
Abstract
Introduction
Innovation
This study provides new insights into the understanding of the mechanisms of action of P144, a peptide that blocks transforming growth factor‐β1 (TGF‐β1) actions. In experimental hypertension, NADPH oxidase‐dependent oxidative stress participates in the pathophysiology of renal fibrosis. Since P144 blocks TGF‐β1 activation of NADPH oxidases and fibrosis, our data identify P144 as a desirable therapeutic agent to reverse renal fibrosis and oxidative stress in hypertension. Despite the fact that the usage of peptides as “druggable” therapeutics constitutes a current challenge, these molecules are likely to be useful tools for attacking protein‐protein interactions, and novel technologies (i.e., nanotechnology) may help solve this limitation.
Hypertension is associated with multiple functional and structural cardiovascular and renal alterations (29). Kidney malfunction causes hypertension, which, in turn, aggravates chronic kidney disease, accelerating its progression (35). Transforming growth factor-β1 (TGF-β1) acts as a key fibrogenic cytokine in many tissues, and its elevated circulating levels have been related to the progression of kidney disease (1). Currently, it is also believed that ROS can be related to actions mediated by this cytokine. In fact, in vitro studies have demonstrated that activation of NADPH oxidase has a role in the TGF-β1-induced effects (8, 28, 30, 41). Moreover, the association of NADPH oxidase with TGF-β1-induced fibrosis has also been observed in different experimental models (31, 32).
The peptide P144 encompasses amino acids 730–743 from the membrane-proximal ligand-binding domain of TGF-β1 type III receptor, also called betaglycan, and is able to block TGF-β1-induced effects in a hepatic failure rat model, as well as in a murine model of scleroderma (14, 36). Furthermore, P144 prevents myocardial fibrosis in experimental hypertension (17). Whether P144 also possesses effects in renal tissue in arterial hypertension and the potential mechanism implicated remains unknown. Thus, we investigated whether a chronic treatment with P144 might inhibit renal NADPH oxidase-mediated oxidative stress. We also studied whether this effect may be associated with its potential renal antifibrotic properties.
Results
Blood pressure values and renal parameters in untreated 10-week-old rats
Untreated 10-week-old Wistar-Kyoto (WKY, n=10) rats and 10-week-old spontaneously hypertensive rats (SHR, n=10) were studied to assess baseline values of the parameters tested in 22-week-old rats in order to determine whether P144 reverses or prevents the development of abnormalities. At 10 weeks, SHR exhibited higher values of systolic blood pressure (SBP), and renal ·O2
− production, p47phox protein expression, and levels of nitrotyrosine (NT) than WKY (Supplementary Table S1; Supplementary Data are available online at
Effects of P144 on blood pressure
SHR is a commonly used model of human essential hypertension and target organ damage. The SBP was measured in order to know the potential antihypertensive effect of P144. At 22 weeks, values of SBP were elevated (p<0.01) in vehicle-treated SHR (V-SHR, 246.7±3.6 mmHg) and P144-SHR (215.6±5.9 mmHg) compared with vehicle-treated WKY (V-WKY, 175.0±3.5 mmHg) and P144-WKY (160.8±5.2 mmHg). In addition, the SBP was higher at 22 weeks than at 10 weeks in both groups (V-SHR: 190.4±7.5 mmHg; P144-SHR: 191.7±4.8 mmHg), which means that the P144-SHR group remained hypertensive during the experimental period. No significant effects of P144 on SBP were observed in P144-treated WKY (P144-WKY, 160.8±5.2 mmHg) compared with V-WKY.
Effects of P144 on renal NADPH oxidase activity and H2O2 production
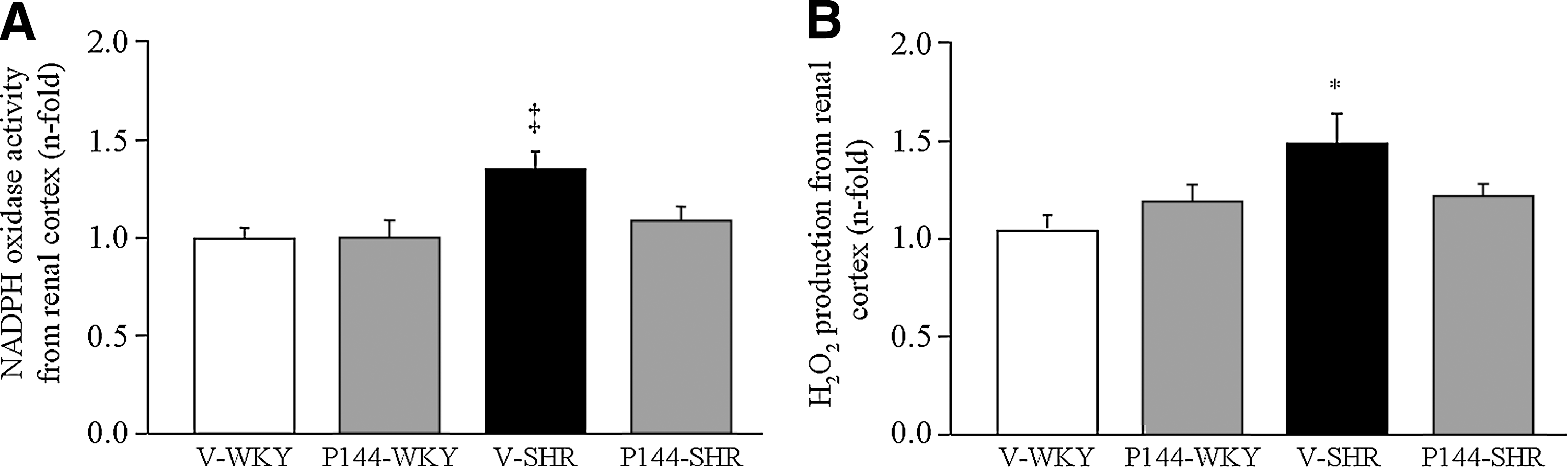
NADPH oxidases play a major role in the increase of oxidative stress in the kidney of SHR (7), and generation of ROS is the best-known function of these enzymes. Renal NADPH oxidase activity (Fig. 1A) and H2O2 production (Fig. 1B) were significantly increased in V-SHR as compared with V-WKY. The administration of P144 reverted the increase in NADPH oxidase activity and prevented the increase in H2O2, as no differences were found in these parameters among P144-SHR, P144-WKY, and V-WKY (Fig. 1).
