
Abstract
Introduction
S
Hypoxic pulmonary vasoconstriction (HPV) allows shifting blood flow from hypoxic to normoxic lung areas, thereby coupling ventilation and perfusion (55). HPV is considered to be responsible for the high pulmonary vascular resistance during fetal life. A similar mechanism for flow matching has been proposed in the fetoplacental vasculature, so that hypoxic vasoconstriction of fetoplacental arteries would divert blood flow to the placental areas with better maternal perfusion (21, 23, 25, 55). On the other hand, the increase in oxygen tension at birth is a key factor stimulating DA constriction, which precedes the permanent closure of the vessel (45, 55). Therefore, oxygen sensing in PA, fetoplacental arteries and DA play crucial roles during fetal life and in the transition to postnatal life.
Some vessels are specialized in detecting and responding to acute changes in oxygen tension. The results from the present study are consistent with a prominent role of neutral sphingomyelinase (nSMase)-derived ceramide in the hypoxic contraction in pulmonary arteries and chorioallantoic arteries and in the normoxic contraction of the ductus arteriosus. These data provide evidence for the proposal that nSMase-derived ceramide is a unifying mediator of acute oxygen-sensing in specialized vascular tissues.
Elucidation of the mechanisms involved in acute oxygen sensing in vascular smooth muscle cells has been the matter of intensive effort. It is now generally accepted that the physiological responses to changes in oxygen requires the existence of an oxygen sensor coupled to a signal transduction system, which in turn activates a variety of effector mechanisms (46, 52, 54). Intriguingly, despite opposite stimuli, hypoxic contraction of PA and fetoplacental arteries and normoxic contraction of the DA appear to share common mechanistic features. For instance, voltage-gated potassium (Kv) channels are inhibited by hypoxia in PA (2, 8, 56) and in fetoplacental arteries (20) and by normoxia in DA (34, 47), which suggest that these oxygen-sensitive K+ channels constitute a conserved effector mechanism in these cells. The inhibition of Kv channels likely contributes to depolarization of the cell membrane and subsequent activation of voltage-gated L-type calcium channels and, accordingly, calcium channel blockers inhibit the hypoxic vasoconstriction in PA (8, 32, 53) and fetoplacental arteries (25) and the normoxic DA contraction (11, 33). In addition, other common mechanisms for HPV and normoxic DA contraction have been reported, such as release of calcium from intracellular stores (24, 27, 37), voltage-independent calcium entry (22, 26) and increase in Rho kinase activity (22, 26, 43).
The chicken embryo is a suitable model for studying developmental vascular biology (13). As compared to other models, the responses to normoxia in the DA (1, 7) and to hypoxia in PA (58) and chorioallantoic arteries (CA), the avian homologue of human fetoplacental arteries, (28) are consistent, robust, and reproducible in the chicken. Moreover, they do not require an agonist-induced pretone which preclude the possible influence of a preconstriction agent on the intracellular signaling mediating the responses to oxygen changes. Interestingly, in the chicken DA, the pulmonary and the aortic parts of the vessel can be morphologically and functionally distinguished. Thus, the pulmonary side (pDA) is a muscular artery which responds to normoxia with contraction, while the aortic side (aDA) is an elastic artery, which responds to normoxia with relaxation (1, 4, 7). This offers an excellent opportunity to comparatively search for an exclusive oxygen-sensitive mechanism in different portions of the same vessel.
Ceramide can be generated in cells via de novo biosynthesis pathway involving serine palmitoyl transferase or be synthesized from membrane sphingomyelin by sphingomyelinases (SMase), which are activated by multiple membrane receptors and nonreceptor stimuli (18). We have previously found that neutral SMase (nSMase)-derived ceramide acts as a critical mediator in the HPV response in rats by increasing reactive oxygen species (ROS) production via NADPH oxidase (8, 14). However, the possible role of ceramide in other oxygen-sensing tissues remains to be explored. Here we show that two opposite stimuli, such as hypoxia (in PA and CA) and normoxia (in pDA) increase ceramide production to promote vasoconstriction in chicken vessels. Moreover, inhibition of nSMase also prevented the HPV and oxygen-induced DA contraction in human tissues.
Results
Hypoxic contraction is reduced by nSMase inhibition in PA and CA
In chicken PA incubated under normoxic conditions, exposition to hypoxia led to a triphasic (contraction–relaxation–contraction) response. The second contraction reached a plateau after 10–12 min (Fig. 1A). To ascertain the role of nSMase in HPV, a second challenge to hypoxia was elicited in the absence (vehicle) or in the presence of an anticeramide antibody (15B4) or the nSMase inhibitor GW4869. Both treatments inhibited the contraction induced by hypoxia in PA (Fig. 1A, B), the inhibitory effects of GW4869 being concentration-dependent (12.5%±5.7%, 25.7%±5.1%, and 56.3%±7.2% for 0.1, 1, and 10 μM). Moreover, downregulation of nSMAse by siRNA also almost fully inhibited HPV (Fig. 1C, D) but had no effect on the contractile response to exogenous addition of bacterial SMase (Fig. 1D). CA responded with a transient contraction during the 10-min exposure to hypoxia (Fig. 1E). Similar to HPV, the hypoxic CA contraction was reproducible and inhibited by GW4869 (Fig. 1E, F). Unlike the hypoxic responses, the contractions induced by endothelin-1 (ET-1, 30 nM) were not affected by GW4869 in either PA or CA (Fig. 1G).
