
Abstract
Relevance of Stroke and Lack of Treatment Options
There are various risk factors for stroke, including nonmodifiable ones such as age, race, or genetic background, and modifiable ones such as hypertension, poor diet, smoking, lack of exercise, obesity, hypercholesterolemia, and diabetes (22). In the clinic, stroke is presented in two major forms: either ischemic (∼85% of the cases), in which a vessel becomes occluded by a blood clot or by vascular dissection, or hemorrhagic caused by intracerebral (∼10%) or subarachnoidal (∼3%) bleedings. In this review, we will focus on the more common ischemic stroke.
Already 5 min after the onset of ischemia, neurons begin to die. Therefore, blood flow should be restored as early as possible. For this, recombinant tissue plasminogen activator (rt-PA) is applied intravenously, in order to lyse the occluding blood clot. Indeed, it is the only drug that is currently approved for acute stroke treatment and, significantly, neuroprotective therapies are not available at all. Moreover, only a limited number of patients benefit from rt-PA, as rt-PA may be administered to patients unless several requirements are met. First, a computed tomography (CT) scan should be performed to exclude a hemorrhagic stroke, in which case rt-PA would exacerbate bleeding. Second, rt-PA should be administered within the first 3–4.5 h after the cerebrovascular incident. This, however, represents a challenge given the very narrow time window to recognize the stroke, transportation to the hospital, and completing the CT examination. Third, several additional contra-indications exist, for example, an age older than 80 years, increased risk of bleeding, previous surgeries, and so on. Thus, the use of rt-PA is finally restricted to only approximately 10% of all stroke patients. For this reason, other improved thromobolytic drugs are currently under investigation. They are characterized by longer half life and higher fibrin specificity and, hence, a possibly lower risk of bleeding. However, recent clinical trials were disappointing (38). Therefore, in order to find effective treatment options in stroke, we need to identify novel mechanism-based targets. One such promising candidate mechanism is oxidative stress.
The Oxidative Stress Hypothesis
Oxidative stress refers to a relative surplus of reactive oxygen species (ROS) caused by excessive ROS generation and/or impaired ROS degradation. Under conditions of oxidative stress, the overload of ROS becomes detrimental and possibly contributes to various acute and chronic diseases, such as myocardial infarction, hypertension, atherosclerosis, cancer, neurodegeneration, and inflammation. Consequently, oxidative stress has also been suggested as a key underlying mechanism of ischemic stroke (11).
The brain is particularly sensitive to oxidative stress because of its high oxygen consumption and low oxidative defence capacity (20). In stroke, ischemia-derived nutrient and oxygen deficiency lead to cellular energy failure and neuronal damage. Reperfusion even aggravates this damage, because the freshly arriving oxygen will serve as a substrate for new ROS production (10).
Based on the hypothesis that oxidative stress plays a major role in neuronal damage caused by ischemic stroke, clinical trials using antioxidant treatments in an attempt to scavenge ROS were performed. However, despite promising preclinical results and even a first rather promising, although underpowered, Phase III trial (37), these studies eventually failed to show a significant amelioration in stroke patients (56). Possible explanations for the negative outcome of antioxidants in clinical trials are detailed elsewhere (71).
However, one important consideration is that ROS are not always associated with deleterious effects and under physiological conditions, they appear to also have essential functions, for example, within the innate immune response (24), cellular signaling, control of cellular proliferation and differentiation, oxygen sensing, vascular tone, and angiogenesis. Consequently, interfering with these beneficial roles of ROS in a too broad and unspecific manner can result in pathologies as well, a phenomenon termed as reductive stress.
Since these nonspecific antioxidative therapies were unsuccessful in the clinic, the oxidative stress hypothesis still remains unproven. Nevertheless, the failure of antioxidant trials, even in stroke patients (15), does not disprove the involvement of ROS in ischemia-reperfusion (I/R) injury. Rather, it may demonstrate that a much deeper understanding of the underlying mechanisms and more focused, molecular approaches on relevant molecular sources of oxidative stress are needed.