Effects of P144 on renal NADPH oxidase subunits expression
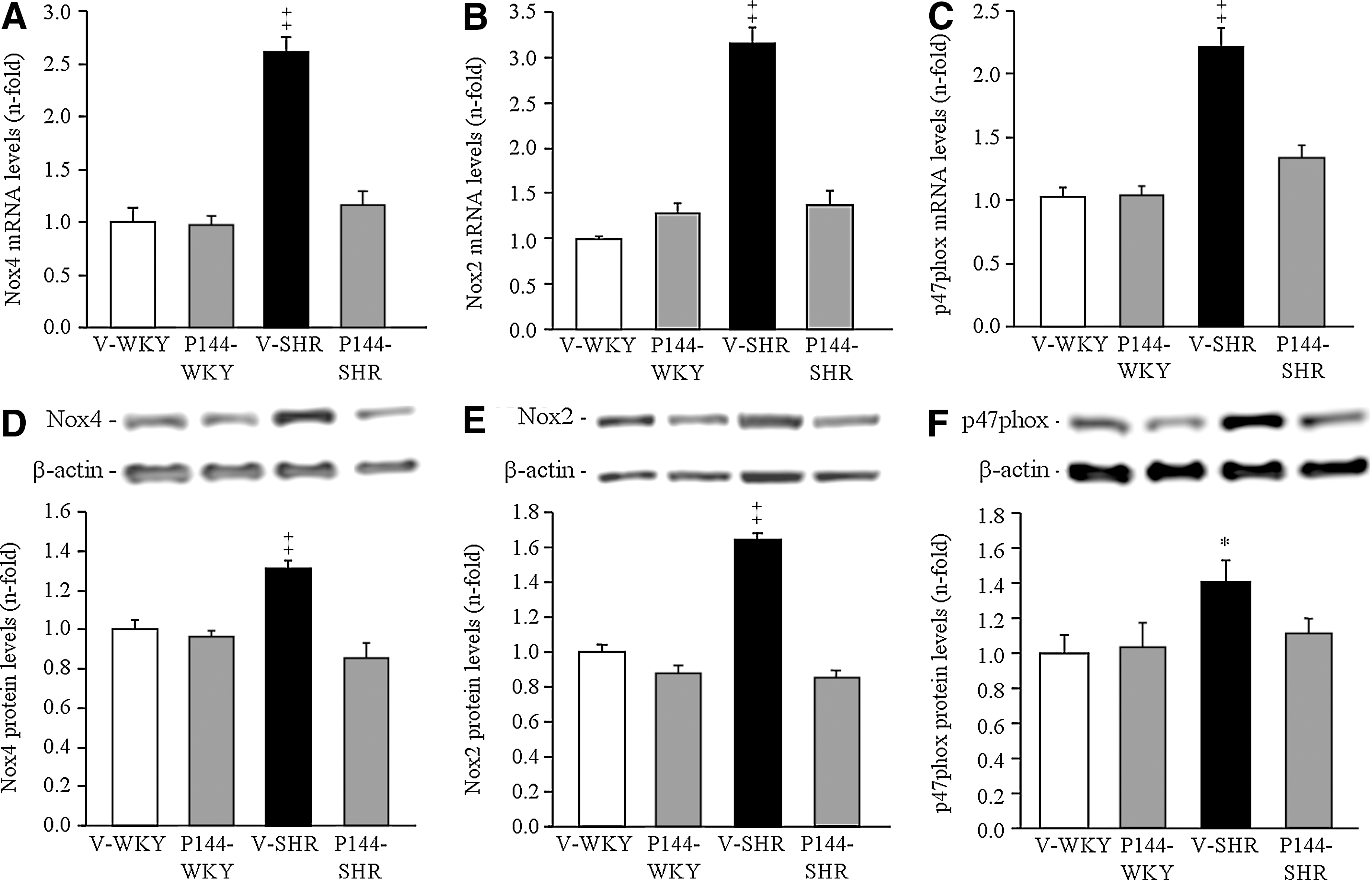
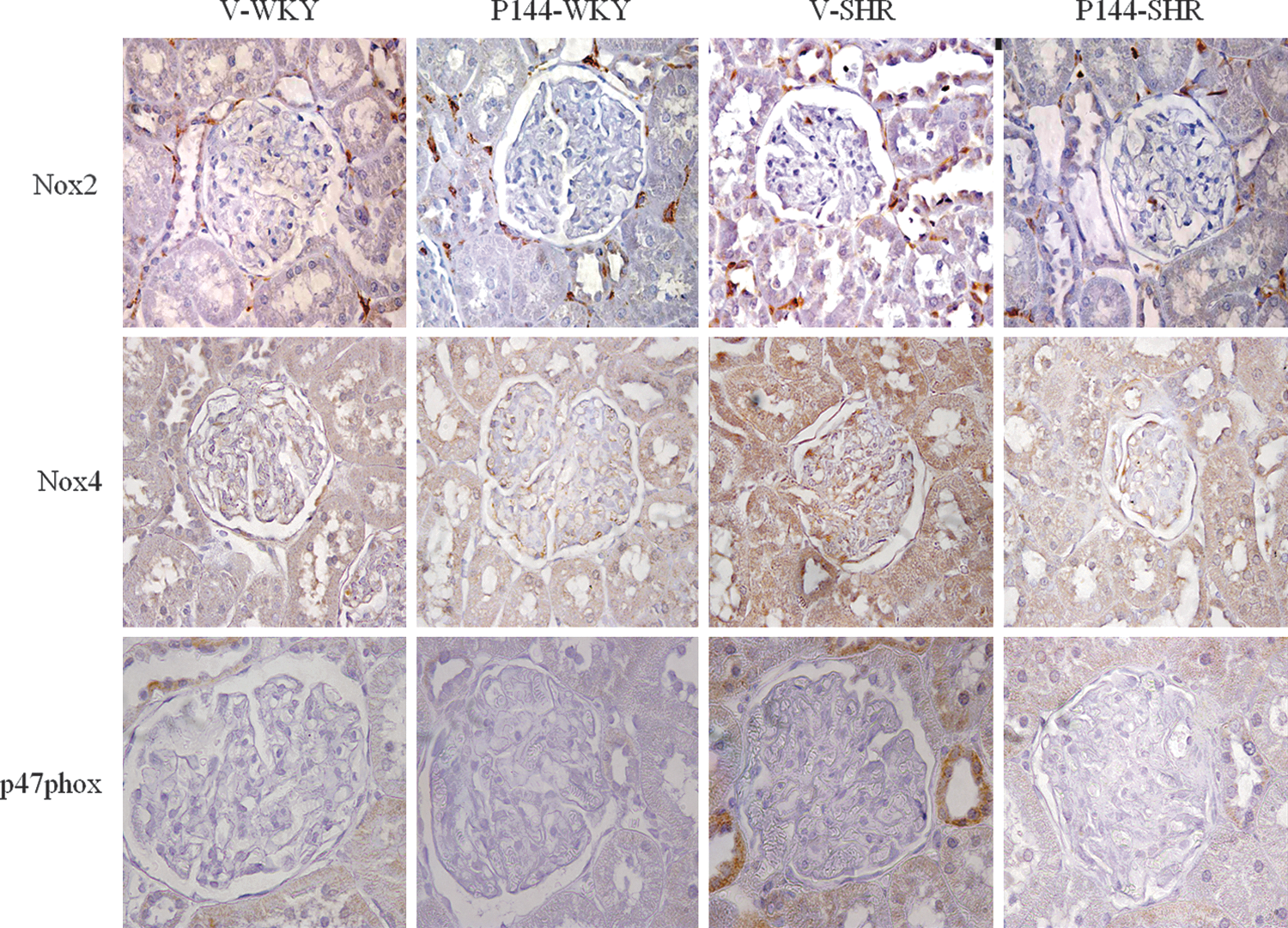
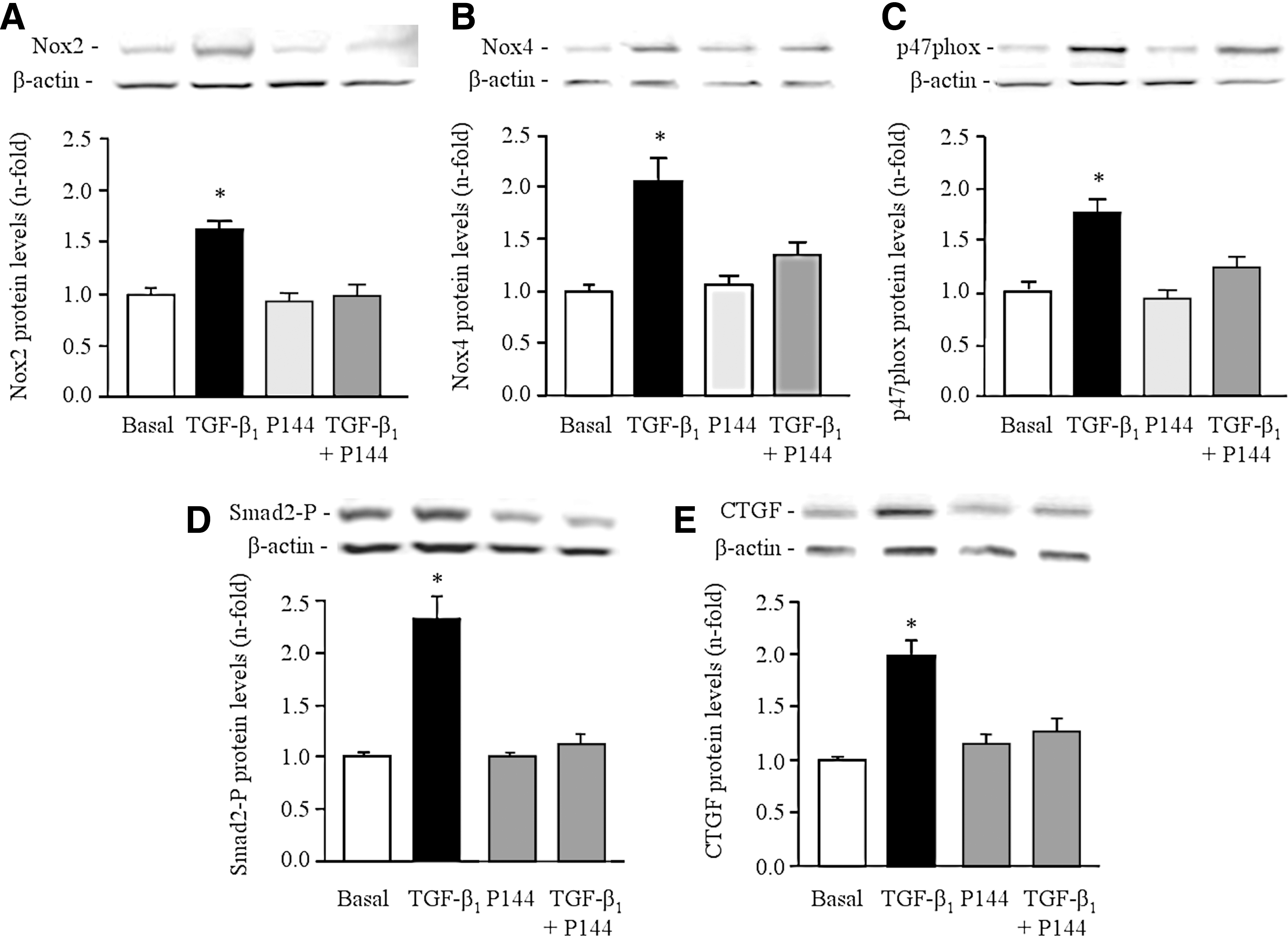
To study the participation of the NADPH oxidase system in the pathogenesis of renal oxidative stress, the expression of Nox4, Nox2, and p47phox was investigated. Real-time PCR showed a significantly greater mRNA abundance for Nox4 (Fig. 2A), Nox2 (Fig. 2B) and p47phox (Fig. 2C) in the renal cortex of V-SHR than in V-WKY. Similarly, protein expression of Nox4 (Fig. 2D), Nox2 (Fig. 2E), and p47phox (Fig. 2F) was significantly higher in V-SHR than in V-WKY. The chronic administration of P144 reduced the mRNA and protein expression of these subunits in P144-SHR (Fig. 2). The immunolabeling of Nox2, Nox4, and p47phox was stronger in V-SHR than in V-WKY (Fig. 3). Weaker immunolabeling could be detected in P144-SHR compared with V-SHR. The presence of Nox2 and p47phox was mainly localized in the tubules, although some weak signal could occasionally be detected in the glomeruli (Fig. 3). On the other hand, Nox4 immunostaining was strongly observed in distal convoluted tubules and in glomeruli (Fig. 3).
