
Abstract
Reactive oxygen species (ROS) exert various biological effects and contribute to signaling events during physiological and pathological processes. Enhanced levels of ROS are highly associated with different tumors, a Western lifestyle, and a nutritional regime. The supplementation of food with traditional antioxidants was shown to be protective against cancer in a number of studies both in vitro and in vivo. However, recent large-scale human trials in well-nourished populations did not confirm the beneficial role of antioxidants in cancer, whereas there is a well-established connection between longevity of several human populations and increased amount of antioxidants in their diets. Although our knowledge about ROS generators, ROS scavengers, and ROS signaling has improved, the knowledge about the direct link between nutrition, ROS levels, and cancer is limited. These limitations are partly due to lack of standardized reliable ROS measurement methods, easily usable biomarkers, knowledge of ROS action in cellular compartments, and individual genetic predispositions. The current review summarizes ROS formation due to nutrition with respect to macronutrients and antioxidant micronutrients in the context of cancer and discusses signaling mechanisms, used biomarkers, and its limitations along with large-scale human trials. Antioxid. Redox Signal. 19, 2157–2196.
III. Biomarkers of Oxidative Stress
VII. Dietary Fashion and Oxidative Stress
VIII. Antioxidant Supplementation and Oxidative Stress: A Complicated Riddle
I. Introduction
W

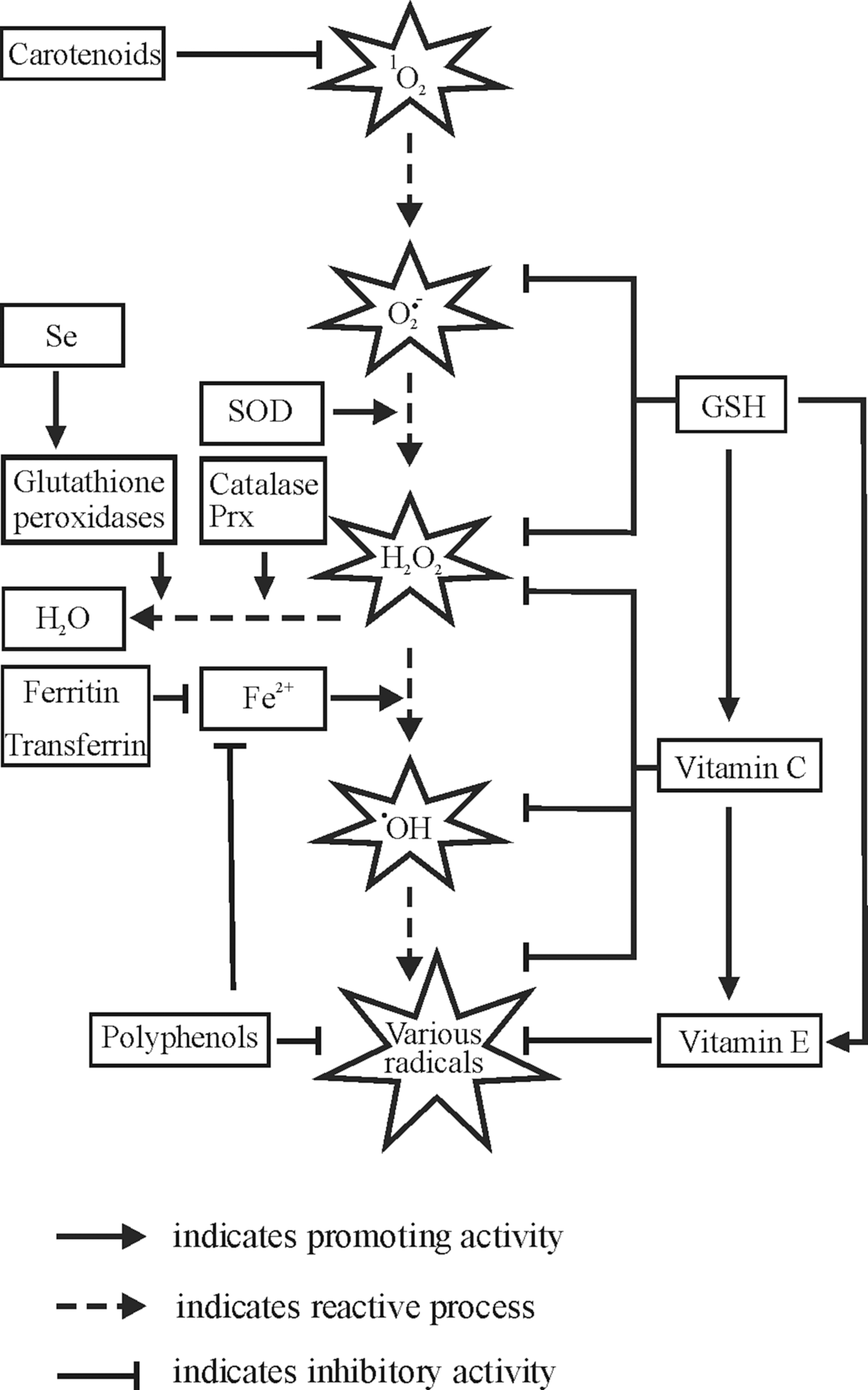
The dismutation of O2 −• to oxygen and hydrogen peroxide (H2O2) may occur spontaneously, but in mammalian cells, it is usually catalyzed by superoxide dismutases (SODs) (141). Although H2O2 is a relatively stable molecule, it may be converted to several other ROS, including hydroxyl radicals (•OH), which in turn react at extremely high rates with almost any biomolecule they encounter (35, 156). The formation of •OH from H2O2 can be catalyzed by Fe2+ in a so-called Fenton reaction, by other transition metals such as Cu2+ in Fenton-like processes as well as by nonphysiological stimuli, including ultraviolet (UV) radiation (169).
Due to research within the last decades, it is now also universally recognized that ROS, especially O2 −• and H2O2, may act as second messengers for signaling processes in different organisms, ranging from bacteria to mammals (96). It has been shown that ROS modulate diverse physiological processes, including the regulation of growth factor signaling, the hypoxic response, and inflammation. Thereby, ROS signaling involves primarily the chemical reactions with lipids and proteins. The action of ROS is balanced by the antioxidative capacity of the cells or an organism that is brought about by the antioxidant enzymes and specific low-molecular-weight antioxidants. However, it appears that a concentration threshold exists above which ROS cannot be kept in the low range of concentrations necessary for intracellular signaling by the endogenous antioxidant defense system. The resulting disturbance in the prooxidant–antioxidant balance in favor of the former is commonly referred to as oxidative stress. Further, dysregulated ROS formation was shown to occur in a number of diseases such as type II diabetes, cardiovascular diseases, or cancer (60, 85, 124, 180, 398). These diseases are frequently associated with a Western lifestyle and a nutritional regime. Importantly, formation and degradation of ROS appear to involve the different compartments of cells in a more or less specialized manner. It appears to be of special interest that the compartment-specific production of ROS due to different nutritional regimes may have an impact on the pathogenesis and/or the outcome of several diseases, especially cancer. Therefore, it is the aim of this review to summarize the current status of the role of nutrition on ROS formation with specific consideration of the cellular compartments and its role during cancer.
II. Intracellular Sources of ROS
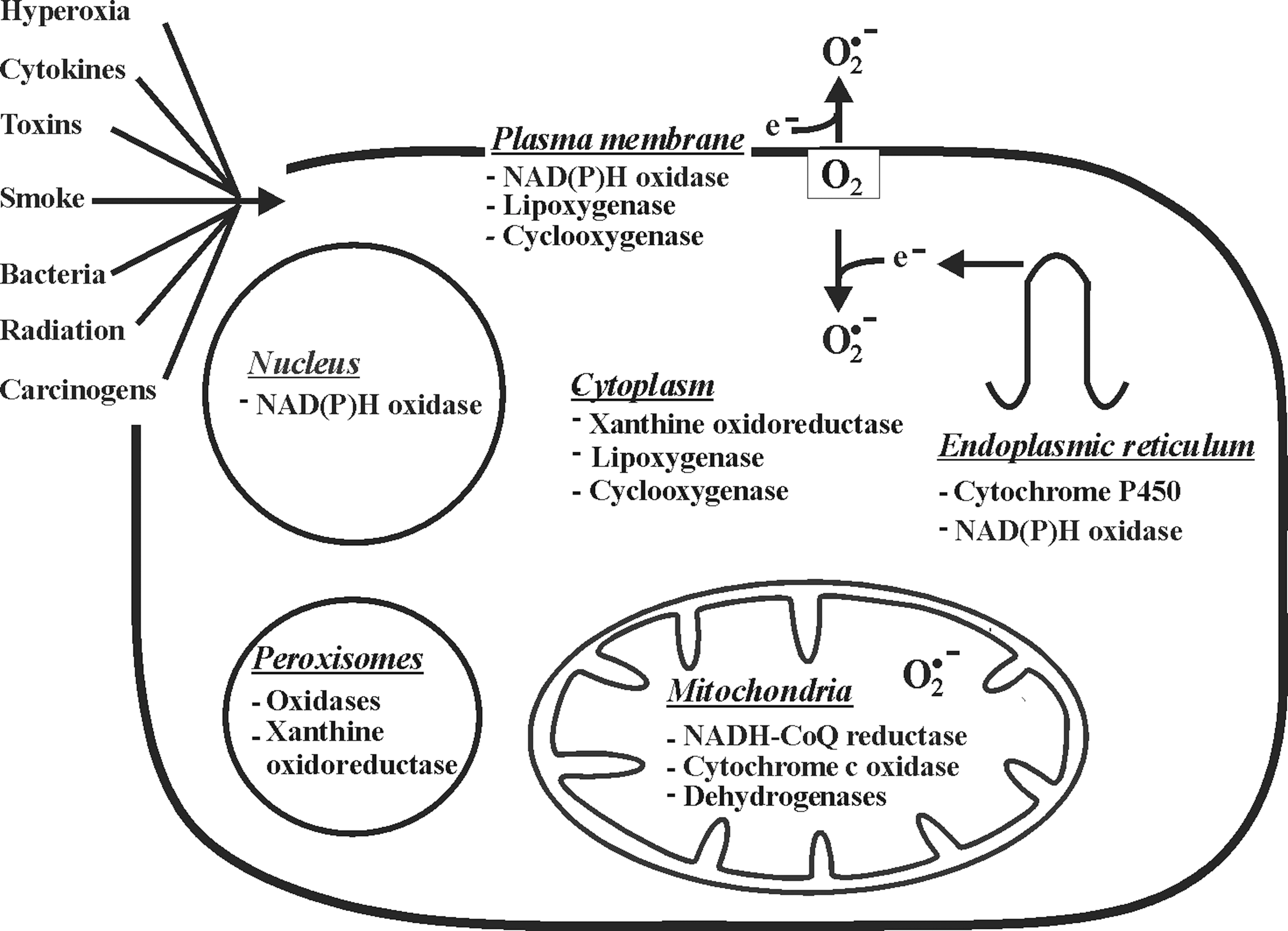
The fact that ROS can be both signaling molecules and toxic agents makes it necessary for an organism to produce them in a well-regulated manner to maintain homeostasis at the cellular level. Cellular production of ROS can be either nonenzymatic or enzymatic (Fig. 2). The major nonenzymatic triggers for ROS formation in vivo are ionizing and UV radiation, toxic chemicals, and drugs. While the majority of intracellular ROS is generated as byproducts of respiratory chain function, various enzymes such as NADPH oxidases, xanthine oxidoreductase (XOR), several peroxisomal oxidases, enzymes of the cytochrome P450 family, cyclooxygenases, and lipoxygenases were found to contribute to ROS production.

Due to localization of the numerous potential ROS sources within different cellular compartments, it is evident that ROS can be generated in the mitochondria, cell and nuclear membranes, cytoplasm, endoplasmic reticulum (ER), lysosomes, and peroxisomes (Table 1; sites of O2 −• production are shown in Figure 3). Depending on reactivity and membrane permeability, ROS have different diffusion distances (379). Thus, species with low membrane permeability such as O2 −• or with very high reactivity such as •OH are largely restricted to the compartments where they are generated. Although H2O2 was considered to be membrane permeable, recent data indicated that its diffusion is limited (241). Therefore, H2O2 transmembrane gradients in vivo are formed mainly due to the different formation/consumption rates on either side of a membrane as well as due to specific membrane transport (14, 35). Since the formation of ROS within each cell compartment has been extensively discussed in several other recent reviews (13, 42, 253, 318, 357), we will give only a summarized overview here.
DHODH, dihydroortate dehydrogenase; KGDH, α-ketoglutarate dehydrogenase; PDH, pyruvate dehydrogenase; XOR, xanthine oxidoreductase.
Mitochondria are known to be major sources of ROS (253). Interestingly, there seems to be a sex difference in ROS formation and degradation between the mitochondria from males and females (367). This appears to contribute to the prolonged lifespan of women. Indeed, it was shown that the female mitochondria produce less ROS than male, and that females display a higher expression of antioxidant enzymes due to the action of estrogens (367). The mitochondrial electron transport chain (ETC) can leak electrons to oxygen, constituting an important source of O2 −•. In addition to five more minor sites producing ROS in mammalian mitochondria (42), it is usually believed that the majority of mitochondrial ROS are formed at complex I (NADH-CoQ reductase) and complex III (cytochrome c oxidase) of the ETC, where electrons can react with oxygen to produce O2 −•. While complex I generates O2 −• only within the mitochondrial matrix, complex III generates O2 −• not only in the matrix but also in the intermembrane space (263). Moreover, the mitochondrial flavoprotein enzymes, in particular pyruvate dehydrogenase and α-ketoglutarate dehydrogenase, have also been recognized as sources of O2 −• (53). It was recently shown that dihydroortate dehydrogenase, an enzyme associated with the mitochondrial ETC and required for pyrimidine synthesis, is involved in N-(4-hydroxyphenyl) retinamide-induced ROS production (129). Further, the direct reduction of molecular oxygen to H2O2 was described to occur within the intermembrane space of mitochondria due to the action of the redox enzyme p66Shc (114). For this function, p66Shc utilizes reducing equivalents from the ETC through the oxidation of cytochrome c. The fraction of total oxygen that is diverted into mitochondrial ROS production varies according to different estimates and is between 0.15% and 4% (42).
The main ROS producers in the cellular plasma membrane appear to be NADPH oxidases (47). Production of O2 −• by the NADPH oxidase complex in phagocytes is a well-known phenomenon of the oxidative burst involved in bacteriocidal activity (21). The heterodimeric transmembrane parts of the classical NADPH oxidase complexes from phagocytes consist of the NOX2 and p22phox. As of yet, further NOX proteins have been identified, and currently the NOX family members are designated NOX1–5 and DUOX1/2 (47). While the expression of NOX2 appears to be confined to polymorphonuclear cells, macrophages, and endothelial cells, NOX1 seems to be present in the plasma membrane of various cell types (3, 21, 47). In general, NOX activity is supposed to be tightly controlled by regulatory subunits. Among these are the classical NOX2 regulators p47phox and p67phox, their homologs NOXO1 and NOXA1, the DUOX1/2 regulators DUOXA1 and 2, and the GTPase Rac. Recently, several other proteins have been identified that were proposed to have a regulatory function on NOX activity under certain conditions, thus adding more complexity to the NADPH oxidase network. With respect to nutritional signaling, especially those nonphagocytic cell NADPH oxidases appear to be important, and they seem to be involved in production of those ROS that are considered to act as second messengers during angiogenesis, hypoxic responses, oxidative protein modification (281), and wound healing; for example, a DUOX-generated tissue gradient of H2O2 was found to be responsible for wound detection in zebrafish (262).
The actions of cyclooxygenases and lipoxygenases during conversion of arachidonic acid, which can be released into the cytoplasm from membrane phospholipids due to the activity of phospholipase A2, also contribute to intracellular formation of ROS (68). Another cytoplasmic source for O2 −• is XOR, a molybdoflavoprotein, which exists in the two interconvertible forms xanthine oxidase (XO) and xanthine dehydrogenase (XDH). Both catalyze the two terminal reactions of the purine degradation pathway, that is, the conversion of hypoxanthine to xanthine and xanthine to uric acid (142). During these reactions, XDH can reduce oxygen and NAD+, whereas XO only reduces oxygen to yield O2 −• (365). Interestingly, the subcellular localization of XOR remains enigmatic; XOR was initially described to be solely cytosolic (154), but later it has been detected also in the peroxisomes of hepatocytes, in endocytic vesicles, and occasionally on the rough ER of Kupffer and liver sinusoidal cells (105).
