO could cause destructive host immunopathology to exacerbate disease.
The divergent results from Mtb DosS studies by four independent groups add to the complexity of these heme sensor kinases, and further studies on the role of DosS as a redox sensor, or sensor of O2 (or both) is eagerly awaited. Also, an important question that emerged from these studies is how met DosS is reduced to the active ferrous DosS state to induce the Dos regulon? It is evident that DosS and DosT are key regulators of the adaptive responses to fluctuating NO, CO, and O2 levels in vivo. However, much remains to be learned about the role of these gases in Mtb disease and that more multidisciplinary in vitro and in vivo approaches are needed to better understand the mechanisms of pathogenesis.
Several other important questions regarding the function of heme sensor proteins remain to be answered. To date, knowledge of the intracellular redox environment of many microbes, the identity of main redox couples and buffers, and the behavior of these redox couples under different environmental conditions remain sparse. This is in part due to a lack of appropriate tools to measure the ambient intracellular redox state of the microbial cell noninvasively. Also, a remaining understudied issue is the identification of redox couples capable of reducing the oxidized heme iron of redox sensors. Nonetheless, despite our incomplete understanding of these events, much progress has been made in establishing model heme proteins such as EcDos, FixL, CooA, and more recently DosS and DosT, which forms a solid foundation for the examination of new heme proteins that react with NO, CO, and O2. It is likely that additional heme proteins with new functional properties will be identified in the coming years.
In conclusion, a challenge faced by heme investigators is the design of innovative in vitro and in vivo experimental approaches that upon integration will yield new experimental tools and concepts, and shed light on the true physiologically function of heme-based sensor proteins.
Footnotes
Acknowledgments
We thank Joel N. Glasgow for critical editing of this manuscript. Research in the laboratory is supported in whole or in part by National Institutes of Health Grants AI058131, AI076389 (to AJCS) and CA131653 (to JRL). This work is also supported by the University of Alabama at Birmingham (UAB) Center for AIDS Research, and UAB Center for Free Radical Biology (AJCS). AJCS is a Burroughs Welcome Investigator in the Pathogenesis of Infectious Diseases.
Author Disclosure Statement
No competing financial interests exist.