
Abstract
Introduction
Txl-2, a member of the thioredoxin (Trx) and nucleoside diphosphate kinase (NDPk) family, was cloned and characterized in 2003 (37). It is mainly expressed in testis, and at lower levels in lung and brain, while more ubiquitously expressed at very low levels in other tissues (37). While Txl-2 function in cancer remains largely unknown, other Trx family members have been reported to be critical in the regulation of tumor development and progression. For instance, Trx-1 is overexpressed in a number of human primary tumors, including lung, colorectal, and cervical cancers and leukemia (23). Specifically, an increased expression of Trx-1 in CRC has been shown to be associated with poor prognosis in patients with liver metastasis (30). We have recently demonstrated an increased expression of Txl-2 in colon cancer (25), but its specific role in colon cancer and the underlying mechanisms are still unknown.
Innovation
Thioredoxin (Trx) system has been increasingly linked to cancer malignant phenotypes in the recent years. In this work, we provide further evidence to this scenario by reporting another Trx family member thioredoxin-like protein 2 (Txl-2), which is upregulated in colon cancer with a strong correlation with patient prognosis. Txl-2 was initially identified as the target of the monoclonal antibody MC3, previously isolated in our laboratory, which detects colon cancer with high sensitivity and specificity. We demonstrate now that the most abundant isoform Txl-2b promotes colon cancer metastasis. Our studies show that interaction between Txl-2b isoform and the small GTPase family member Ran mediates colon cancer cell invasion and metastasis via phospho-Akt activation and induction of matrix metalloproteinases expression. Taken together, our data uncover Txl-2 as a novel biomarker and target molecule for human cancer and offers a novel mechanism of how Trx family members regulate metastasis.
It is well-recognized that mRNA splicing can lead to production of protein isoforms with oncogenic properties (32).Three different isoforms have been identified for Txl-2, including full length Txl-2 (Txl-2a), Txl-2b (splicing variant lacking exons 1 and 5), and Txl-2c (splicing variant lacking exons 1, 4, and 5) (25, 37). Of these, Txl-2b was upregulated in colon cancer tissues compared with normal mucosa, while Txl-2c was almost exclusively present in colon cancer tissues, and full length Txl-2 seldom occurred in both normal mucosa and colon cancer tissues (25). Thus, it would be interesting to investigate whether these three transcript variants show differential effects on the functional properties of colon cancer cells.
