
Abstract
Aims:
Acetaminophen (APAP)-induced liver injury is mainly due to the excessive formation of reactive oxygen species (ROS) and reactive nitrogen species (RNS) through the formation of a reactive intermediate, N-acetyl-p-benzoquinone imine (NAPQI), in both humans and rodents. Here, we show that the indole-derived synthetic compound has a protective effect against APAP-induced liver injury in C57Bl/6 mice model.
Conclusion:
Our data suggest that the indole-derivative, NecroX-7, directly binds to NAPQI when hepatic GSH levels are very low and the NAPQI–NecroX-7 complex is secreted to the blood from the liver. NecroX-7 shows more preventive and similar therapeutic effects against APAP-induced liver injury when compared to the effect of N-acetylcysteine in C57Bl/6 mice. Antioxid. Redox Signal. 18, 1713–1722.
Introduction
Innovation
NecroX-7 is an indole-derived potent antioxidant molecule, which can be bound to other types of reactive molecules such as DPPH, ABTS, and especially NAPQI. It is well known that NAPQI is a major intermediate of APAP, which causes necrosis of hepatocytes in rodent and human. Thus, blocking NAPQI formation or eliminating NAPQI are novel strategies for the treatment or prevention of APAP-induced liver injury instead of GSH replenishment.
Since 1974, N-acetylcysteine (NAC) has been used as a rescue medication for APAP poisoning by providing cysteine for GSH synthesis (5, 26). NAC is a comparatively safe drug for APAP-induced liver injury (AILI) and is currently being used to treat acute viral hepatitis and other acute hepatic failures (10). However, some studies show that a prolonged treatment with a high dose of NAC (600 to 1200 mg/kg per day) interferes with the normal hepatic mitochondrial metabolic processes and impairs liver regeneration after AILI by interrupting nuclear factor (NF)-κB signal pathways (22, 34, 37). Although there have been numerous studies on the detailed mechanisms of action of the effect of NAC on APAP toxicity, a new generation of drugs with improved efficacy and decreased interruption of normal hepatic function should be developed.
Indole-derived chemicals, such as melatonin and its derivatives, showed in vitro antioxidant activity and ameliorated oxidative stress-related organ damage in neuron, kidney, and liver (27, 32). In the case of melatonin, it protected mice from the APAP toxicity without disturbing the antipyretic and anti-analgesic effects of APAP (14). Matsura and his colleagues suggested that melatonin could act downstream of NAPQI formation and upstream of GSH conjugation to NAPQI. They concluded that melatonin protected APAP-induced liver injury probably via its antinitrosative, antioxidative, and anti-inflammatory activities (21). Taken together, indole derivatives might have a potential to treat AILI, and more clear action mechanisms for AILI are needed.
NecroX compounds are indole-derived small molecules which showed antioxidative and antinitrosative effects in H9C2 cardiomyoblasts, and a protective effect against ischemia-reperfusion injury in beagle dogs (4, 15). NecroX-7 was recently developed as a potent antioxidant molecule based on the indole moiety. There are small differences in the functional group of the indole backbone, and that change of functional group showed different cell permeability and efficacy to the radical reactivity. It was demonstrated that NecroX-7 can prevent cells from oxidative stress-related necrotic cell death (4, 15, 24). However, little is known about the action mechanisms of NecroX-7 on specific types of necrotic cell death. In this study, we tested NecroX-7 on AILI in mice because ROS and RNS formation by an excessive APAP consumption is the main cause of hepatic necrosis. We found that the binding of NecroX-7 to NAPQI can attenuate APAP-induced hepatic injury in C57Bl/6 mice.