The ER is also an important compartment where intracellular ROS production takes place (24, 121). Various oxygenases and oxidases (e.g., cytochrome P450 enzymes, flavin-containing monooxygenases, and prolyl and lysyl hydroxylases) involved in a multitude of functions are important sources of ROS formation. Especially, during protein folding, ROS and glutathione disulfide (GSSG) are formed to a major extend as byproducts from the action of endoplasmic oxidoreductin 1 (ERO1) sulfhydryl oxidases (15, 304). In addition to ERO1, the NADPH oxidases NOX4 and NOX5 are believed to be important ROS producers in the ER (281). In line, iron deposits in the ER and in close contact to the nucleus (58) may be involved in the production of •OH by a Fenton reaction (212).
Lysosomes also have a key role in ROS generation. An ETC in lysosomes, necessary for intralysosomal proton accumulation, promotes a three-electron reduction of O2, thus giving rise to formation of •OH (265). Additionally, lysosomes contain reduced iron liberated by autophagic degradation of metalloproteins that can favor a Fenton reaction. Unlike iron, lysosomal copper is probably not redox-active because it forms complexes with various thiols (195).
Among the ROS-producing enzymes found in peroxisomes are various flavin-dependent oxidases (13). Substrates of these enzymes are long-chain fatty acids and their derivatives, hydroxyacids, amino acids, purines, and polyamines (13). The result of their activity is mostly the production of H2O2; it has been estimated that about 35% of all H2O2 produced in the rat liver derives from peroxisomal oxidases (318). The most important metabolic process in peroxisomes contributing to formation of H2O2 is β-oxidation of fatty acids by three acyl-CoA oxidases, that is, palmitoyl-CoA oxidase, trihydroxycoprostanoyl-CoA oxidase, and pristanoyl-CoA oxidase (318). Interestingly, some reports suggested that a peroxisomal XO formed by the proteolytic conversion of XDH upon a number of stress stimuli provides an important enzymatic source not only for H2O2 but also for O2 −•(318).
Finally, the nucleus is a potential place for ROS production, and apparently, it is especially vulnerable to ROS action due to its nucleic acid contents. Nuclear NOX-dependent ROS production was shown to be involved in oxidative stress-responsive gene modulation. Among the NOX isoforms, NOX4, though predominantly localized in the ER, can also be found in the nuclei of several cell types, indicating that NOX4-dependent ROS production in the nucleus may contribute to regulation of redox-dependent transcription factors and gene expression involved in cell growth, differentiation, senescence, and apoptosis (357). Further, chelatable iron that can favor a Fenton reaction has also been found in the nucleus (280), and other metal ions such as Cu2+ are thought to be present on the chromosomes and might also accelerate the production of ROS (285). In line, nuclear ferritin known to sequester free iron was found to ameliorate oxidative damage in embryonic avian corneal epithelial cells (51). Thus, each compartment is involved in ROS generation and therefore may be involved to a different extend in the modulation of cell growth, differentiation, senescence, and apoptosis, and thus in carcinogenesis.
III. Biomarkers of Oxidative Stress
A. Direct measurement of ROS production
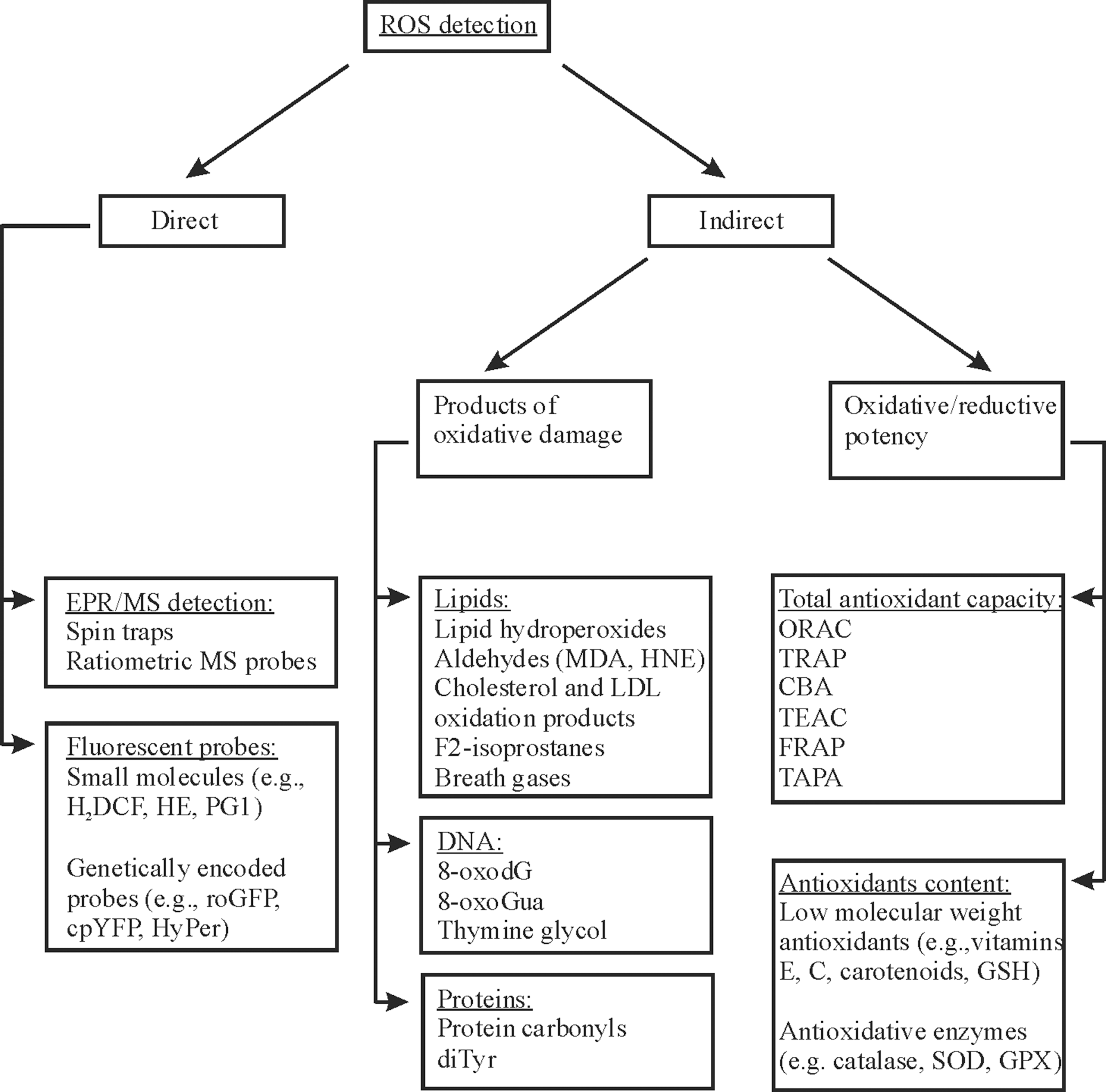
The direct measurement of ROS produced in vivo is not really standardized yet, and a number of possible assays employ electron paramagnetic resonance (EPR) techniques and dyes or probes that become fluorescent upon reaction with ROS (88, 237, 311) (Fig. 4). Intracellular ROS can be measured in vivo either with small-molecule fluorescent probes (173, 240, 300) or with fluorescent probes based on genetically encoded constructs (237). The most common small molecule used is 2′,7′-dihydrodichlorofluorescein (H2DCF), which is oxidized by H2O2 to fluorescent 2′,7′-dichlorofluorescein (DCF) (173). The small probes like DCF have the advantages of simplicity and a high signal-to-background ratio, but they are very susceptible to artificial side reactions. For example, oxidation of H2DCF can occur also in the absence of H2O2 being stimulated by metals, peroxidases, and cytochrome c (237). Moreover, during the reaction, an intermediate DCF•− radical is formed that can react with oxygen forming O2 −• with the consequence of an artificial amplification of the fluorescence signal (173). Hydroethidine (HE) is the most popular fluorogenic probe to detect intracellular O2 −• by formation of fluorescent 2-hydroxyethidium. However, HE was also shown to be an unreliable indicator for intracellular O2 −• formation (399). Thus, there is a demand for improved chemical probes, and these are now being developed. For example, Peroxy Green 1 (PG1) and Peroxy Crimson 1 (PC1) have recently been proposed for an improved selective measurement of H2O2 in cells (240, 300). However, the increase in selectivity was gained at the expense of sensitivity, since the H2O2-mediated boronate deprotection reaction is a relatively slow and irreversible process, and therefore PG1/PC1 cannot be used for detection of transient changes in the intracellular H2O2 concentrations (300).
Some limitations of the chemical probes may be overcome by the development of genetically encoded redox probes that are more specific and allow quantitation of ROS production within live cells (237). The basis for these redox-sensitive probes is green and yellow fluorescent proteins (GFP and YFP). Introduction of specific cysteine residues and other changes into GFP and YFP resulted in a strong redox-dependent change in fluorescence and development of reduction–oxidation-sensitive GFPs (roGFP1 and roGFP2) and circularly permuted YFP (cpYFP) (137, 272). The roGFPs seem to be best suited to determine the steady-state redox conditions and to document redox changes that happen in the frame of minutes (237). Fusing of roGFP2 to glutaredoxin (Grx1-roGFP2) enabled development of a more sensitive probe for the measurement of glutathione (GSH) redox potential (125), whereas coupling of roGFP2 to yeast peroxidase Orp1 (Orp1-roGFP2) and cpYFP to the regulatory domain of Escherichia coli OxyR (HyPer) improved fluorescence imaging of H2O2 in live cells (30, 126). Although cpYFP was initially described to be also suitable for intracellular O2 −• detection (375), another report claimed that cpYFP is highly responsive to pH, but not to O2 −• in mitochondria (319). Thus, genetically encoded redox fluorescent probes represent a new generation of ROS-detecting probes that still require improvement, especially with respect to specficity.
Another approach to measure ROS in vivo is EPR where spin traps react selectively with free radicals to generate a relatively stable product (298, 393). In particular, spin trapping with nitrones such as 2,2,6,6-tetramethyl-1-piperidinyl-1-oxyl (TEMPO) can generate, upon reaction with ROS, more-persistent nitroxide adducts detectable by EPR (366). A 5-diethoxyphosphoryl-5-methyl-1-pyrroline-N-oxide (DEPMPO) and a 2-(2-carboxyethyl)-2-methyl-4-phenyl-2H-imidazole-1-oxide (MCPIO) spin trap can be used for the detection of •OH and O2 −• by EPR and mass spectrometry (MS) (298). The latter has the advantage to detect also EPR-silent probes. Recently, a ratiometric MS approach was developed to assess mitochondrial H2O2 in fruit flies in vivo by using MitoB probe (72). MitoB consists of a triphenylphosphonium (TPP) cation driving its accumulation within mitochondria, conjugated to an arylboronic acid that reacts with H2O2 to form a phenol, MitoP. Quantifying the MitoB/MitoP ratio by liquid chromatography–tandem mass spectrometry (LC-TMS) allowed measuring the H2O2 levels within mitochondria (72).
B. Indirect measurement of ROS generation
Despite recent advances, direct detection of intracellular ROS is still rather problematic due to their high reactivity and instability. All the optical methods for ROS measurements, described above, are not applicable to most animal models in vivo and to humans (254). Moreover, analyzing ROS in living animals and humans by conventional EPR has significant limitations due to the use of high-frequency microwaves. Therefore, ROS generation is usually indirectly assayed by detecting specific and relatively stable biomarkers in various biological fluids (serum or plasma, urine, saliva, semen, etc.), which are convenient to obtain without invasive procedures. These biomarkers are generated due to specific reactions of ROS with certain biomolecules (268). The most commonly used indicators of high ROS levels are products of lipid peroxidation, DNA oxidation, and protein oxidation (Fig. 4).
1. Lipid peroxidation markers
Lipid hydroperoxides are nonradical intermediates of lipid peroxidation, and their measurement may provide information about ROS levels (Fig. 5A). Several chromatographic, enzymatic, and colorimetric methods have been developed for their detection (40). Since analysis of lipid hydroperoxides is quite difficult, because they are very unstable and decompose rapidly, these methods can be used only with cautious interpretation of obtained results (147). Among the most widely used membrane lipid peroxidation markers are 4-hydroxy-2-nonenal (HNE) and malondialdehyde (MDA). Both are relatively stable α,β-unsaturated aldehydes that are formed during decomposition of unstable dienic hydroperoxides; however, some α,β-unsaturated aldehydes are permanently formed under physiologic conditions during apoptosis and proliferation, and therefore their appeareance does not necessarily need to reflect oxidative stress. The usual test for MDA production is thiobarbituric acid-reactive substances (TBARS), a spectrophotometric assay that quantifies a chromogen produced by the reaction of MDA with thiobarbituric acid. Although MDA is often used as a marker for ROS-mediated lipid peroxidation, the MDA levels give usually an overestimation of ROS formation. This is due to the nutrient-derived peroxides and peroxide decomposition products that can be absorbed through the gut and therefore influence MDA levels (19). Moreover, MDA can be produced also by the degradation of endoperoxides and not only during lipid peroxidation. Thus, MDA/TBARS appears not to be a reliable test system for in vivo ROS measurements, and the European Food Safety Authority recently disapproved the use of TBARS as a biomarker for lipid peroxidation (147).

Low-density lipoproteins (LDL) play an important role during lipid and cholesterol metabolism. While oxidation products of cholesterol such as 7β-hydroxycholesterol may predict the development of lung cancer (210), the last decade showed that measurement of oxidized LDL (oxLDL) appears to be a good indicator for atherosclerosis and coronary artery disease (363). The oxLDL measurements with immunoassays provide a reliable and easy method, although standardization of the available oxLDL assays is still needed (147).
The F2-isoprostanes (especially 5-, 8-, 12-, 15-F2-isoprostanes), which originate from the peroxidation of arachidonic acid residues in lipids, are often considered to be the best indicators of lipid peroxidation in humans (320). They are not influenced by the diet and can be assessed in various biological fluids, including urine, plasma, synovial fluid, bronchoalveolar fluid, bile, lymph, and cerebrospinal fluid. Urine is considered most suitable for F2-isoprostane measurements, since isoprostanes in plasma have a shorter half-life, and their levels vary widely during the day. Indeed, increased levels of urinary 15-F2t-isoprostane were shown to be associated with a risk for breast cancer and hepatocellular carcinoma (309, 386). Further, the most abundant F2-isoprostane is 8-isoprostaglandin-F-2 (8-iso-PGF2), and increased 8-iso-PGF2 concentrations, as detected by gas chromatography–mass spectrometry (GC-MS), LC-MS, and enzyme immunoassays, have been associated not only with cancer but also with atherosclerosis, pulmonary diseases, and Alzheimer disease (250). However, the use of F2-isoprostanes also has its limitations. Extensive auto-oxidation of arachidonic acid occurs in blood plasma leading to artificial generation of these compounds (305). Moreover, though F2-isoprostanes are quite stable in isolated samples of body fluids, once released into circulation, they are rapidly metabolized in vivo (76). Another way of assessing whole-body lipid peroxidation is based on the analysis of exhaled hydrocarbon gases (alkanes), for example, ethane and pentane formed during decomposition of peroxyl radicals (88).
2. Markers indicating DNA, protein, and amino acid oxidation
The direct reactions of ROS with DNA bases as well as the action of membrane lipid peroxidation products such as MDA lead to formation of numerous DNA adducts; some of the most common are shown in Figure 5A and B (74). Among the DNA adducts, 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-hydroxydeoxyguanosine, 8-oxodG) is probably the best characterized and most widely used as a biomarker of oxidative DNA damage (67). Several methods for quantifying 8-oxodG are available, including HPLC, GC-MS, and LC-MS (67). However, since 8-oxodG can arise from degradation of oxidized dGTP in the DNA precursor pool and since under certain conditions oxidative DNA damage occur without changes in 8-oxodG levels (296), the measurement of 8-oxodG alone is not enough to correctly assess the ROS-dependent DNA damage. The simultaneous measurement of numerous oxidation products, including 5-hydroxymethyl-2′-deoxyuridine, 5,6-dihydroxy-5,6-dihydrothymine (thymine glycol), and 8-oxo-7,8-dihydroguanine (8-oxoGua), in the same sample seems to be a more correct indication for oxidative DNA damage than the detection of a single product (87).
There are fewer studies assessing the potential of ROS-dependent protein and amino acid modifications. However, the formation of the protein carbonyl (PCO) groups, the tyrosine and phenylalanine oxidation products resulting in o,o′-dityrosine (di-Tyr) and orthotyrosine, has been reported as relevant and are widely used. PCOs may be generated by the oxidation of several amino acid side chains (e.g., Lys, Arg, Pro, and Thr) as well as by interaction of these amino acids with α,β-unsaturated aldehydes such as MDA and HNE (76). PCOs can be detected in plasma and are chemically stable (88). Likewise, di-Tyr is also stable and detectable in human plasma and urine (34). Among the methods developed to quantify di-Tyr are HPLC, GC-MS analysis after derivatization, and LC-MS/MS analysis with atmospheric pressure chemical ionization or electrospray ionization (271).