The ability to invade tissues and metastasize to other sites is one of the most critical clinical parameters in predicting the progression of cancer to a more aggressive phenotype. We hypothesized that Txl-2 may mediate colon cancer progression and especially metastasis based on the following reasons. First, it is well established that the cytoskeleton plays a crucial role in tumor metastasis (15). Txl-2b was found to possess intrinsic microtubule binding capability, and therefore, might be a novel regulator of microtubule physiology (37). Second, Txl-2, also named nm23-H9, belongs to the NDPk family that has been extensively studied in regard to their roles in cancer metastasis. Ten genes belonging to the NDPk family have been identified in humans (4). Among them, the nm23-H1 was the first identified metastasis suppressor gene (5), whereas nm23-H4, nm23-H6, and nm23-H7, were found to be overexpressed in gastric and colon cancer (38). However, the role of Txl-2 (nm23-H9) in colon cancer metastasis remains largely unknown.
In the present study, we examined the Txl-2 expression in 243 cases of colon cancer tissues and found that overexpression of Txl-2 strongly correlated with the histological grade and prognosis of colon cancer patients. Knockdown of Txl-2 expression significantly inhibited the invasive and metastatic abilities of colon cancer cells. The differential effects of each Txl-2 isoform on cancer cell invasion and metastasis were further investigated. Overexpression of the most abundant isoform Txl-2b (88%) can promote cancer cell invasion and metastasis. Txl-2b harbors a regulator of chromosome condensation-1(RCC1) motif which is absolutely required for a newly discovered interaction with Ran. We further explored the downstream signaling events that participated in the Txl-2b-Ran mediated invasion and metastasis.
Results
Upregulation of Txl-2 in colon cancer correlates with tumor staging and patient survival
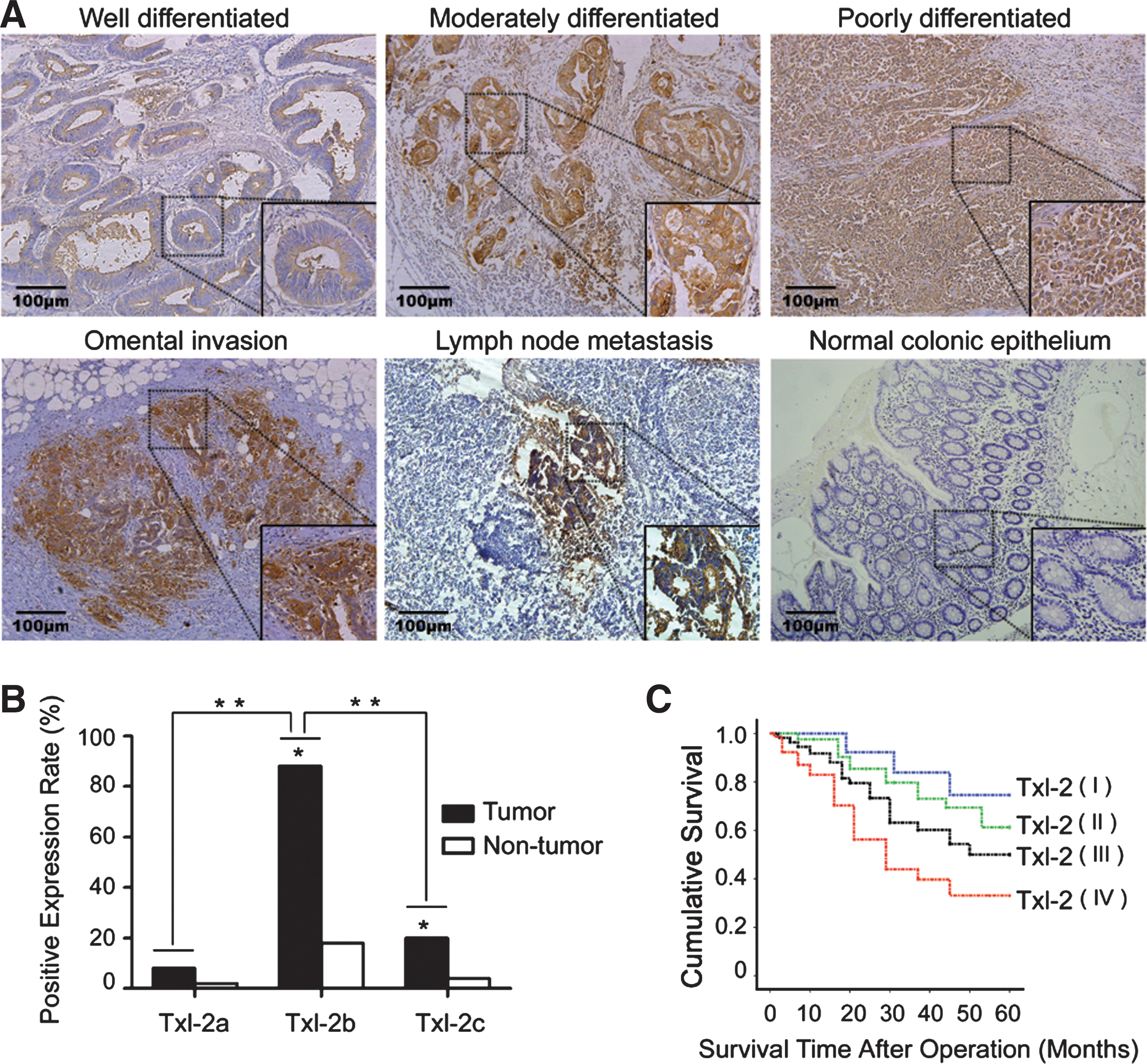
The expression of Txl-2 in human colon cancer tissues and adjacent nontumorous tissues was examined in 243 colon cancer patients. We found that Txl-2 expression was increased in colon cancer cells compared to adjacent nontumorous tissues, and normal colonic epithelium (Fig. 1A), and the increased expression was significantly correlated with the degrees of cancer cell differentiation (Table 1, p=0.019). As shown in Table 1, the increased Txl-2 expression also correlated with tumor stages consisting of tumor size (T) (p<0.001), lymph node invasion (N) (p<0.001), and distant metastasis (M) (p=0.022). To determine whether Txl-2 isoforms have differential expression rates, a semiquantitative reverse transcription polymerase chain reaction (RT-PCR) analysis was performed in 50 pairs of colon cancer samples using the method we reported previously (25). We found that Txl-2b was the most frequently presented Txl-2 isoform in tumor tissues (88%, 44/50), as compared to Txl-2a (8%, 4/50, p<0.001) and Txl-2c (20%, 10/50, p<0.001) (Fig. 1B). These results supported our previous findings that the total Txl-2 mRNA level was elevated in colon cancer tissues compared with paracancerous tissues and adjacent normal tissues in a relatively small number of patients (25). Furthermore, we investigated whether Txl-2 expression can be an independent prognostic factor. In general, the mean