Results
NecroX-7 protects primary hepatocytes from the APAP-induced cell death and ROS/RNS production
APAP toxicity is due mainly to the production of ROS and RNS, thus we tested a group of indole-containing small molecules that display antioxidant and antinitrosative effects. To validate candidates for AILI treatment, the compounds were screened against tert-butylhydroperoxide (t-BHP) (Supplementary Table S1; supplementary data are available online at
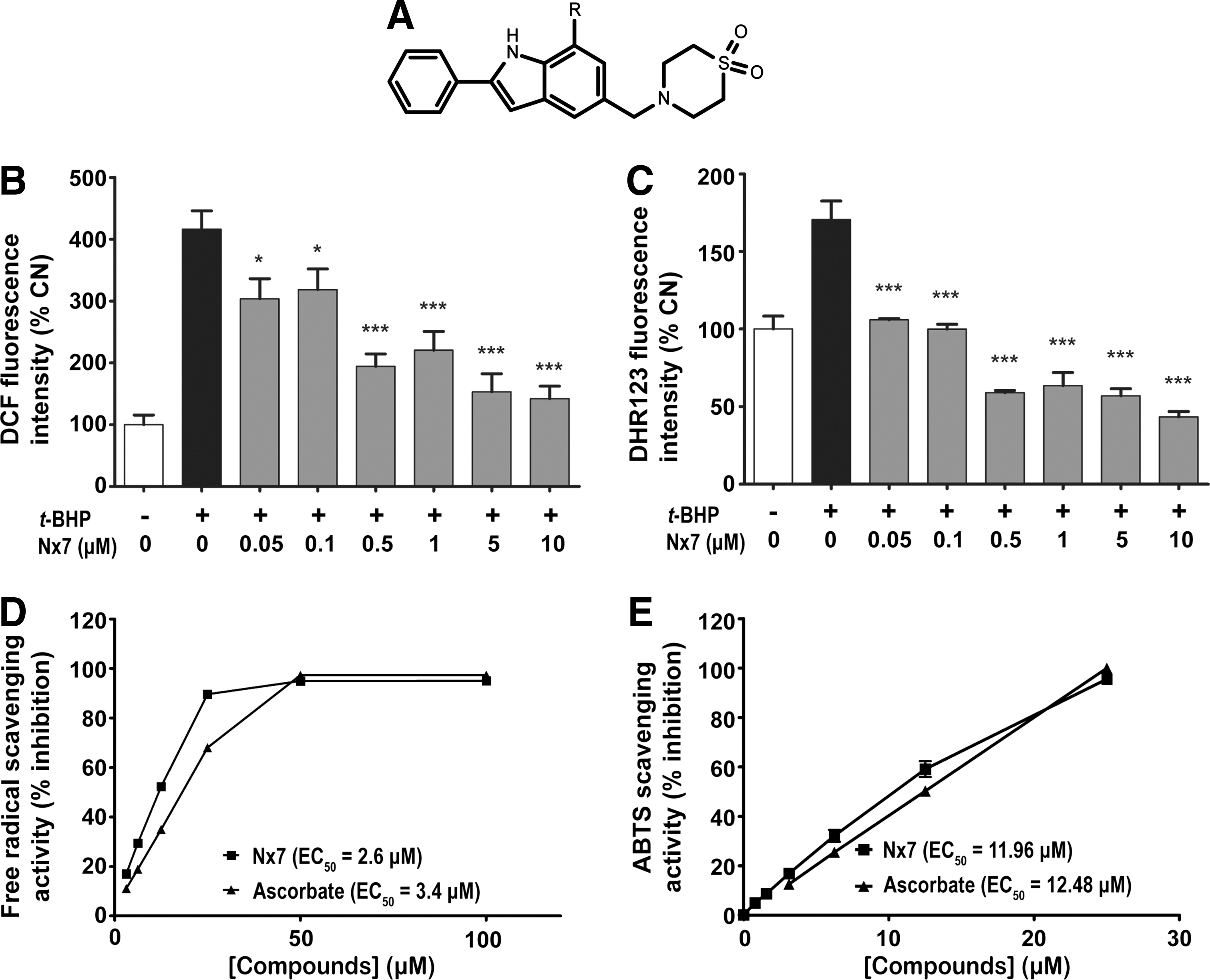
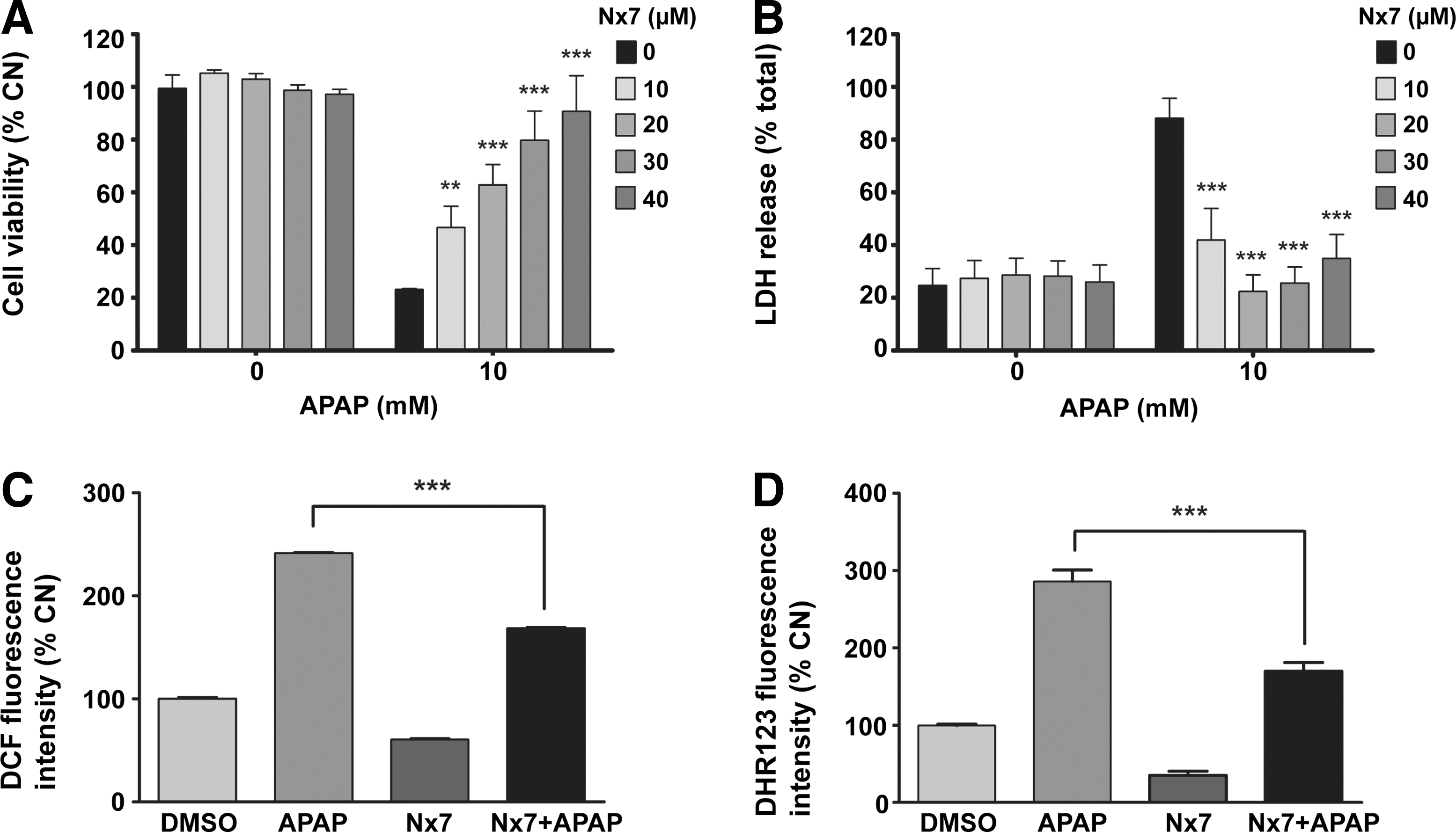
To determine whether NecroX-7 can be used for the treatment of AILI, the levels of APAP-induced cell death and ROS/RNS production were measured in primary hepatocytes isolated from C57BL/6 mice. NecroX-7 improved the viability of hepatocytes (Fig. 2A) and decreased the release of lactate dehydrogenase (LDH) in a dose dependent manner (Fig. 2B). At concentrations higher than 10 μM, NecroX-7 significantly prevented APAP-induced cell death in hepatocytes. Moreover, it was discovered that 25 μM NecroX-7 significantly reduced the DCF-DA (Fig. 2C) and DHR123 (Fig. 2D) fluorescent intensity induced by APAP.