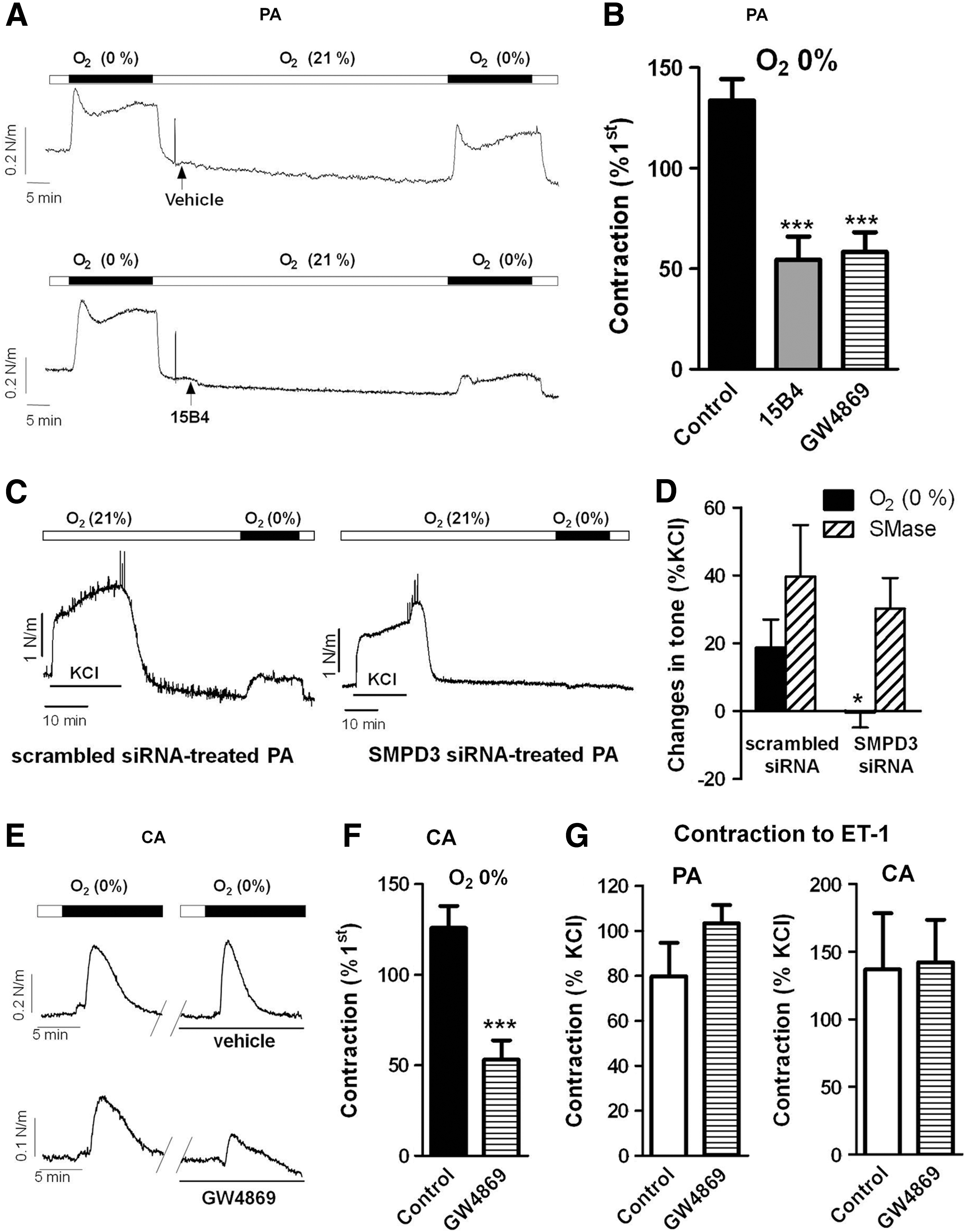
Normoxic contraction of the DA is reduced by nSMase inhibition
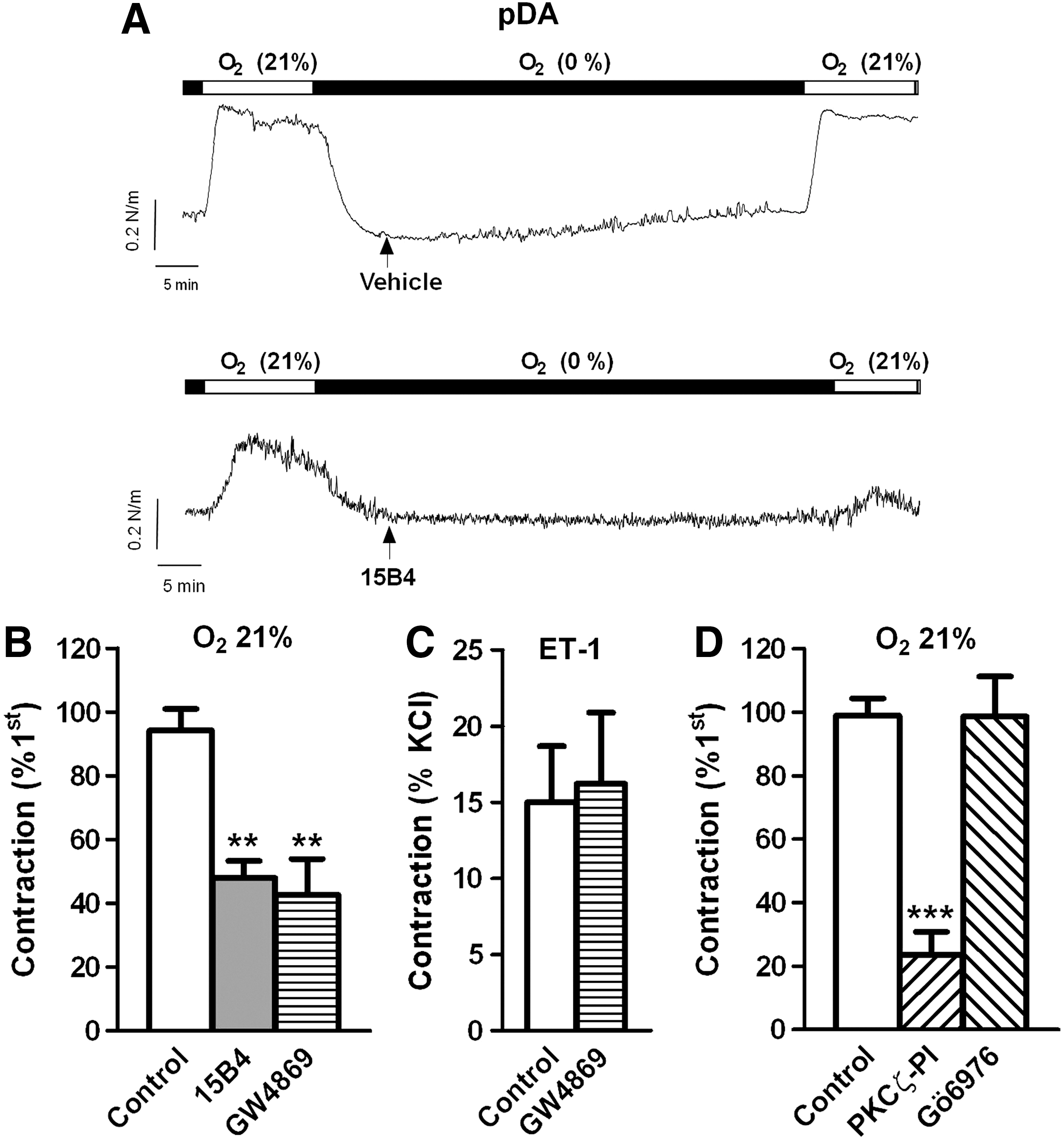
Exposure to normoxia in chicken pDA caused a reproducible contraction, which was inhibited by the anticeramide antibody and the nSMase inhibitor (Fig. 2A, B). Again the contraction induced by ET-1 in the DA was not affected by GW4869 (Fig. 2C). In another set of experiments, the normoxic contraction was tested in the presence of a PKCζ peptide inhibitor (PKCζ-PI, 10 μM) or Gö6976 (0.1 μM) an inhibitor of classic and novel protein kinase C (PKC) isoforms but without effect on the atypical isoform PKCζ. PKCζ-PI reduced the normoxic contraction of the DA, while Gö6976 was without effect (Fig. 2D).
Increase in ceramide content by hypoxia in PA and by normoxia in DA
In freshly isolated PA smooth muscles cells (PASMC) incubated in normoxic conditions, exposure to hypoxia for 10 min led to a marked increase in ceramide content measured by the fluorescence of permeabilized cells immunostained with an anticeramide antibody (Fig. 3A). DA smooth muscle cells (DASMC) isolated from the pDA or the aDA were incubated in hypoxia and exposed to normoxia for 10 min. Interestingly, normoxia markedly increased ceramide content in smooth muscle cells isolated from pDA but not in those isolated from aDA (Fig. 3B). In Figure 3C, the confocal images of nonpermeabilized PASMC show that ceramide can be exposed at the extracellular surface and bound by the anticeramide antibody. The increased in ceramide content after hypoxia in PA was further confirmed using ultra high performance liquid chromatography–mass spectrometry (UHPLC-MS). Figure 3D shows that hypoxia increased significantly the most abundant ceramide (d18:1/16:0) and a similar trend was observed for the other ceramides. A marked increase in this ceramide was also found in PA after exposure with bacterial SMase for 10 min, which served as a positive control (0.08±0.008 vs. 5.8±3.8 ceramide/phosphatidylcholine [PC]) in control and SMase-treated PA, respectively; n=5; p<0.05). Moreover, in PASMC exposed to hypoxia, ceramide levels recovered after 10 min of re-exposure to normoxia.