follow-up time of the patients with colon cancer was 41.8 months with a median value of 50 months. The 5-year survival rate of the 243 patients was 47%. Importantly, the survival rates based on cumulated survival and survival time after operation were inversely correlated with Txl-2 expression as shown by the Kaplan–Meier postoperative survival curve (Fig. 1C, log-rank test: p<0.001). Further multivariate analysis demonstrated Txl-2 expression as an independent prognostic factor (Table 2). Adjusted hazard ratio (HR) was set to 1.00 in the Txl-2 negative (I) expression group; the adjusted HR of weak positive (II), moderate positive (III), and strong positive (IV) groups were 1.74, 1.96, and 3.02, respectively. All these data pointed to an important role for Txl-2 as an independent prognostic factor for patients with colon cancer.
p value when expression levels were compared using Kruskal–Wallis test.
p value when expression levels were compared using Mann–Whitney test.
TNM, tumor-node-metastasis staging classification system. From National Comprehensive Cancer Network (NCCN) guidelines. Available at:
Multivariate analysis was used to evaluate whether Txl-2 expression is an independent prognostic factor for colon cancer patients.
Adjusted HR was set to 1.00 in the Txl-2 negative (I) expression group.
T, extent of the tumor; N, nearby (regional) lymph nodes; from NCCN guidelines. Available at:
HR, hazard ratio; Txl-2, thioredoxin-like protein 2.
Depletion of Txl-2 inhibits in vitro invasive and in vivo metastatic properties of colon cancer cells
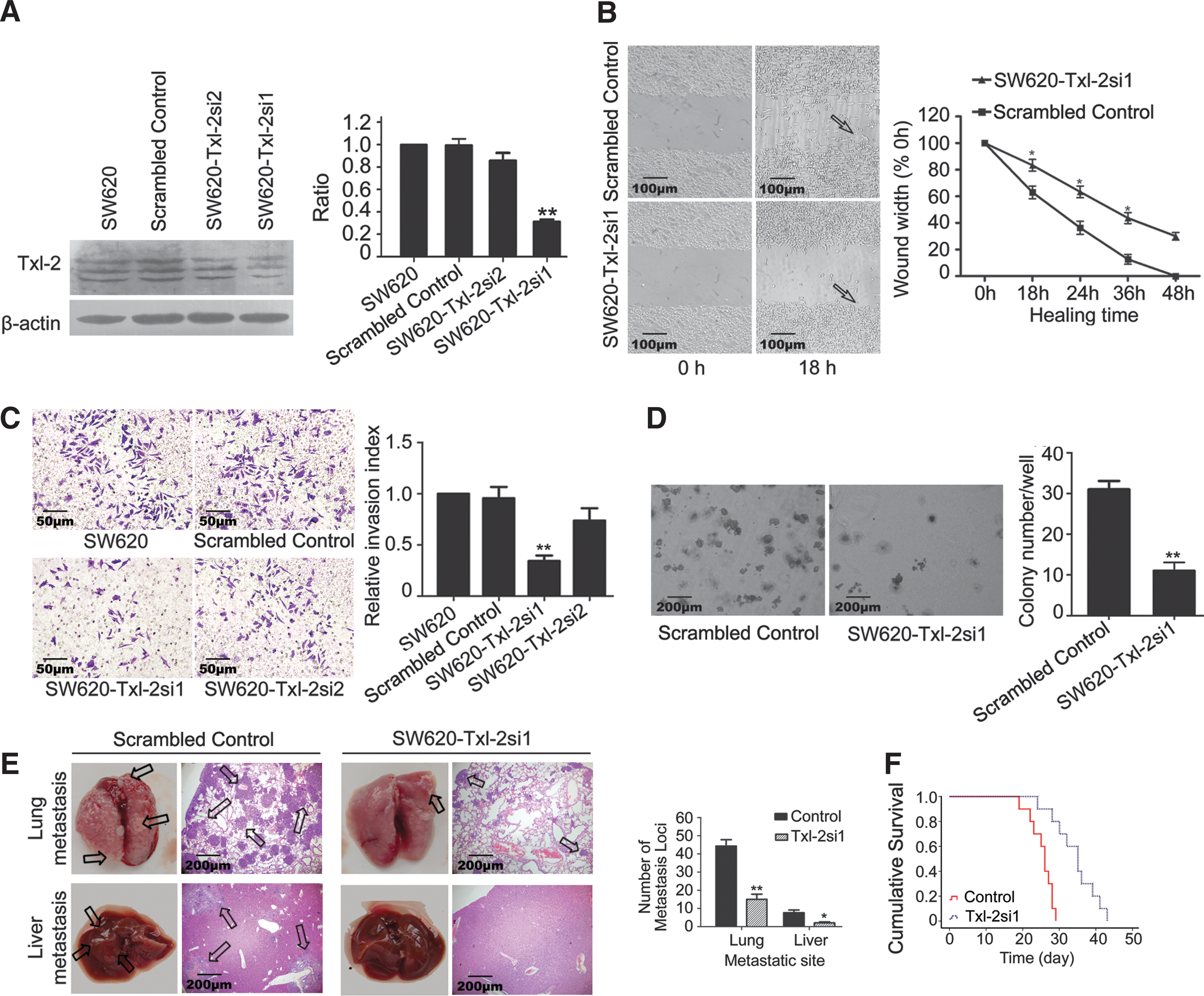
To explore the impact of Txl-2 on colon cancer invasion and metastasis, we knocked down Txl-2 expression in the SW620 colon cancer cell line which has a high metastatic potential (19) by using Txl-2-specific siRNA targeted to the overlapping sequences of three Txl-2 isoforms, aiming to knock down simultaneously the expression of all Txl-2 isoforms. Two Txl-2 siRNA expression vectors named pSilencer Txl-2 (#si1 and #si2) were constructed, and the knock down efficacy was compared based on Txl-2 expression using specific Txl-2 antibodies able to recognize the three isoforms (37). We found that SW620-Txl-2si1 cells were able to inhibit Txl-2 expression by 3.1-fold compared to the scrambled control (p=0.0004), whereas SW620-Txl-2si2 did not result in significant Txl-2 downregulation (Fig. 2A). To test the impact of Txl-2 on cell migration, we performed a wound healing assay by using SW620-Txl-2si1. As shown in Figure 2B, Txl-2 downregulated cells exhibited longer healing time than the control cells. Moreover, as compared to control cells, the Txl-2 downregulated cells showed a reduced cell invasive ability in the Transwell invasion assay (Fig. 2C). Specifically, SW620-Txl-2si1 cells showed the strongest inhibition of cell invasion over the SW620-Scrambled control cells (2.85-fold, p=0.006). Moreover, these SW620-Txl-2si1 cells significantly reduced the number of colonies in a colony formation assay in soft