
Abstract
Faecalibacterium prausnitzii is one of the most abundant commensal microbes in the human gut. It is an important supplier of butyrate to the colonic epithelium, and low numbers of faecalibacteria have been associated with severe inflammatory bowel disease. Previous studies revealed that F. prausnitzii shuttles electrons extracellularly to oxygen in systems containing flavins and thiols. Since this electron shuttling to oxygen strongly stimulates growth, the present studies were aimed at elucidating the role of riboflavin as an extracellular electronophore of F. prausnitzii. We show that F. prausnitzii can use riboflavin as a mediator for extracellular electron transfer (EET) to the anode of microbial fuel cell systems. However, this bacterium relies on exogenous riboflavin, since it does not secrete this compound as shown by the analysis of a spent growth medium using cyclic voltammetry (CV). Importantly, CV showed that riboflavin can undergo fully reversible redox cycling under physiologically relevant conditions. Lastly, riboflavin is shown to mediate the electrochemical oxidation of the main bacterial reducing equivalent NADH. Based on our present observations, we hypothesize that riboflavin is of major importance as a redox mediator for bacterial EET and growth in the human gut. Antioxid. Redox Signal. 17, 1433–1440.
Introduction
Innovation
Faecalibacterium prausnitzii is one of the most abundant beneficial microbes in the human gut. Here, we have employed microbial fuel cell systems to investigate how F. prausnitzii exploits the redox-active vitamin riboflavin, which is commonly present in the gut lumen, as an electronophore for extracellular electron transfer (EET). We show that faecalibacteria can use riboflavin to drive the chemical oxidation of NADH, and that this vitamin undergoes a fully reversible redox cycle under physiologically relevant conditions. Our findings thus highlight the importance of riboflavin as a redox mediator for bacterial EET and growth in the human gut.
Most of the bacteria capable of extracellular electron shuttling were thus far isolated from geological niches, such as marine sediments. A completely different environment is the mammalian gut, one of the most diverse microbial ecosystems known, which harbors up to 1012 microbes per gram of fecal content (1). Importantly, the environmental conditions prevailing in the gut would favor EET, since nutrients are readily available; the oxygen tension is negligible; the redox potential is low; and redox-active compounds such as flavins are abundant (2, 6). Interestingly, flavins are not only derived from the host diet but also derived from some commensal microbes in the gut, such as Escherichia coli and Bacteroides, which can produce riboflavin, which is a well-known redox-active vitamin (5). For example, it was previously shown that L. lactis can exploit riboflavin as a soluble redox mediator (5). This characteristic is likely to offer an advantage when present in dairy products where riboflavin is readily available (6). More recently, we have shown that one of the most abundant commensal gut microbes, Faecalibacterium prausnitzii, is capable of EET to oxygen in a system containing flavins and thiols (4). The present studies were therefore aimed at determining how F. prausnitzii can exploit riboflavin as a redox mediator for EET. Through the use of microbial fuel cells, we show that the electrochemical redox reaction of riboflavin can be linked to the NAD+/NADH redox couple. Our findings thus highlight the potential benefit of EET for F. prausnitzii cells growing in the human gut.