NecroX-7 directly binds to NAPQI in the liver and the complex is secreted into the blood
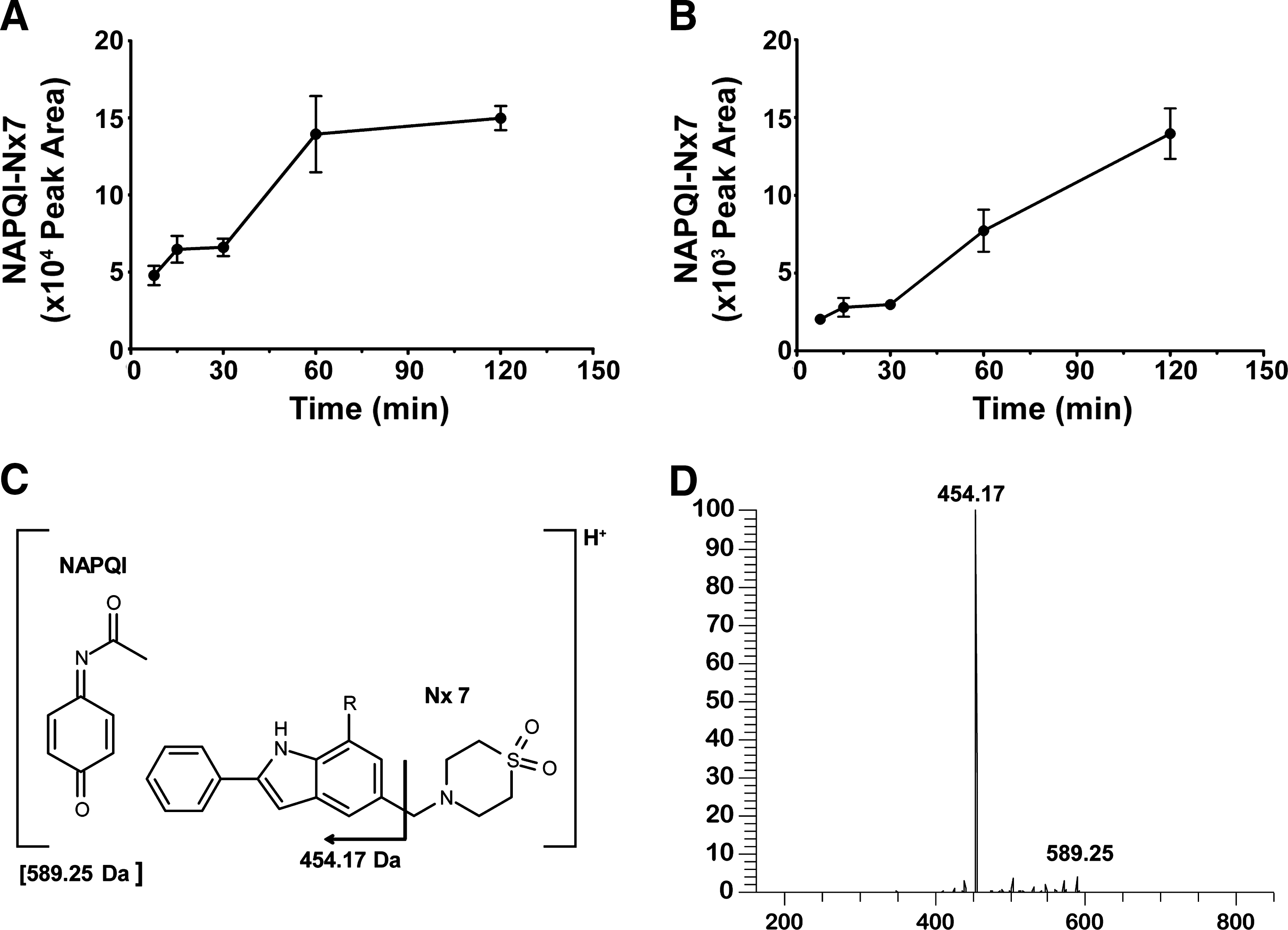
We hypothesized that NecroX-7 might bind to NAPQI, the reactive metabolite of APAP, because of the ROS/RNS scavenging activity of NecroX-7 and its ability to bind directly some radicals. To test our hypothesis, we analyzed liver tissues and plasma samples after NecroX-7 and APAP injection, using liquid chromatography-mass spectrophotometry (LC-MS/MS). We found four different complexes with a same molecular mass of 589.25 Da in the liver (Fig. 3A) and plasma samples (Fig. 3B) in different amounts. The NAPQI–NecroX-7 complex was increased both in liver and blood in a time-dependent manner. The major conjugation sites are expected to be the phenyl moiety of NAPQI and the indole ring of NecroX-7 (Fig. 3C). In addition, the simple mixing of NecroX-7 with NAPQI also generated the same conjugated complexes as those generated in vivo in the absence of enzymes or liver homogenates (data not shown). This indicates that NecroX-7 binds to NAPQI directly without the requirement of cofactors or proteins. The identity of the complexes was confirmed by high-resolution LC-MS analysis and MS/MS analysis of the protonated complex molecules (Fig. 3D). All four of the complexes identified by LC-MS exhibited an [M+H]+ ion mass matching the theoretical mass of 589.25 (C32H37N4O5S) and the same characteristic fragment ion at 454.17 Da upon collision-induced dissociation, corresponding to the loss of thiomorpholine dioxide.