Increase in ROS production by hypoxia in PA and by normoxia in DA
We have previously reported that the contraction of rat PA in response to acute hypoxia and ceramide is associated with NADPH oxidase-mediated ROS generation (14). Likewise, hypoxia increased ROS production in chicken PA as determined by 2,7-dichlorofluorescein (DCF) fluorescence (Fig. 4A) and this increase was inhibited by the NADPH oxidase inhibitor VAS2870. The inset in Figure 4A shows that hypoxia-induced ROS increase was reversible after returning to normoxia. In the pDA, exposure to normoxia also led to an increase in ROS production (Fig. 4B). When the pDA were incubated with the NADPH oxidase inhibitor apocynin (300 μM), showed a decrease in ROS levels upon the normoxic challenge (Fig. 4B). Furthermore, incubation with apocynin or with the more selective inhibitor VAS2870 similarly reduced the hypoxic vasoconstriction in PA (Fig. 4C) and the normoxic contraction of the pDA (Fig. 4D). In addition, HPV response was markedly impaired by the presence of the mitochondrial electron transport chain inhibitors rotenone (complex I, 30 μM) and myxothiaxol (complex III, 10 μM) as compared to parallel controls (23%±12%, 16%±10%, and 148%±22% of the first hypoxic response, respectively). Likewise, rotenone inhibited hypoxia-induced ROS increase in PA (25%±4% and −11%±15% in control vs. rotenone-treated PA; p<0.05).

Expression of SMPD3
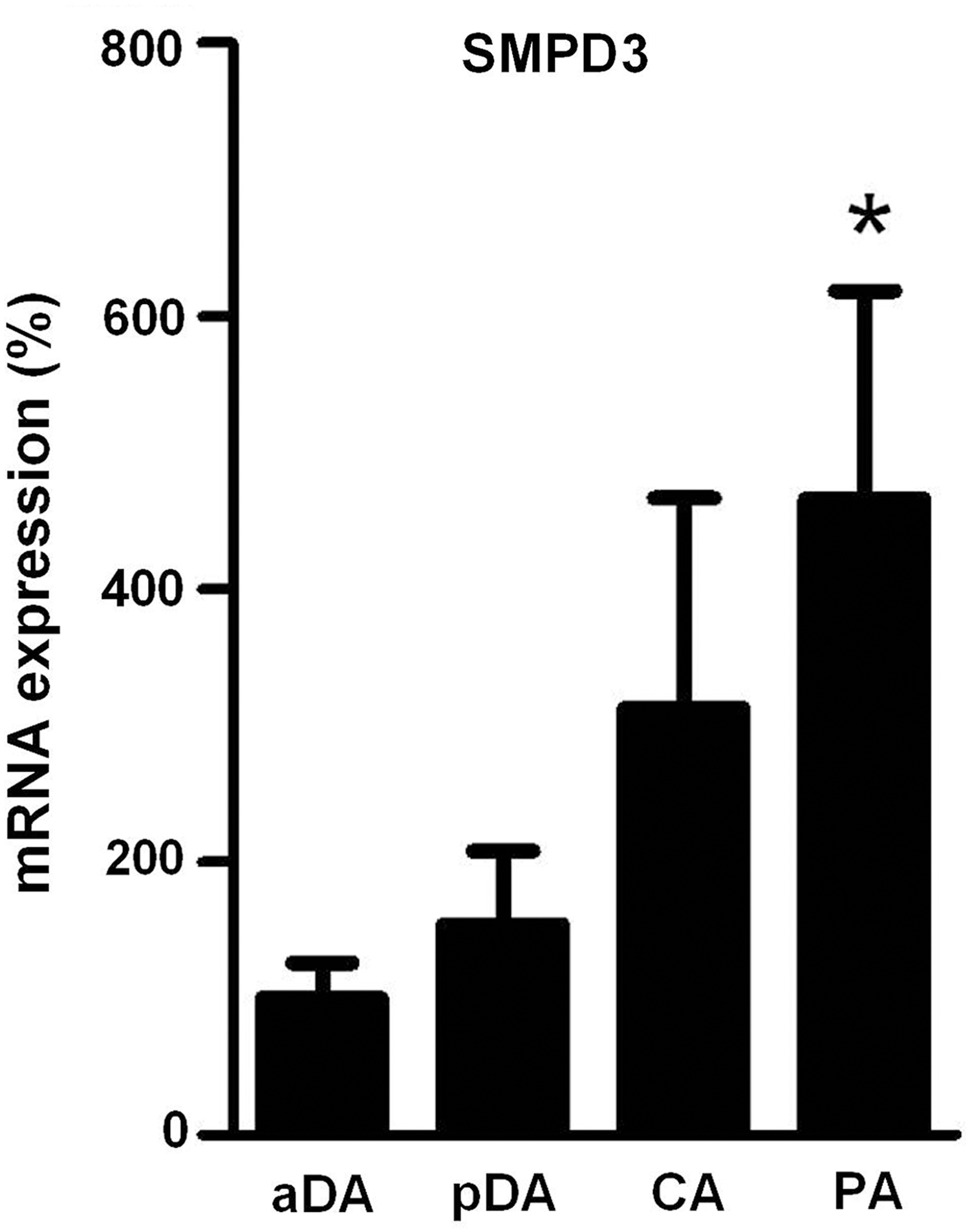
As compared to PA and pDA, aDA was relatively insensitive to changes in oxygen in terms of ceramide production, which could reflect changes in the expression of nSMase. To test this possibility, mRNA levels of nSMase in pDA, aDA, PA, and CA were quantified. Quantitative real-time–polymerase chain reaction (RT-PCR) analysis revealed a reduced expression in aDA as compared to PA. No significant differences were found between aDA and pDA (Fig. 5).