NADPH Oxidases As a Relevant Source of Oxidative Stress in Stroke
A superior approach for using antioxidants and scavenge ROS in a generalized, nonselective manner would be to specifically inhibit the disease-relevant enzymatic source(s) of ROS formation and leave physiological ROS signaling intact. In the central nervous system, ROS can derive from different sources, including mitochondria, xanthine oxidase, uncoupled nitric oxide synthase (NOS), and cyclooxygenase (68). However, NADPH oxidases are the only known enzyme family with the single known function of producing ROS. All other enzymes produce ROS as a by-product or on uncoupling.
NADPH oxidases are multicomponent protein complexes containing a catalytic NOX subunit that transfers electrons from NADPH to oxygen, thereby forming ROS (6). Five NOX isoforms exist, NOX1-5. Two additional proteins, DUOX1 and DUOX2, also contain an oxidase domain, which in this case is linked to a peroxidase-like domain. Their roles in vascular biology are unclear. NOX homologues differ not only in their tissue and (sub)cellular localization, which is an important determinant of their function, but also in their requirements for regulatory subunits and the nature of the ROS produced (46). Since many reviews on NADPH oxidases have been published, we will not discuss in great detail the characteristics of the different NOX homologues but rather refer to recent reviews (49, 59).
Of the five NOX isoforms, NOX3 is most likely not relevant in I/R injury, as it is mainly expressed in the inner ear but not in blood vessels. In contrast, NOX1, NOX2 (previously termed gp91phox), and NOX4 are expressed under physiological conditions in the central nervous system, including intracranial vessels and neuronal tissues (1, 28). Based on mRNA levels, NOX4 is the most widely distributed NOX isoform in the vasculature. Its expression is an order of magnitude higher in cerebral arteries compared with peripheral blood vessels (42). With regard to the NOX5 isoform, it should be noted that NOX5 is not expressed in rats and mice and is, therefore, functionally less characterized than others. Nevertheless, NOX5 is of considerable interest, as it is both expressed in the human vasculature and regulated in a unique manner, that is, direct binding of calcium, which may be of pathomechanistic relevance, as an imbalance in calcium homeostasis appears to be decisive for neuronal cell death after an ischemic stroke. The absence of NOX5 in the rat and mouse genome is, therefore, a noteworthy limitation of all preclinical studies in these species.
Since specific antibodies for the various isoforms are scarce, most papers are limited to measuring NOX mRNA levels. Since NOX activities are also highly regulated at a post-translational level, protein expression and activity data are much more favorable. For example, increased NOX activity does not necessarily require increased subunit expression, although correlation of mRNA levels of NOX2 or NOX4 and NADPH oxidase activity has been observed in transgenic mice (4). After an ischemic stroke, NOX2 and NOX4 are up-regulated within 24 h. It appears that in this process, NOX2 protein levels increase within microglia (23), and NOX4 protein levels increase in neurons and endothelial cells (33, 65) (Fig. 1). This up-regulation might give a first indication of the implication of NOX2 and 4 in stroke pathology, and, hence, these two NOX isoforms present a potential target in stroke therapy. Therefore, several research groups started investigating the role of NADPH oxidases in stroke.