NecroX-7 does not disturb the metabolic process of APAP
We next checked the APAP metabolites to determine whether NecroX-7 affects normal APAP metabolism due to the direct binding with NAPQI. To identify the effect of NecroX-7 on the metabolism of APAP, the levels of APAP metabolites, such as APAP-glucuronide, APAP-sulfate, and APAP-cysteine, were determined. There were no significant changes in the time-dependent profiles of these metabolites despite of the NecroX-7 treatment (Figs. 4 and 5). These water-soluble products are mainly formed in the liver and then excreted to the urine after going through bile and blood (2). Hepatic GSH level was depleted 30 min after APAP treatment in both groups (Fig. 4D). Decrease of GSH in NecroX-7-treated mice reflects that NecroX-7 may have an action after GSH depletion. We also examined the NADPH/NADP+ ratio, which is indicative of the activation of CYP2E1 because the reduction of APAP to NAPQI is a NADPH-consuming process (2). There was no significant change in the NADPH/NADP+ ratio after treatment with NecroX-7 (Supplementary Fig. S1). In addition, NecroX-7 did not inhibit CYP2E1 enzymatic activity which was performed with mouse liver microsomal protein (Supplementary Fig. S2).These results suggest that NecroX-7 does not disrupt APAP metabolism, but inhibits the action of NAPQI by direct binding.


NecroX-7 protects mice from the APAP-induced hepatotoxicity and lethality
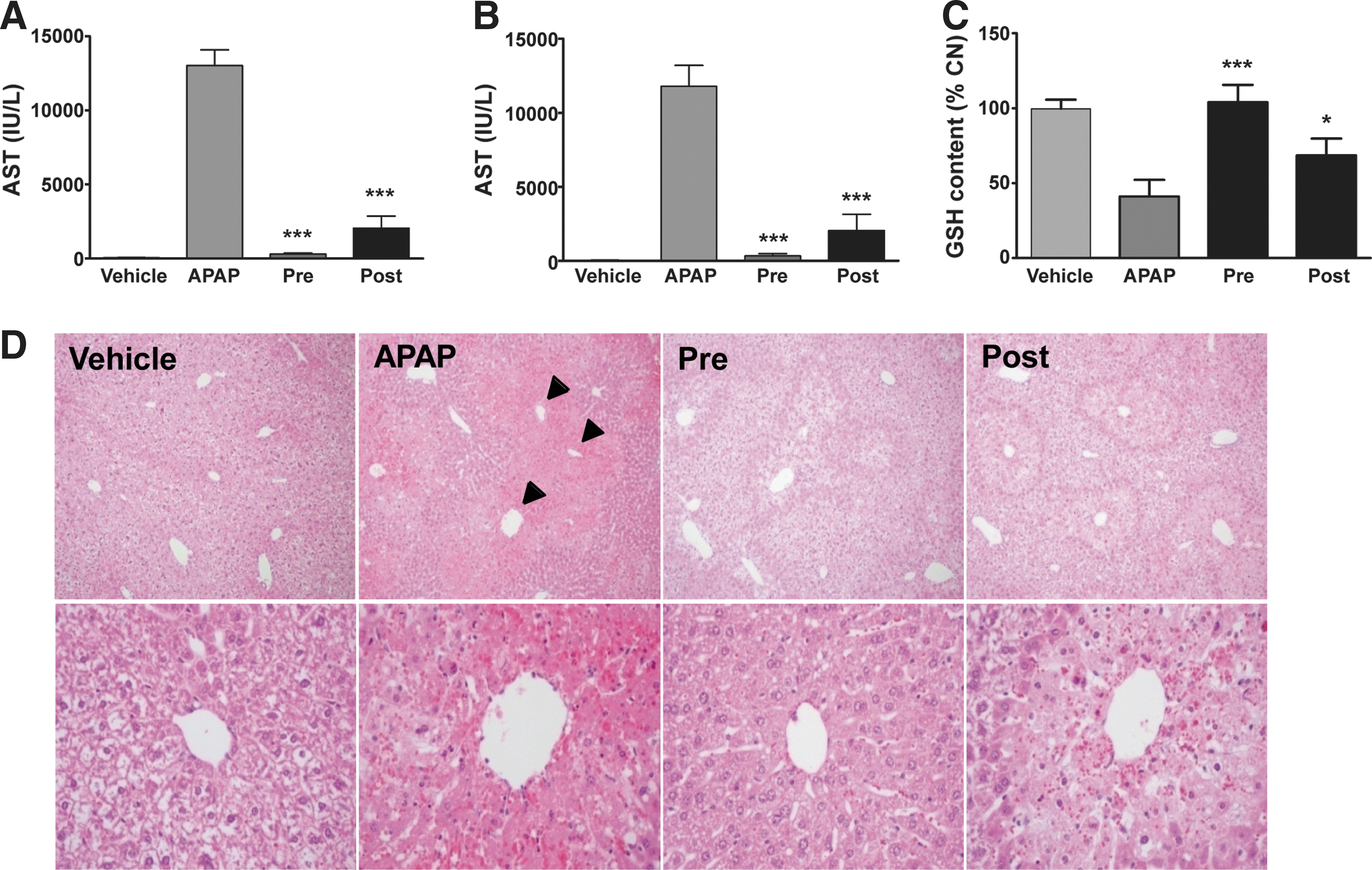
To determine whether the binding of NecroX-7 and NAPQI protects against AILI in C57BL/6 mice, we injected NecroX-7 2 h before or 1 h after the APAP injection. Twenty-four h after APAP treatment, the plasma aspartate aminotransferase (AST) (Fig. 6A) and alanine aminotransferase (ALT) (Fig. 6B) levels were markedly increased in the APAP-treated mice, but the mice treated with both APAP and NecroX-7 exhibited similar levels as that of normal. The hepatic GSH in the NecroX-7-treated groups were recovered to the normal levels, whereas the level was approximately 40% of the normal level in the APAP-treated group (Fig. 6C). Histological analysis of the APAP-treated mice revealed gross necrosis of the hepatocytes around the central vein, whereas the NecroX-7-treated mice showed diminished hepatic necrosis (Fig. 6D).
The hepatic necrosis caused by APAP is characterized by the increased production of superoxide and peroxynitrite, the levels of which are proportional to the levels of activation of c-Jun N-terminal kinase (JNK) signaling and 3-nitrotyrosine (3-NT) formation (9, 30). In mice treated with APAP for 24 h, the levels of phosphorylated JNK and 3-NT were drastically increased, and the pre-treatment with NecroX-7 decreased these levels to normal. The same effect