Endogenous and exogenous ceramide mimic the effects of hypoxia in PA and CA, and the effects of normoxia in the DA
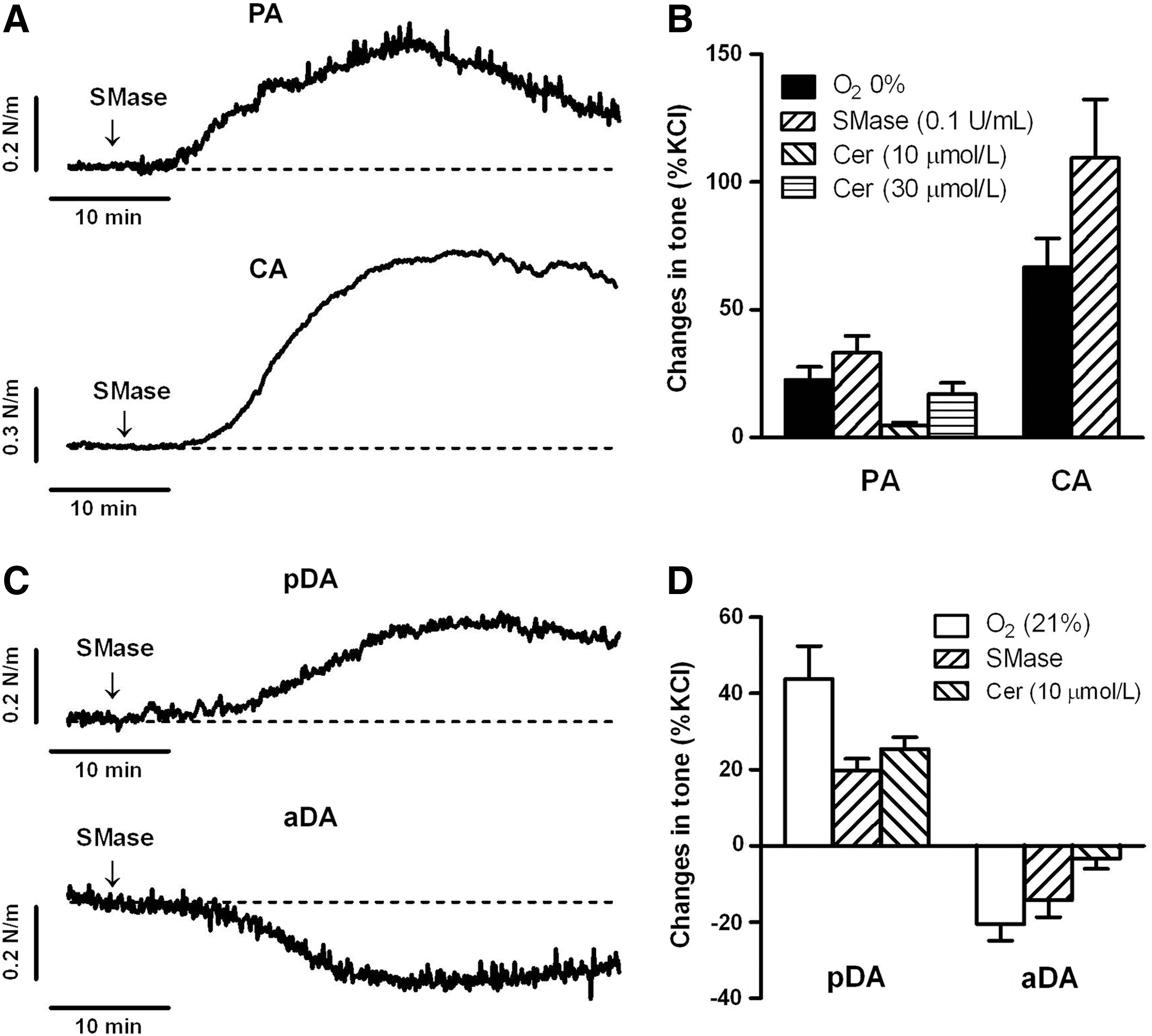
In another set of experiments, vessels were exposed to C6-ceramide (10 or 30 μM) or to SMase from Bacillus cereus (100 mU/ml), that cleaves membrane sphingomyelin and release endogenous ceramide. Addition of C6-ceramide or SMase contracted PA and CA (Fig. 6A), mimicking the effects of acute hypoxia (Fig. 6B). C6-ceramide and SMase also contracted pDA, while relaxed aDA (Fig. 6C), mimicking the responses to normoxia in pDA and aDA (Fig. 6D). In addition, the incubation with the PKCζ-PI (10 μM) reduced the contraction of the pDA induced by C6-ceramide (2.2%±5.2% of KCl-contraction, n=6; p<0.05 vs. control).
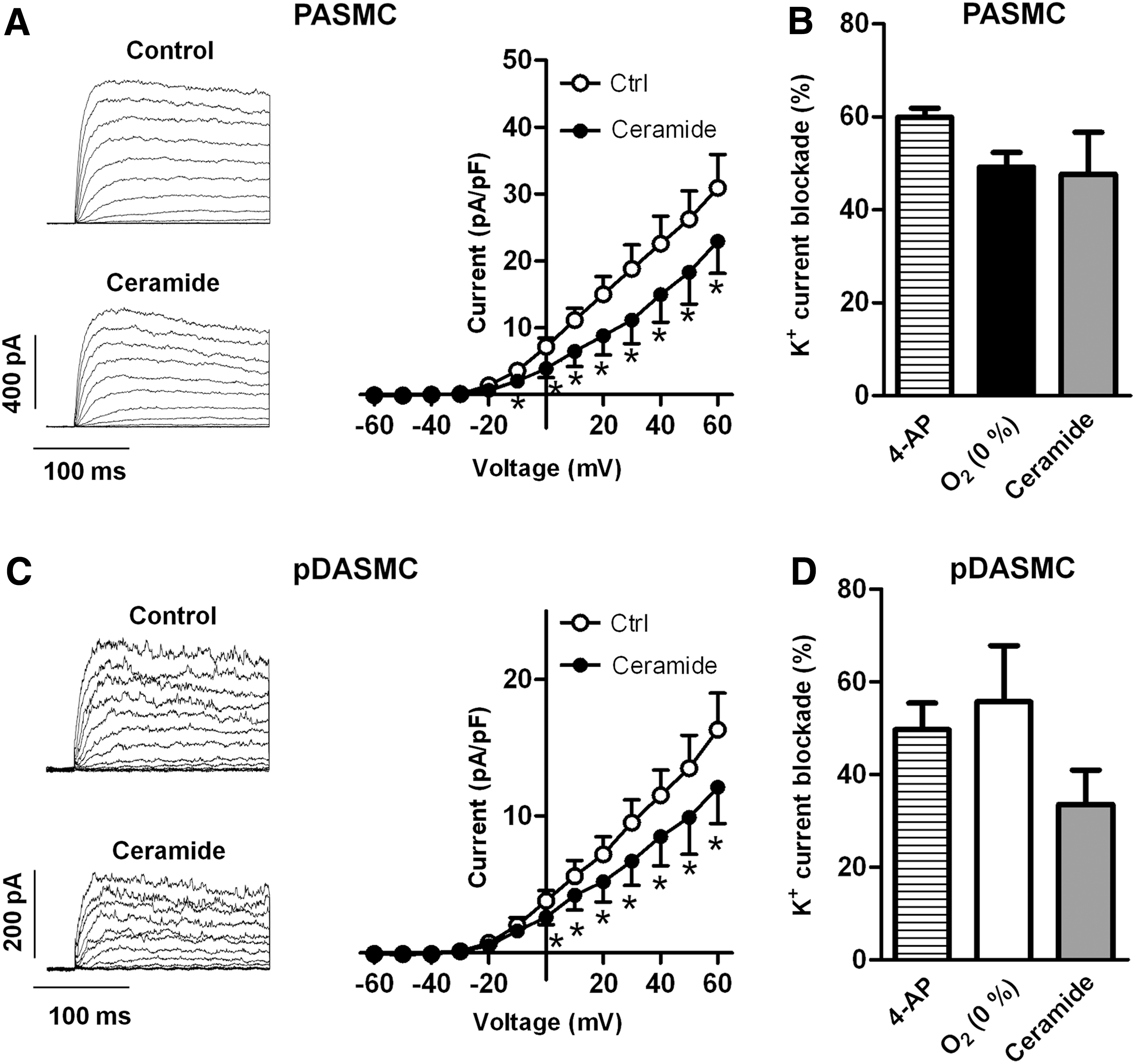
Ceramide inhibits Kv currents in PA and pDA
In agreement with previous studies in rat PASMC, exposure to ceramide (Fig. 7A, B) or to acute hypoxia (Fig. 7B) inhibited Kv currents in chicken PASMC. The inhibitory effects of hypoxia and ceramide were comparable to those elicited by the Kv channels blocker 4-aminopyridine. Interestingly, ceramide also inhibited the potassium current present in pDASMC (Fig. 7C), mimicking the effects of 4-aminopyridine and normoxia (Fig. 7D) in these cells.