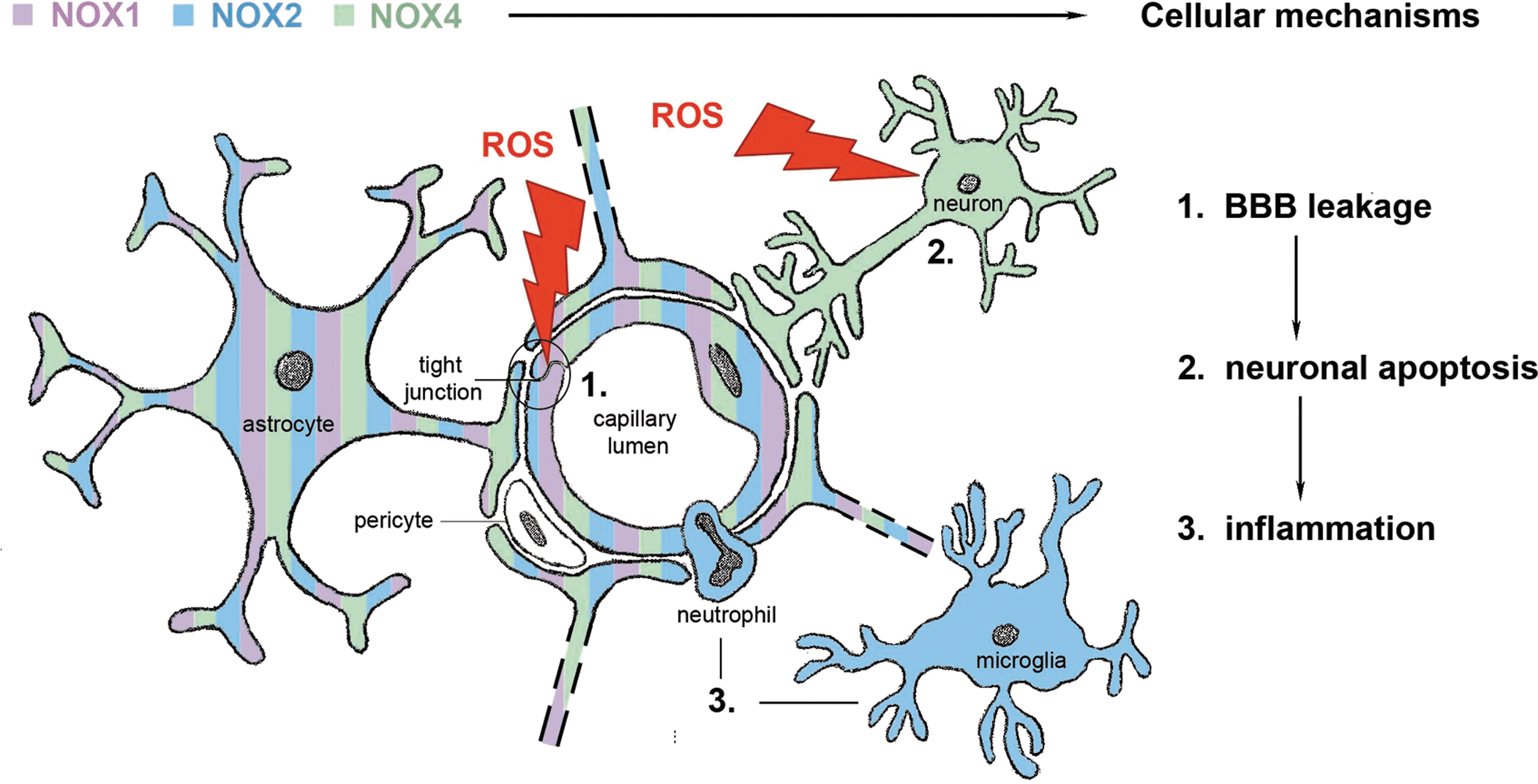
Lack of Quality Control in Preclinical Stroke Research
In 1996, rt-PA treatment has been approved for clot lysis in stroke. Since then, no innovative stroke treatment has made it to the clinic. O'Collins et al. provided an overview of the numerous attempts of stroke researchers to develop neuroprotective drugs. They come to the alarming conclusion that 1026 experimental treatments have been investigated, from which 114 have even been clinically tested in stroke patients, but that none of them has fulfilled their initial expectations (44). The main reason is a quality issue with most, if not all, preclinical stroke studies.
In preclinical models, stroke is induced artificially in young, healthy animals; whereas in humans, stroke happens in older patients, often as a consequence of several underlying pathologies (e.g., hypertension, atherosclerosis, atrial fibrillation, etc.) (19), and spontaneous recanalization can occur. Thus, neither permanent nor transient animal models of ischemic stroke fully mimic the clinical situation.