3. Total antioxidant capacity
As an alternative to the measurement of single or a limited number of oxidative damage products, the overall oxidative status can be assessed on the basis of the oxidative/reductive potency of a given specimen (e.g., serum or urine) (325). Depending upon the reactions involved, the total antioxidant capacity (TAC) assays used can roughly be classified into two types: assays based on hydrogen atom transfer (HAT) and assays based on ET (150). The majority of HAT-based assays apply a competitive reaction scheme, in which the antioxidant and substrate compete for thermally generated peroxyl radicals through the decomposition of azo-compounds. These assays include oxygen radical absorbance capacity, total radical trapping antioxidant parameter (TRAP), and crocin bleaching assays. The most frequently used targets of oxidation in these assays include luminol, phycoerithin, and 20–70-dichlorofluorescin. The ET-based assays measure the capacity of an antioxidant in the reduction of an oxidant, which changes color when reduced depending on the sample's antioxidant concentrations. ET-based assays include the Trolox equivalence antioxidant capacity (also referred to as total antioxidant status), ferric ion reducing antioxidant power (FRAP), total antioxidant potential assay using Cu2+ as an oxidant, and 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical-scavenging capacity assay. All these assays differ from each other in terms of substrates, probes, reaction conditions, and quantitation methods. Therefore, the results from different assays are often very controversial (88, 238). It can be concluded that assessment of plasma TAC alone cannot be considered as a valid tool for evaluating such a complex biological process as oxidative stress.
4. Measurement of antioxidants
A number of studies measured the levels/activities of various antioxidants as indirect markers of changed ROS production. The enzymes commonly assayed as biomarkers of oxidative stress both in tissue samples and in biological fluids include SOD, glutathione peroxidase (GPX), and catalase (see section V) (88). Under conditions of oxidative stress, low-molecular-weight antioxidants such as vitamins E and C and carotenoids are consumed, and therefore their levels often decrease (see section VI). The most commonly used methods to evaluate the levels of low-molecular-weight antioxidants both in body fluids and in tissue samples are LC-MS and GC-MS (88). Measurement of S-glutathionylated proteins combined with depletion of reduced GSH and a decreased GSH:GSSG ratio are useful indicators of oxidative stress (76), especially in cancer (259). However, recently, the use of GSH levels to indicate whether a cell is under oxidative stress was challenged by a study showing that oxidized glutathione is transported to cellular vacuoles in yeast and mammalian cells (248).
Several methods have been optimized to identify and quantify glutathione forms in human samples, including spectrophotometric, fluorometric, and bioluminometric assays, often applied to HPLC analysis, as well as the more recently developed GC-MS and HPLC-electrospray ionization-MS techniques (76). However, the reported levels of GSH, GSSG, and S-glutathionylated proteins in blood vary widely (1 to >500 μM for GSSG), which indicates that current methods of detection are not standardized and affected by various artifacts (76, 308).
C. The limitations of oxidation markers in use
It is well known that oxidative stress-related markers, such as 8-oxodG, HNE and MDA, are commonly elevated in tumor patients and often correlate well with risk factors, such as viral infection and inflammation (223, 323, 390). For example, the correlation between multiple oxidative stress biomarkers and breast cancer, squamous cell carcinomas, leukemia, and colorectal cancer was found (106, 221, 390, 400). As discussed later in more detail, there are data showing that diet composition, especially the supplementation with antioxidants, influence the plasma or urinary levels of oxidative stress markers (see sections VI and VII) (48, 101, 232, 247). Some studies have evaluated oxidative stress biomarkers as particular cancer risk factors and simultaneously related those factors to nutrient intake. In particular, it was found that an antioxidant micronutrient cocktail can modulate oxidative stress in patients with a history of sporadic colorectal adenoma (149). In another study, the serum TAC was decreased with an increasing carbohydrate intake, but was increased with a higher vitamin A and vitamin C intake in breast cancer patients (390). It was also found that the intake of chicken and high-fat dietary products was associated with increased levels of plasma lipid peroxides, while vitamin E intake was associated with decreased lipid peroxides in women with breast cancer (364). By contrast, several other studies showed that cancer-relevant biomarkers of oxidative stress were unchanged by diet. For instance, increased consumption of selenium-rich food and subsequent increase in blood Se concentrations did not affect biomarkers of oxidative stress in healthy Australian males (387). No differences in the DNA and lipid oxidative damage markers and antioxidant capacity between intervention groups of healthy, nonsmoking men receiving 2, 5, or 8 servings/day of vegetables and fruit were found (45).
The use of biomarkers in human studies requires an ability to sensitively, specificly, and reproducibly measure oxidative damage to the relevant molecular targets (132). The substances used as biomarkers should be stable, generated only by ROS action, and not influenced by their presence in food. As it was discussed before, the existing biomarkers fulfil these demands only to some extent. It is known that different biomarkers do not measure identical aspects of oxidative stress, and while there are usually good correlations between the concentrations of several peroxidation products, including MDA, F2-isoprostanes, conjugated dienes, GSH, and PCOs, these parameters are often not correlated with other biomarkers such as the concentration of antioxidants and DNA fragmentation (88, 320). Even more, the usual ROS markers insufficiently reflect intracellular processes leading to ROS generation and are not intracellular compartment specific. Indeed, the biomarkers of lipid oxidation are generated in the plasma membrane as well as in various intracellular membranes. DNA oxidation markers cannot be considered specific to the nuclear compartment not only because of mitochondrial DNA (mtDNA) oxidation but also due to oxidation in the nucleotide pool, since single-nucleoside triphosphates are less protected from oxidative damage than the chromatin-bound genomic DNA. Although some types of oxidative products of nucleotides are not amenable to DNA synthesis (e.g., thymine glycol and 5′,8-purine cyclodeoxynucleoside), other species may be tied into the nucleic acids (322). However, none of the analytical methods used to measure DNA adducts identifies where the oxidative DNA damage is located (76).
D. Compartment-specific ROS measurement
The activity of compartment-specific antioxidant enzymes (see section V) in correlation with a disease could be very informative concerning the details of intracellular generation of ROS. A good example of such enzyme that has compartment-specific isoforms is SOD, and changes in the cytosolic Cu/ZnSOD (SOD1), mitochondrial MnSOD (SOD2), or extracellular SOD (EcSOD; SOD3) activity or expression can be correlated to ROS levels. Elevated MnSOD levels were found in tissues of patients with gastric carcinomas and adenomas (182) as well as squamous cell carcinomas, whereas no significant changes in Cu/ZnSOD were detected (106, 391). By contrast, increased Cu/ZnSOD activity was found in tissue samples from patients with colorectal cancer (331), in leukemia patients (400), and in patients with breast cancer (22, 288). EcSOD (but not Cu/ZnSOD or MnSOD) expression was found to be significantly decreased in lung cancer (350). However, the obvious problem in such studies is the limited availability of healthy human tissues in which antioxidant enzyme activities can be assayed. Although the changes in antioxidant enzymes may correlate the ROS levels to intracellular compartments, the significance of these changes is often difficult to interpret, since expression of antioxidant enzymes is often induced by oxidative stress itself, and, moreover, their decreasing activities and levels upon oxidative stress, probably due to structural modification by free radicals, were also reported (67, 230). For instance, the levels of catalase, a rather peroxisome-specific antioxidant enzyme, were found to be induced (288), decreased (331), and unaffected (22) in different types of cancer. In line, while high MnSOD expression correlates with a poor prognosis, advanced progression, and an invasive and metastatic phenotype of gastrointestinal tumors (182), it is a factor correlated with favorable survival in patients with buccal mucosal squamous cell carcinomas (106).
While the biological significance of changes in antioxidant enzymes is under discussion and a common view has not been reached, methods for quantitative detection of compartment-specific ROS generation in live cells is needed. Recently, a new class of compartment-specific reporters for H2O2 imaging was generated (300, 337). Thereby, a mutant of the DNA repair protein O6-alkylguanine-DNA alkyltransferase (SNAP tag) was targeted to the different compartments and later conjugated with PG1. The SNAP-PG adducts respond to H2O2 with an increase of green fluorescence in the target organelle. However, these probes react irreversibly with H2O2, and cannot be used to monitor transient changes in H2O2 concentrations. By contrast, genetically encoded ROS probes such as roGFPs (Fig. 6) and HyPer can be targeted to distinct subcellular compartments and react in a reversable manner.

Another important progress was made with generation of mitochondrion-targeted probes for ROS detection (Fig. 7) (82). The promising strategy for fluorescence imaging of mitochondrial ROS in living cells is to create dyes that contain both a ROS-responsive element and a mitochondrion-targeting moiety. Mitochondria Peroxy Yellow 1 (MitoPY1) is an example of such a dye (81). Reaction of MitoPY1 with H2O2 triggers a fluorescence increase by its conversion to MitoPY1ox. Mitochondrial O2 −• may be assayed by using mitochondrion-targeted HE (Mito-HE, MitoSOX red), a TPP+ conjugated to HE via an alkyl chain. Further, a recently developed mitochondrion-targeted fluorescent lipid peroxidation probe MitoPerOx was shown to report mitochondrial lipid peroxidation in isolated liver cells (284). Highly reactive ROS, including •OH, peroxynitrite (ONOO−), and hypochlorous acid (HOCl), can be measured in mitochondria by rhodamine-like fluorophores capped with either a 4-aminophenyl aryl ether (MitoAR) or a 4-hydroxy aryl ether (MitoHR) group (187). However, all the limitations attributed to the small fluorescent probes discussed before (see section III.A) are also applicable to mitochondrion-specific small probes (237, 254, 399). Alternatively, genetically encoded redox probes such as roGFPs and Grx1–roGFP2 were successfully targeted to mitochondria in several cell lines in vitro (237) and in vivo in transgenic mice (128). Those analyses could corroborate that mitochondria are most reduced organelles, whereas the ER and Golgi are much more oxidized. Accordingly, the more oxidized compartments appear to react faster when cells are challenged by a reducing agent or environment (Fig. 6). Further, the use of genetically encoded redox probes and a new chemical strategy to trap the in vivo redox state of the transgenic biosensor enabled quantitative mapping of the GSH redox potential and H2O2 in the cytosol and mitochondria during development and aging in Drosophila (9).

Despite recent progress, it should be noted that the optical approaches to compartment-specific ROS measurement are often applicable only to cultured cells or to optically amenable organisms such as zebrafish or worms. Two-photon confocal microscopy technique is needed to probe deeper beneath the surface in nontransparent transgenic animals expressing fluorescent proteins (254). As an alternative to optical ROS detection, the use of mitochondrion-targeted nitrone traps for EPR detection of ROS (Mito-DEPMPO) was proposed (139) as well as the ratiometric MS probe targeted to the mitochondrial matrix (MitoB), which showed an increase in mitochondrial H2O2 production with age in Drosophila (72).
The potential for the development of markers indicating high ROS levels in a compartment-specific manner was confirmed by the effects of antioxidants designed to selectively remove ROS from mitochondria in vivo (53). The most extensively studied mitochondrion-targeted antioxidant is mitoquinone (MitoQ); it contains an antioxidant quinone moiety that is covalently linked to a lipophilic TPP+ via a 10-carbon alkyl chain. The TPP+ part allows that MitoQ efficiently accumulates in mitochondria in a membrane potential-dependent manner (333). MitoTempol and MitoVitE are other promising mitochondrion-targeted antioxidants derived from tempol (and antioxidant nitroxide) and vitamin E, respectively, covalently attached to TPP+. Mitochondrion-targeted peptides and plastoquinone derivatives have been developed as well (332, 343). Mitochondrion-targeted antioxidants were used in a number of in vivo studies in rats and mice and found to be protective against oxidative damage in many different pathologies, such as diabetes, cardiovascular disease, cancer, and neurodegenerative disorders (61, 122, 217). In line, MitoQ oral administration decreased protein oxidative damage markers as well as mitochondrial markers of oxidative damage in several animal models (342), though it did not influence expression and activity of mitochondrial MnSOD in wild-type mice (306). It was recently shown that mitochondrion-targeted antioxidants could protect pancreatic β-cells against oxidative stress under glucotoxic and glucolipotoxic conditions (207). However, the results of two phase II human trials (with Parkinson's disease and chronic hepatitis C patients) are less convincing (333). In this respect, it should be taken into account that presence of high quantities of TPP+-conjugated probes in the mitochondria may lead to quenching, mitochondrial uncoupling, and changes in the mitochondrial function (53, 165).
IV. ROS and Tumorigenesis
High levels of ROS are often associated with various pathological conditions, including cardiovascular dysfunction, diabetes, and premature aging (60, 338). The finding that oxidative stress is also a common characteristic of several tumors links elevated levels of ROS and oxidative stress with tumorigenesis. Increased ROS production in cancer cells is believed to be caused by increased metabolic activity, mitochondrial dysfunction, as well as alterations of gene expression (124, 274). ROS-mediated oxidative stress is proposed to promote tumorigenesis by harmful processes such as DNA, protein, and polysaccharide damages, peroxidation of lipids, as well as various epigenetic changes, which all modify the cellular function at the genomic, proteomic, and metabolomic levels and can result in uncontrolled proliferation and apoptosis. Potentially, each of these steps during tumorigenesis can contribute to further generation of ROS, which then can cause more mutations, directly interfere with signaling pathways, thereby activating oncogenes and/or inactivating the tumor suppressor genes (Fig. 8). Since the role of ROS in cellular signaling, tumorigenesis, and cancer therapy was recently discussed in several reviews (124, 226, 294, 398), we would describe it here only briefly.
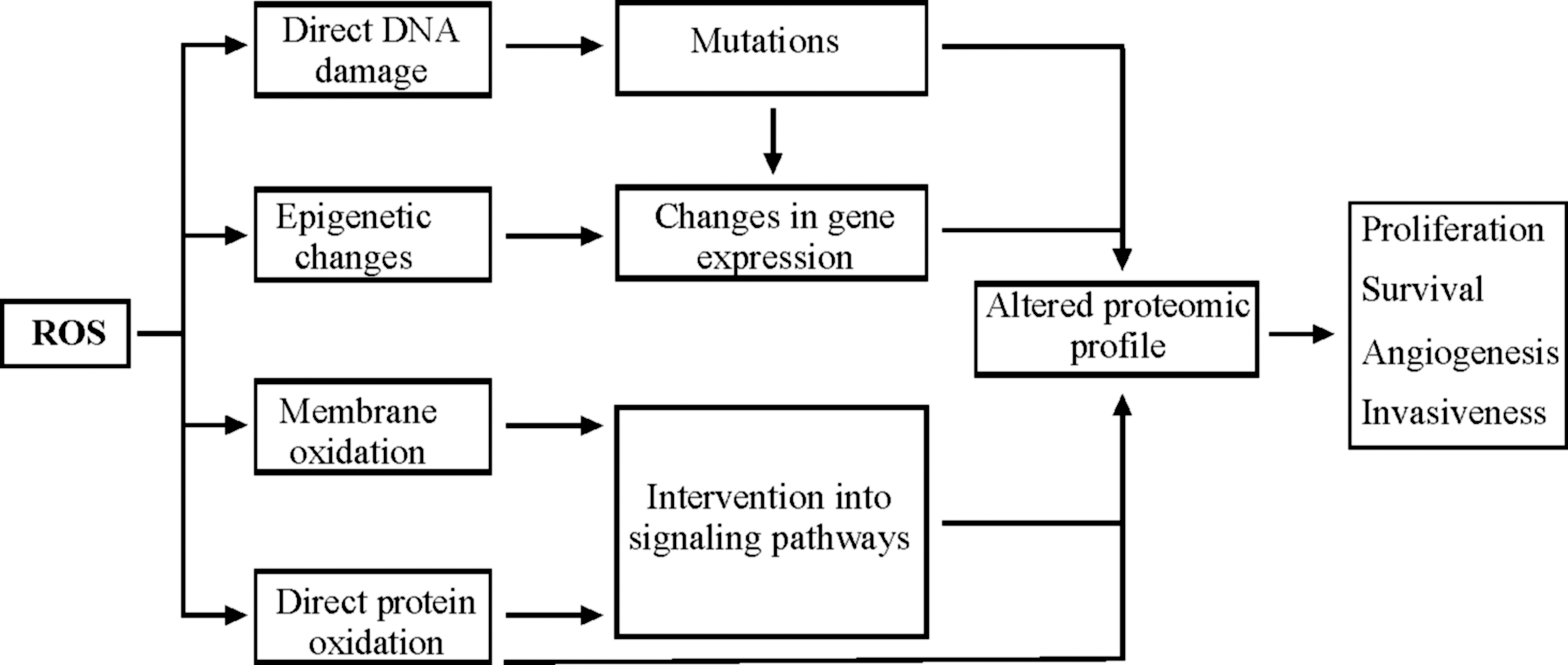
A. ROS-induced oxidative damage
Different ROS have an unequal ability to damage DNA and proteins. While •OH are known to cause most damage to DNA by reacting with all four bases, 1O2 selectively attacks guanine (381). It is unlikely that O2 −• and H2O2 react with DNA bases directly, though they contribute to DNA damage by forming •OH. The ROS mostly responsible for protein damage is •OH that acts by abstracting a hydrogen atom from the protein polypeptide backbone (359). The abstraction of a hydrogen atom leads to the formation of a carbon-centered radical initiating the formation of various products such as peroxyl radicals, alkyl peroxide, and alkoxyl radicals (338). In addition, lipid peroxidation contributes to the formation of DNA adducts (see section III.B.2, Fig. 5A) as well as etheno-adducts and propano-adducts, which are also considered to be genotoxic (258, 359).