was observed even when NecroX-7 was given after APAP administration (Fig. 7A). Given these results, it appears that NecroX-7 blocks the downstream effects of NAPQI formation without disturbing APAP metabolism via glucuronidation and sulfation (Fig. 7B). We next compared the protective effect of NecroX-7 with that of NAC, used clinically to treat APAP over dosing, by observing the survival of the mice for 72 h. Although the mice started to die 12 h after the APAP injection, mice treated with NecroX-7 showed a significant improvement in the survival rate, even at a 10 mg/kg dose. NecroX-7 greatly improved the survival rate of the mice in comparison to 200 mg/kg of NAC (Fig. 7C). In addition to its preventive effect when administered prior to APAP, NecroX-7 also successfully protected against AILI even when given 2 h after APAP (Fig. 8). The therapeutic effects observed in the post-treated groups are relevant to the treatment of APAP-intoxication because liver injury caused by APAP is seen in patients who have already ingested a toxic dose of APAP (5, 6).


Discussion
ROS and RNS are ubiquitous intermediates in our body and are generated during normal physiological processes, and multiple defense systems have evolved to protect our cells from these potentially damaging free radicals (33). However, unregulated ROS production results in damage to DNA, proteins, and lipids that causes cellular apoptosis and necrosis in cases of acute liver injury, ischemia/reperfusion, and other inflammatory diseases (12, 20, 28). In APAP-induced acute liver injury, formation of ROS and RNS from mitochondria are the main causes of hepatic failure (19). In previous studies, NecroX compounds showed strong antioxidant capacity and protected ischemia-reperfusion damage of liver in a Beagle dog model (4, 16). In addition to the ROS/RNS scavenging ability of NecroX-7, which showed binding affinity to the some radicals such as DPPH and ABTS, we showed that the direct chemical-mediated covalent binding to NAPQI could protect APAP-induced liver injury.
NAC is a pro-drug used to supply cysteine residue for the GSH replenishment, which is used to overcome the inefficient absorption of orally administered cysteine. Treatment with NAC showed some beneficial outcome after late administration in AILI, which indicates additional mechanisms of cellular protection independent GSH replenishment (37). Formation of hypotaurin from NAC has been shown protective in cell injury by acting as radical scavenger. However, the hepatoprotective role of hypotaurin is not completely understood (17). Therefore, novel therapeutic agents with a well-known mechanism are necessary to prevent accidental or intentional liver injury and death caused by APAP misuses.