The majority of studies are performed in rodents due to economical and ethical reasons, whereas pigs and primates would be better suited because of their brains' higher proportion of white matter and gyrencephalic structure, which is more similar to humans (26). Moreover, commonly used preclinical ischemic stroke models are diverse. The method of vessel occlusion, for example, differs from mechanical or thermal to embolic or chemical (Fig. 2). Most preclinical studies use the mechanical model of transient middle cerebral artery occlusion. In addition, in many studies, the observation period stops after about 24 h poststroke. This per definition will not address long-term morbidity, mortality, and relevant neurological outcomes.

Significantly, these limitations are not specific to stroke research and hold true for almost all cardiovascular indications, where progress is poor in general. All major drug groups have run out of patent, documenting an obvious 20 year-long innovation block. Thus, the successful translation of preclinical research, in general, requires conceptual changes and quality improvement similar to clinical research in the 70s.
Quality Management in Preclinical Stroke Studies
A group of basic and clinical researchers along with representatives from the pharmaceutical industry formed the Stroke Therapy Academic Industry Roundtable (STAIR) to design preclinical study criteria that should be followed to improve the quality of stroke studies and their later translation into practice (60). Since human populations are more heterogeneous compared with laboratory animals and most stroke patients have diverse comorbidities, experimental animals should ideally also present comorbidities in order to better mimic clinical conditions. Furthermore, old animals should be included, because stroke patients are also mostly elderly people. At present, however, preclinical studies mainly use young animals, presumably to reduce time and costs. Significantly, not only age but also gender plays a role in stroke pathology and/or drug metabolism. Thus, both male and female animals should be assessed in stroke studies. Further aspects that may have hampered translation from animal to clinical studies are the endpoints that are assessed in animals. While the infarct size is often the major investigated parameter in animals, neurological outcome is the most important endpoint in humans. Accordingly, functional studies should be performed up to several weeks postcerebral I/R and survival monitored to meet the criteria of clinical trials, rather than stopping the observation period 24 h after ischemia. In addition, at least one permanent and one transient model should be performed to validate a stroke mechanism. Other quality parameters include the verification of reduced blood flow on vessel occlusion (<0.12 ml/g/min) and/or reperfusion, as well as monitoring rectal temperature, because hypothermia likely has neuroprotective effects in experimental models (26), although there is actually no evidence from clinical trials (14). The most important quality criteria for preclinical stroke studies are summarized in Figure 3. Of course, these strict requirements only apply for true translational and not for early proof of principle or purely mechanistic studies.

Similar to all preclinical experiments, randomization (only performed in 36% of all stroke studies), allocation concealment (11%), and blinded analysis of the results (29%) should be performed (54). Mortalities per experimental group need to be explicitly stated, as this is both a quality criterion for the animal surgery and may distort data representation when only the phenotype of the surviving animals is reported.
Moreover, it has recently been recommended to conduct preclinical studies both internationally and multicenter (16, 17). Finally, another aspect that should not be neglected is publication bias (55). Negative findings are often not publishable or more difficult to publish, and the literature is, thus, in favor of positive results, which may overestimate efficacy of any experimental therapy.