agar relative to SW620-Scrambled control (2.81-fold, p=0.006, Fig. 2D). Next, tail vein metastatic assay in nude mice was conducted to examine the metastatic ability of SW620-Txl-2si1 cells in vivo. Notably, compared with SW620-Scrambled control, i.v. inoculation of SW620-Txl-2si1 cells led to a significant reduction of visible tumors in liver and lung, which correlated with a lower number of metastasis loci (p<0.05, p<0.01, respectively, Fig. 2E). In addition, mice inoculated with SW620-Txl-2si1 cells presented a prolonged survival time with the median survival time of 35.0 days compared with 26.0 days of the mice injected with SW620-Scrambled control cells (p<0.001, Fig. 2F). Together, these studies demonstrate that Txl-2 promotes invasion and metastasis of colon cancer cells.
Differential effects of Txl-2 isoforms on colon cancer cell invasion and metastasis
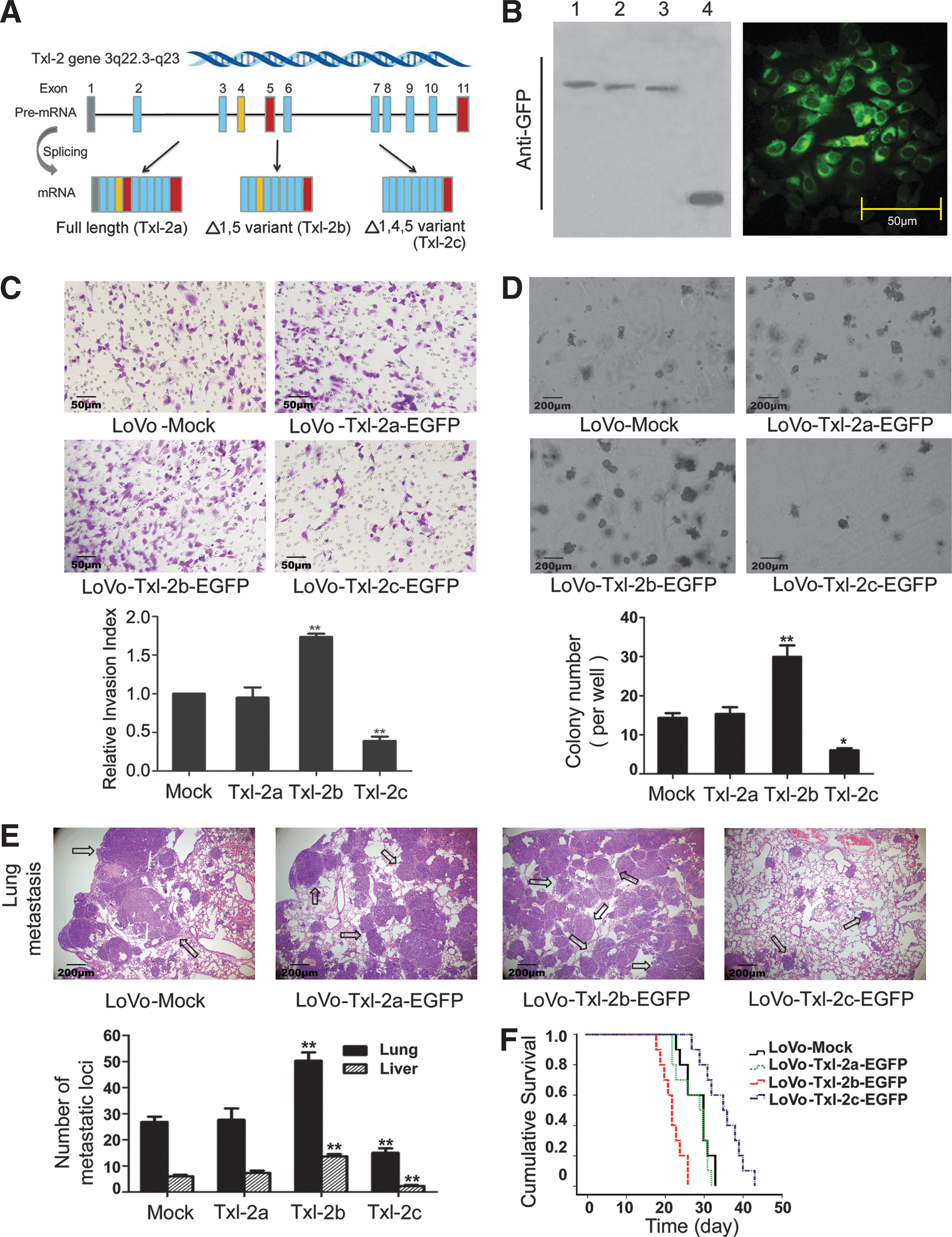
To delineate the distinctive functions of the three Txl-2 isoforms (Fig. 3A) (25, 37), expression constructs of each Txl-2 transcript variant (pEGFP-N3-Txl-2a, b, and c) were stably transfected into LoVo cells, a cell line lacking endogenous Txl-2 expression. The stable expression of the differentTxl-2 isoforms was confirmed by Western blot and immunostaining with anti-GFP antibodies by showing Txl-2a, b, and c-EGFP fusion protein expression, respectively (Fig. 3B). By using these stably transfected cells, we investigated the differential effects of the Txl-2 isoforms on cell invasion (Fig. 3C) and soft agar colony formation (Fig. 3D) that we had observed for Txl-2 siRNA. Among the three Txl-2 isoforms, the strongest stimulatory effect was seen for Txl-2b. In particular, enforced expression of Txl-2b-EGFP in LoVo cells increased the cellular invasion (1.87-fold, p=0.002, Fig. 3C) and increased anchorage independent growth in soft agar (2.10-fold, p=0.07, Fig. 3D). Interestingly, no obvious change was observed for cells transfected with Txl-2a-EGFP, while a reverse effect was noted for cells with Txl-2c-EGFP. Cells with Txl-2c-EGFP significantly suppressed the cellular invasion (2.59-fold, p=0.004, Fig. 3C) and the colony growth (2.38-fold, p=0.01, Fig. 3D) when compared with the control cell lines, respectively. These findings were further supported by in vivo metastatic assay. Inoculation of Txl-2b-EGFP transfected cells through tail vein led to a significant increase in visible tumors in liver and lung which also correlated to the number of metastasis loci (both p<0.01). In contrast, Txl-2c-EGFP transfected cells resulted in less visible tumors in liver and lung (both p<0.01, Fig. 3E). Interestingly, mice inoculated with Txl-2b-EGFP cells presented a shortened survival time with a median survival time of 22.0 days compared to the 30.0 days of the mock control (p<0.001, Fig. 3F), while Txl-2c-EGFP inoculation prolonged the median survival time to 35.0 days (p<0.05, Fig. 3F). These results demonstrate that the most abundant isoform Txl-2b exerts the major effects on promoting cancer cell invasion and metastasis, whereas Txl-2c has an inhibitory role.