In line, a number of reports indicate that nuclear DNA damage appears to be important in tumorigenesis. In comparison to healthy individuals, an almost 50% higher level of the DNA oxidation product 8-oxoGua has been detected in the lung, breast, and prostate cancer patients and has been implicated to cause the Ras and p53 mutations found in both lung and liver cancer cells (352). In comparison to its nuclear counterpart, mtDNA is even more vulnerable, since it lacks histones, has limited repair capacity, and lies in a closer proximity to the sites of mitochondrial ROS generation (54). In line, mtDNA mutations have been observed in a great number of human cancers (54). Although mutation of almost all genes within the mitochondrial genome is affiliated with various types of tumors (244), one particular region in the mtDNA, which is called displacement loop (D-loop), is known as a mutational hotspot. The D-loop mutation is associated with hepatocellular carcinoma (348), ovarian cancer (214), breast cancer (349), colorectal cancer (206), and melanoma (347).
Epigenetic changes such as DNA methylation can also be influenced by ROS (102). For example, ROS-mediated DNA base modifications such as 8-oxodG formation adjacent to a cytosine may prevent methylation of the latter (353). The resulting hypomethylation may contribute to induction and suppression of gene expression, thus enabling the cells to achieve novel characteristics that may contribute to tumorigenesis. ROS are also associated with hypermethylation (102) that can lead to increased overall mutation rates by downregulating expression of repair genes such as O-6-methylguanine-DNA methyltransferase (MGMT) and mutL homolog 1 (MLH1) (398).
B. ROS-dependent regulation of signaling pathways
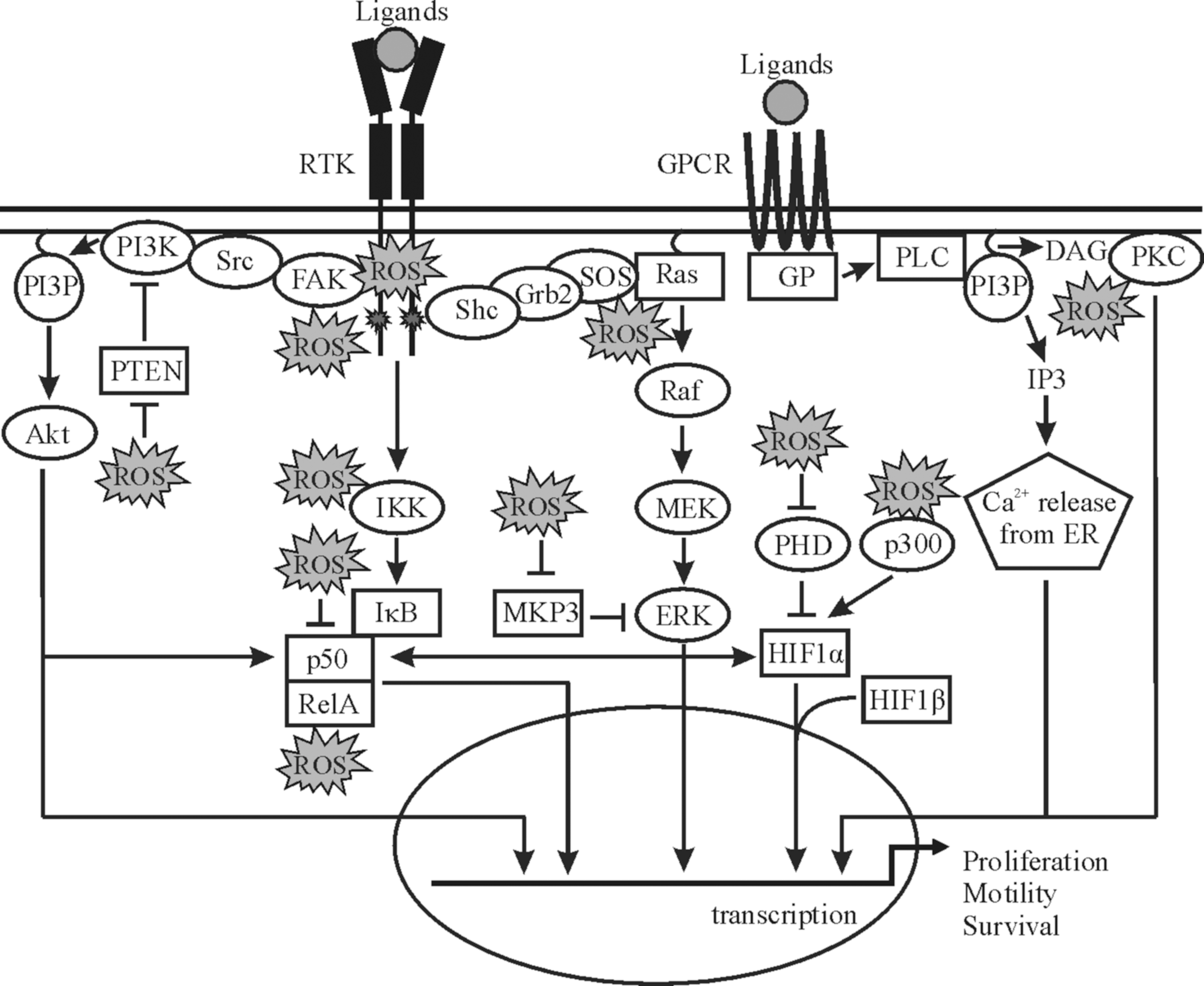
The involvement of ROS in the carcinogenesis is also connected to the physiological role of ROS in several major signaling cascades (175, 249, 371). Interestingly, growth factors, hormones, and coagulation factors, including platelet-derived growth factor (PDGF), angiotensin II, and insulin, have been shown either to lead to the generation of ROS or to use them at least partially as messengers. Therefore, an aberrant accumulation of ROS within the cell may imitate the signaling function of these stimuli and lead to a ligand-independent modulation of these signaling cascades. Although not all of the ROS-mediated interventions are tumorigenic, a majority of them are supposed to be. This appears often to be due to stimulation of various components of the mitogen-activated protein kinases (MAPK) and/or phosphatidylinositol 3-kinases (PI3K)/Akt signaling cascades, while a number of other pathways, including protein kinase C (PKC), wnt/β-catenin, hedgehog, and Notch signaling, can be also modulated by ROS (73, 197, 371, 389). Since intracellular pathways can be influenced by ROS in a bewildering number of ways (294), we will provide here only some typical examples that are also presented in Figure 9.
1. MAPK signaling
Although the mechanisms are not fully resolved, ROS initiate activation of the extracellular signal-regulated kinase (ERK) and c-Jun NH2-terminal kinase (JNK) MAPK cascades by causing autophosphorylation of growth factor receptors such as epidermal growth factor receptor (EGFR) and PDGFR in a ligand-dependent manner (185). In addition, various components from the major MAPK signaling cascades were shown to be responsive toward changes in the cellular ROS levels. In particular, a direct activation of the ERK1/2 cascade is achieved by oxidative modification of Ras at Cys118 (196). This modification inhibits GDP/GTP exchange, thereby activating Ras. In addition, ROS appear also to indirectly affect ERK1/2, since MAPK/ERK kinase 1/2 (MEK1/2) inhibitors are found to thwart ROS-mediated ERK1/2 activation (233), thus pointing to a regulation at the level of MEK1/2 or, for example, mitogen-activated protein kinase phosphatase (MKP3), a negative regulator of ERK1/2. With the latter, it was shown that ROS can promote proteosomal degradation of MKP3, thereby activating ERK1/2 (62).
2. PI3K/Akt signaling
ROS such as H2O2 were found to activate protein kinase B (PKB), also known as Akt in different cell types (236). Biologically relevant concentrations of ROS were found to activate growth factor receptors, including EGFR and PDGFR, known to stimulate the PKB/Akt pathway (236). ROS were also found to initiate PKB/Akt signaling by promoting tyrosine phosphorylation of the nonreceptor tyrosine kinase called focal adhesion kinase (FAK) in a PI3K-dependent manner, since FAK acts as an upstream mediator of PI3K activation (334). PI3K activation is counter-regulated by the tumor suppressor phosphatase and tensin homolog deleted on chromosome 10 (PTEN). An inactivation of PTEN would enhance PKB/Akt activation, and indeed, it was found that ROS can inactivate PTEN by oxidation of Cys124 (200). Further, loss of PTEN leads to depletion of antioxidant enzymes, resulting in increased oxidative stress (151).
3. PKC signaling
Certain isoforms of PKC have been reported to be modulated by ROS. For example, the isoforms α, βI, and γ of cPKC, δ, and ɛ of nPKC, and ζ of aPKC were catalytically activated upon treatment of cells with H2O2 (190). Although the kinase is unknown yet, the catalytical activation of these PKC isoforms was due to phosphorylation of tyrosine residues located in the catalytic domain (190). It is plausible that also a conserved cysteine-rich region found in PKCα typically responsible for binding diacylglycerol may also be involved in PKC activation (179).
4. Protein tyrosine phosphatases
Another important regulatory mechanism of ROS action is oxidation of the catalytic cysteine residues of protein tyrosine phosphatases (PTP), which leads to their reversible inactivation (273). Both the growth factor-independent general cellular redox state and extracellular ligand-stimulated ROS production can cause PTP oxidation. Classical PTPs, which specifically dephosphorylate protein phosphotyrosine, as well as dual-specificity phosphatases, including MAPK phosphatases, are subject to ROS-dependent oxidation (273).
5. NF-κB signaling
A number of transcription factors were shown to act either positively or negatively in either a direct or an indirect response to ROS; among those ROS-sensitive transcription factors are nuclear factor kappa B (NF-κB), nuclear factor erythroid-2-related factor-2 (Nrf2), Sp1, redox factor 1 (Ref-1), cJun, hypoxia-inducible factor-1α (HIF-1α), and hypoxia-inducible factor-2α (HIF-2α) (198, 336). Activation of redox-sensitive transcription factors can lead to expression of genes associated with survival and further malignant transformation of cancer cells. In particular, a number of tumor suppressor proteins such as p53, forkhead box O (FoxO), retinoblastoma, p21, and p16 were shown to play a role in activating the expression of genes encoding antioxidant enzymes, including MnSOD and catalase, in response to oxidative stress (371).
In particular, NF-κB activation appears to be ultimately linked with ROS, since a number of agents activating NF-κB are also able to trigger ROS formation. ROS were found to mediate phosphorylation of inhibitor of NF-κBα (IκBα) kinases IKKα and IKKβ, leading to release of free NF-κB dimers, which could further translocate to the nucleus (174). Further, Toll-like receptor 2 (TLR-2) was shown to be involved in the activation of NF-κB by H2O2 in cardiac myocytes (104). Likewise, tumor necrosis factor-α initiated a redox-dependent phosphorylation of Ser276 on RelA. It was then found that phosphorylation of Ser276 that was necessary for NF-κB activation is mediated by the catalytic subunit of protein kinase A, which was induced by ROS (164). By contrast to the activating action of ROS, it was shown that with p50, another member of the NF-κB family, oxidation, S-glutathionylation, or formation of sulfenic acid at Cys62 results in reduced DNA-binding abilities (228, 282).
6. HIFα signaling
Early findings demonstrating that addition of H2O2 to cells grown under hypoxia destabilized HIF-1α (374) and HIF-2α (63) support the concept that ROS play an important role in HIF signaling. The redox processes modifying both HIF-1α and HIF-2α appeared to act in a direct and indirect manner. Direct regulation affects the C-terminal transactivation domain in which the Cys800 of HIF-1α and the Cys848 of HIF-2α seem to be critical for transactivation (180). Similar to activator protein-1 (AP-1), the oxidation/reduction state of these cysteines is dependent on the presence of Ref-1 (58). In addition to Ref-1, other coactivators such as steroid receptor coactivator-1 and transcription intermediary factor 2 were found to interact with the HIF-1α and HIF-2α C-terminus in a redox-dependent manner (58), thus again supporting a role for ROS in modulating HIFα activity.
The indirect action of ROS in hypoxia occurs predominantly via regulation of prolyl hydroxylases (PHD), redox-sensitive kinases, and phosphatases. The PHDs that are necessary for oxygen-dependent HIF degradation belong to a family of dioxygenases that require oxygen, Fe2+, 2-oxoglutarate, and ascorbate [reviewed by (168)]. These enzymes act only when iron is in the Fe2+ state, which is lost during hydroxylation of the substrate. Therefore, Fe2+ is likely regenerated by a radical cycling system in which ascorbate reduces Fe3+ to Fe2+, thus rendering the enzyme active (186). In addition to HIF-1α, ROS derived from ER-localized NOX4 were shown to stabilize HIF-2α by preventing its hydroxylation, thus allowing formation of transcriptionally active HIF-2 (83).
The HIFα signaling is known to undergo a crosstalk with both the PI3K/Akt and MAPK cascades where ROS appear to play a mediatory role [reviewed by (84)]. Subsequently, antioxidants or inhibitors of NADPH oxidases prevented the signaling via PI3K/Akt to HIF-1α (120). In addition, ERK1/2 can phosphorylate HIF-1α (84) and contribute to HIF-1α induction by a number of nonhypoxic stimuli known to induce ROS production such as angiotensin-II (302), prostaglandin E2 (107), or shock waves (373). However, it appears that the ROS sensitivity of ERK1/2 and the subsequent induction of HIF-1α are stimulus- and cell type-dependent (120, 373). Further, the p38 MAPKs and the p38 upstream kinases MKK3 and MKK6 (181) were shown to be involved in the induction of HIF-1α by thrombin (120) and chromium (VI) (109).
C. Dual role of ROS in tumorigenesis
Together, the mechanisms for redox regulation of the major intracellular signaling pathways are complex and not yet resolved until the last detail, thus requiring further investigations. Since the function and regulation of the major intracellular cascades such as MAPK and PKB were comprehensively reviewed (57, 289, 294), the present review will not describe how each of the pathways is involved into tumorigenesis. In brief, depending on the kinetics of activation and inactivation, the subcellular localization, the formation of complexes with partner proteins, and the availability of substrates, these pathways largely contribute to cell cycle progression, apoptosis and survival, angiogenesis, invasion, and metastasis (124).
The general picture of ROS involvement into malignant transformation is further complicated by the fact that while moderately increased ROS levels lead to tumor-promoting mutations of oncogenes and tumor suppressor genes as well as to induction of signaling pathways responsible for carcinogenesis, excessive ROS levels can suppress tumors (189). A number of studies demonstrated the ability of ROS to induce apoptosis of cancer cells; some studies also found that high ROS levels correlate with decreased proliferation of transformed cells, decreased tumor angiogenesis, and metastasis [listed in (124)]. A threshold concept for cancer cell therapy implies that it should be possible to preferentially accumulate ROS in cancer cells above the cellular tolerability level, thus inducing their death (189, 277). In accordance with this concept, a recent study demonstrated that piperlongumine, an alkaloid from Piper longum, increases ROS levels and apoptotic cell death in both cancer cells and normal cells engineered to have a cancer genotype, but it has little effect on either rapidly or slowly dividing primary normal cells (287). Therapeutic strategies exploiting the role of ROS both as an inducer and an inhibitor of tumorigenesis are being developed (294, 351). For example, procarbazine, motexafin gadolinium, elesclomol, 2-methoxyestradiol, and imexon are used to increase ROS, while minodronate and histamine are used to eliminate ROS (124). Similar to many chemotherapy drugs, radiation therapy also stimulates ROS generation, thus eliminating cancer cells (20).
V. The Antioxidant System
Under normal physiological conditions, formation and removal of ROS are balanced to avoid oxidative stress. In principle, an antioxidant can be any substance that delays/prevents formation or promotes removal of ROS or oxidative damage of a target molecule (127). The antioxidant defense system consists of a number of molecules among which several enzymes, such as SODs, GPXs, catalase, and simpler molecules such as GSH, represent major players (Fig. 10).