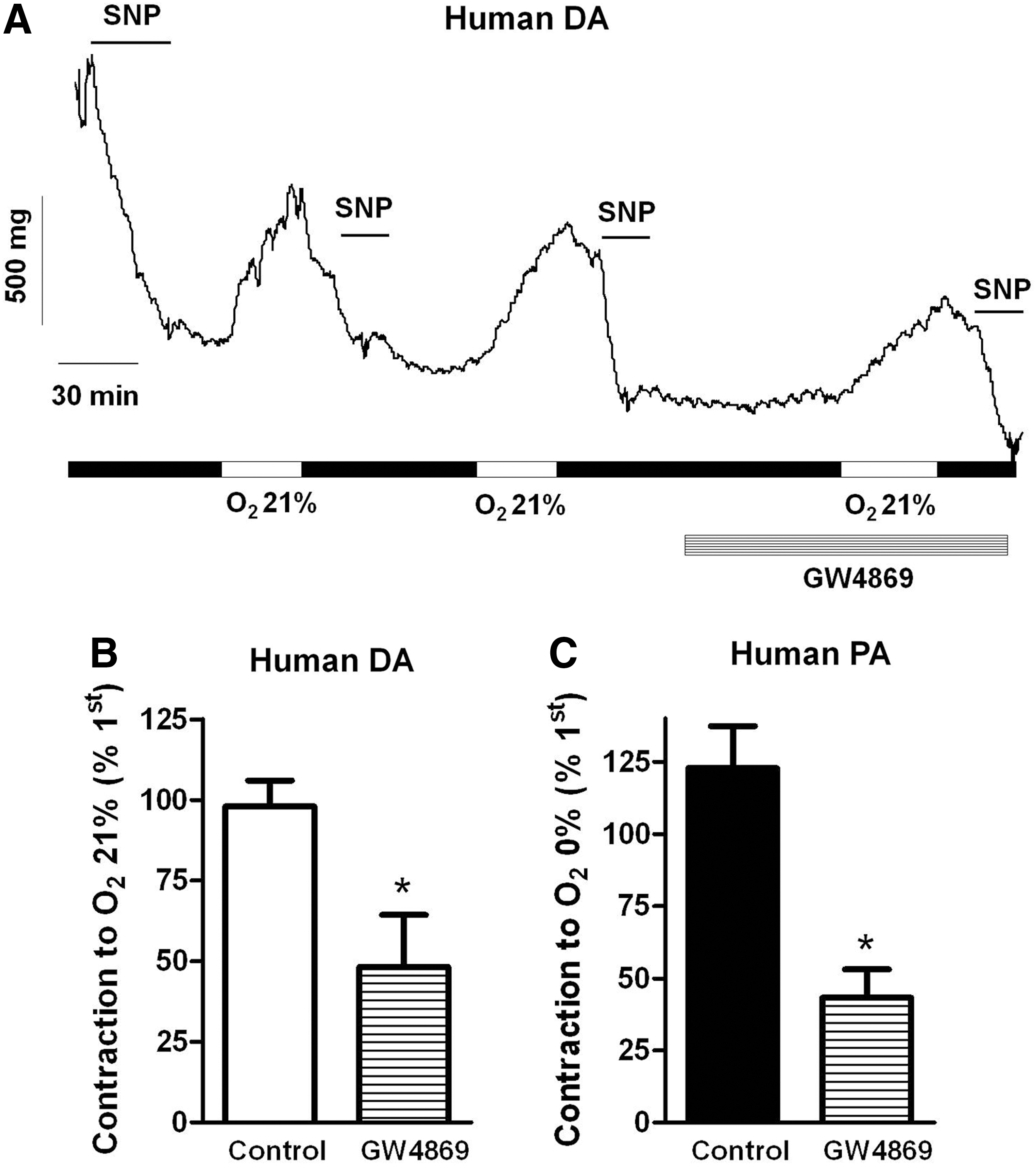
Effects of GW4869 in human vessels
Finally, we tested the effects of GW4869 in HPV and in the normoxic DA contraction in human tissues. The drug inhibited both the normoxic contraction of the DA (Fig. 8A, B) and the hypoxic contraction of the PA (Fig. 8C). Moreover, exogenous addition of ceramide (10 μM) also contracted human DA (22%±5% of the normoxic response, n=4).
Discussion
In the present article, we show that ceramide is involved in acute vascular oxygen sensing. Pharmacological and genetic inhibition of the ceramide-generating enzyme nSMase reduced the hypoxic vasoconstriction in PA and CA and the normoxic contraction of the DA. Moreover, ceramide content and ROS production were increased by hypoxia in PA and by normoxia in DA. Accordingly, ceramide mimicked the contractile responses of hypoxia in PA and CA and those of normoxia in the DA. In addition, ceramide inhibited Kv currents present in PASMC and DASMC. Finally, the role of nSMase in acute oxygen sensing was confirmed in human PA and DA.
Hypoxic PA contraction
HPV allows shifting blood flow from hypoxic to normoxic lung areas, thereby coupling ventilation and perfusion (46, 49). Despite intensive effort, the precise mechanisms involved in HPV have not been fully clarified. While there is general consensus that the oxygen sensor resides in the mitochondria (46, 49, 52, 54), a variety of effector mechanisms have been reported to play a role in HPV, including calcium release from intracellular stores, closure of Kv channels, opening of voltage-gated calcium channels and nonselective cation channels and calcium sensitization via Rho kinase activation (2, 30, 43, 44, 46). However, the most contentious area concerns the signaling mechanisms that link the mitochondrial sensor to the effectors. Thus, some authors propose that ROS decrease during hypoxia, while others propose that ROS increase during hypoxia (49, 51, 54). In previous studies, we have shown that acute hypoxia increases ROS in rat PA through an integrated signaling pathway, which includes activation of nSMase, increase in ceramide production and PKCζ-dependent NADPH oxidase activation (8, 14). Our results showing a NADPH-dependent Kv channel inhibition by hypoxia (8, 14) have been more recently confirmed by Mittal et al. (35). Herein we show that acute hypoxia also increases ceramide and ROS production in chicken PASMC. Moreover, the nSMase inhibitor GW4869, the siRNA against SMPD3 (the gene encoding nSMase2), and the NADPH oxidase inhibitors apocynin and VAS2870 reduced HPV in chicken PA. GW4869 also inhibited HPV in human PA, which suggest that this mechanism of oxygen sensing is widely present in vertebrates. HPV was also inhibited by the anticeramide-antibody which reflects that ceramide may be exposed in the outer leaflet of the plasma membrane since antibodies are not able to cross the membrane. This was confirmed by immunocytochemistry in nonpermeabilized cells. Although it is generally believed that ceramides in the plasma membrane are segregated into rigid domains or rafts, they may be rapidly transferred across the membrane to the outer leaflet and vice versa flip-flop (3, 12, 31). Thus, ceramide generated as a second messenger may recruit its signaling targets (e.g., PKCζ) to the inner surface of the membrane (19) but also be exposed to the extracellular surface.