The Trx domain of Txl-2b is essential for inducing invasion and metastasis of colon cancer cells
Txl-2 is composed of an N-terminal Trx domain followed by one NDPk domain, both separated by a interface domain (25, 37). To investigate whether both Trx and NDPk domains modulate the functional properties of Txl-2 isoforms in cancer cell invasion and metastasis, we analyzed the role of the Trx domain by generating a redox inactive Txl-2b variant (Txl-2bCS) in which the cysteine residues at the active site of the Trx domain were replaced by serine residues (W

Txl-2b interacts with the small GTPase Ran
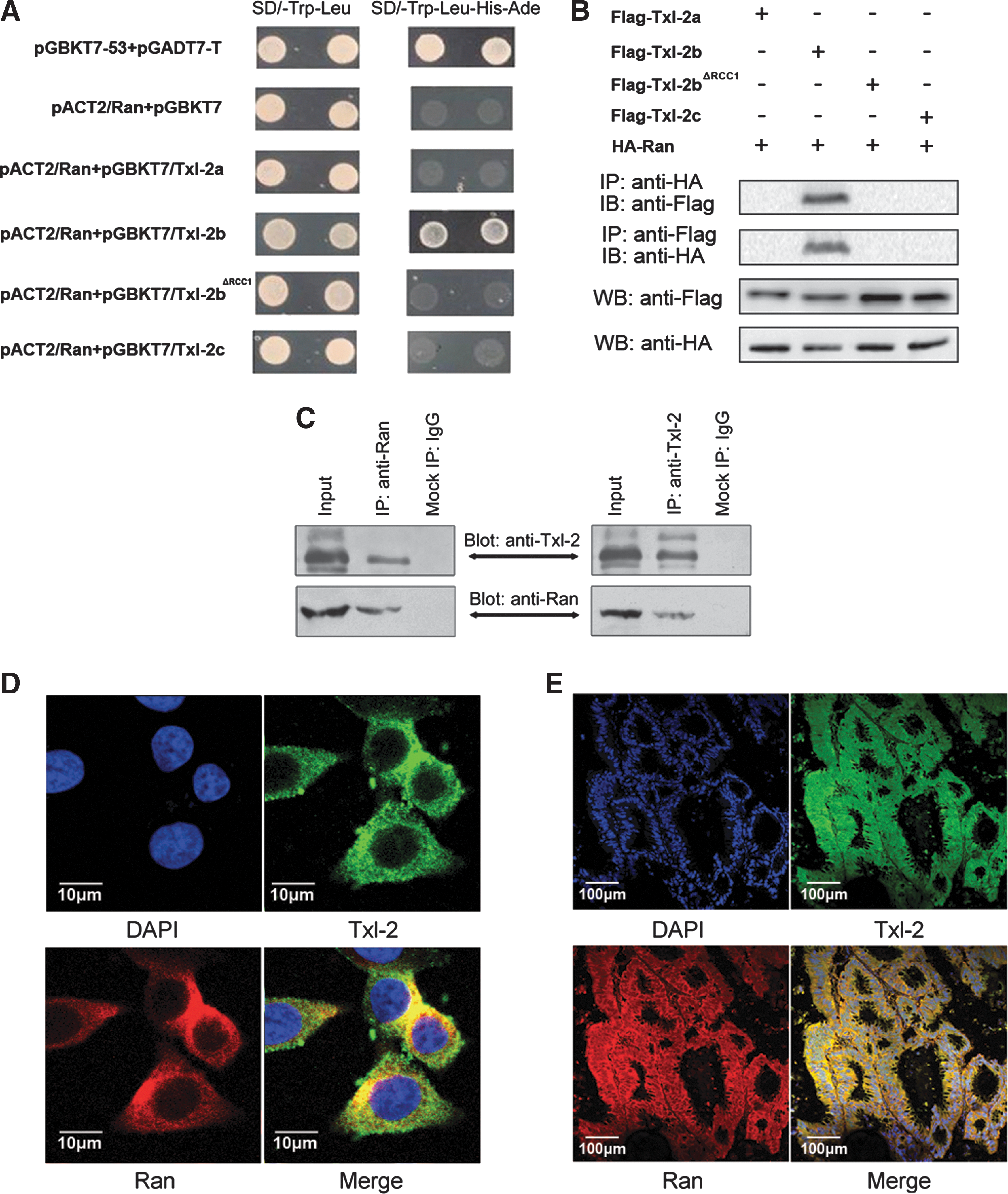
Next, to gain deeper insight into the molecular mechanisms mediating Txl-2 function, we set to identify potential binding partners of Txl-2. A search in the PROSITE database (
A paired immunohistochemical study for Txl-2 and Ran expression in 50 cases of colon cancer tissues.
By chi-square test.
Phospho-Akt mediates the Txl-2b-Ran-matrix metalloproteinase signaling induced tumor cell invasion
Ran is a well-recognized downstream effector of the PI3K signaling pathway that mediates invasion and metastasis of cancer cells (20). Thus, the involvement of Txl-2b interaction with Ran in cancer cell metastasis and the PI3K pathway was further investigated. We found that SW620 cells with decreased levels of Txl-2 showed a lack of Akt activation as detected by a specific antibody to phosphoserine 473 of Akt (Fig. 6A), whereas overexpression of Txl-2b resulted in an elevated Akt activation (Fig. 6B). Interference of Txl-2b and Ran interaction by deleting the RCC1 signature in Txl-2b strongly abolished the enhanced invasive potential of Txl-2b overexpressing LoVo cells, accompanied by a decrease in the levels of matrix metalloproteinase (MMP) and phospho-Akt (Ser 473) (Fig. 6B, D). The same effect was also noticed when downregulating Ran expression by siRNA in Txl-2b transfected cells (Fig. 6D, E, and Supplementary Methods and Suppl. Fig. S1; Supplementary Data are available online at
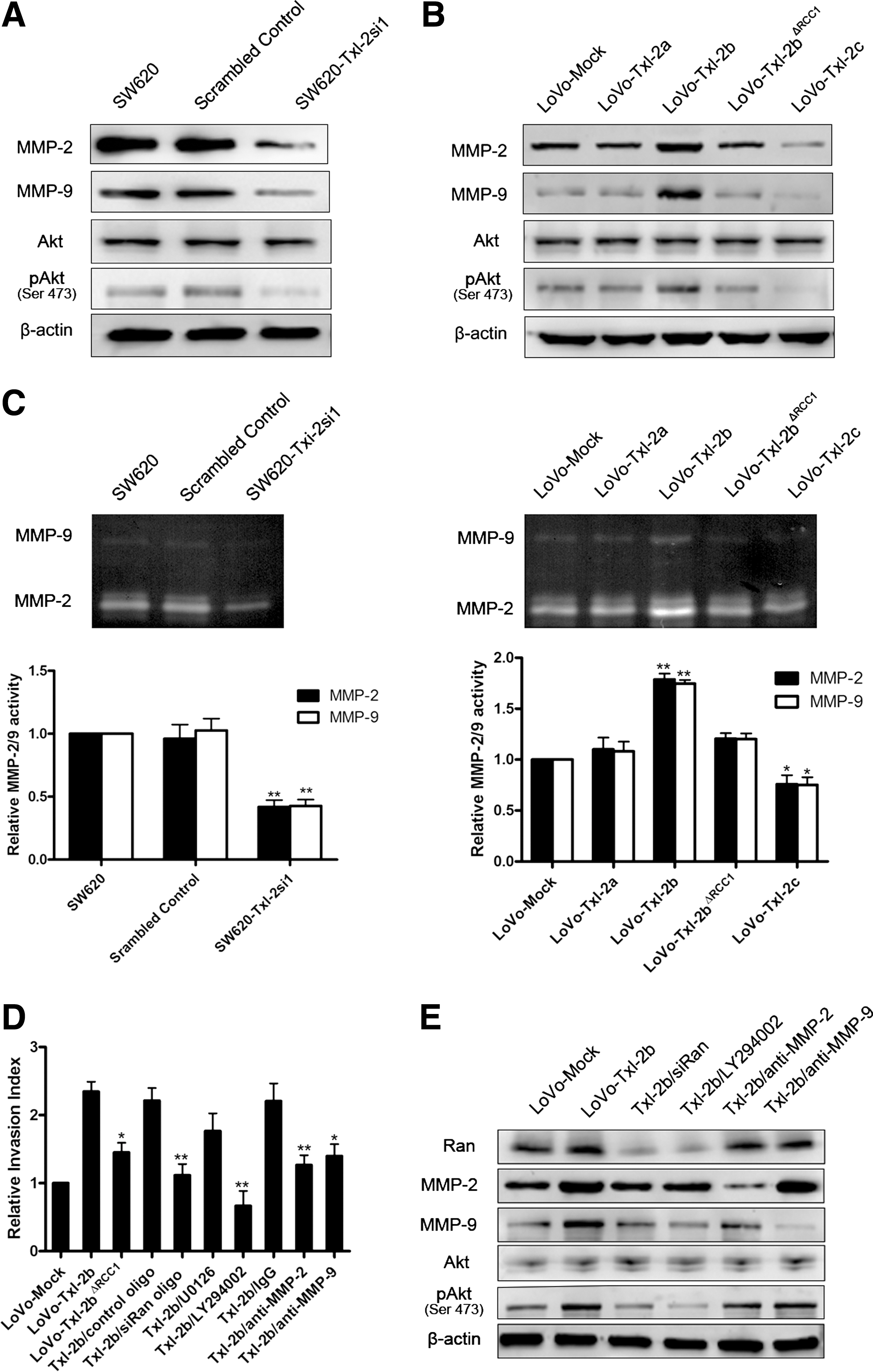
Discussion
In the present study, we demonstrate that Txl-2 plays a key role in colon cancer cell progression and metastasis. Txl-2 overexpression was significantly correlated with the degrees of cancer cell differentiation and tumor staging. More importantly, Txl-2 expression was inversely correlated with survival time of colon cancer patients. Taken together, our data suggest that Txl-2 could be a novel prognostic factor for patients with CRC. Normally, Txl-2 is predominantly expressed in testis and at much lower levels in other somatic tissues, while our previous study has identified Txl-2 in several cancer tissues (25). Therefore, Txl-2 might be a novel member of the cancer-testis antigens family (24), which is characterized by a restricted expression to testis germline cells in normal tissues and their expression being dysregulated in different tumor malignancies.