Results
The extracellular riboflavin concentration sets a limit to EET by F. prausnitzii
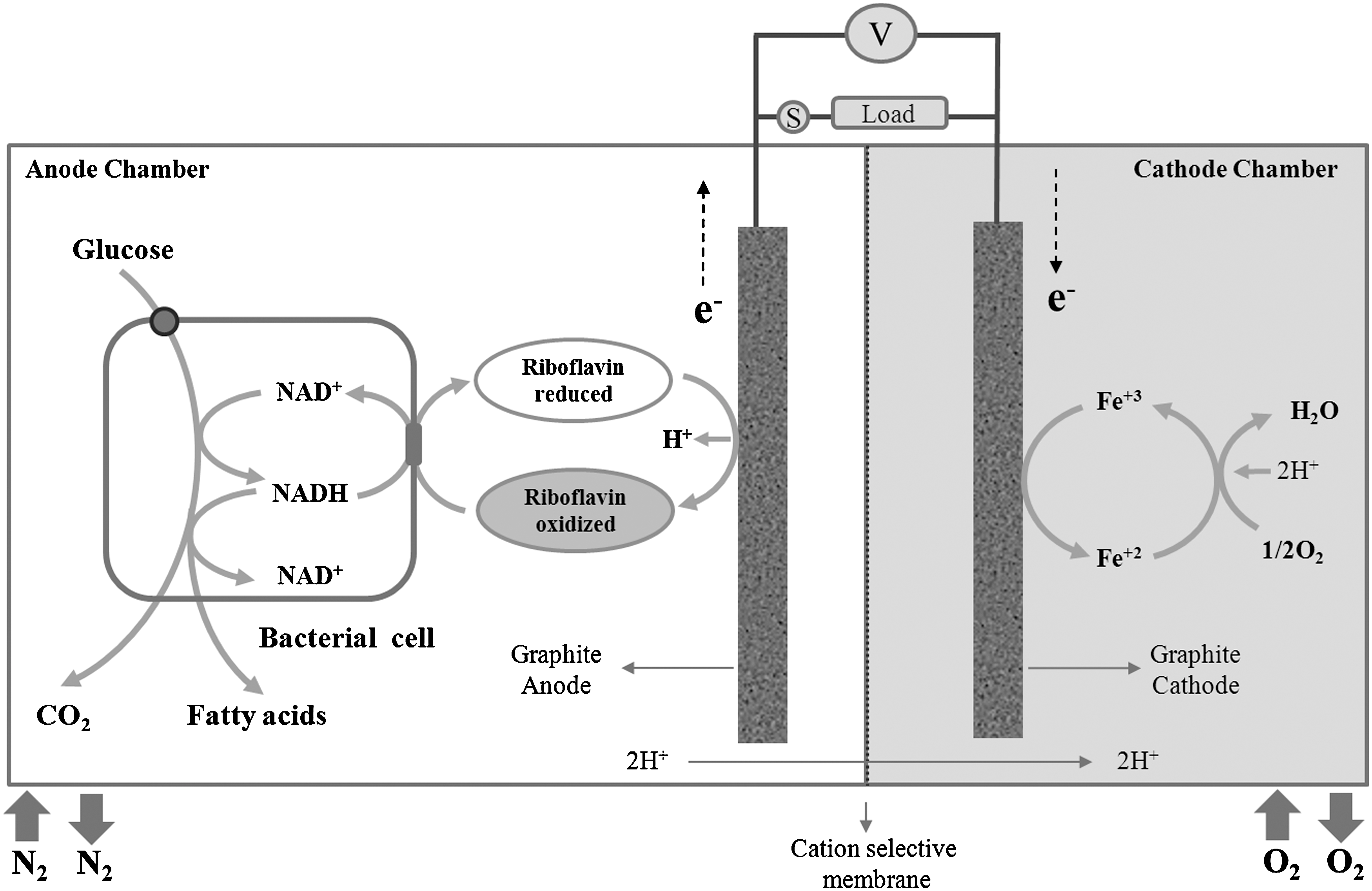
To investigate how F. prausnitzii can employ riboflavin as an electron shuttle, a two-compartment microbial fuel cell system was developed (Fig. 1). In this system, the oxidative metabolism of F. prausnitzii (NAD+/NADH; Eo

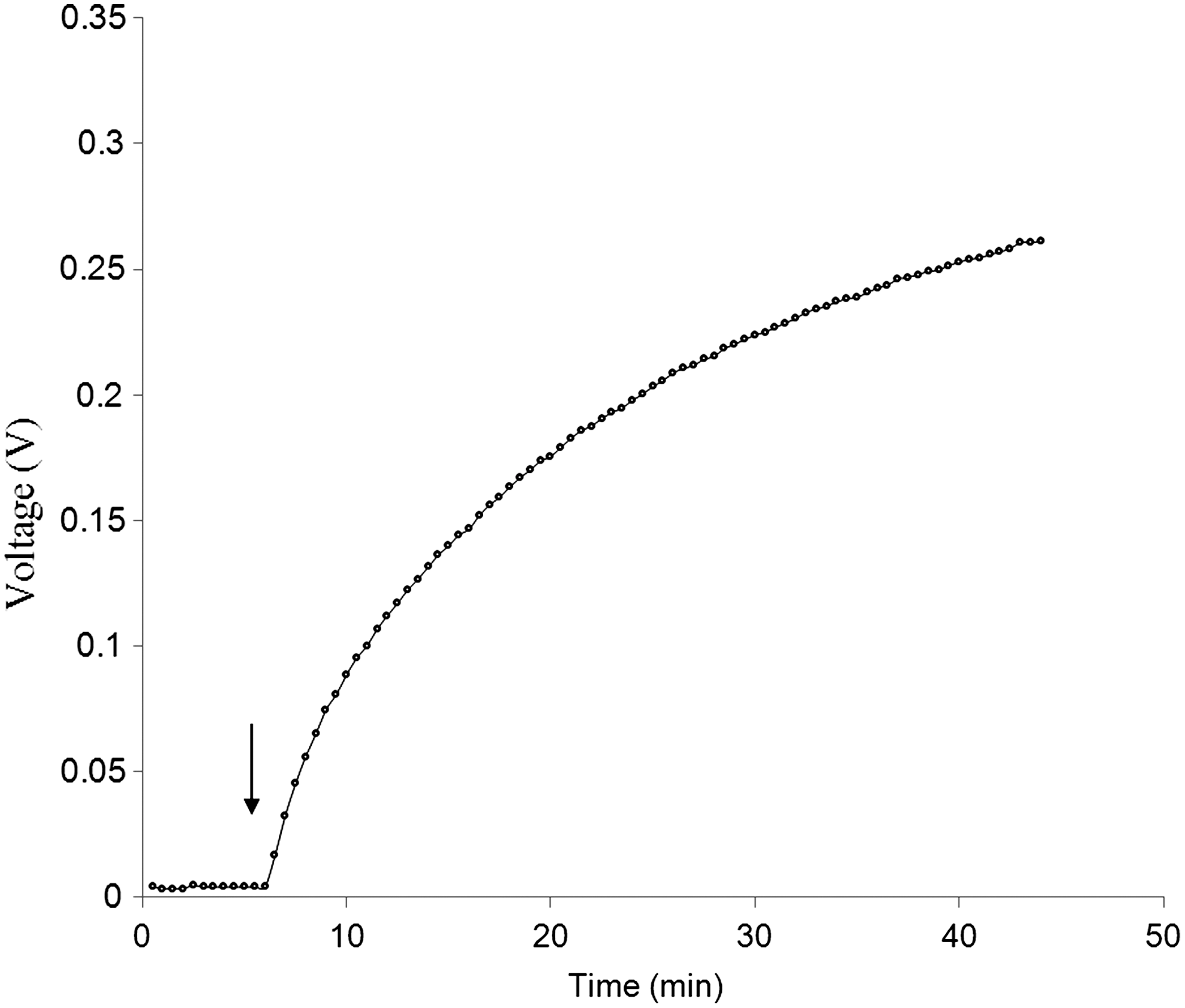

). No significant current was produced by F. prausnitzii when glucose was omitted from the anode chamber even if riboflavin was added to 400 μM. However, a rapid increase in current was observed when 0.1 M glucose was subsequently introduced into the anode chamber ( ). In control experiments, current production was not observed when F. prausnitzii was omitted from the anode chamber, neither in the presence nor absence of 0.1 M glucose and up to 400 μM riboflavin (
). In control experiments, current production was not observed when F. prausnitzii was omitted from the anode chamber, neither in the presence nor absence of 0.1 M glucose and up to 400 μM riboflavin ( ).
).
Current production by riboflavin-mediated chemical oxidation of NADH
To mediate bacterial EET, the redox couple of reduced and oxidized riboflavin must be coupled to the NAD+/NADH redox couple (7). The proof of principle that this is possible was obtained by monitoring the effects of the addition of riboflavin to the bacterial reducing equivalent NADH in the anode chamber of the fuel cell depicted in Figures 1 and 2. In this case, the chemical oxidation of NADH was coupled to ferricyanide reduction via riboflavin. Figure 5 shows that when NADH was introduced into the anode chamber, a rapid increase in current was observed due to direct oxidation of NADH. Importantly, the current increased several fold when riboflavin was introduced into the anode chamber (Fig. 5). This observation shows that riboflavin is capable of facilitating electron transfer from NADH to the graphite electrode.

). Current was not produced when 200 μM riboflavin was introduced into the anode chamber alone ( ).
).