Hypoxic CA contraction
It has been proposed that, similar to HPV, hypoxic fetoplacental vasoconstriction would divert fetal blood flow toward regions receiving better maternal perfusion (20, 23, 55). Moreover, resembling HPV, inhibition of Kv channels has been reported to contribute to hypoxic fetoplacental vasoconstriction (20). The avian homologous of mammalian fetoplacental arteries are CA which perfuse the late embryonic organ for gas-exchange, the chorioallantoic membrane. Like human fetoplacental vessels (20), chicken CA constrict in response to acute hypoxia (28). Herein we show that the hypoxic contraction of CA is decreased after nSMase inhibition. In agreement with previous reports, our observations argue in favor of a common mechanism for hypoxic vasoconstriction in PA and fetoplacental arteries.
Normoxic DA contraction
At birth, closure of the DA is crucial for the adaptation from fetal to postnatal life. In full term, infants the increase in oxygen tension at birth leads to DA constriction which precedes the anatomical and permanent closure of the vessel (45, 55). Thus, the DA behaves exactly opposite to PA and CA, since it contracts during normoxia and relaxes to hypoxia. Paradoxically, the effector mechanisms responsible for HPV and the normoxic contraction of the DA seem to be virtually the same. These include: Kv channel inhibition (33), Rho kinase activation (22, 26), activation of store operated calcium entry encoded by transient receptor potential cation channels channels (22) and calcium release from intracellular stores (27). In contrast with the discrepancies reported in the HPV field, most evidences support a role of increased ROS in the normoxic DA contraction (55). As mentioned for the PA, the reported mechanisms for oxygen sensing/signaling are similar in mammalian and chicken DA (7, 17). In the present study we show that, similar to HPV, the normoxic contraction of the pDA was attenuated by nSMase and NADPH oxidase inhibitors. Furthermore, oxygen increased ceramide production in SMC isolated from pDA but not in those from aDA, the portion of the vessel that lacks a normoxic contraction (1, 11). In rat PA, we have found that hypoxia-induced p47phox phosphorylation and ROS production were prevented by inhibition of nSMase or PKCζ (14), a well known target of ceramide (5, 36, 40). Likewise, contraction of pDA induced by normoxia and ceramide were reduced by a PKCζ inhibitor.
One potential limitation of our study was the use of the same hypoxic and normoxic conditions for all vessels studied. Our hypoxic conditions yielded pO2 values of ∼3 kPa, comparable to those estimated to occur in the fetal PA and DA (15). However, our normoxic conditions (∼18 kPa), representative of normoxic ventilation, may approach those detected in PA (46, 48), but exceed the pO2 values that the DA sense after the newborn starts breathing.
Ceramide mimics the effects of changes in oxygen concentration
In our previous study, we showed that nSMase2 mRNA expression was ∼10-fold higher in PA compared to mesenteric arteries (8). Thus, the specific increase in ceramide production observed in oxygen sensing in vascular cells could reflect differences at the level of nSMase expression. Herein we found a trend for higher expression in the mRNA expression of SMPD3 in PA, pDA, and CA as compared to aDA, although only the former reached statistical significance. Unfortunately, we could not study possible differences at the protein level due to the lack of commercial antibodies for chicken nSMase2. Therefore, the specific responses observed in oxygen sensing cells may be partly attributable to differences nSMase expression. In a striking similarity with the effects of changes in oxygen tension, addition of SMase and ceramide contracted PA, CA, and pDA, while relaxed aDA. These effects resemble those induced by hydrogen peroxide which inhibits the oxygen-sensitive Kv currents and causes contraction of the PA (14) and the pDA (7), while relaxes the aDA. These results support the notion that oxygen sensitive responses require not only ceramide production but also the presence of exclusive oxygen-sensitive effector mechanisms targeted by ceramide.
Kv channel modulation
Despite the functional contribution of Kv channels in HPV, it is still a matter of controversy, it is generally accepted their modulation by changes in oxygen tension. Therefore, we focus on Kv channels as well recognized oxygen sensing effectors in PA (2, 56), DA (34, 47), and fetoplacental arteries (20). Accordingly, Kv channels were inhibited by hypoxia in chicken PA (present study) and by normoxia in pDA (7). In addition, ceramide inhibited Kv currents present in SMC from chicken PA and pDA, again mimicking the effects of these opposite stimuli. Interestingly, the nature of the Kv channel subunits most likely to generate the oxygen-sensitive Kv current in both PASMC and DASMC appears the same (i.e., Kv1.5 and Kv2.1) (55). It has been proposed that hypoxia decreases Kv current by shifting the PASMCs to a more reduced redox status, while the inhibition of Kv currents induced by normoxia in the DASMC results from a more oxidized environment (34, 53). In agreement with this idea, reducing and oxidizing agents (at relatively high concentrations) mimic the effects of hypoxia in PA and normoxia in DA, respectively (38). A still unanswered question in this model is how a more reduced redox status in the PA and an oxidized environment in the DA lead to the inhibition of virtually the same Kv channels. Moreover, the reducing agent GSH inhibits not only the hypoxic-sensitive Kv currents in PASMC but also the hypoxic-insensitive Kv currents in mesenteric arteries (57). Alternatively, we propose that inhibition of Kv channels by opposite stimuli involves a common mediator (i.e., ceramide and ROS). In agreement with our proposal, not only ceramide (present study) but also t-butylhydroperoxide (at concentrations in the micromolar range), inhibit Kv currents and cause vasoconstriction in both PA and DA (7, 14, 42). Moreover, inhibition of endogenous H2O2 production by increasing intracellular catalase through the patch pipette prevented the inhibition of Kv currents induced by hypoxia in PA and by normoxia in DA (7, 14, 42). Another argument in favor of our proposal arises from the fact that ceramide inhibits the hypoxic-sensitive Kv currents in PASMC but not the hypoxic-insensitive Kv currents in mesenteric arteries (36).
Therefore, based on this and previous reports (8, 11, 14, 36) we propose a common signaling pathway for oxygen sensitive vessels involving the activation of nSMase. The ceramide generated via activation of PKCζ leads to NADPH-derived ROS production. Among other targets, ROS inhibit Kv channels and lead to vessel contraction.
Unresolved issues
Although the identity of the oxygen sensor remains unknown, substantial body of evidence supports the concept that mitochondria acts as the oxygen sensor in specialized oxygen sensing tissues (46, 52, 54). In agreement with this idea, hypoxic contraction in rat (14) and chicken (present study) PA and the normoxic contraction of the DA (7, 17) are blunted by inhibitors of complex I and III of the mitochondrial electron transport chain. Moreover, the complex I inhibitor rotenone prevents hypoxia-induced ceramide and ROS production in rat (14) and chicken (present study) PA. However, the mechanisms matching the sensor to nSMase remain unknown. Intriguingly, nSMase is a redox-sensitive enzyme that can be activated by an increase in ROS (18, 29). Thus, one possibility is that in both situations, hypoxia in PA and normoxia in DA, nSMase is activated by mitochondrial-derived ROS. It is conceivable that normoxia increases mitochondrial ROS generation in the DA by providing more oxygen as substrate. Likewise, hypoxia is able to increase mitochondrial-derived ROS in PA and probably in systemic arteries (52). Therefore, activation of nSMase, production of ceramide and subsequent NADPH oxidase-derived ROS might represent an amplification pathway downstream the sensor (14, 39, 41) present only in specialized oxygen sensing cells.