Effects of P144 on renal levels of NT
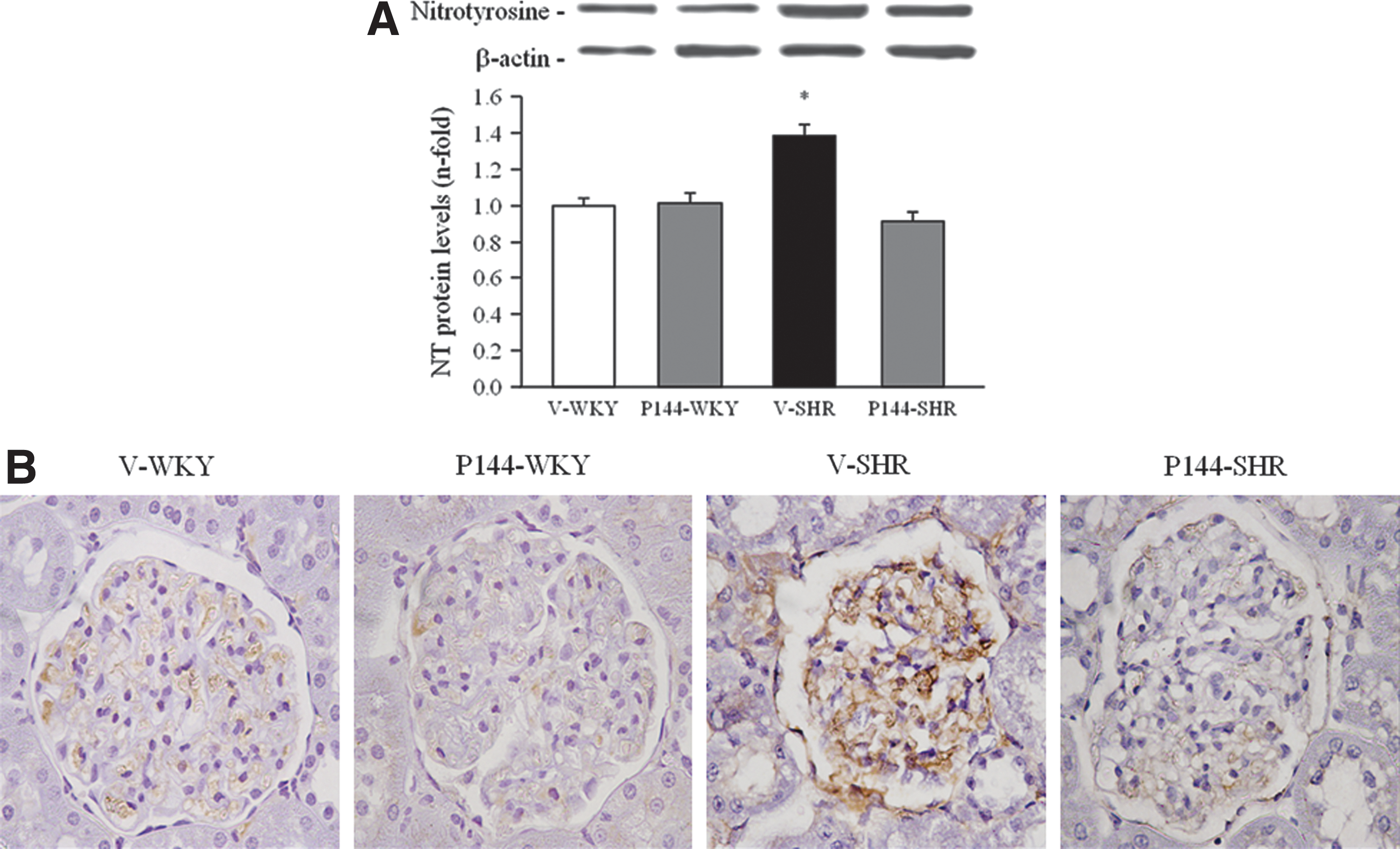
Oxidative stress has been described as one important factor involved in the progression of renal injury in SHR, and the level of NT is considered a marker of in vivo generation of peroxynitrite. Renal levels of NT were higher (p<0.05) in V-SHR than in V-WKY (Fig. 4A). The administration of P144 significantly reversed the levels of NT in P144-SHR. The V-SHR group exhibited the strongest NT immunolabeling compared with V-WKY mainly localized in the glomeruli (Fig. 4B). Finally, treatment with P144 reverted this effect, as a noticeable decrease in the intensity of NT staining is observed in the glomeruli of SHR-treated rats (Fig. 4B). These results were confirmed by NT detection by Dot blot (data not shown).
Effects of P144 on redox status
Levels of thiobarbituric acid-reactive substances (TBARS) and total antioxidant capacity (TAC) were determined to evaluate the effect of P144 on redox status. As illustrated in Supplementary Figure S1, TBARS content was markedly increased in V-SHR and was decreased by chronic treatment with P144 (Supplementary Fig. S1). TAC was significantly decreased in V-SHR compared with V-WKY (Supplementary Fig. S1). Treatment with P144 increased TAC in P144-SHR to the levels of V-WKY.
Superoxide dismutase (SOD) is an enzyme that acts as a first line of defence against oxygen free radical-mediated damage by catalyzing the dismutation of ·O2 − to H2O2. Our results indicate that SOD activity was significantly decreased in V-SHR compared to V-WKY and that P144 treatment increased it up to the levels of V-WKY (Supplementary Fig. S1).
Effects of P144 on NO production
The expression of eNOS may be indicative of NO production, and its reduction is a potential mechanism involved in the development of hypertension in SHR. In fact, mRNA and protein levels of eNOS were significantly reduced in V-SHR compared with the V-WKY group (Supplementary Fig. S2). In this line, the levels of eNOS phosphorylation and the NO production were also significantly lower in V-SHR than in V-WKY (Supplementary Fig. S2). With regard to the effect of chronic treatment with P144, it did not modify the expression of renal eNOS (mRNA and protein) in any of the studied groups while it significantly induced eNOS phosphorylation and increased NO production in P144-SHR up to the levels of V-WKY (Supplementary Fig. S2).
Effects of P144 on renal TGF-β1, receptor subtypes of TGF-β1, and extracellular matrix proteins expression
Renal fibrosis reflects the balance between collagen deposition and degradation, and several proteins are implicated in this equilibrium (25). The mRNA levels of TGF-β1, were significantly higher in the kidneys from V-SHR than in those from V-WKY (Fig. 5). Accordingly, the mRNA levels of structural molecules such as fibronectin and type I collagen were significantly increased in V-SHR compared with the V-WKY group (Fig. 5). In addition, the enzymes that catalyze different steps in the collagen synthesis and maturation, biglycan, BMP1, and LOX were significantly higher in the kidneys from V-SHR than in V-WKY (Fig. 5). The chronic administration of P144 was accompanied by a significant decrease in the levels of these mRNAs in P144-SHR compared with V-SHR. No significant differences were observed among V-WKY, P144-SHR, and P144-WKY (Fig. 5). Our findings also showed that the ratio between MMP-1/TIMP-1 expression was significantly decreased in V-SHR (0.45±0.06 AU) compared with V-WKY (1.05±0.27 AU). Chronic treatment with P144 did not modify the ratio between MMP-1/TIMP-1 expression in either WKY (P144-WKY: 0.99±0.31 AU) or SHR (P144-SHR: 0.37±0.11 AU) compared with vehicle.