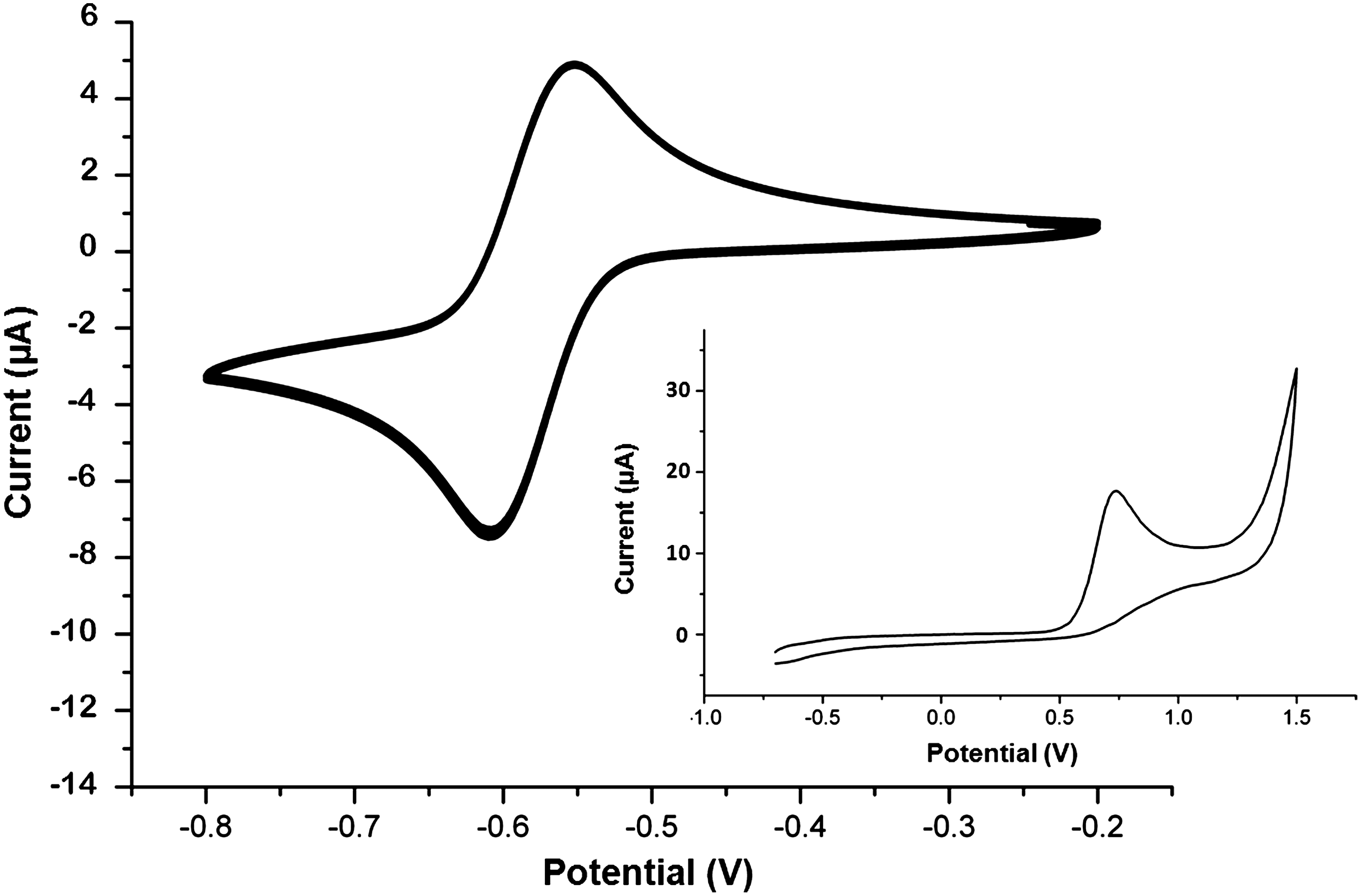
Cyclic voltammetry reveals a fully reversible redox cycle of riboflavin
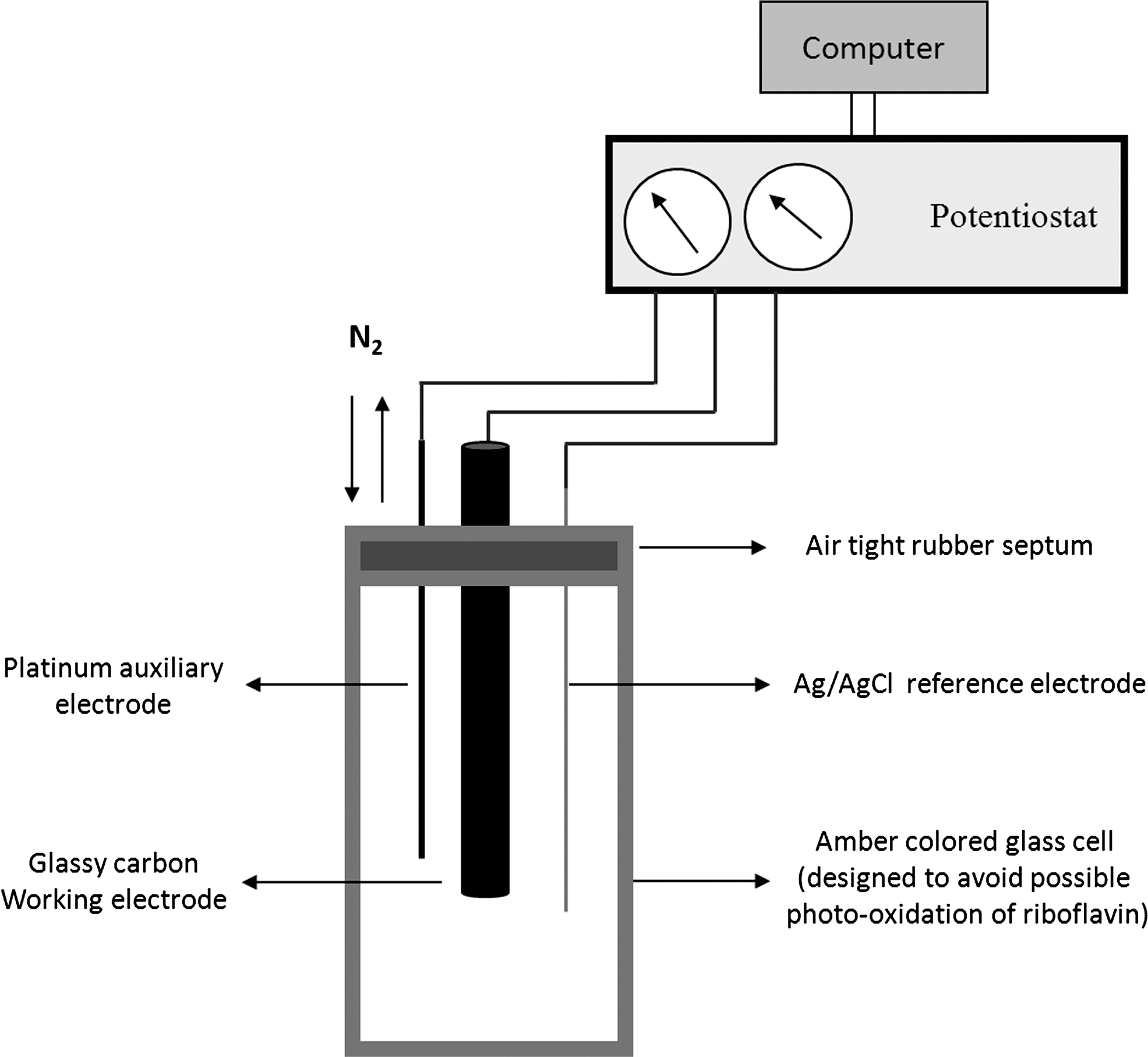
The redox behavior of riboflavin and its role as redox mediator were further studied by cyclic voltammetry (CV) in a small bioreactor (Fig. 6). The cyclic voltammogram of riboflavin shows that this redox mediator can undergo a fully reversible redox cycle under the conditions defined by the fuel cell. The calculated midpoint redox potential under these conditions was −0.58 V versus Ag/AgCl (Fig. 7). The fully reversible redox cycle and stability under physiological conditions clearly show that riboflavin is an excellent potential redox mediator for EET.