To explore its functions, we knocked down Txl-2 in the colon cancer cell line SW620, which has a high metastatic potential (19) and performed in vitro and in vivo functional studies showing that depletion of Txl-2 significantly inhibits the metastatic potential of colon cancer cells. Subsequently, the effects of the different Txl-2 isoforms on cancer cell invasion and metastasis were investigated. Interestingly, the most abundant isoform Txl-2b (88%) showed the strongest stimulatory effect on cellular invasion, anchorage independent growth in soft agar, and in vivo metastasis. Compared to Txl-2b, Txl-2a seemed to have little effect but, in contrast, Txl-2c played the opposite function on cancer cells. These results demonstrate that Txl-2b is the major isoform responsible for the Txl-2 function in cancer cell invasion and metastasis. As for Txl-2c, considering that it only appears along with Txl-2b (25), we deem it might function via a negative feedback manner for Txl-2b. Interestingly, alternative splicing is not a rare phenomenon involved in human pathology caused by Trx family. Differential isoforms of thioredoxin reductase 1 (TrxR1) were found in different malignancies and could be a valuable tool in the diagnostics and characterization of tumors (36). Isoform-specific modulation of tumor phenotypes was also reported in other well-studied oncogenic proteins, such as CD44, OPN and Survivin (6, 9, 41).
We investigated the role of the Trx and NDPK domains on the functional properties of Txl-2b by introducing inactivating mutations in the respective active sites of both domains. Notably, only the redox-inactive Trx domain, but not the mutated NDPk domain, strongly inhibited Txl-2b induced invasion and colonization of colon cancer cells, suggesting that the Trx domain plays a prominent role in its oncogenic properties of Txl-2b. The redox activity of Trx resides in the sequence of its conserved active site Cys-Gly-Pro-Cys (CGPC), which undergoes reversible oxidation of the two cysteine residues from a dithiol to a disulfide form (23). Several functions have been discovered for Trxs in human cancer, mostly dependent on its redox activity, including regulation of transcription factor activity, modulation of apoptosis, and regulation of cell motility, angiogenesis, and metastasis (23, 39). We previously found that the dimeric conformation of Txl-2 is maintained by disulfide bonds (37). The fact that an intact redox active site in the Trx domain is essential for Txl-2b induced metastasis demonstrates that Txl-2b may function in cancer cell metastasis in a redox-dependent manner. Nevertheless, we still cannot rule out the possibility of the NDPk domain playing a role in the prometastatic effect of Txl-2b since many reports suggest that the histidine protein kinase activity cannot fully explain the functions of NDPk family members in cancer (2, 45).
We further proceeded to explore the downstream events of Txl-2b induced cancer cell invasion and metastasis. Ran, a small GTP-binding protein from the Ras superfamily, is a well-known molecule that regulates mitotic spindle formation (35). Recently, emerging evidence has been reported on Ran associating with various cancer malignant phenotypes. Activation of the Ran GTPase is subjected to growth factor regulation and can give rise to cellular transformation (26). Ran is an effector of the invasive and metastatic phenotype induced by osteopontin (20) and its overexpression is also associated with local invasion and metastasis in clear cell renal cell carcinoma and epithelial ovarian cancer (1, 3). Our previous study also discovered Ran overexpression in colon cancer (12) and its prometastatic function (unpublished data). In the present study, we found that Txl-2b exerts its proinvasive and metastatic function, at least partially, through its binding to Ran. Among the three isoforms of Txl-2, only Txl-2b binds Ran, as demonstrated by both yeast two-hybrid assay and coimmunoprecipitation. Deletion of RCC1 signature in Txl-2b abolished its interaction with Ran and cancer cells no longer exhibited the enhanced invasive potential, indicating that Txl-2b interacts with Ran via its RCC1 domain and that this interaction is critical for colon cancer cell metastasis. Additionally, inhibition of Ran expression by siRNA oligo also blocked Txl-2b mediated tumor cell invasion and metastasis.
It is not clear why Txl-2b is the only isoform that interacts with Ran. The dimeric conformation of Txl-2 is maintained by disulfide bonds (37) and, therefore, it is reasonable to speculate that Txl-2 could be organized as either homodimer or heterodimer composed by different combinations of the various splicing variants. It is possible then that only homodimers composed by Txl-2b isoforms acquire the necessary spatial conformation to interact with Ran, while homodimers of other splicing variants as well as heterodimers are organized in a different spatial conformation; thus, precluding the interaction with Ran. For instance, the feedback regulatory mechanism proposed above for the Txl-2c inhibitory function in metastasis, could be explained by a possible interference of the Txl-2c hetero/homodimers with Txl-2b and Ran or other effectors. Further studies are required to validate this hypothesis.