NecroX-7 decreased t-BHP- and APAP-induced cell death and ROS/RNS production in HepG2 and primary hepatocytes. Some studies showed that indole derivatives have antioxidant activity (7, 32). It can be estimated that indole derivatives would have a protective effect on ROS-related disease such as AILI. However, ROS and RNS production is following process after NAPQI formation. Once NAPQI is produced, that binds to the cysteine residues of intracellular proteins and produces ROS/RNS continuously. Thus, it is not appropriate way to give an antioxidant agent to the patients for the treatment of AILI. As shown in Figure 6, NecroX-7 protected necrosis of hepatocytes around the central vein when treated before APAP, and blocked progress of necrosis when treated after APAP. Conversion of APAP to NAPQI mainly occurred in hepatocytes, and highly reactive NAPQI leads hepatocytes to necrosis through GSH depletion and massive ROS/RNS production. Thus, scavenging NAPQI is a novel and promising strategy in an early stage of AILI, and NecroX-7 will attenuate progress of AILI when treated after APAP over-consumption. As shown in Figures 4 and 5, NecroX-7 did not disturb APAP metabolism and GSH depletion 1 h after APAP treatment. It means that NecroX-7 acts as a NAPQI-scavenger when hepatocellular GSH is extremely low by APAP administration.
Saito and his colleagues showed that metallothionein (MT), a ubiquitous heavy metal binding and cysteine-rich protein, protects AILI by binding NAPQI directly (31). The induction of MT by ZnCl2 did not affect the metabolic activation of APAP but attenuated AILI. They concluded that MT acts as a second line of defense behind GSH to scavenge NAPQI and prevent the action of NAPQI (31). In our results, NecroX-7 completely protected AILI when treated before APAP and also after APAP administration. It shows that NecroX-7 may be used as a preventive drug for the AILI and also for the treatment of AILI. NecroX-7 showed low toxicity and did not induce liver and renal damage in mice and rats (data not shown, please see the patent about the NecroX-7 entitled ‘Indole and indazole compounds as an inhibitor of cellular necrosis', patent number: KR2008-0080519).
In summary, we report a new compound that protects against APAP-induced acute liver failure in the rodent model by directly binding to NAPQI. NAPQI is considered to be a key molecule in pathogenesis. Therefore, sequestering this highly toxic molecule with NecroX-7 may provide a new therapeutic strategy to treat AILI. NecroX-7 has some benefits. First, NecroX-7 did not disturb normal respiration of the mitochondria (Supplementary Fig. S3), suggesting that NecroX-7 has a low toxicity in normal cells. Binding of NecroX-7 with NAPQI is able to prevent the toxic side effects of APAP with low side effects. Second, NecroX-7 had more preventive than and similar therapeutic effect to NAC in our animal model. There are needs for the development of novel antidote for AILI with lesser side effects than NAC. Therefore, NecroX-7 is a promising drug for the treatment of APAP-induced organ failure.
Materials and Methods
Materials
NecroX-7 was kindly offered from LG Life Biosciences (Daejeon, Korea). Monochlorobimane was from Molecular Probes (Eugene, OR), and all other chemicals used in these experiments were purchased from Sigma-Aldrich (St. Louis, MO) or described in detailed methods. NAPQI was purchased from Sigma-Aldrich (St. Louis, MO). Cell culture reagents were purchased from Gibco-BRL (Grand Island, NY) and culture dishes were from Corning Inc. (Corning, NY).
Animals
Male C57Bl/6N mice (20–25 g) were obtained from the Nara Biotech Co. in Seoul, Korea. They were maintained under controlled conditions (25°C, 55% humidity and 12 h light/dark cycle) and fed a standard laboratory chow. All experimental procedures were in accordance with the guidelines issued by the Institutional Animal Care and Use Committee of the Chungnam National University School of Medicine (Daejeon, Korea).
Mice model of acetaminophen-induced liver injury
Mice were administered 20 ml/kg phosphate-buffered saline or 400 mg/kg of acetaminophen intraperitoneally (I.P.) after overnight food deprivation. Some mice received 10 ml/kg D.W or NecroX-7 orally (50 mg/kg) in the D.W 2 h before or 1 h after APAP. Twenty-four hours after the acetaminophen treatment, blood of the mice was collected and the mice were sacrificed to obtain their livers. A portion of the liver was fixed in formalin solution for histological sections. The serum was obtained from the whole blood by centrifugation after coagulation.
Measurements of serum AST and ALT
Serum alanine aminotransferase (ALT) and aspartate transaminase (AST) content were assayed as markers of hepatotoxicity. Blood was collected from the heart under general anesthesia with 0.4 mg/g of avertin. ALT and AST levels were measured on a Hitachi 7180 autoanalyzer using Wako reagents (Wako Pure Chemical Industries, Osaka, Japan).