A number of nonvascular cells, such as the glomus cells of the carotid body, neuroepithelial bodies in the lungs or chromaffin cells of the fetal adrenal medulla are also able to acutely respond to changes in oxygen tension. The results from the present study raise the question of whether nSMase-derived ceramide might also participate in oxygen sensing in these tissues. However, it has been reported that the response to hypoxia in the carotid body is not affected by the nSMase inhibitor GW4869 (16). The possible role of ceramide in other oxygen sensing tissues warrants future studies.
Materials and Methods
Animal experiments were performed in accordance with the Spanish legislation and the procedures were approved by the review board of the Complutense University of Madrid. The Human Studies Committee of Hospital Gregorio Marañón and Hospital General Universitario de Valencia approved the use, after informed consent, of discarded DAs and lung tissue, respectively, excised during surgery. DAs derived from five neonates with hypoplastic left heart syndrome (n=3) or coartaction of the aorta (n=2), while lung tissue was obtained from three adult patients with lung carcinoma surgery.
Reagents
Unless stated otherwise, drugs and reagents were obtained from Sigma-Aldrich Quimica. Ethanol (HPLC-grade) and ammonium (Reagent-grade) were purchased from Panreac Quimica, formic acid (98%) and LC-MS grade methanol were purchased from Fluka Analytical. The mouse anticeramide 15B4 antibody was from Alexis (Grupo Taper). Drugs were dissolved in distilled water except C6-ceramide, GW4869 and rotenone in DMSO and myxothiazol in ethanol. Final vehicle concentrations were ≤0.1%.
Egg incubation and tissue isolation
Fertilized eggs of White Leghorn chickens were incubated at 38°C, and automatically rotated once per hour (Brinsea Polyhatch incubator). Embryos were incubated for 19–20 days of the 21-day incubation period or allowed to hatch, and then transferred to a brooder chick box (Novital Brooder Chick Box, PY333) for up to 3 days. During the grow-out period, chickens were provided ad libitum access to water and a standard starter diet. Animals were euthanized by decapitation, and the DA, the chorioallantoic membrane, or the lungs were removed and immersed in ice-cold Krebs buffer (composition in mM: NaCl 118, KCl 4.75, NaHCO3 25, MgSO4 1.2, CaCl2 2.0, KH2PO4 1.2 and glucose 11). Right and left DA and CA were sampled at 19–20 days of incubation (noninternally pipped embryos), while PA were isolated from 2- to 3-day-old chickens.
Vessel and cell isolation
Vessels were isolated as previously described (1, 11, 28, 58). Briefly, secondary branches of the PA and tertiary branches of the CA were carefully dissected free of surrounding tissue and cut into rings (1.8–2 mm length). The DAs were divided in two segments referred to as pulmonary and aortic DA (pDA and aDA, respectively).
PASMC and DASMC were isolated by enzymatic digestion as previously described (10, 11). For cell isolation, endothelium-denuded vessels were dissected into a physiological salt solution (PSS) of composition (in mM): NaCl 130, KCl 5, MgCl2 1.2, CaCl2 1.5, glucose 10, HEPES 10 (pH 7.3 with NaOH). PA rings were incubated in a Ca2+-free PSS containing (in mg/ml) papain 1, dithiothreitol 0.8 and albumin 0.7 for 5–7 min. DA rings were initially incubated at 4°C in Ca2+-free PSS containing elastase I (0.28 mg/ml) for 5 min. Thereafter, DA was incubated at 37°C in a low Ca2+ (10 μM) PSS containing collagenase I (1 mg/ml), collagenase XI (1 mg/ml), papain (0.15 mg/ml), and ditiothreitol (1.5 mg/ml) for additional 5 min. After enzymatic incubation, tissues were washed in Ca2+-free PSS and disaggregated using a wide bore, smooth-tipped pipette. Cells were stored in Ca2+-free PSS (4°C) and used within 8 h (7, 10). To maintain a hypoxic environment, solutions for DASMC digestion and storage contained the oxygen scavenger sodium dithionite (0.8×10−3 M; pH adjusted to 7.4 with NaOH) (7, 10).
Normoxia and hypoxia
For contractile tension recording, the chambers were filled with Krebs buffer maintained at 37°C. To achieve normoxic conditions, solutions were continuously aerated with 21% O2–5% CO2–74% N2 (pO2=17–19 kPa). Hypoxia was induced by gassing the chamber with 95% N2/5% CO2 (pO2=2.6–3.3 kPa). For the other in vitro experiments, cells and isolated vessels were placed in a chamber of 0.5 ml and perfused with a normoxic (oxygen in equilibrium with room air) or a hypoxic PSS at a rate of 2 ml/min. In these experiments, hypoxia was achieved by vigorously bubbling the PSS solution with 100% N2 in a film coated reservoir. This led to an oxygen concentration of 3%–4% in the chamber as measured with an oxygen Clarck electrode (WPI Instruments).
Recording of arterial reactivity
Contractile responses in endothelium-intact PA, CA, and DA rings mounted in a wire myograph were recorded as previously reported (1, 11, 36). Human DA rings (internal diameter 3–5 mm) were mounted in conventional organ baths. PA and CA were mounted under normoxic condition; while DA rings were maintained under hypoxic conditions (see above). After an equilibration period of 30 min, chicken vessels and human resistance PA (internal diameter 300–400 μm) were distended to a resting tension corresponding to a transmural pressure of 2.66 kPa, while human DA were mounted between two hooks under a tension of 30 mN. Preparations were firstly stimulated by raising the K+ concentration of the buffer (to 80 mM) in exchange for Na+. Vessels were washed three times and allowed to recover before a new stimulation.
In preliminary experiments, we ascertained that vessels were able to respond to successive challenges to hypoxia (for PA and CA) or normoxia (for DA) in the absence of a preconstrictor agent. Therefore, each vessel was exposed to two hypoxic (PA and CA) or normoxic (DA) challenges. The second challenge was examined after 1 h incubation with vehicle (control), anticeramide antibody (15B4, 200 ng/ml), the nSMase inhibitor GW4869 (10 μM), the classic PKC inhibitor Gö6976 (0.1 μM), PKCζ-PI (0.1 μM), the NADPH oxidases inhibitors apocynin (300 μM), and VAS2870 (30 μmol) or the mitochondrial electron transport chain inhibitors rotenone (complex I, 30 μM) and myxothiaxol (complex III, 10 μM) and the contractile responses were expressed as a percentage of the first challenge. In some experiments, the effects of C6-ceramide (10 μM), SMase from B. cereus (100 mU/ml) or ET-1 (30 nM) were tested in the absence and the presence of GW4869. The contractile responses induced by these agents were expressed as a percentage of the initial response to KCl.
Human DA showed and intrinsic contractile tone that was not reversed upon continuous wash-out with hypoxic Krebs solution. Therefore, to study oxygen-induced contractions all human DA rings were initially relaxed by sodium nitroprusside (SNP, 100 μM). Thereafter, DAs were washed three times with drug free Krebs solution. After 40 min equilibration rings were challenged to normoxia. Similar procedure was used before subsequent exposures to normoxia.