Similar to Ras and Rho GTPases, exchange of GDP for GTP leads to a conformational change in the Ran GTPase that greatly enhances affinity to downstream effectors (35). In particular, GEFs facilitate exchange of GDP with GTP to promote GTPase activation (34). It is noteworthy that similar to the action of GEFs, redox agents have been shown to stimulate Ran guanine nucleotide dissociation, which results in an increased rate of GDP-GTP exchange and an accelerated activation of Ran function (17, 18). Besides redox regulation of Ran, growing evidence indicates that NDPk family members also interact with Ras-related family members and influence malignant cancer phenotypes (10, 16, 28, 42). In this context, it is tempting to speculate that Txl-2 acts as a GEF to Ran in a redox-dependent manner and facilitates Ran activity and the downstream effectors, maybe also aided by its NDPk domain.
MMPs belong to a family of enzymes that degrade the extracellular matrix components and, therefore, play an important role in tissue repair, tumor invasion, and metastasis. There is evidence that Trx family members can regulate MMP expression and stimulate tumor cell invasion (13, 31). MMP-2 and MMP-9 are the most important proteases for the degradation of the main constituents of the basement membrane, type IV collagen. Importantly, upregulation of MMP-2 and MMP-9 accelerates cell migration and invasion in CRC (29). In our study, MMP-2 and MMP-9 expression and activity were upregulated in Txl-2b overexpressing cells, accompanied by increased levels of phosphorylated AKT. However, no significant changes were detected in cells overexpressing the deletion mutant Txl-2bΔRCC1, suggesting that MMP-2 and MMP-9 might be regulated by the interaction between Txl-2b and Ran.
Interactions between different transcription factors provide fine-tuning of the transcriptional regulation of MMP promoter activity. The expression of most MMPs members, including MMP-2 and MMP-9, has been shown to be regulated by activator protein 1 (AP-1) and nuclear factor-κB (NF-κB) (8, 14). Interestingly, many Trx family members regulate the DNA-binding activity of transcription factors, such as AP-1, NF-κB, and p53 (23, 33, 39). Besides, NF-κB and AP-1 can also be regulated by some members of the NDP kinase family, such as Nm23-H2 and Nm23-H5 (21, 22). Indeed, we have found increased phosphorylation of the NF-κB kinase IKK in Txl-2b overexpressing cells (unpublished data). It is then possible that Txl-2b, through modulating NF-κB or AP-1 transcriptional activities, induces MMP-2 and MMP-9 gene expression. Dysregulation of PI3K/Akt pathway plays an essential role in cancer development (44). Akt activation was found through Txl-2b and Ran interaction. Since NF-κB and AP-1 are the downstream targets of PI3K/Akt (44), and Akt can also be a downstream target of NF-κB (27), further studies are ongoing to explore the interplay between them.
In summary, our data demonstrate that Txl-2 is overexpressed in colon cancer and its levels correlate with cancer progression, providing evidence that Txl-2 may be a novel effector of invasion and metastasis in colon cancer. Based on our findings, we propose a mechanism by which the splicing variant Txl-2b mediates tumor invasion and metastasis through directly interaction with Ran via PI3K pathway and modulation of MMP-2/9 expression. Inhibition of the Txl-2b-Ran-MMP signaling arises as an attractive target for colon cancer therapy. In normal tissues, Txl-2 is mainly expressed in testis and this work demonstrates that dysregulated expression of Txl-2 promotes cancer progression; thus, categorizing Txl-2 as a putative novel cancer-testis antigen. Cancer-testis antigens are potentially suitable targets for tumor vaccines because of their high immunogenicity in vivo, even in cancer-bearing patients, and their restricted normal tissue distribution (24). Therefore, our data provide a novel biomarker and a target molecule for the diagnosis and treatment of colon cancer.
Materials and Methods
Cell culture and transfection
Human colon adenocarcinoma cell lines SW480, LoVo and human 293T cells were preserved by our institute. Cells were cultured in RPMI 1640 medium (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (Hyclone, Logan, UT), 100 μ/ml penicillin, and 100 μg/ml streptomycin (Sigma-Aldrich, St Louis, MO) at 37°C with a humidified atmosphere of 95% air and 5% CO2. Cell transfection was performed with Lipofectamine ™ 2000 (Invitrogen) as described in the manufacturer's protocol.
Antibodies and reagents
The rabbit anti-Txl-2 polyclonal antibody able to recognize the three isoforms was produced by us previously (37). Goat polyclonal anti-Ran antibody was from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-MMPs antibodies, phospho-Akt pathway antibody sampler kit, PI3 Kinase inhibitor LY294002, and the MEK1/2 inhibitor U0126 were from Cell Signaling Technology (Beverly, MA). Protein A/G agarose was obtained from Calbiochem (Cambridge, MA). Anti-GFP antibody was obtained from eBioscience (San Diego, CA). Anti-Flag and anti-HA antibodies were from Santa Cruz Biotechnology. Anti-βactin antibody was from Sigma-Aldrich.
Tissue collection
Formalin-fixed, paraffin-embedded tumor tissue, and corresponding normal tissue blocks were collected from 243 colon cancer patients who underwent surgical resection during 2004 in the Department of Pathology at the Tianjin Union Medicine Centre, China. All cases of colon cancer and adjacent nontumorous tissues were diagnosed clinically and pathologically. This study was approved by the Ethics Committee of Tianjin Union Medicine Centre. Enrolled patients were followed up at least 5 years for survival calculations.
Immunohistochemical staining
Txl-2 immunostaining was performed by an avidin-biotin method as described previously (25). The expression of Txl-2 was determined by assessing the percentage of decorated tumor cells and the staining intensity semiquantitatively. The percentage of positive cells was rated as follows: 0 (no cells), 1 (1%–25%), 2 (26%–50%), 3 (51%–75%), or 4 (76%–100%). The staining intensity was rated as follows: 0, no signal; 1, weak; 2, moderate; and 3, strong staining. A total score of 0–12 was then calculated and graded as I: score 0 and 1, II: 2, 3 and 4, III: 6 and 8, and IV: 9 and 12.