F. prausnitzii relies on exogenous riboflavin for EET
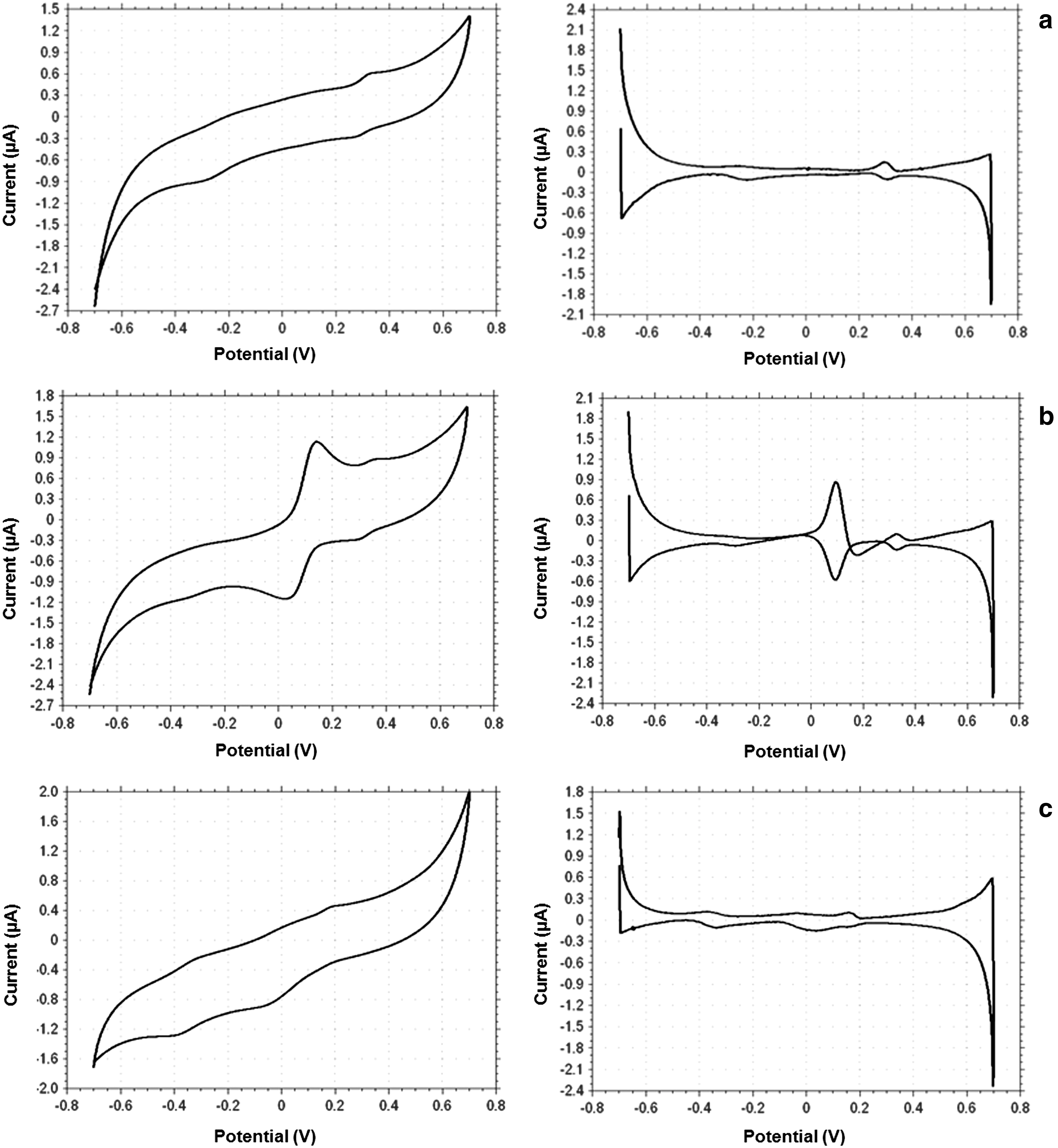
Considering the importance of riboflavin for optimal growth of F. prausnitzii in environments with low levels of oxygen, it was relevant to know whether this bacterium can produce and secrete riboflavin. Although not reported previously (4), this was still a possibility. To investigate the possible secretion of redox-active riboflavin, CV was performed on spent broth derived from 24-h-old cultures. Figure 8a shows the cyclic voltammogram of the spent broth and the corresponding first-derivative voltammogram. When compared to the cyclic voltammograms of spent broth with 50 μM of added riboflavin (Fig. 8b) or fresh broth containing 0.13 μM of riboflavin (Fig. 8c), it is clearly evident that the cyclic voltammogram of the spent broth without added riboflavin did not show a redox wave assignable to riboflavin (Fig. 8a, b). Nevertheless, a minor redox process was observed at +0.3 V (vs. Ag/AgCl; Fig. 8a). The addition of 50 μM riboflavin to the spent broth (Fig. 8b) showed that the redox process at +0.3 V was independent of the presence or absence of riboflavin. These findings show that F. prausnitzii did not secrete riboflavin in amounts above the detection limit. It thus seems that F. prausnitzii cannot exploit self-secreted riboflavin for EET, at least under the growth conditions employed in the present studies.