Introduction of siRNA by reverse permeabilization
Reverse permeabilization was used to introduce siRNA into isolated chicken PA (6). Briefly, PA were exposed to three successive solutions (4°C) containing (in mM) (i) 10 EGTA, 120 KCl, 5 Na2ATP, 2 MgCl2, 20 HEPES and 50 nM siRNA (pH 6.8; 30 min); (ii) 120 KCl, 5 Na2ATP, 2 MgCl2, and 20 HEPES and 50 nM siRNA (pH 6.8; 180 min); and (iii) 120 KCl, 5 Na2ATP, 10 MgCl2, and 20 HEPES and 50 nM siRNA (pH 6.8; 30 min). Subsequently, PA were bathed in a fourth solution containing (in mM) 120 NaCl, 5 KCl, 5 Na2ATP, 10 MgCl2, 5.6 glucose, and 10 HEPES (pH 7.1, 4°C), in which [Ca2+] was gradually increased from 0.001 to 0.01 to 0.1 to 1 mM every 15 min. Vessels were then placed in DMEM culture medium supplemented with
A mixture of four siRNAs (including one Cy3-labelled siRNA) targeting different regions of chicken nSMase was used. After 24 h incubation, some vessels were examined using fluorescence microscopy to assess siRNA uptake. In preliminary studies, siRNA efficiency was measured by RT-PCR confirming a reduction in SMPD3 mRNA levels by 51%±12% at 72 h, as compared to vessels exposed to random siRNA (scramble).
Immunofluorescent detection of ceramide content
Freshly isolated PASMC or DASMC were allowed to settle on gelatine-coated coverslips for 20 min. PASMC were initially perfused with normoxic PSS for 25 min and thereafter, some coverslips were kept in normoxia and others changed to hypoxic PSS during the last 5 min. On the contrary, DASMC were initially perfused with hypoxic PSS for 25 min, and then randomly changed to normoxia or kept in hypoxia for the last 5 min. Thereafter, cells were immediately fixed with 4% paraformaldehyde, washed with phosphate-buffered saline (PBS) and permeabilized with 0.4% Triton and 3% bovine serum albumin bovine for 1 h. Cells were incubated with 1:30 dilution of mouse anticeramide antibody at 4°C over-night, and then for 15 min at 37°C. Coverslips were washed with blocking buffer and exposed to FITC-conjugated anti-mouse secondary antibody for 2 h a 37°C. After washing, immunofluorescent signals were viewed using an inverted nonconfocal fluorescent microscope. Fluorescence was quantified using ImageJ (ver 1.32j, NIH,
Determination of ROS
The fluorescent dye DCF was used to asses ROS production. Although this dye is far the commonest probe used to detect ROS, it can be oxidized by other free radicals (50). Thus, measurements were compared with parallel time controls, positive controls (t-butylhydroperoxide) and experiments in the presence of ROS generating system inhibitors. Endothelium denuded PA and pDA segments were incubated with the membrane-permeable diacetate form of DCF (DCF-DA, 10 μM) for 60–90 min. Vessels were then placed in the stage of a fluorescent inverted microscope (Leica DM IRB), superfused with PSS (2 ml/min). Preparations were allowed to equilibrate for 30 min under normoxic (for PA) or hypoxic (for DA) conditions, in the absence or in the presence of VAS2870, apocynin or rotenone. Finally, vessels were illuminated through the luminal surface using a 450–490 nm band-pass filter. The emitted fluorescence was filtered using 515 nm long-pass emission filter. Images were taken at 1 min intervals with a Leica DC300F color digital camera. Fluorescence was quantified using ImageJ. Intensity values are reported as a percent of the initial values after subtracting background. After fluorescence values were stable, preparations were challenged with a hypoxic (for PA) or a normoxic (for DA) solution in the continuous absence or presence of apocynin or VAS2870.
RT-PCR analysis
Total RNA was isolated and purified from PA, CA, aDA, and pDA homogenates using RNeasy Fibrous Tissue Mini kit (Qiagen). Total RNA was reverse transcribed into cDNA using iScript™ cDNA Synthesis Kit (BioRad) following manufacturer's instructions. RT-PCR was performed using a Taqman system (Roche-Applied Biosystems) in the Unidad de Genómica (Universidad Complutense de Madrid). Specific primers were designed for the chicken nSMase gene SMPD3.
Lipid extraction and ceramide species measurement
Quantitative evaluation of the proportion of ceramide to the main PC (34:2) was performed by UHPLC-MS. Samples were thawed and 100 μl ethanol added to every tube and vortex-mixed for 5 min. The suspensions were transferred into 0.5 ml microcentrifuge tubes where 5–10 μg of glass beads (acid-washed, 150–212 μm; Sigma-Aldrich) had been prepared. Lipids were then extracted by vigorous shaking with a TissueLyser LT from Qiagen for 15 min, at 50 rpm. Tubes were further centrifuged at 15400 g and 15°C for 20 min, and 80 μl from the supernatant was transferred to HPLC vials with insert. The samples were analyzed with an UHPLC-QTOF MS from Agilent Technologies, equipped with a 1290 series LC system and a 6550 iFunnel QTOF MS detector. About 0.5 μl from each sample was injected (in triplicate) onto the column, a Zorbax Eclipse Plus C8, 2.1×150 mm; 1.8 μm (Agilent Technologies) kept at 80°C. Compounds were eluted with an 8 min linear gradient for the mobile phase at 0.6 ml/min, from 50% ammonium formate 10 mM (pH 6.6) and 50% methanol to 100% methanol. Data files were processed with MassHunter Qualitative Analysis B.05.00 to clean of background noises and unrelated ions by the Molecular Feature Extraction tool. Data pretreatment, including alignment and filtering was performed in MassProfiler Professional (B.12.01; Agilent Technologies). The result was a matrix with all the compounds in the samples sorted by their characteristic retention time and neutral mass, and the abundance of each compound for each sample. The list of exact masses was exported to public databases (METLIN—
Electrophysiological studies
Membrane currents were recorded with an Axopatch 200B and a Digidata 1322A (Axon Instruments) using the whole-cell configuration of the patch clamp technique. Currents were evoked by applying depolarizing steps, normalized for cell capacitance and expressed in pA/pF as previously described (10, 11). Current-voltage relationships were constructed by measuring the currents at the end of the pulse. For recording optimal Kv currents, cells were superfused with an external Ca2+-free PSS (see above) and a Ca2+-free pipette (internal) solution containing (mM): KCl 110, MgCl2 1.2, Na2ATP 5, HEPES 10, EGTA 10, pH adjusted to 7.3 with KOH. All experiments were performed at room temperature (22°C −24°C). Experiments were performed under normoxic or hypoxic conditions for PASMC or DASMC, respectively.
Data analysis
Data are expressed as means±SEM; n indicates the number of samples from different animals or patients. For multiple comparisons, statistical analysis was performed using a one or two way ANOVA followed by a Bonferroni post hoc test, otherwise using a two-tailed Student's t-test for paired or unpaired observations. Differences were considered statistically significant when p<0.05.
Footnotes
Acknowledgments
This work was supported by the Spanish Ministerio de Ciencia e Innovación (predoctoral FPI grant to D.M-C.; research grants SAF2010-22066-C02-02 to A.C.; SAF2008-03948 and SAF2011-28150 to F.P.V.; SAF2011-26443 to J.C.; Juan de la Cierva contract to L.M.), the Spanish Ministry of Education (predoctoral FPU grant to J.M.-S.), Spanish Ministry of Economy and Competitiveness (research grant CTQ2011-23562 to F.J.R.), and Marie Curie European Reintegration Grant within the 7th European Community Framework Programme (PERG05-GA-2009-249165 to F.P.V. and L.M.). A.F. was funded from the European Union 7th Framework Programme (FP7/2007–2013) under grant agreement no. 264864. The authors thank Dr. Coral Barbas for her generous support and advice for the determination of ceramides.
Author Disclosure Statement
No competing financial interests exist