GSH is a three-residue peptide (γ-
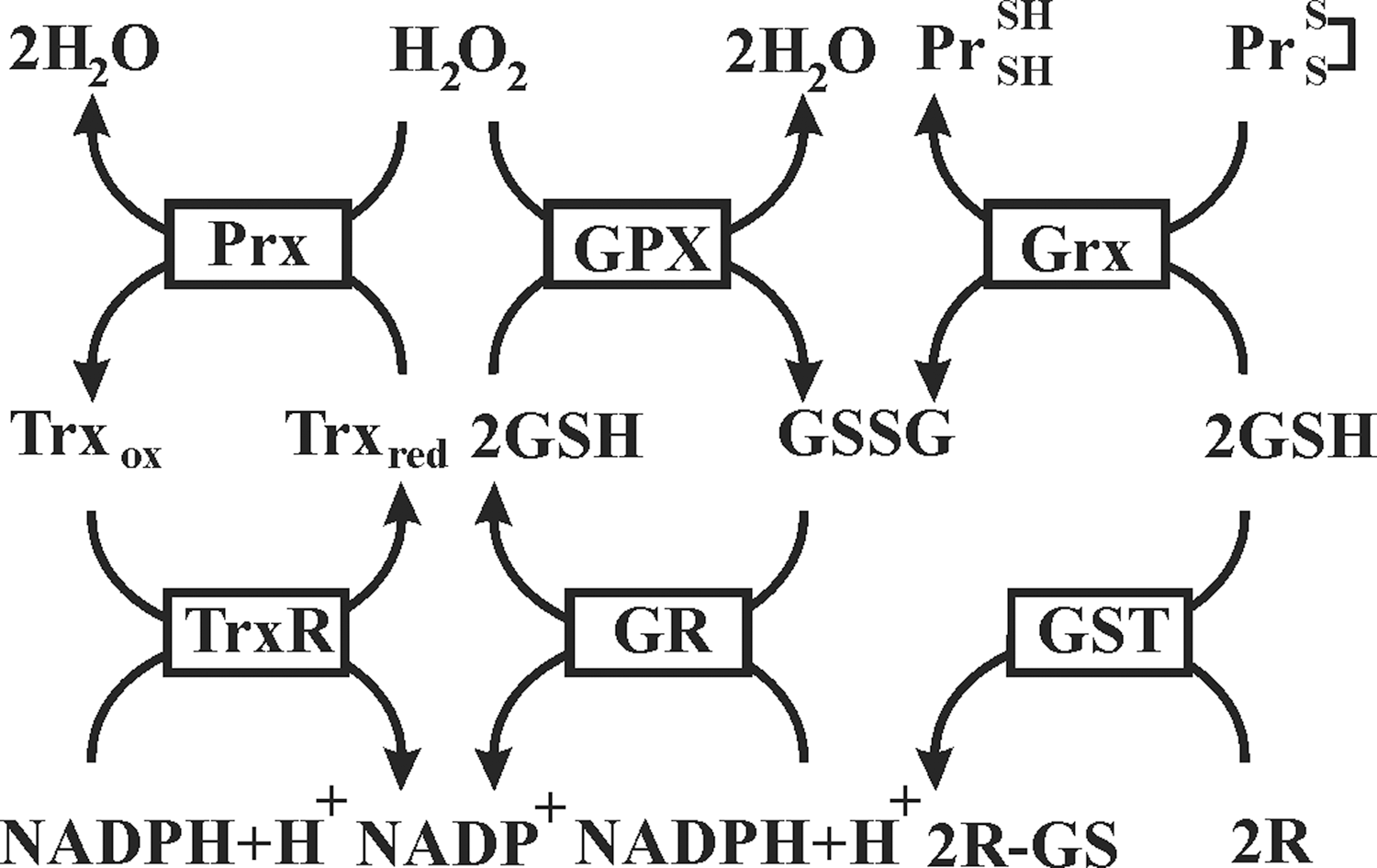
Among antioxidant enzymes, SODs catalyze the dismutation of O2 −• to H2O2, and catalase stops formation of •OH by converting H2O2 to oxygen and water. Peroxidase activity is also characteristic for antioxidant proteins of the peroxiredoxin (Prx) family. Prx are ubiquitous thiol-specific proteins that react with H2O2 at an exceptionally high rate (385). In addition to H2O2, Prx also reduce and detoxify peroxynitrite and a wide range of organic hydroperoxides. It is believed that Prx2 because of its high reaction rate and abundance trap almost all H2O2 in vivo (379). Oxidized cysteine residues of Prx are specifically reduced by thioredoxins (Trx), thiol–disulfide oxidoreductases that can be oxidized upon oxidative stress caused by a variety of stimuli (148). Oxidized Trx (as well as other oxidized cellular proteins) can be reversibly reduced by thioredoxin reductase (TrxR) in a NADPH-dependent manner (148).
Sequestration of free iron, necessary for a Fenton reaction, by iron-binding proteins such as ferritin and transferrin, is another way to reduce intracellular ROS production. Dietary micronutrients also contribute to the antioxidant defense system. Several water-soluble molecules, such as vitamin C, and lipid-soluble molecules, such as vitamin E, are considered to have antioxidant activity (see section VI). Some dietary minerals have antioxidative properties due to their role as cofactors of the corresponding antioxidant enzymes. In particular, selenium is a component of peroxidases GPX1–4 and TrxRs (see section VI.E) (29).
A. Intracellular and extracellular localization of antioxidants
Various components of the antioxidant system are unequally distributed between different intracellular compartments (Fig. 12 and Table 2). For example, in eukaryotic cells, the Cu- and Zn-containing Cu/ZnSOD is located mainly in the cytosol and peroxisomes, whereas another Cu-, Zn-dependent form of SOD, known as EcSOD, is secreted and forms a glycosylated heterotetramer bound to the extracellular matrix (141). In contrast, an Mn-dependent MnSOD is found in the mitochondrial matrix.
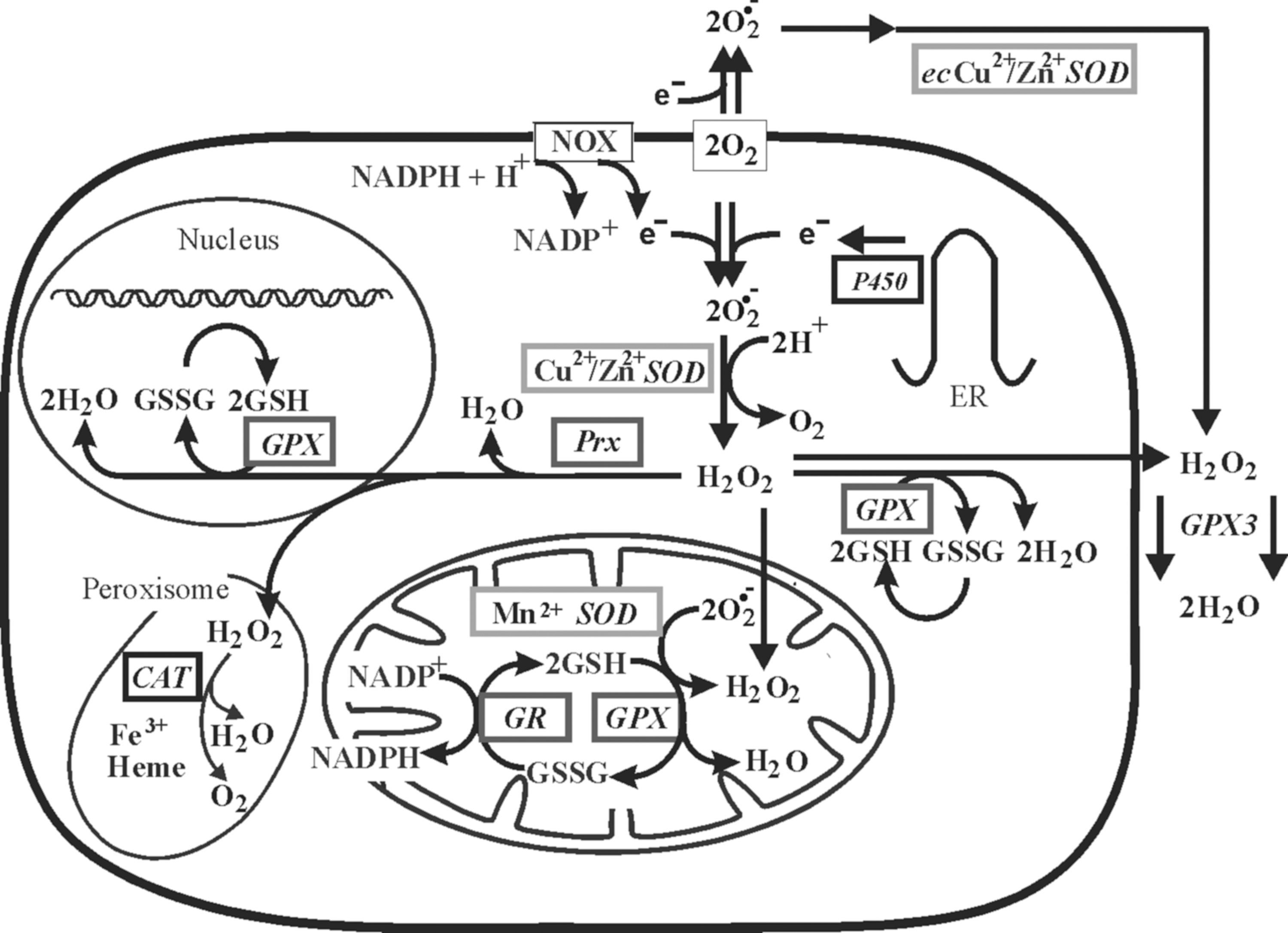
GPX, glutathione peroxidases; GSH, glutathione; Prx, peroxiredoxin; SOD, superoxide dismutase; Trx, thioredoxin; TrxR, thioredoxin reductase.
GPX1 is the ubiquitous enzyme that reduces cytosolic, mitochondrial, and, in some cells, peroxisomal peroxides, while very similar GPX2 is an epithelial-specific form with highest expression in the intestine (219). A secreted GPX3 form expressed in the lung and kidney and measurable in plasma can be important in the protection against external peroxides (219). There are three different isoforms of GPX4 (also known as phospholipid–hydroperoxide GPX) derived from a single gene: cytosolic (c-GPX4), mitochondrial (m-GPX4), and nuclear (n-GPX4) (155, 266). Unlike other GPX, GPX4 has a broad range of substrates; in addition to H2O2, these include derivatives from cholesterol and cholesteryl esters and thymine hydroperoxide (155).
Prx1, 2, and 4 function mainly in the cytoplasm, but are also expressed in the nuclei; Prx1 is also present in the mitochondria and peroxisomes, whereas Prx4 is found in lysosomes (116, 157, 267). While Prx3 is localized in the mitochondria (267), Prx5 is found in the mitochondria, cytoplasm, nuclei, and peroxisomes (324). Similarly, Trx1 as well as TrxR1 can be found in the cytoplasm and nucleus. By contrast, Trx2 and TrxR2 are localized to mitochondria, where they appear to have a protective function against oxidative stress (116, 267).
In addition to antioxidant enzymes, several mitochondrial electron carriers such as cytochrome c and ubiquinol possess antioxidant activity. Interestingly, the inner mitochondrial membrane appears to contain a high amount of the antioxidant vitamin E (134), whereas GSH and vitamin C are important antioxidants in the mitochondrial matrix (53). Lysosomes contain high concentrations of low-molecular-weight antioxidants such as GSH and vitamin C (195).
Data concerning oxidative stress induced by depletion of glutamate and glutamine indicate that the nuclear compartment may have a better protection against oxidative stress than the cytoplasm or mitochondria (117). Indeed, the nucleus contains antioxidants such as GSH, n-GPX4, Trx1, TrxR1, and Prx1, 2, 4, and 5. Data from Prx1-deficient cells show a higher DNA damage (93), and together with nuclear expression of GSTpi in cancer tissues associated with anticancer drug resistance indicate that the regulation and functions of the nuclear antioxidant system are distinct from similar processes in the cytoplasm (116).
B. The functions of antioxidant enzymes unrelated to their ROS-removing activities
Beside their activities in removing excessive ROS described above, antioxidant enzymes are involved in a wide range of cellular processes. The balance between ROS generation and removal is necessary to keep physiological processes running. The floodgate model proposes that while low fluxes of H2O2 are scavenged by antioxidant enzymes, primarily Prx, higher fluxes cause localized Prx inactivation, enabling redox-sensitive signaling proteins such as PTPs to be targeted (384). It was recently found that Prx1 associated with the membranes is transiently phosphorylated on Tyr194, and thereby inactivated both in cells stimulated by growth factors and in those at the margin of healing cutaneous wounds in mice (382). Thus, the localized inactivation of Prx1 allows the transient accumulation of H2O2 around the membranes, where signaling components are concentrated, while preventing the toxic accumulation of H2O2 elsewhere. In addition, GPX4 was found to be involved in an apoptosis-inducing factor (AIF)-mediated cell death pathway (321) and to be pivotal for male fertility (356). As another example, the Trx/TrxR system supports the activity of ribonucleotide reductase, inhibits apoptosis signal-regulated kinase-1, and controls a pathway for methionine sulfoxide reductase B2 reduction mediated by metallothionein (116).
Although excessive ROS generation is still considered one of the causes for malignant transformation of normal cells, this does not necessarily mean that antioxidants have antitumorigenic properties. While almost any chemical reducing an oxidant's activity in vitro could be classified as an antioxidant, this may occur independent of its ability to have beneficial effects in vivo (127). In addition, some molecules such as vitamin C and plant polyphenols, including quercetin, which are considered to be antitumorigenic, have an ability to act both as an antioxidant or a prooxidant depending on their concentration (see section VIII.A) (130, 296, 362). In fact this again seems to be counterintuitive, many antitumor agents act by inducing ROS production in cancer cells (see section IV.C). One of the possible ways of how elevated ROS inhibit cell proliferation is p16-dependent sustained activation of PKC δ (290, 346). In line, antioxidants not only protect normal cells but also can favor tumor growth. In particular, they were found to promote survival of cells that are metabolically impaired and therefore lack attachment to the extracellular matrix (315).
Together, it becomes clear that ROS are required for normal cellular functions while its excessive generation leads to development of diseases, including cancer. Certain threshold concentrations of ROS exist at which either survival or cell death responses may be triggered; these may differ dependent on the cell/tissue type and/or the cellular compartment in which ROS are generated.
VI. Traditional Antioxidant Nutrients
A. Vitamin E
Vitamin E is a term for a family of structurally related lipid-soluble components from which the naturally occurring members, the tocopherols and tocotrienols, are physiologically relevant with respect to ROS. They possess an ability to quench ROS in cell membranes and other lipid environments, thereby protecting against oxidation of polyunsaturated fatty acids (PUFA) (Fig. 13A). The best-known form of vitamin E is α-tocopherol, which is highly active in neutralizing peroxyl radicals. The reaction of peroxyl radicals with α-tocopherol is 10,000 times faster than with PUFA (49). During interaction with peroxyl radicals, α-tocopherol donates a phenolic hydrogen atom and generates an α-tocopheroxyl radical that is relatively stable (41). Under normal conditions, the α-tocopheroxyl radical does not react with the membrane components. Rather, it reacts with a second radical to form a nonradical product, or it can be reconverted to α-tocopherol by vitamin C or other water-soluble reductants (41).