Assessment of cell viability and plasma membrane integrity
Cell viability was determined by Cell Counting Kit-8 assay (Dojindo, Kumamoto, Japan), as described (3). Integrity of plasma membrane was assessed by measuring extracellular release of lactate dehydrogenase (LDH). LDH release was measured using a CytoTox 96® Nonradioactive Cytotoxicity Assay kit (Promega, WI), according to the manufacturer's guidelines.
Western blot analysis
Antibody against phospho-T183/185 JNK1/2 was purchased from Invitrogen (Camarillo, CA). Anti-JNK, 3-nitrotyrosine, and anti-tubulin antibodies were from Santa Cruz Biotechnology (Santa Cruz, CA), Abcam (Cambridge, UK), and Sigma-Aldrich, respectively. Protein concentration was determined by the Bradford method. Western blotting was performed as described elsewhere.
Analysis of GSH content in cells and livers
Livers from mice were obtained after treatment of APAP with or without NecroX-7 and homogenized in 20 vol of cold 50 mM Tris buffer (pH 7.4), followed by sonication. The reduced form of glutathione level was determined by staining with 100 μM of monochlorobimane, and calculated with mean fluorescence intensity relative to protein concentration (13).
Immunohistochemistry of liver sections
At the end of the experiments, livers of mice were fixed overnight in 10% formalin at 4°C, and embedded in paraffin. Paraffin-embedded tissue sections were deparaffinized and the sections were incubated with the primary antibody (1:100–1:200) at 4°C overnight. From this point until color development, the DAKO LSAB+ kit (DAKO Corporation, Carpinteria, CA) was used as a suggested protocol. The slides were counterstained with Gills Hematoxylin II and immersed in ammonia blue. Then the slides were dehydrated and mounted with Permount (11).
LC-MS/MS analysis of APAP and its metabolites
APAP, and its sulfate, glucuronide, glutathione, and cysteine conjugates were determined as previously with a little modification (18). All LC-MS/MS experiments were carried out on an Agilent 1200 HPLC system (Agilent Technologies, Santa Clara, CA) coupled with and Thermo LTQ linear ion trap mass spectrometer (Thermo Scientific, San Jose, CA) equipped with an electrospray ionization (ESI) source. For high resolution LC-MS analysis, a 12 Tesla Fourier-transform ion cyclotron resonance mass spectrometer (FT-ICR MS) equipped with ESI source was used. The LC separation was achieved using a Luna C18 RP column (150 mm×2 mm I.D., 5 μm 100 Å particles, Phenomenex, Torrance, CA) with gradient elution of mobile phase A (2 mM ammonium acetate) and mobile phase B (methanol) at a flow rate of 0.2 ml/min. Mobile phase composition was changed from 0% B at 0 min to 90% B at 8 min and maintained for 6 min. The metabolites separated by LC were detected by mass spectrometer in the positive ESI mode using the selected reaction monitoring (SRM).
Isolation of primary hepatocytes
Primary mouse hepatocytes were prepared from 20–25 g C57Bl/6N mice by collagenase 0.8 mg/ml) perfusion. Isolated cells were further purified with a percoll density gradient and plated appropriate density to the culture plates (96-well plates) in medium 199 (Sigma, USA) (35). Cells were allowed to attach for 3–6 h, and, media was changed to remove unattached cells or dying cells.
Statistical analysis
Results are reported as means±SD or SE. Statistical analyses between two measurements were determined by the two-tailed unpaired Student's t-test, and survival curves were analyzed with a log-rank test. Statistical significance was assessed using GraphPadInstat 3.05 software (San Diego, CA). P values less than 0.05 were considered significant.
Additional Methods
Determination of intracellular ROS/RNS, NADPH/NADP+ ratio, free radical scavenging activity, total antioxidant activity, oxygen consumption rate and CYP2EI activity are described in Supplementary Methods.
Footnotes
Acknowledgments
We thank Dae-Sik Lim for help generating the final figures; Kyu Lim and Byung-Doo Hwang for advice on the biochemical experiments; and Chul-Ho Lee for advice on the animal experiments. We are grateful Dr. J. Thomas LaMont at Harvard Medical School for the manuscript editing.
This work was supported by a National Research Foundation grant (2011-0027839, 2012-0005767) and a grant from the Korea Healthcare Technology R&D Project, Ministry of Health & Welfare (A10058) by the Korea government.
Author Disclosure Statement
Soon-Ha Kim is a participant of the patent about the anti-necrotic uses of indole derivatives (Korean patent number: KR2008-0080519). The other authors declare no conflict of interest.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.