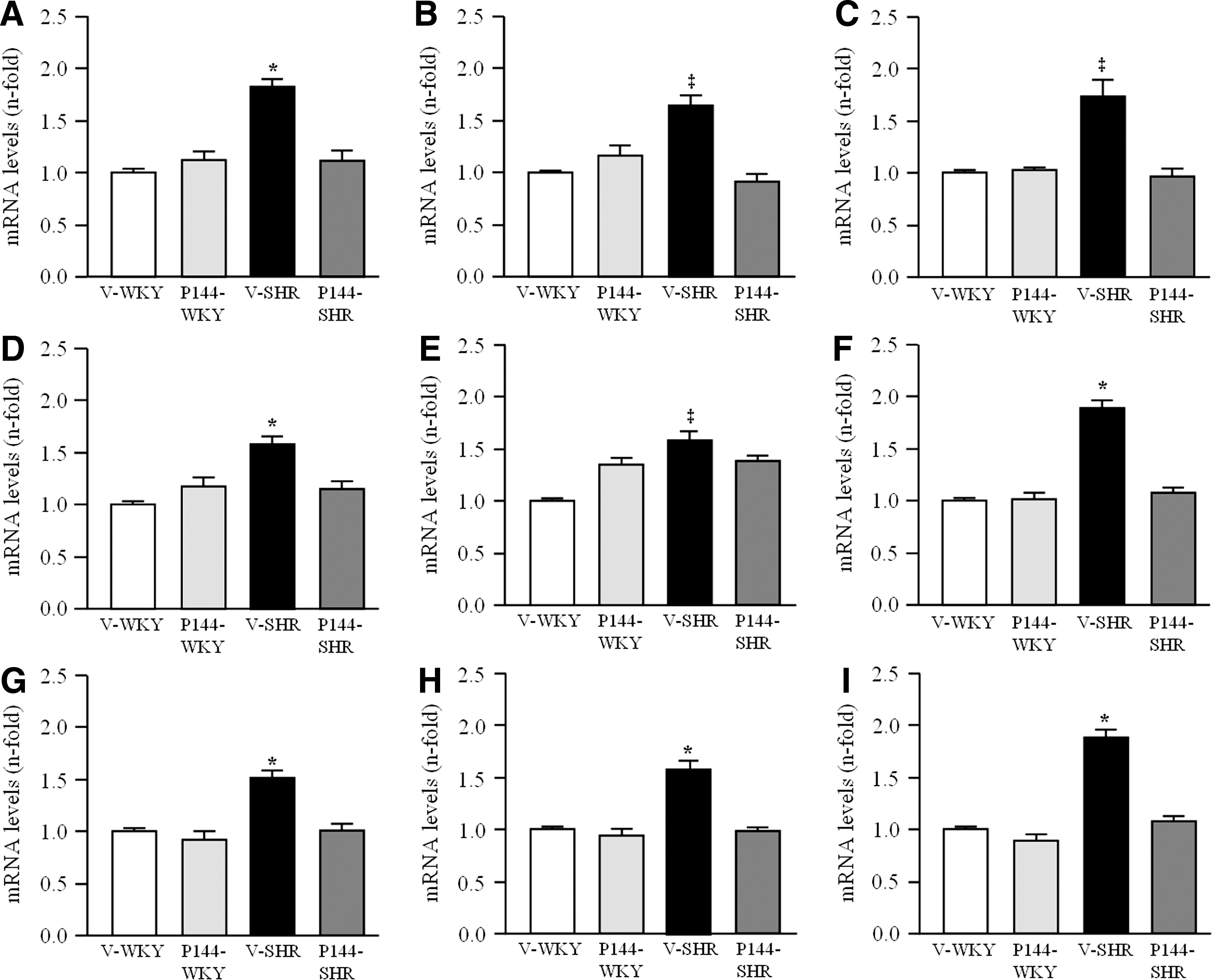
Basal expression of type I, II, and III receptors of TGF-β1 was analyzed in order to assess their potential involvement in the effects of P144 on TGF-β1-mediated actions. The expression of the three receptors subtypes of TGF-β1 was higher in V-SHR than in V-WKY (Fig. 5). Chronic treatment with P144 significantly reduced the expression of receptors I, II, and III of TGF-β1 (Fig. 5) in P144-SHR compared with V-SHR.
Effects of P144 on renal CTGF expression
CTGF is an important downstream mediator of TGF-β1 and is associated with the renal fibrosis described in experimental hypertension (21). Renal CTGF mRNA levels (Fig. 6A) and protein expression (Fig. 6B) were increased (p<0.05) in V-SHR compared with the V-WKY group. Likewise, V-SHR showed stronger immunostaining for CTGF in glomeruli and tubuli than V-WKY (Fig. 6C). P144 treatment diminished (p<0.05) CTGF mRNA and protein expression, as well as the immunostaining intensity in P144-SHR compared with V-SHR.

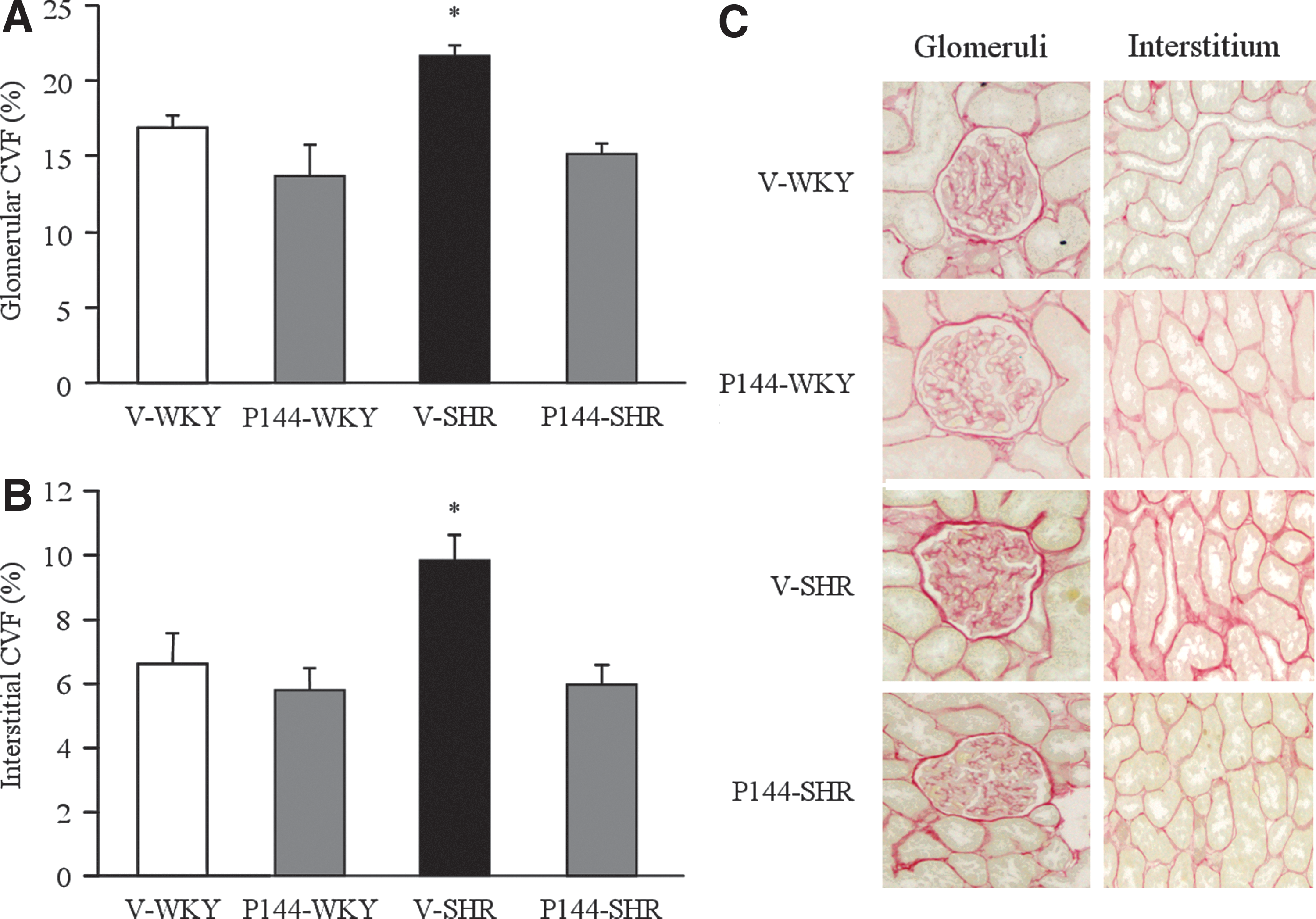
Effects of P144 on renal histomorphological alterations
Histological evaluation was performed to determine the degree of renal fibrosis in SHR as well as the organ-protective effects of P144. Quantification of CVF in renal tissues by computer-assisted analysis revealed a larger area occupied by both glomerular (Fig. 7A, C) and interstitial (Fig. 7B, C) fibrosis in V-SHR compared with V-WKY. Chronic treatment with P144 significantly reduced extension of the glomerular and interstitial fibrotic area in P144-SHR compared with V-SHR, whereas no significant effect was observed in P144-WKY compared with V-WKY rats. Furthermore, the fibrotic levels detected in P144-SHR normalized to similar levels of those found in V-WKY animals.