RNA preparation and semiquantitative RT-PCR
Fifty cases of fresh colon cancer samples were collected, including the adjacent normal tissues. mRNA was prepared using Trizol (Invitrogen) according to the manufacturer's instructions. Semiquantitative RT-PCR and the quantification were performed as previously described (25).
Plasmid construction
pSilencer3.1-H1 (Ambion, Austin, TX) was used for construction of human Txl-2 siRNA expression vectors pSilencer Txl-2 #1 and #2 according to the manufacturer's protocol. The open reading frame of human full-length Txl-2 and Txl-2b were cloned into pGEX-4T-1 expression vector as previously described (37). The products were then subcloned into the vector pEGFP-N3 (BD Biosciences Clontech). cDNA of Txl-2c was obtained by two-step overlapping PCR using two sets primers. For generation of vectors withmutated Txl-2b, cysteine residues in the active site of Trx domain were replaced with serine residues (Cys->Ser) in pEGFP-N3-Txl-2bCS, while in pEGFP-N3-Txl-2bHP, the histidine residue in the active site of NDPk domain was replaced with proline (His->Pro). Double mutant vector of both active sites in Trx and NDPk domain was also generated using the above described mutations and named pEGFP-N3-Txl-2bDM. These site-specific mutations were conducted by PCR using three primer sets. The RCC1 signature (MCSGPSHLLIL) between residues 161 and 171 of Txl-2b was deleted to generate the pEGFP-N3-Txl-2bΔRCC1by PCR. All the primers used in this section are listed in Supplementary Table S1. All the constructs were verified by sequencing in both directions.
Invasion assay
Cell invasion assays were performed using Transwells (8 mm pore size; Corning Costar Corp., Acton, MA) as previously described (20). In some cases, inhibitors or MMP-2 or MMP-9 antibodies (10 μg/ml; Cell Signaling Technology) were also added to the upper and lower chambers.
Soft agar colonization assay
Cells were plated in triplicate in a 24-well plate at 5×103 cells/well, as previously described (20). All colonies composed of greater than 50 cells were counted in each well using a digital imaging system (Olympus Corp., Tokyo, Japan).
Immunoprecipitation
Cells were lysed in a buffer containing 150 mM NaCl, 25 mM Tris-HCl, pH 7.45, 0.5% Nonidet P-40, 0.1% sodium dodecyl sulfate (SDS), 1 mM dithiothreitol, 0.2 mM phenylmethylsulfonyl fluoride (pH 7.5), and a protease inhibitor cocktail. Cell lysates were precleared with agarose beads for 2 h at 4°C and mixed with anti-Txl-2, or anti-Ran-coated protein A/G agarose overnight at 4°C. The agarose was extensively washed with a high-stringency wash buffer containing 0.1% SDS, and the associated proteins were stored for further immunoblot analysis.
Yeast two-hybrid assay
Yeast two-hybrid assay was done using the Matchmaker System (Clontech, Palo Alto, CA) following the manufacturer's protocols. Full-length Ran cDNAs were cloned into the pACT2 plasmid (Clontech) where they were expressed as fusion proteins to the GAL4 activation domain. For the bait protein, Txl-2a, b, and c cDNA and the deletion mutant Txl-2bΔRCC1 were cloned into the pGBKT7 plasmid, respectively, and expressed as a fusion to GAL4 DNA binding domain. Plasmids were cotransformed into AH109 yeast strain and positive clones were observed on high-stringency dropout plates (-Leu/-Trp/-His/-Ade).
Immunofluorescence staining
Cells were plated onto glass coverslips and fixed with 4% paraformaldehyde for 20 min and permeabilized with 0.1% Triton X-100 in phosphate-buffered saline for 15 min. Blocking solution was applied for 1 h at room temperature. Rabbit polyclonal anti-Txl-2 antibody (1: 150) and goat polyclonal anti-Ran antibody(1: 200)were applied at 4°C overnight. FITC-conjugated and Cy3-conjugated secondary antibodies were loaded and incubated for 2 h at room temperature. Immunostaining signal and DAPI-stained nuclei were visualized by the FLUOVIEW laser scanning confocal microscope (Olympus Corp.).
Gelatin zymography
The enzymatic activity of MMP-2 and MMP-9 was determined using a gelatin zymography assay kit (GENMED Scientifics, Inc., Shanghai, China), as described previously (43). In brief, the conditioned medium containing equal amounts of protein was separated on a 10% SDS-polyacrylamide gel electrophoresis containing 0.1% gelatin. Gels were placed in 2.5% Triton X-100 for 30 min, incubated for 24 h in buffer (50 mM Tris-HCl, pH 7.4, 5 mM CaCl2) and stained with Coomassie blue R-250.
Tail vein metastatic assay
Every experimental group had eighteen 4 to 6-week-old BALB/C-nu/nu nude mice obtained from the Shanghai Laboratory Animal Center of China. Each mouse was injected via tail vein with 2.5×106 cells. Four weeks later, 8 mice were sacrificed, and 4 μM paraffin-embedded sections of lung and liver tissues were prepared. The sections were stained with hematoxylin and eosin and examined for the presence of metastatic tumor foci under microscopy. The rest of the mice (n=10 per group) were maintained for survival analysis and studied until they were dead.
Statistical analysis
All statistical analyses were performed using SPSS16.0 software (Chicago, IL). Mann–Whitney U-test for two groups and Kruskal–Wallis H-test for multigroups were used in the analysis of immunohistochemical results. Survival was assessed by the Kaplan–Meier method and statistical analysis performed by Log-Rank testing. Numerical data are presented as the mean±SEM. The differences between groups were analyzed using the Student's t -test when comparing only two groups or assessed by one-way analysis of variance when more than two groups were compared. p<0.05 was considered as statistically significant.
Footnotes
Acknowledgments
We thank Prof. Peisong Gao (Division of Allergy and Clinical Immunology, Department of Medicine, the Johns Hopkins University) and Prof. Jianping Jin (Department of Physiology, Wayne State University School of Medicine) for their contribution to the preparation of this manuscript. We also thank Yunmin Li (Department of Health Statistics, The Fourth Military Medical University, China) for his assistance in statistical analysis. This work was supported by grants from National Key Basic Research and Development Program (2010CB529300, 2009CB521805, 2010CB529306) and National Science Foundation of China (NSF: 81201929, 30971337, 81272650).
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.