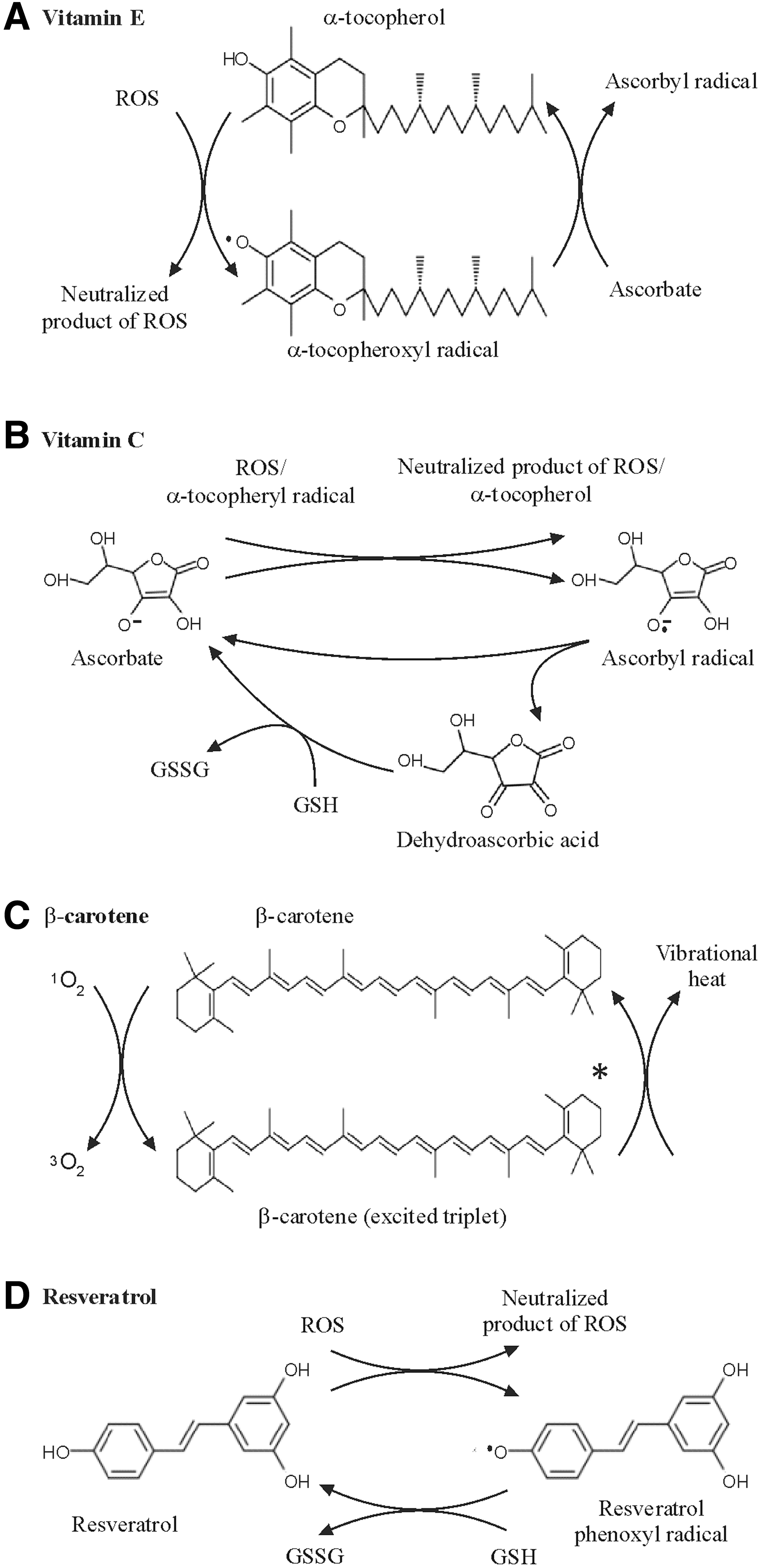
The protective role of vitamin E against ROS was indicated in several studies. By using a TBARS assay as a marker for ROS-mediated MDA production, it was found that dietary supplementation of vitamin E reduced the plasma or urinary TBARS levels (48, 232, 247). However, a number of other studies did not show the protective effect of vitamin E against oxidative damage in human subjects (261), and it should be considered that results from TBARS assays, as discussed in section III.B1, are no longer considered reliable to determine oxidative damage. In line, it has been argued that at physiological concentrations, α-tocopherol does not act as an antioxidant in vivo (261).
In general, vitamin E has been found to decrease malignant transformation in cell cultures and to suppress tumor development in animal models (Table 3) (6, 27, 41, 340). Tocopheryl phosphate, a more-active, natural derivative of α-tocopherol, was shown to have even more potent effects than α-tocopherol itself in terms of inhibition of cell proliferation and regulation of gene expression in vitro (261). It should be noted however that the tocopherols (α, β, γ, and δ-tocopherol) can decrease cell proliferation and inhibit tumor progression not necessarily due to their antioxidative properties, but via inhibition of several protein kinases, reduction of vascular endothelial growth factor production, decreased inflammation, etc. (18, 261, 299).
Reported protective effect is marked as (+), lack of protection or adverse effects are marked as (−).
ATBC, α-Tocopherol β-Carotene Study; CARET, β-Carotene and Retinol Efficacy Trial; HOPE, Health Outcomes Prevention Evaluation; HPS, Heart Protection Study; MDA, malondialdehyde; NPCT, Nutritional Prevention of Cancer Trial; PHS, Physician Health Study; ROS, reactive oxygen species; SCID, severe combined immunodeficiency; SELECT, Selenium and Vitamin E Cancer Prevention Trial; SU.VI.MAX, Supplémentation en Vitamines et Minéraux Anti-oxydants; TPA, 12-O-tetradecanoylphorbol-13-acetate; TRAMP, transgenic adenocarcinoma of the mouse prostate; WACS, Women's Antioxidant Cardiovascular Study; WHS, Women's Health Study.
Epidemiological studies have been unable to point out a strong link between tumor suppression and vitamin E supplementation. For example, while one nested case–control study carried out in the United States (prostate, lung, colorectal, and ovarian cancer screening trial) found a significant inverse association between serum α-tocopherol and prostate cancer risk, primarily among smokers (377), another case–control study (prostate testing for cancer and treatment, UK) found no evidence that vitamin E was associated with the overall prostate cancer risk or more aggressive prostate cancer phenotypes (113). Moreover, neither dietary nor supplemental intakes of nutrients often suggested for prostate cancer prevention, including vitamin E and selenium, were significantly associated with prostate cancer risk in the prospective cohort study Prostate Cancer Prevention Trial (PCPT; USA and Canada, 1994–2003) (192). Regular vitamin E supplement use for more than 10 years in a particularly large prospective cohort Cancer Prevention Study II involving 991,522 U.S. adults was associated with a reduced risk of bladder cancer mortality, but regular intake for a shorter duration was not protective (163).
The Linxian large-scale randomized intervention trial, carried out in China, involved thousands of malnourished people showing that α-tocopherol was potentially associated with reduction of stomach cancer when supplemented together with β-carotene and selenium (286). In total contrast, the α-tocopherol–β-carotene study (ATBC) found that α-tocopherol is associated with a higher frequency of stomach cancer while being protective against prostate and colorectal cancer; no effect was seen on lung and pancreatic cancer (8, 293). The beneficial and adverse effects of supplemental α-tocopherol in ATBC disappeared during postintervention follow-up (369). Low-dose supplementation of various antioxidants, including vitamin E, lowered total cancer incidence and all-cause mortality in men in a randomized, placebo-controlled Supplémentation en Vitamines et Minéraux Anti-oxydants (SU.VI.MAX) trial among French adults (140). A number of other randomized intervention trials with vitamin E, either alone or in combination, did not influence or prevent cancer development (119, 211).
B. Vitamin C
Vitamin C/
It was reported that individuals suffering from severe ascorbic acid shortage show elevated levels of 8-oxodG, a prominent product of DNA oxidation (101). Ascorbic acid supplementation was also reported to have a protective role on peroxidation of plasma lipoproteins induced by acute smoking (138), while another study in which ascorbic acid was given to individual smokers failed to decrease urinary excretion of 8-oxodG (283). In line, the reduction of ROS and the inhibitory role of ascorbic acid in tumorigenesis have been a controversial issue for over two decades (36). Although several early studies suggested that ascorbic acid could provide protective effects against tumorigenesis (36, 52), later studies suggested that such an antitumor role of ascorbic acid is hitherto inconclusive (Table 3) (50, 145, 163).
C. Carotenoids
Carotenoids are a large group of tetraterpenoid organic pigments that naturally occur in various photosynthetic organisms. Although they can readily react with ROS such as peroxyl radicals or alkoxyl radicals donating electrons, their principle role seems to be 1O2 quenching (234). One carotenoid molecule is supposed to quench about 1000 1O2 molecules (Fig. 13C) (191). Among over 700 carotenoids identified so far, ∼40 are normally found in human diet, but not all of them are uniformly retained in our body. Actually <20 carotenoids and related metabolites were detected in human tissues and plasma (264). The prominent members are α-carotene, β-carotene, lycopene, lutein, and β-cryptoxanthin. Although their role as antioxidants is well evident in vitro, the results obtained in vivo are less convincing (191, 232).
Probably due to its additional role as a precursor of vitamin A, the most studied carotenoid is β-carotene. It is speculated to have antitumorigenic properties, but the results of epidemiological studies are inconclusive, as some support its protective role, (80, 159, 170) while others do not (Table 3) (177, 312). The protective effect of β-carotene was tested in combination with α-tocopherol in the ATBC study, and after eight years of dietary supplementation, no protective effect against cancer was found (8, 293). It was even concluded that such supplementation might be potentially harmful, because β-carotene was found to be associated with increased cancer, most notably lung, prostate, and stomach, in the ATBC study (8, 369), as well as higher lung cancer mortality in the β-carotene and retinol efficacy trial (CARET) (118). In general, a recent meta-analysis of randomized controlled trials showed that β-carotene supplementation has no beneficial effect on cancer prevention, but it was associated with an increased risk not only of lung cancer but also of gastric cancer at doses of 20–30 mg/day, in smokers and asbestos workers (89), as well as significantly increased the risk of urothelial cancer (167).
By contrast, α-carotene has been reported to show more protection against skin, lung, liver, and colon cancer than β-carotene in murine models (256, 264). Apart from α- and β-carotene, several other carotenoids such as lycopene, lutein, and β-cryptoxanthin have also been reported to be potentially tumor suppressing (Table 3) (264, 327). However, the absence of large-scale randomized placebo-controlled human trials precludes us from concluding about the antioxidant and antitumorigenic property of these carotenoids.
D. Polyphenols
Dietary polyphenols are the most abundant antioxidants in human diets (136). There are almost 8000 structural variants of polyphenols ranging from single-ring-containing simple phenolic acid to highly polymerized condensed tannins, naturally present in plants as secondary metabolites. In principle, the aromatic rings of polyphenols that bear one or more hydroxyl moieties enable them to act as ROS scavengers and metal chelators. A large number of in vitro studies suggest that polyphenols are highly efficient antioxidants, since the reaction rate constants of several flavonoids with ROS in vitro surpass those of vitamin E (91).
Resveratrol is a polyphenol that has been drawing high interest of nutritionists for decades by being a candidate that may resolve the so-called French paradox (184). The role of resveratrol in diminishing oxidative stress and tumor formation was reported by several studies (Table 3 and Fig. 13D) (103, 202, 317). Pretreatment of mouse skin with resveratrol restored control levels of H2O2 and GSH as well as myeloperoxidase, GR, and SOD activities in the model of 12-O-tetradecanoylphorbol-13-acetate (TPA)-promoted mouse skin tumors (166). Resveratrol exerted a strong inhibitory effect on O2 −• and H2O2 production in macrophages stimulated with lipopolysaccharides or phorbol esters (224). In addition, resveratrol can modulate signal transduction and gene expression by nonantioxidant mechanisms (299). For example, resveratrol was shown to exhibit antitumor properties by downregulating several cellular signaling pathways such as ERK1/2, Akt, and NF-κB, and by activating the deacetylase sirtuin 1 (194, 299). Although resveratrol appears to provide a significant protection against ROS-mediated tumorigenesis, the low bioavailability does not contribute to its wide use (194, 260). Moreover, there are data indicating that resveratrol at low concentrations elicits prooxidant effects and inhibits induction of apoptosis by anticancer drugs such as vincristine and daunorubicin in human leukemia cells (4, 5). However, the promises of resveratrol are apparent warranting large-scale clinical trials that could provide a more concrete knowledge about the effect of resveratrol as an antitumorigenic nutrient.
Quercetin is another prominent member of polyphenols, and some murine studies pointed to a protective role of quercetin; however, several other studies failed to find its protective effect on ROS scavenging in vivo (136, 362). Several studies identified the role of apigenin as an antioxidant and antitumorigenic nutrient (276), and to our knowledge, one ongoing placebo-controlled intervention trial has been undertaken that involves apigenin together with epigallocatechin (119). Protective roles of other dietary polyphenols (curcumin, catechins, isoflavones, anthocyanidins, etc.) against carcinogenesis in various cell culture models were also reported, and their potential role as an antitumorigenic nutrient seems to be a busy field of research (Table 3) (28, 344, 388).
Taken together, dietary polyphenols exhibit a promising role as efficient ROS scavengers, and thereby considered to be potential antitumorigenic nutrients. Although their antioxidant capacity was shown experimentally, some features obviate us from being confident that they act only by protection against ROS-mediated tumorigenesis. Recent studies also suggest that many polyphenols exert their activity indirectly by activating the transcription factor Nrf2 (314). Under basal conditions, Nrf2 is bound with the cytoskeletal anchor Kelch-like ECH-associated protein 1 (Keap1), which targets Nrf2 for ubiquitination and proteasomal degradation (345). Several stimuli, including oxidative stress and nutritional polyphenols such as curcumin and catechins, induce dissociation of the Nrf2-Keap1 complex and nuclear localization of Nrf2, where it activates transcription at the antioxidant response element (ARE) (397). The ARE controls expression of genes involved in antioxidant defense such as GST A1 and A2, heme oxygenase 1, and NAD(P)H:quinone oxidoreductase (314, 397). In line, rats consuming polyphenol-rich apple juice as compared to polyphenol-free control juice showed induced expression of Nrf2 as well as catalase, GPX2, and GR (335). However, induction of Nrf2 needs to be interpreted with caution, since several tumors display enhanced levels of Nrf2 (345). In addition, a recent study showed that Nrf2 contributes to a proliferative phenotype by favoring glutamine metabolism and nucleotide synthesis (242).
The action of polyphenols is also hampered by rapid metabolism in human tissue and by gut bacteria. This may result in very low bioavailability of polyphenols, which could be insufficient to exert any significant antioxidant activity (313). Many metabolic products of polyphenols (e.g., methylated or glucuronated forms) are expected to have reduced or even no antioxidant activity due to the loss of radical scavenging hydroxyl groups (133). In line, a comprehensive recent analysis found no convincing evidence to show that antioxidant effects of polyphenols are relevant for cardiovascular health in humans (147). Nevertheless, protective effects of polyphenols are often observed, though it is suggested that such roles might be substantially localized to the gastrointestinal tract that usually remains glutted with various oxidants (133). In general, the lack of randomized controlled trials with isolated polyphenols using biologically and methodologically valid markers makes it impossible to conclusively evaluate their antitumorigenic potential.
E. Selenium
Selenium is a trace element that plays essential biological roles (295). While excessive intake of Se leads to toxicity known as selenosis, trace amounts of Se are necessary for the proper action of more than 20 selenoproteins (29, 193). Most selenoproteins are enzymes with an active site containing Se in the form of selenocysteine. The importance of Se in the antioxidant system comes from the fact that selenocysteine is present in the catalytic sites from various members of the GPX and TrxR families. Selenoprotein P is also implicated to have antioxidative properties (295). Indeed, Se-containing compounds exerted an ROS-scavenging effect in vitro and in various animal studies (64, 95, 201). Thus, GPX and TrxR were assumed to have protective effects against carcinogenesis (43, 56, 295).
A significant inverse relationship between geographical availability of Se and carcinogenesis has been observed (69), and a similar inverse association has also been found between baseline plasma Se levels and the risk of advanced prostate cancer in many, but not all case–control and prospective cohort studies (Table 3) (11, 152, 192, 204, 279). The antitumorigenic effect of Se and selenoproteins for prostate cancer was then further supported in a mouse model (86) as well as in a study showing that the GPX1 polymorphism can play an important role in predisposition to prostate carcinogenesis (16). Similarly, GPX2 has been reported to inhibit carcinogenesis by exerting hydroperoxide removal and suppression of COX2 expression (26, 44). In line, GPX1 knockout mice are highly sensitive to oxidative stress (78), and GPX2 knockout mice were found to be more susceptible to skin cancer upon γ-irradiation (372).
These data are contrasted by a multicenter, double-blind, randomized, placebo-controlled Nutritional Prevention of Cancer Trial (NPCT), showing that dietary supplementation of Se was associated with a significantly elevated risk for squamous cell carcinoma and nonmelanoma skin cancer (90). However, in the same study, the results from secondary endpoint analyses indicated that Se supplementation is correlated with reduced incidence of lung, colon, and prostate carcinomas (70). This result was in conformity with the Linxian and SU.VI.MAX trials (140, 222, 376). A NPCT substudy found that a 200 μg/day Se treatment decreased cancer by 25%, whereas 400 μg/day of Se had no effect (297). Moreover, the selenium and vitamin E cancer prevention trial (SELECT) showed no protective effects of Se against prostate, lung, and colon carcinogenesis (211). In general, a recent meta-analyses of multiple case–control and randomized studies led to the conclusion that Se is protective against prostate cancer, but only in a relatively narrow range of concentrations (152). Indeed, participants profited from a Se supplementation only when entering the study with a low basal Se status (152). Another meta-analysis of epidemiological data concluded that despite evidence for an inverse association between Se exposure and the risk of some types of cancer, these results should be interpreted with care due to the potential limiting factors of heterogeneity and influences of unknown biases (79). One of these factors is genetic background—a study among smokers demonstrated that depending on a 15-kDa selenoprotein genetic polymorphism, smoking individuals either can benefit from a higher Se intake or have an increased risk for lung cancer (161). Together, the results from a number of ongoing randomized controlled trials (79, 119) should help to better understand the general effect of selenium in cancer.