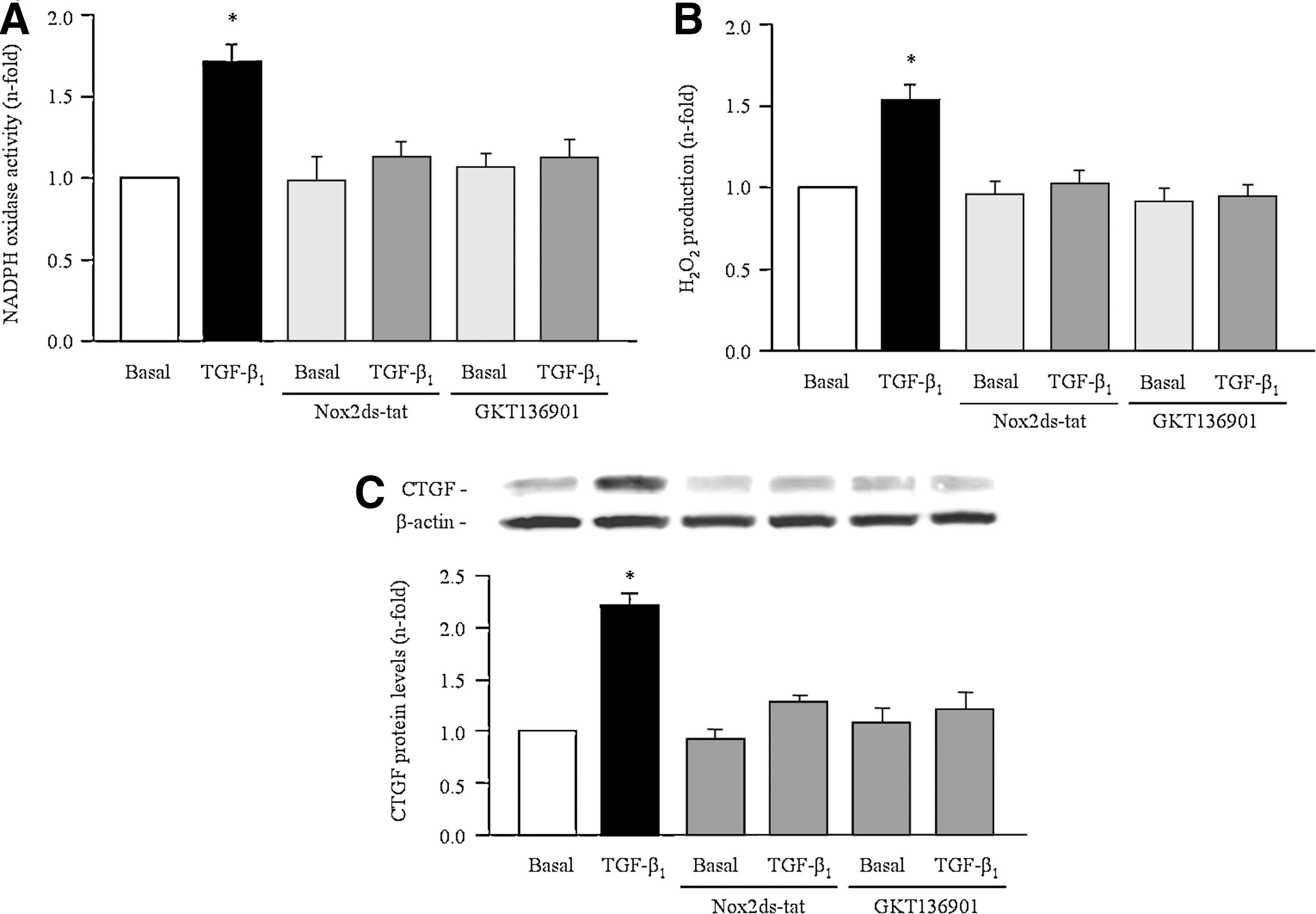
Role of NADPH oxidase on TGF-β1-induced effects in cultured renal cells
To better characterize the role of NADPH oxidases in TGF-β1-induced ROS production and fibrosis, experiments with specific inhibitors for Nox2, Nox2ds-tat, and Nox1/Nox4, GKT136901, were performed. In NRK52E, preincubation with GKT136901 (10 μM) and Nox2ds-tat (50 μM) induced a significant decrease on TGF-β1-induced NADPH oxidase activity (Fig. 8A), H2O2 production (Fig. 8B), as well as on the TGF-β1-induced CTGF expression (Fig. 8C).
Effects of P144 in cultured renal cells
To study the direct implication of P144 in TGF-β1-induced actions, NRK52E cells were stimulated with TGF-β1 and then, NADPH oxidase-dependent ROS production and expression of downstream effectors of TGF-β1, such as CTGF, were determined. These in vitro studies showed that TGF-β1 stimulated (p<0.05) NADPH oxidase-dependent ·O2 − and H2O2 production in NRK52E cells as well as the expression of NT (Fig. 9). These parameters of oxidative stress were significantly inhibited on preincubation with P144 (Fig. 9). As expected, TGF-β1-induced protein expression of Nox4, Nox2, and p47phox was also significantly inhibited by the preincubation with P144 (Fig. 10). Consistently, levels of Smad2-P, a TGF-β1-dependent signaling pathway molecule, and CTGF expression, a growth factor stimulated by TGF-β1 that promotes fibrosis, were also increased (Fig. 10) in NRK52E cells by stimulation with TGF-β1. After the preincubation with P144, the expression of both parameters decreased significantly (Fig. 10).