Discussion
F. prausnitzii is a major human commensal representing ∼20% of the total gut microbiota. The importance of this bacterium for human health is underscored by its anti-inflammatory effects in enteric colitis (8). Upon fermentation of glucose and the consumption of acetate, faecalibacteria produce butyrate as the major fermentative end product (4). In the present studies, we have employed a microbial fuel cell system to define the role of riboflavin as an electron shuttle exploited by F. prausnitzii. In this approach, glucose was used as an electron donor, resting bacterial cells as catalysts and riboflavin as electron mediator. The functionality of the fuel cell system was clearly demonstrated by introducing riboflavin into an anode compartment containing faecalibacterial cells that were pre-energized with glucose. This immediately led to the generation of currents that were proportional to the added amounts of riboflavin (Fig. 4). These experiments further revealed that when glucose, the sole electron donor, was omitted from the anode compartment, significant current production was not observed even in the presence of high concentrations of riboflavin. Together, we can infer from these findings that the added glucose was metabolized by the F. prausnitzii cells, resulting in the generation of electron carriers, NADH in particular. As shown by the fuel cell experiments with NADH and riboflavin, the generation of NADH and its subsequent oxidation would be sufficient to drive the extracellular transfer of electrons to the fuel cell's anode via riboflavin, as is schematically depicted in Figure 2.
Our present findings show that electron transfer from the F. prausnitzii cells to the anode of a fuel cell is dependent on the availability of extracellular riboflavin. This differs from the EET mechanism employed by Geobacter spp., which is capable of shuttling electrons directly to the electrode without the need for an externally added electronophore (9). Consistent with this model, F. prausnitzii was found to be incapable of transferring electrons directly to the electrode, even after extended incubation periods of up to 24 h. This behavior might be related to the normal environmental niche occupied by F. prausnitzii, the human gut, where extracellular nutrients are abundant, and where oxygen diffuses in from the epithelial cells. Such a view is consistent with our previous observation that F. prausnitzii grows optimally in the presence of low amounts of oxygen provided that riboflavin and oxidized thiols are available for electron transfer to oxygen (4). In contrast, free-living organisms growing in soil or marine sediments, such as Geobacter spp., employ mineral respiration (9). Notably, riboflavin-mediated EET is not a general bacterial phenomenon as exemplified for E. coli, which can produce riboflavin, but cannot use this compound as a redox mediator (5). Interestingly, Park and Zeikus (7) have shown that neutral red (NR) can act as a redox mediator for E. coli. In the present studies, we observed that this is not the case for F. prausnitzii, as current was not produced in the fuel cell when 200 μM NR was used instead of riboflavin (data not shown). Lastly, our data show that F. prausnitzii does not secrete riboflavin to detectable levels, unlike other electrogenic bacteria of nongut origin, such as lactococci associated with dairy products or marine bacteria of the Shewanella species (5, 9). Thus, it seems that in the human gut, F. prausnitzii must rely on external riboflavin synthesized by other gut microbes or ingested by the human host.
In conclusion, our present fuel cell experiments show that the human gut commensal F. prausnitzii, which is considered to be important for gut health, can exploit riboflavin as an electron mediator to shuttle electrons to an appropriate electron acceptor. The finding that this electron acceptor can be the graphite anode of a fuel cell allowed a detailed characterization of the extracellular electron shuttling by F. prausnitzii as well as the redox properties of riboflavin. The completely reversible redox cycling of riboflavin makes this compound ideally suited as a redox mediator in the human gut. Nevertheless, it should be noted that the human gut contains many redox-active substances derived from the host, the diet of the host, or gut microbial sources. We therefore hypothesize that the extracellular electron transport of F. prausnitzii can also be mediated by some of these alternative redox-active substances and, most likely, this will also apply to other members of residential gut microbiota.