VII. Dietary Fashion and Oxidative Stress
During the metabolic processes involved in the breakdown of carbohydrates, lipids, and proteins, which are necessary to provide energy, a bulk of ROS is produced. From that point of view, every food can naïvely be considered a prooxidant independent of its constituents. Diets itself also contain several low-molecular-weight prooxidants that upon ingestion can readily trigger reactions causing ROS formation in vivo. The various enzymatic and nonenzymatic antioxidant processes and molecules counteract ROS generation under normal conditions, and often food itself contains several antioxidant components that can contribute to ROS scavenging (Fig. 14).
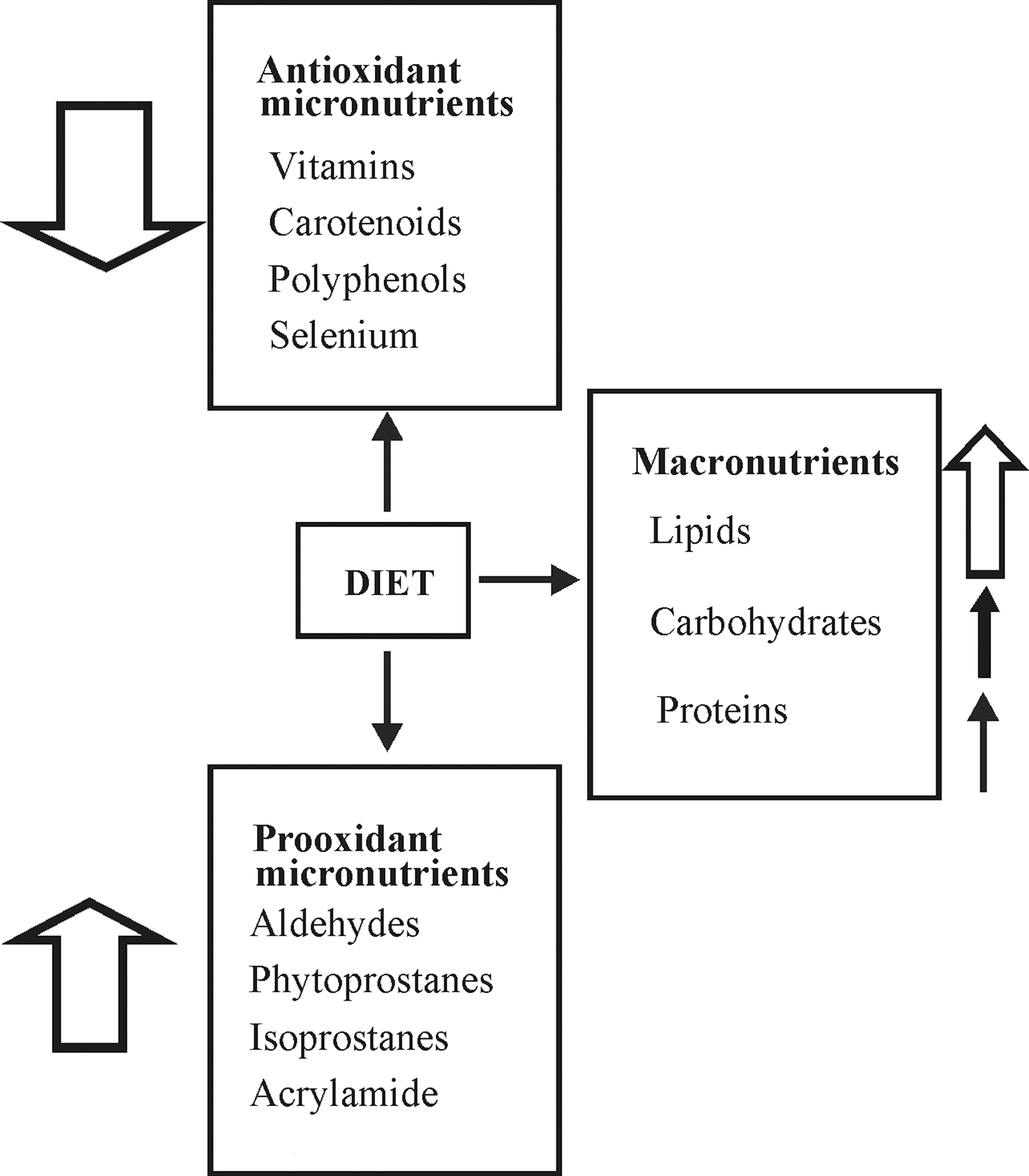
A. Low-molecular-weight dietary oxidants and antioxidants
The majority of the dietary oxidants are often already intermediates of the prestarted radical reactions. These reactions can be initiated already in the food source and aggravated by storage time or preparation conditions. Virtually, every food, whether it is plant-derived or animal-derived, contains such prooxidants (e.g., ROS themselves or aldehydes). In plants, generation of initial ROS can cause oxidation of linolenate (a prominent component of the plant cell membrane), which is a starting point for producing G1-phytoprostanes and MDA, while in animals, a similar reaction sequence forms isoprostanes from arachidonate (251). In addition, carbohydrate-rich food processed at high temperature (for instance, French fries or potato chips) are usually containing different amounts of acrylamide, which can act as a prooxidant by depleting GSH (146). Commonly, a number of beverages are also considered as major source of prooxidants; for example, tea and coffee could generate H2O2 to achieve over 100 μM concentrations (215).
Although the major human intervention trials failed to confirm the proposed roles of the traditional dietary antioxidants (see section VI), it is worth remembering that there are thousands of plant-derived bioactive compounds that are potentially antioxidants, whereas only half a dozen was tested in those trials. During evolution, plants gained the possibility to generate an enormous number of antioxidant molecules to confront the oxidative challenge they encounter upon photoreception. Thus, plant-derived food is generally considered to be rich in antioxidants. Their amount depends on the plant species, life cycle of the plant, postharvest technologies, and soil type (358). Depending on the method used, the potential antioxidant capacity of different fruits and vegetables varies. However, there is a consensus that cruciferous vegetables, legumes (especially soy origin), sweet potatoes, and tomatoes are some of the most antioxidant-rich vegetables. Berries and many other fruits, for example, apple, citrus, and grapes, hold also strong positions in terms of their antioxidant capacity (98, 291). Due to the tremendous varieties in fruits and vegetables worldwide, the World Cancer Research Fund and the American Institute of Cancer Research recommend serving several different fruits and vegetables instead of only one (115). Although animal-derived food is usually not considered to be rich in antioxidants, their role in the antioxidant system is not negligible. For example, GSH is found in fresh meat (330).
B. Macronutrients and ROS generation/accumulation
The macronutrients stimulate ROS production by increasing the postprandial metabolic rate, especially at the mitochondrial level. Acetyl CoA, which is formed by oxidative conversion of pyruvate (generated from glucose during glucolysis), as well as after β-oxidation of fatty acids and metabolism of several glucogenic amino acids, is processed in mitochondria via the tricarboxylic acid cycle, leading to the generation of NADH and FADH2 (Fig. 15). NADH and FADH2 donate electrons to complex I and II, respectively, of the mitochondrial ETC, while leakage of these electrons to oxygen is the main source of O2 •− formation (263). After ingestion of macronutrients, processing of substrates through the ETC is accelerated, and in the absence of sufficient physical activity (i.e., without increased demand for ATP), the mitochondrial membrane potential would be getting more and more reduced, resulting in higher ROS generation (60, 97, 263). Correspondingly, immediately after ingestion of a meal, ROS concentrations in peripheral cells are markedly increased (245, 269). Increased production of ROS would eventually strengthen the existing ROS-mediated activation of various signaling pathways and transcription factors, some of which (i.e., NF-κB, AP-1, and early growth response protein-1 [Egr-1]) promote additional ROS generation (see section IV.B) (10). For instance, ROS-dependent activation of PKC favors NADPH oxidase overexpression (99).
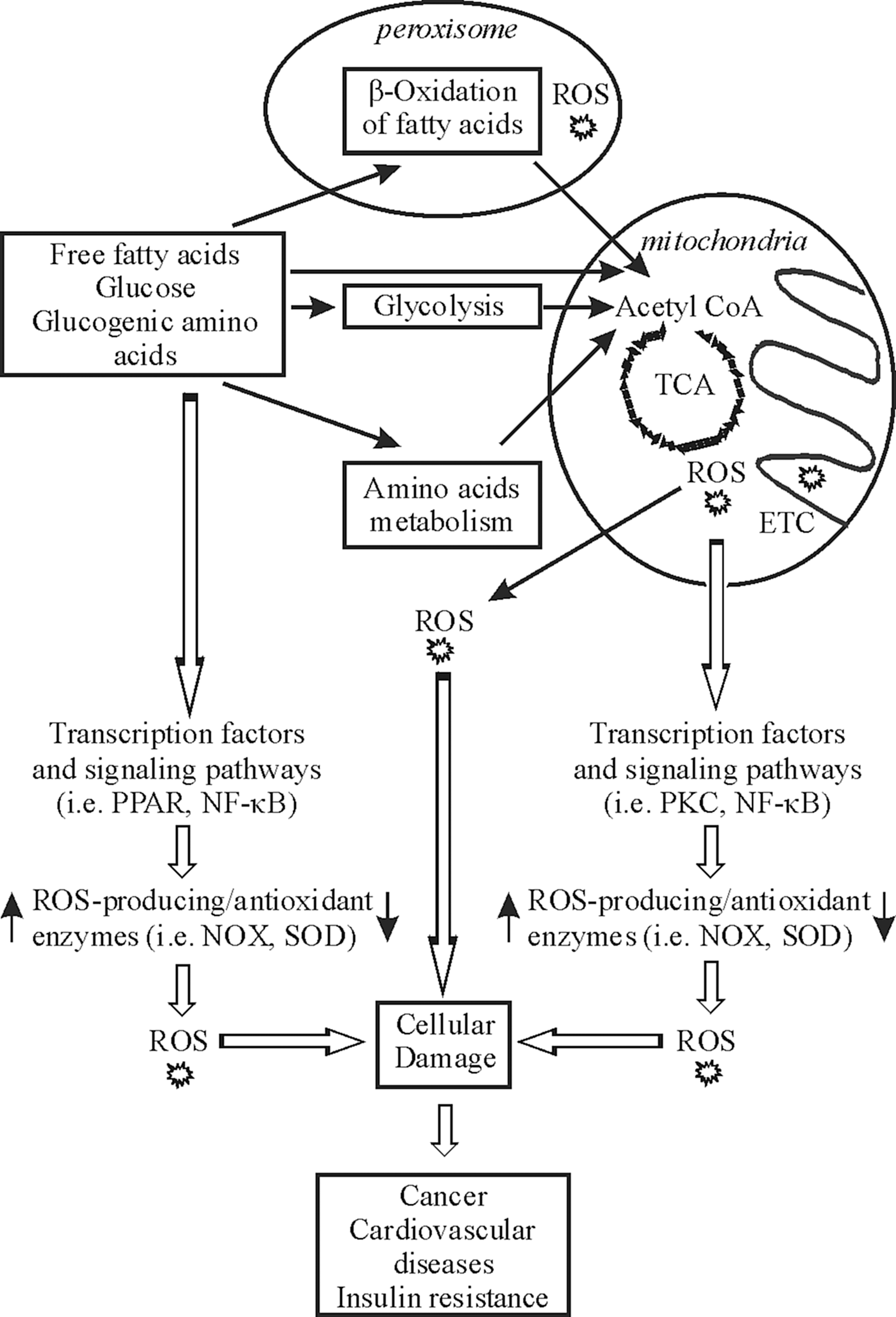
1. Carbohydrate-dependent ROS generation
A consistent increase of various oxidative stress biomarkers was shown after an oral glucose tolerance test (OGTT) as well as after the ingestion of carbohydrate-rich meals in diabetic patients (97). With respect to a healthy population, many, but not all, investigators reported an increase in selected biomarkers of oxidative stress in response to an OGTT and carbohydrate-rich meals (97). Glucose challenge stimulates ROS generation by polymorphonuclear leukocytes and mononuclear cells and causes fall in the α-tocopherol concentration 2 h after ingestion (246). In line, ROS levels were low in polymorphonuclear leukocytes and mononuclear cells of healthy subjects after a 48-h fast (77). In conformity, the glycemic index of carbohydrates is found to be correlated with oxidative stress (269), and an excess intake of highly processed carbohydrates of a high glycemic index (e.g., white bread) results in higher postprandial ROS generation, while leafy vegetables, fruits, nuts, seeds, or unprocessed grain results in lesser glucose excursion and ROS generation (269). Interestingly, glucose was found to upregulate NOX2 activity in polymorphonuclear leukocytes and mononuclear cells in vivo (245).
2. Protein-dependent ROS generation
Protein intake was also found to increase O2 −• production in polymorphonuclear leukocytes and mononuclear cells 1 h after a meal (245). Dietary protein restriction was shown to reduce ROS generation at complex I of the mitochondrial ETC, implicating that proteins, like glucose, are potent dietary macronutrients in terms of ROS production in mitochondria (216). Apparently, protein restriction has also been reported to reduce ROS generation in several rodent in vivo studies (216). Interestingly, chronic restriction of methionine alone was found to reduce mtROS in rats, and a paradoxical increase in GSH was observed, indicating a major metabolic shift toward GSH production with the few available methionine molecules (216, 303). On the other hand, the data concerning protein-dependent postprandial ROS generation are less abundant as compared to glucose intake, and one study did not find a significant increase in oxidative stress after protein intake by young healthy men (97).
3. Lipid-dependent ROS generation
In contrast to glucose and proteins, lipids provide a major platform for ROS generation and accumulation in the plasma and ER membrane. It was shown in a number of studies that various biomarkers of oxidative stress increased in healthy as well as diseased individuals in response to lipid-rich meals (sometimes containing small-to-moderate amounts of carbohydrate and protein), with maximal values observed 2–4 h after feeding (97). It was suggested that ingestion of lipids (which produce a significantly greater amount of FADH2 than carbohydrate and protein, thus leading to leakage-susceptible reverse electron flow from mitochondrial complex II to complex I) would promote exacerbated ROS production compared to other macronutrients (97). In human prostate cancer cells, androgens increase the mitochondrial oxidation of fatty acids, leading to increased production of ROS (208). The fatty acids show differential features of oxidizability according to their type. Saturated fatty acids normally provide more protection against ROS than PUFA. The composition of different PUFAs vastly depends on the dietary intake of ω-3 and ω-6 PUFAs. The susceptibility of fatty acids to oxidation is often thought to be directly dependent on their degree of unsaturation, but some studies suggest that the relation between the chemical structure and susceptibility to oxidation is not as straightforward as hypothesized from theoretical viewpoints (301). Indeed, the ω-3 group, especially eicosapentanoic acid (EPA) and docosahexaenoic acid (DHA), provides better protection against oxidative damage than the ω-6 group by being less susceptible to oxidation (231, 301, 341). Interestingly, DHA has been found not only to lower blood pressure and heart rate but also to inhibit the NOX2, which is a major cellular ROS generator in both phagocytic and nonphagocytic cells (225). Thus, a critical balance between EPA and DHA is highly recommended for more protection, and this can primarily be achieved by a selective dietary intake.
4. Indirect stimulation of ROS production by macronutrients
Beside the direct immediate effect on ROS production, mostly via the function of mitochondrial ETC, fatty acids and carbohydrates may have long-term indirect effects via regulation of expression of either ROS-generating or antioxidant enzymes. There are at least three transcription factor families, peroxisome proliferator-activated receptors (PPAR), liver X receptors, and sterol response element-binding proteins, which play a major role in the fatty acid-induced regulation of gene transcription (360). The classic example of a transcription factor, which can be directly activated by fatty acids, is PPAR (360). PPAR is a nuclear hormone receptor transcription factor that upon activation binds to specific motifs within the promoters of the corresponding genes. PPARγ was found to have an antioxidant effect by altering the expression of enzymes that participate in the production or elimination of ROS, that is, Cu/ZnSOD and NADPH oxidase (153, 158). In a murine model of type 2 diabetes, PPARγ activation exerts an antioxidant effect in vivo (23). In another study, the decreased expression of PPARγ in peripheral blood mononuclear cells in morbidly obese persons with insulin resistance after a high-fat meal was associated with the decrease of SOD and GR activities and the increase in the concentration of free fatty acids (111). Although the molecular basis of the association between PPARγ and SOD has not been determined, the presence of a functional PPAR-response element in the Cu/ZnSOD promoter suggests that Cu/ZnSOD expression may be directly regulated by PPARγ activation (392).