Discussion
The main findings of this study are as follows: (i) chronic treatment with P144 reverses NADPH oxidase-dependent ROS production, NADPH oxidase subunits expression, and associated oxidative stress in the kidney of SHR; (ii) chronic treatment with P144 reverses renal CTGF up-regulation, the main mediator of TGF-β1; (iii) chronic treatment with P144 reduces fibrosis in the renal cortex of SHR; and (iv) P144 is able to block TGF-β1-stimulated NADPH oxidase-dependent ROS production and subunits expression as well as TGF-β1-dependent intracellular pathway in NRK52E cells. Collectively, our findings suggest that the inhibition of the NADPH oxidase-mediated oxidative stress is associated with the ability of P144 to block TGF-β1-induced fibrosis in the kidney of hypertensive rats.
TGF-β1 type III receptor or betaglycan is the most abundant TGF-β1 binding protein at the cell surface (27). Betaglycan potentiates TGF-β1 binding to the signaling type I and type II receptors and is involved in ligand presentation (26). Different experimental approaches demonstrated that the specific blockade of TGF-β1 prevented renal damage (22, 40). P144, a synthetic peptide from the sequence of the extracellular region of betaglycan, with potential binding activity to TGF-β1, has been developed (12). The antifibrotic effect of P144 has been previously described in several organs, where the activation of this cytokine presents a pathophysiological role (14, 17, 36). Nevertheless, the mechanisms associated to the effects of P144 remain to be elucidated in the pro-fibrotic mechanism of TGF-β1 in the kidney.
Available data suggest the contribution of oxidative stress in the development of renal damage in SHR (19, 42). Among the diverse sources of ROS, accumulating evidence indicates that NADPH oxidase activity is increased in different animal models of renal injury (37, 45, 47). Accordingly, we have shown here that the oxidative stress (increased TBARS and renal NT levels and reduced TAC and renal SOD) found in untreated V-SHR was associated with an increase in NADPH oxidase activity and expression. Chronic treatment with P144 normalized all parameters to the levels of V-WKY, suggesting that the modification of redox balance might be one of the consequences of the interference of P144 on TGF-β1-induced signaling pathways and effects. Interestingly, our findings showing that the treatment with P144 in hypertensive rats promoted the phosphorylation of eNOS and increased the levels of NO metabolites (NOx), supporting another mechanism responsible for the protective effect of P144 on renal fibrosis in hypertension.
Several in vitro studies have demonstrated that oxidative stress is downstream of the actions of TGF-β1 (8, 28, 30, 41). Accordingly, the specific blockade of diverse NADPH oxidase subunits reinforces the evidence that TGF-β1 stimulates ROS production and CTGF expression through these homologs, supporting the connection between TGF-β1 and fibrosis through the activation of NADPH oxidases. Furthermore, diverse experimental approaches have suggested that an up-regulation of TGF-β1 expression might be associated with the activation of the NADPH oxidases. First, a study in apolipoprotein E-deficient mice has demonstrated that elevated systemic TGF-β1 causes vascular alterations through NADPH oxidase activation (6). Second, treatment of Dahl salt-sensitive rats with atorvastatin reduced renal TGF-β1 and inhibited NADPH oxidase activity (47). In the same model, the administration of kallikrein improved renal function and reduced TGF-β1 expression, which was associated with a decrease in NADPH oxidase activity (45). In agreement with these studies, we have observed that chronic blockade of TGF-β1 actions by P144 reversed NADPH oxidase overactivity observed in V-SHR. Accordingly, treatment with P144 was associated with a significant reduction in the expression of NADPH oxidase subunits and NT in the renal cortex of P144-SHR. In vitro findings from the current study extended these data by showing that P144 inhibits the ability of TGF-β1 to stimulate NADPH oxidase-dependent oxidative stress in epithelial cells of the rat kidney. These experimental findings allow us to suggest that the effect of P144 may be due to the blockade of TGF-β1 pathways, independently of the hemodynamic factor. Thus, P144 treatment appears to limit oxidative stress by lowering NADPH oxidase abundance in the kidney. Although multiple studies support the detrimental role of Nox2 up-regulation in vascular and renal damage (3), some recent publications have reported a possible protective role of Nox4 in the vascular system (34, 38), whereas another study (2) shows that Nox4 does not promote renal disease development but may rather have a small, limited protective effect. Therefore, further studies with more specific Nox4 inhibitors would be necessary to better analyze the potential renal effects of Nox4. In fact, Cucoranu et al. have recently demonstrated that Nox4 silencing prevents TGF-β1-mediated fibrosis in human cardiac fibroblasts (8).
Our findings demonstrate a higher content of collagen in the renal cortex of V-SHR than in V-WKY rats. Accordingly, the higher collagen deposition found in the kidneys of V-SHR is also associated with the up-regulation of different genes that are traditionally participating in the fibrotic process, including fibronectin, BMP1, biglycan, LOX, and collagen type I. Furthermore, the levels of CTGF, a growth factor that is related with the expression of many of these fibrotic molecules (20, 40), were also increased in the tubular and glomerular areas of V-SHR. In addition, several studies support the notion that TGF-β1 constitutes a key molecule in the renal fibrosis associated with the chronic progressive renal disease (1, 48). In fact, increased levels of TGF-β1 in SHR have been reported (21, 39). In agreement with this, we found that TGF-β1 was up-regulated in V-SHR. On the other hand, the fibrogenic potential of TGF-β1 is also due to its ability to inhibit collagen degradation (44). Nevertheless, chronic treatment with P144 did not modify the ability of TGF-β1 to inhibit collagen degradation in SHR in the conditions analyzed in the present study. Thus, our current findings demonstrate, for the first time, that chronic treatment with P144 prevents renal fibrosis in SHR by interfering mainly in the TGF-β1-dependent signaling pathway and collagen synthesis in the kidney of SHR.
Several studies have shown that the reduction of NADPH oxidase activation is associated with renal protection in different animal models of chronic renal disease (31 –33). The present study shows that oxidative stress is already increased in young hypertensive rats with normal renal structure (CVF). Our data in treated hypertensive rats allow us to suggest that the chronic treatment with P144 reversed renal oxidative stress and prevented the development of fibrosis in line with other studies, showing that antioxidant therapy (e.g., apocynin, tempol, or N-acetylcysteine) reduced renal injury associated with a reduction of oxidative stress (4, 18, 46). These studies allow us to suggest that the effect of P144 may be, in part, due to the modification of redox balance in the renal cortex of SHR as a consequence of the interference of P144 on TGF-β1-induced signaling pathways and effects.