Notes
Bacterial strains and culturing conditions
F. prausnitii strain A2-165 (DSM 17677) was maintained at 37°C on yeast extract, casitone, fatty acid and glucose (YCFAG) agar (4) in an anaerobic tent. For the microbial fuel cell experiments, the bacterial cells were grown anaerobically in 50 ml of YCFAG broth to an optical density at 600 nm (A600) of ∼0.8. Cells were harvested by centrifugation, washed in 50 mM potassium phosphate buffer (pH 7.0) and suspended in 0.5 ml of the same buffer. The suspension of resting bacterial cells obtained was used to inoculate the anode chamber of the microbial fuel cell. Cell-free spent broth (24 h) was used for CV. All buffers and media were made anaerobic by flushing, immediately after autoclaving, with oxygen-free nitrogen for 30 min.
Construction of a microbial fuel cell
A custom-made two-chambered, microbial fuel cell was fabricated from borosilicate glass bottles with 65 ml working volumes for the cathode and anode chambers (Fig. 1). The two compartments were separated by a CMI-7000S cation exchange membrane (Membranes International, Inc.), using a 20 cm diameter septum. Graphite slabs (dimensions of 5×1.5×0.2 cm) were used as cathode and anode. The electrodes were connected to the external circuit with insulated copper wire and the bare connections were sealed with a nonconductive epoxy resin. The resistance between the wires and the electrodes was less than 2 Ω. The anode chamber contained 50 mM potassium phosphate buffer (pH 7.0) as anolyte, 0.1 M glucose and a Ag/AgCl reference electrode. The cathode chamber contained 100 mM potassium-phosphate buffer (pH 7.0) with 50 mM potassium ferricyanide as catholyte. The assembled fuel cell was maintained at 37°C and the anode and cathode chambers were purged continuously with nitrogen gas and air, respectively. The reduction potential of potassium ferricyanide is Eo
Current production by riboflavin-mediated chemical oxidation of NADH
The chemical oxidation of NADH was coupled to reduction of potassium ferricyanide using riboflavin as electron mediator in the above-mentioned fuel cell system. The anode and cathode compartments were purged with nitrogen and air respectively and maintained at 37°C. The fuel cell was operated in open circuit mode until the maximum OCV was achieved. For current production, the cell was switched to the closed circuit mode using a fixed external load of 150 Ω resistance. After a specified period, NADH (1 mM final concentration from 325 mM stock prepared in 50 mM potassium phosphate buffer pH 7.0 and deoxygenated by purging with N2 for 10 min) was added as electron donor. After ca. 10 min incubation, 200 μM of riboflavin was injected into the anode chamber as redox mediator. In control experiments 200 μM of riboflavin alone was used in the anode chamber.
Coupling of bacterial redox-metabolism with the riboflavin and ferricyanide redox couples
The bacterial cell pellet (prepared as described above) was inoculated in the anode chamber containing 50 mM potassium phosphate buffer and 0.1 M glucose. The bacterial cells were incubated for 10 min after which 200 μM riboflavin was added as redox mediator. The current production was determined by operating the fuel cell in closed circuit mode as described above.
Cyclic voltammetry
CV was carried out using a potentiostat (model 600C; CH Instruments) in a single-chambered cell with a working volume of 4 ml. The assembled reactor contained a 3 mm diameter Teflon shrouded glassy carbon working electrode, an Ag/AgCl reference electrode and a platinum wire auxiliary electrode (Fig. 6). Potassium phosphate buffer (100 mM, pH 7.0) with 5 mM NaCl was used as electrolyte. The cell was purged with N2 for 10 min before running experiments. A scan rate of 50 mV s−1 was employed under quiescent conditions.
Footnotes
Acknowledgment
M.T.K. was supported by a grant from the Graduate School GUIDE of the University of Groningen.
Author Disclosure Statement
M. Tanweer Khan, Wesley R. Browne, Jan Maarten van Dijl, and Hermie J.M. Harmsen declare no competing commercial interests or any other conflicts of interest.