Additionally, fatty acids and glucose have the ability to alter a number of pathways leading to activation and nuclear translocation of the downstream transcription factors such as AP-1 and NF-κB (73, 360). Among target genes of AP-1 and NF-κB are antioxidant as well as ROS-generating enzymes. Therefore, NF-κB can, on the one hand, be induced by a macronutrient-dependent burst in ROS generation, and, on the other hand, be induced by macronutrients indirectly via regulation of classical intracellular signaling pathways such as PKC. While the immediate action of macronutrients increases ROS production, their long-term effects on regulation of transcription factors may cause not only induction of ROS generation but also increased ROS scavenging.
5. Generation of ROS upon deficiency of macronutrients
It has been shown that nutrient starvation increases ROS, especially intracellular levels of H2O2 (316). Another study demonstrated that O2
−• formation was selectively induced by starvation of glucose,
VIII. Antioxidant Supplementation and Oxidative Stress: A Complicated Riddle
The affiliation of ROS in tumorigenesis was widely reported for more than three decades and almost universally accepted (124, 226, 371, 398). This led to the establishment of antioxidants as chemopreventive agents against cancer, and it was believed that tumorigenesis can be prevented by a dint of antioxidant supplementation. Primary observations were so compelling that an antioxidant hype emerged throughout the world, and high doses of antioxidant supplements were investigated in hundreds of chemoprevention studies. About a dozen of these studies were huge randomized trials with various combinations of the most prominent antioxidants (so-called traditional antioxidants) (Table 3) (119). So far, more than 10 large-scale trials have been accomplished (119). Except polyphenols, almost all the traditional antioxidants have been involved in the accomplished trials in various combinations and various doses. The results obtained were enigmatic, showing, in general, no corroboration of the current knowledge and of the data derived from nonhuman models as well as from observational epidemiologic studies. Meta-analyses of randomized controlled trials, including tens of thousands subjects, found no overall association between the consumption of antioxidant supplements and cancer risk (255). The situation is aptly implicated as the antioxidant paradox (131). Interestingly, the paradox is not only limited to the field of oncology, rather it is also reported for other diseases, for example, diabetes and cardiovascular diseases (147, 328).
A. Type, dose, combination, and duration of exposure to antioxidants determine their effects
The reasons for the lack of protective effects of antioxidants in human trials are not known. Among the possible explanations are the wrong type, dose, combination, and/or duration of exposure. Indeed, in almost all of the large-scale trials with antioxidants, a significantly high dose of each antioxidant was applied either alone or in limited combinations. The question in these studies was only whether a specific amount and combination of antioxidants can prevent tumorigenesis in a specific randomized population, but mechanisms of action have drawn less attention. In this respect, it should be remembered that some antioxidants, including vitamin E, vitamin C, and quercetin, can have prooxidative properties at high concentrations. Meta-analysis of the dose–response relationship between vitamin E supplementation and total mortality indicates that administration of α-tocopherol at high doses (≥400 IU/day) may be associated with increased mortality rate, an observation attributed to the prooxidant effects of α-tocopherol at these concentrations (239). It was also shown that β-carotene acts as a prooxidant in the presence of chronic oxidative stress such as that resulting from smoking (149, 229). In line, quercetin at concentrations in the range of 1–40 μM reacts with ROS and chelates metal ions, whereas at concentrations higher than 40 μM, it increases oxidative stress (362).
Moreover, one antioxidant component can stipulate another's presence or even block its action. For example, supplementation of mildly hypercholesterolemic subjects with vitamin E lowered the plasma levels of the antioxidant ubiquinol-10 (172). On the other hand, the beneficial effects of multiple dietary antioxidants combined may be greater than the effect of each of those compounds examined individually (119). In particular, the natural combination of phytochemicals and vitamins in fruits and vegetables is believed to be responsible for their potent antioxidant activity (213). For example, it was calculated that vitamin C in apples contributes only to about 0.4% of their TAC (92).
Since neoplastic lesions are multistage processes and may take several decades to become clinically detectable, the duration of antioxidant therapy in human trials, though always at least several years, could have been insufficient for evaluating their efficiency in preventing cancer. Indeed, in colorectal cancer prevention study performed by the American Cancer Society that lasted for 5 years, it was found that a regular multivitamin use at enrollment was not associated with the risk of colorectal cancer, whereas regular multivitamin use 10 years before enrollment was associated with a reduced risk, no matter whether participants still used multivitamins at enrollment or had stopped (162). These results are consistent with the hypothesis that a past, but not recent, vitamin use may be associated with a reduced risk of colorectal cancer, though it should be taken into account that multivitamin users were more likely than nonusers to practice health conscious behaviors in general, including exercising more (162). The effects of a recent antioxidant use may only be detectable at a subclinical level, whereas a reduction in the risk of clinically detectable disease may require longer exposures. Therefore, a useful alternative to the clinical endpoints is the use of carcinogenesis biomarkers, cell cycle markers, and markers of oxidative stress/inflammation (119). In line, one recent study showed that an antioxidant micronutrient cocktail (800 mg
B. The sensitivity to the antioxidant action has individual variations
Not only the dose, duration, and/or the way of antioxidant administration, but also individual nutritional as well as the health status of participants can strongly influence the outcome of trials. The Linxian study that corroborated to the protective role of a combination of antioxidants was carried out with a severely undernourished population. On the other hand, many enrolled subjects in the Western countries consumed antioxidants independently of participation in trials. The lack of effects of antioxidants in the Italian population may be explained by the simple suggestion that Italians already receive the benefits of a Mediterranean diet rich in phenolic antioxidants; thus, this population may be selected not to respond in a beneficial way to antioxidant supplements (131). In general, there are no large clinical trials in which only patients that demonstrate elevated ROS levels were recruited, though only these subjects are expected to benefit from the action of antioxidants. In some particular subgroups of patients, the positive effect of antioxidants was indeed shown. For example, the combination of γ-tocopherol and vitamin C significantly reduced the progression of atherosclerosis in hypercholesterolemic smoking men who had low baseline serum vitamin E levels (310).
Single-nucleotide polymorphisms in genes encoding important enzymes regulating the oxidative status of a cell can also contribute to enhanced oxidative stress in some individuals. For instance, a genetic polymorphism in the MnSOD gene can potentially cause impaired protein targeting and is found to be associated with prostate (203), breast (12), and ovarian (270) cancer. Similarly, differences in GST P activity caused by a genetic polymorphism have been found to correlate with oxidative DNA damage and breast cancer (227). Interestingly, various studies have revealed that genetic variations in several of the antioxidant-related genes are very common throughout the world and may vary among different ethnic and racial communities (1, 31, 135, 188). Since antioxidant supplementation trials did not take into account the single-nucleotide polymorphisms within the genes for the antioxidant enzymes in the corresponding populations, this could be one of the explanations of these trials' contradictory results.
C. The effects upon ROS removal differ depending on their type, concentration, and intracellular localization
The failure of antioxidant trials can be also viewed as evidence against reductionism in science, especially in such a multifaceted connection between ROS and cancer. The research of the last decade indicated that there is a tremendously wide array of mechanisms through which ROS could contribute to tumorigenesis. While direct damage of DNA molecules by ROS and induction of epigenetic changes can contribute to a genetic program that is favorable for tumorigenesis, it became also clear that the effect of ROS often depends on the stage of tumorigenesis. In this regard, it should be noticed that several studies involved smokers, a portion of whom could have suffered from the subclinical stage of cancer. Since many antitumor agents act by inducing ROS production in cancer cells, supplementation of smokers with high doses of antioxidants could be harmful, and it may partially explain the adverse effects observed in ATBC or CARET. Further, the protective or tumorigenic effects could also depend on the type of ROS involved. For example, O2 −• and H2O2 downregulated caveolin-1 expression and inhibited cell migration and invasion, whereas •OH upregulated caveolin-1 expression and promoted cell migration and invasion of the human lung carcinoma H460 cells (218).
Increasing evidence suggests that a number of antioxidant compounds, which were previously thought to have antitumor properties due to their ROS scavenging activity, are more likely to work through cell-to-cell signaling, inflammatory enzyme activity, or gene regulation (252, 261, 299, 368). New data also indicate that some substances that were thought to be carcinogenic due to the stimulation of oxidative stress may indeed act via other mechanisms (13). For example, the prolonged administration of peroxisome proliferators, which include some hypolipidemic drugs (clofibrate), herbicides, plasticizers, and leukotriene antagonists, leads to development of hepatic carcinomas in rodents. Peroxisome proliferators sustain a PPARα-dependent activation of the H2O2-generating enzymes, aldehyde oxidase-1 and cytochrome P450 4A, and a decline in the activity of H2O2-degrading catalase and GPX, leading to the accumulation of the oxidative stress marker lipofuscin and the formation of 8-oxodG–DNA adducts in liver cells (178, 292). However, the current view favors the idea that hepatic carcinogenesis promoted by peroxisome proliferators is not only due to hepatic oxidative stress but also related to the abnormalities in cell growth and communication (13, 59).
It is also possible that orally administered antioxidants may be inaccessible to the source of free radicals in various cellular compartments. For most cell types, antioxidant molecules to be effective should be specifically localized either in the plasma membrane, cytoplasm, mitochondria, peroxisomes, nuclei, or other organelles (see sections II and V.A). For example, water-soluble molecules, such as vitamin C, are radical scavenging agents in the cytoplasm, whereas lipid-soluble molecules, such as vitamin E, β-carotene, and polyphenols, appear to act primarily as antioxidants within the lipid environments. The slow rate of tocopherol movement between the bilayer leaflets could imply that in the absence of an active uptake mechanism, exogenously applied tocopherols stay in the outer leaflet of the cell membranes (18). Therefore, lipid-soluble antioxidants have a low ability to scavenge ROS produced in the cytoplasm and various intracellular organelles (144). Although vitamin C has better access to the intracellular compartments as compared to lipid-soluble vitamins, it has a narrow absorption window with a maximum at 200 mg, and therefore an increase in the orally administered dose will not increase its bioavailability (235).
Resveratrol can behave either as an antioxidant or as a prooxidant depending on the selective intracellular microenvironment. As an antioxidant, resveratrol, in particular, decreases mitochondrial production of O2 •− in various cell types (355). It was recently shown that the prooxidant activity of resveratrol is dependent on its interaction with chromatin-bound copper that leads to ROS generation through Fenton-type reactions with subsequent oxidative DNA breakage and apoptosis (252). Such resveratrol-induced DNA damage is enhanced in low pH environments characteristically observed in tumors due to the Warburg effect. Wrong intracellular localization of antioxidants can be also harmful (131). For example, correct disulfide-bridge formation and folding of proteins in the ER are aided by having an environment more oxidized than the rest of the cell; therefore, an increase of antioxidants in the ER may disrupt the localized oxidation necessary for its physiological function (121). Thus, when using antioxidants, their different action in various cellular compartments as well as their influence on different signaling cascades needs to be considered.
D. Diet, oxidative stress, and cancer
Still, the protective effects of antioxidant-rich food with respect to tumorigenesis are widely reported (2, 183, 394). A selected dietary fashion can result in less ROS generation and provide us with less vulnerability to oxidative stress. These beneficial effects could be achieved primarily by dietary intervention, but not by the micronutrient supplementation (213). Dietary intake does not involve pharmacologic doses of a single constituent and is unlikely to elicit adverse effects such as a high-dose supplementation of various nutrients does. Most importantly, the form of a specific antioxidant present in normal diet could be a vital factor that specifically contributes to its function, and might be overlooked in supplementation studies where the source and form vary from study to study. In line, while pharmacological doses of single antioxidants have demonstrated harmful effects in smokers, two food-based antioxidant-rich diets were found to be safe in a randomized controlled trial in male smokers (176). As a new tool for investigating the relationship between diet and cancer risk in population studies, the measurement of plasma TAC has been proposed (325). A comprehensive food database consisting of the TAC for typical foods as well as other dietary items such as traditional medicine plants, herbs, and spices (more than 3100 in general) was recently published and can be used in future investigations on diet and cancer (55). Recent data of the European Prospective Investigation into Cancer (EPIC) study, including a total of 521,457 subjects, indicate that a high intake of antioxidant compounds from different sources of plant foods as measured by TAC, assessed by TRAP and FRAP assays, is associated with a reduction in the risk of gastric cancer (326). This promising result highlights the importance of synergistic interactions between plant food components as compared to the use of single antioxidants.
As an example, we would shortly discuss the diets of two healthiest communities of the world: the Okinawans and the Mediterranean basin dwellers (39, 378). The Okinawan diet is in principle a low caloric one with the total daily caloric intake as low as 1100 kcal. In a typical Okinawan diet, the carbohydrate intake accounts for 85% of total calorie, while protein and fat intake are 9% and 6%, respectively (378). Another feature of traditional Okinawan diet is the high amount of fruits and vegetables in their cuisine, which are highly rich in antioxidants, for example, sweet potato, legumes, soy products, bitter melon, and seaweeds (378). Fish holds a moderate position in the traditional Okinawan diet, and meat consumption is significantly lower than in the rest of Japan. Clearly, the traditional Okinawan diet promises lesser threat of oxidative stress and oxidative stress-induced tumorigenesis. However, dietary changes were observed in the traditional Okinawan diet for some decades associated with a marked increase in the mortality rate due to cancer (243, 378).
The Cretan diet represents the traditional Mediterranean diet of the 1960s, and the population consumes higher amount of fat (37% of total energy) than the Okinawan population, which is actually striking, given their second lowest rate of incidence of cancer or cardiovascular disease just behind the Okinawans (330). However, it seems to be likely the fatty acid composition holds the key. A large portion of fat in the Cretan diet comes from olive oil, fish, and chicken. Olive oil is rich in monounsaturated fatty acid (MUFA) instead of PUFA; fish oil is one of the higher sources of ω-3 fatty acids, and the chicken contains more ω-3 fatty acids because of being grass- and worm-fed in Crete (330). As a result, the fatty acids are mainly MUFA, and due to this, the ω-3/ω-6 fatty acid ratio is extremely high (329). Another feature of the Cretan diet is the regular consumption of a high amount of fruits considered to be rich in antioxidants. Unlike in the Okinawan population, wine consumption is a common dietary pattern in Crete, which is thought to provide them with resveratrol. Although the beneficial effect of resveratrol is not yet well proven, it is extensively reported to be a potential antioxidant and antitumorigenic phytochemical (see section VI.D) (370).
However, it is also not always clear to what extent the dietary fashion could provide antioxidant support. Antioxidant molecules' absorption, bioavailability, and function are found to be highly dependent on the food microstructure. Several food components can be responsible for limited bioaccessibility and bioavailability of antioxidants (275). It is also imperative to unravel whether any food component either directly or indirectly promotes antioxidant absorption that might present a stumbling block to antioxidant supplementation.
IX. Conclusion
The menace of reactive derivatives of oxygen probably has been present since the very beginning of evolution of aerobic cells. Concomitantly, aerobic cells were also evolved with a regulated system that could neutralize the threat to an effective extent. The regulation must be tight enough to curb oxidative stress, but slack enough to condone basal generation and accumulation of ROS to fulfill their physiological functions. Human body possesses an antioxidant system where some components are synthesized de novo, while others are meant to be obtained through diet, and the absorption of such components is also evolutionarily selected. Our current genome profile is believed to be established some 200 thousand years ago, and the dietary fashion and lifestyle seem to have highly influenced our current genetic profile at that time. The last 10,000 years there has not been any significant change in the genomic profile, considering that the spontaneous mutation rate is ∼0.5% per million years (329). By contrast, especially the last 60 years have brought about a lot of changes in the food amount and composition with PUFAs, excess proteins and cereals dominating our diet. While the caloric intake increased dramatically with a normal Western diet, energy expenditure has been reduced. Chronic alcoholism and tobacco smoking increased severely keeping pace with the ever-worsening environmental toxicity. Thus, our established antioxidant capability is confronted with more difficulties than it is supposed to encounter, underpinning, at least in part, increased occurrence of several chronic diseases such as cancer. Although the results from the large-scale studies of direct nutritional supplementation with particular antioxidants with respect to cancer are heterogeneous, the risk for cancer might be prevented or delayed by using a selective, recommended dietary fashion that considers a balanced energy intake and expenditure, as well as individual genetic predispositions, preferentially from an early stage of life. In addition, the little evidence from the large-scale studies creates the demand to further increase our knowledge about the interconnection between nutrition, lifestyle, and the dynamics of ROS/antioxidant action, as well as to improve and to standardize the techniques measuring reliable biomarkers.
Footnotes
Acknowledgments
Work of the authors is supported by grants from the Biocenter Oulu, the Academy of Finland, Sigrid Juselius Foundation, and Nordic Center of Excellence MitoHealth programme on Food, Nutrition, and Health.