Chronic treatment with P144 moderately reduced but does not normalize SBP in P144-SHR; thus, this group of rats remained hypertensive through the experimental period. Since the direct target of P144 is TGF-β1, the potential role of this cytokine in the pathogenesis of hypertension needs to be further analyzed. TGF-β1 levels are increased in experimental hypertension (21, 39). Moreover, chronic treatment with an anti-TGF-β1 antibody has been associated with a reduction in blood pressure in 9-weeks old male Dahl salt-sensitive rats (9). In addition, systemic overexpression of TGF-β1 was associated with increased blood pressure and endothelial dysfunction in an atherosclerotic surrounding in apoE−/− (6). On the other hand, it seems that the contribution of TGF-β1 to the development of target organ damage in essential hypertension was independent of blood pressure values (10, 24). We can speculate that P144 may be exerting its protective effects on the kidney by preventing TGF-β1-mediated profibrotic and pro-oxidant effects through a mechanism that does not involve a hypotensive action. This is in agreement with other studies demonstrating that the beneficial cardiovascular effects of diverse antihypertensive drugs (e.g., calcium channel blockers, angiotensin II type 1 receptor blockers, or renin inhibitors) are due to their specific effects on particular signaling pathways independent of their blood pressure lowering effects (15, 23, 43). In addition, results from the present study showed that chronic treatment with P144 normalized levels of TGF-β1 and renal parameters of extracellular matrix metabolism despite a moderate reduction of blood pressure values.
In conclusion, the present results support that P144, which blocks the TGF-β1 pathway, exerts a protective effect against renal fibrosis in experimental hypertension. The mechanism involved seems to be associated with a decrease in NADPH oxidase-dependent oxidative stress in kidneys. Our data identify P144 as a desirable therapeutic agent for the prevention of renal damage in hypertension by attenuating fibrosis and oxidative stress, among other effects. The use of this new agent in combination with current antihypertensive drugs could help protect target organs in arterial hypertension. Further studies should be performed to better address this possibility.
Materials and Methods
Animal model
One group of untreated WKY (n=10) and one group of untreated SHR (n=10) were sacrificed at 10 weeks of age to determine the baseline values of the different parameters tested. Ten-week-old male WKY (n=10, V-WKY) and 10-week-old male SHR (n=10, V-SHR) received the vehicle solution (saline) intraperitoneally and were then sacrificed after 12 weeks at the age of 22 weeks. In addition, 10-week-old WKY (n=10, P144-treated WKY or P144-WKY) and 10-week-old SHR (n=10, P144-treated SHR or P144-SHR) were treated with intraperitoneal P144 (1 mg/kg body weight per day) and then sacrificed after 12 weeks. Kidneys were carefully dissected and processed for the different studies.
In vitro studies
In vitro experiments were performed in a rat proximal tubular epithelial cell line NRK52E, purchased from American Type Culture Collection. Cells were incubated in reduced serum media containing TGF-β1 (10 ng/ml) in the absence or presence of P144 (200 μg/ml), for 24 h before harvesting. After the treatments, cells were washed in phosphate-buffered solution (Invitrogen), centrifuged at 1100 g for 5 min at 4°C, and homogenized for protein extraction.
Measurement of blood pressure
SBP was measured in all rats every 2 weeks by the standard tail-cuff method using an LE5007 Pressure Computer (Letica Scientific Instruments).
NADPH oxidase-dependent ROS production
NADPH oxidase activity was measured as an index of ·O2 − generation by the lucigenin (5 μM)-enhanced chemiluminescence method in 25 μg of renal tissue protein and 5 μg of cell homogenates. The effect of several inhibitors, including 5 μM diphenylene iodonium (DPI), a flavoprotein inhibitor, 1 μM oxypurinol, a xanthine oxidase inhibitor, and 1 μM rotenone, a mitochrondial electron transport inhibitor were evaluated. DPI completely prevented ·O2 − production from renal cortex and NRK52E cells homogenates, whereas rotenone or oxypurinol were ineffective, thus identifying NADPH oxidase as an important enzymatic source of ·O2 − production (Supplementary Fig. S3). H2O2 production was determined in 25 μg of tissue protein and in cell culture medium using the Amplex Red hydrogen Peroxide/Peroxide Assay Kit (A22188; Molecular Probes) according to the manufacturer's instructions.
Real-time PCR
Total RNA was extracted from the renal cortex automatically by the ABI PRISM® 6100 Nucleic PrepStation (Applied Biosystems) method. 500 nanograms of total RNA were taken to perform a reverse transcription using the RNA SuperScript® VILO™ cDNA Synthesis Kit (Invitrogen). A real-time PCR was performed using a TaqMan® Gene Expression Assay (Applied Biosystems) and Quantitec SYBR Green PCR Kit (Quiagen).
Western blot
Proteins were separated by electrophoresis and incubated with specific primary antibodies. Protein expression was visualized with the Enhanced Luminol-based Chemiluminescence-Advanced chemiluminescence system (Amersham Biosciences). Membranes were analyzed using the Chemidoc Detection System and analyzed by Quantity One software (Bio-Rad) obtaining densitometric arbitrary units.
Immunohistochemistry
Kidney paraffin sections were incubated with primary antibodies for Nox4, Nox2, p47phox, NT, and CTGF o/n at 4°C. Sections were then incubated with the EnVision+ System-Labeled Polymer-HRP Anti-Rabbit (for Nox4, Nox2, p47phox, and CTGF detection) or Anti-Mouse (for NT detection) for 1 h at room temperature. Signal development was performed with H2O2, using 3,3′-diaminobenzidine as a chromogen. The immunodetection of each protein was repeated at least in a minimum of three preparations from different animals.
Histomorphological analysis
Histological evaluation was performed by two independent observers in a blinded fashion. Kidney sections were stained with collagen-specific picrosirius red (Sirius red F3BA in aqueous picric acid). Quantitative morphometric evaluation of glomerular and interstitial fibrosis was performed in 50 glomerular and 10 interstitial random fields of renal slides of each rat (n=10 animals per group).
Statistical analysis
Statistical significance was determined using Student's t-test or a one-way ANOVA followed by a Scheffe's test once normality had been proved (Shapiro–Wilks test). A p-value<0.05 was considered statistically significant.
Additional material is available in the Supplementary Data.
Footnotes
Acknowledgments
This work was funded through the agreement between the Foundation for Applied Medical Research (FIMA) and UTE project CIMA, Department of Education of Government of Navarra (87/2007), Ministry of Science and Innovation (RECAVA RD06/0014/0008) (SAF2008-04228), and European Union (InGenious HyperCare, LSHM-CT-2006-037093). The authors thank Ana Montoya, Raquel Ros, Laura Martínez and Idoia Rodríguez, and the Morphology and Imaging Department for their technical support.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.