Knockout Models to Validate the Role of NOX in Stroke
Knockout (KO) models are powerful tools for determining the role of different NOX isoforms, allowing assessment of a causality-effect relationship. A general artifact is that in KO mice the gene is deleted from the onset of stroke whereas any therapy is likely to initiate only hours after the ischemic event. Apart from this, even truncated proteins or splice variants with residual activity may be generated, depending on the KO strategy. When taking the example of our own NOX4 KO model, we deleted the exons that code for the protein region which is essential for NADPH binding (Fig. 4). Thus, NADPH oxidase activity will be zero, irrespective of any truncated protein being present (33). In addition, abolishing the expression of one NOX isoform may result in counter-regulation of other NOX isoforms and ROS formation. Moreover, most NOX proteins have several binding partners such as the highly abundant p22phox. It is unclear whether p22phox's sole function relates to NOX. If not, the lack of one binding partner of p22phox may have bystander effects on other p22phox-binding proteins. Moreover genes flanking the target gene or genetic background (due to random recombination) may also lead to misinterpretations (18). Most genetically modified mice have been generated on a 129 substrain, which was the first to be easily genetically manipulated but has poor breeding efficacy. Therefore, embryonic stem cells from a 129 strain are implanted into blastocysts of the well-breeding C57BL6 strain. The resultant chimeric mouse will always have some remaining genetic information from the 129 strain, even after extensive backcrossing onto C57BL6 mice and extensive SNP analysis. Thus, it cannot be excluded that some NOX KO mice display a phenotype that is mistakenly attributed to the deleted NOX gene. In this context, also the sub-strain on which the mice have been backcrossed, that is, C57BL6J versus C57BL6N showing different mitochondrial abnormalities, may represent an important factor. Finally, not all publications clearly state whether littermate mice have been used as wild-type controls.

No implication of NOX1
With regard to a possible role of NOX1 in stroke, three studies have been published that all used the same NOX1 KO mouse strain (30, 31, 33), although not all studies reported details according to the STAIR criteria. Jackman et al. concluded that NOX1 is relevant for angiotensin-stimulated ROS production but has no impact on overall ROS production, stroke size, and neurological outcome (30). However, after a subgroup analysis, the authors postulated that cortical strokes were four times increased in NOX1 KO versus control mice, possibly indicating that NOX1 may play a protective role by limiting cortical damage. Another group reported opposite findings, that is, reduced infarct size in NOX1 KO mice (31). This effect was only visible when the ischemic period lasted 1 h but not when lasting longer. Interestingly, antioxidant treatment showed additional neuroprotective effects in NOX1 KO mice and reduced infarct size to the same extent as in apocynin-treated wild-type mice. Thus, the effect of scavenging ROS by antioxidants was unlikely mediated by NOX1 but rather by other NOX isoforms or non-NOX sources of ROS. We observed neither a difference in infarct volume nor in neurological behavior when investigating NOX1 KO mice in stroke (33). Collectively, NOX1 appears to have no or only a very minor effect on neurological damage after stroke. This is consistent with the fact that NOX1, at least at the mRNA level, is not up-regulated after cerebral I/R.
NOX4, the key player
Using our NOX4 KO mice, we unambiguously showed that NOX4 is a pathologically relevant NOX isoform in ischemic stroke in mice (33). In this study, we tried to adhere to the STAIR criteria as much as possible, for example, by using mice of different ages and both gender, transient and permanent stroke models, and a long follow-up period, including evaluation by magnetic resonance imaging. The deletion of NOX4 dramatically protected mice from brain damage, resulting in a 75% reduction in infarct size, as did intrathecal application of a pharmacological NOX inhibitor in wild-type mice (33). Significantly, the drug was applied after the induction of ischemia, which from a clinical point is therapeutically much more relevant than the prevention of brain damage before the onset of ischemia as is the case in KO mice. Certainly, the intrathecal route of administration is not optimal, and an intravenous administration would be much more desirable but was not possible due to pharmacokinetic limitations of VAS2870, the inhibitor used in our study. Moreover, this compound may have potential off-target effects (61), which may make it less attractive for further development into a drug.
However, that NOX4 is a promising target in stroke was recently confirmed by another group using a transgenic mouse model overexpressing NOX4 in endothelial cells (5). Their findings are supportive of those seen in our global NOX4 KO in that they found increased infarcts when compared with wild-type mice. Mechanistically, this was hypothesized to be due to suppression of endothelial NOS by NOX4 (5). In accordance with these findings and the role of endothelial NOX4, infarcts are smaller in endothelial cell-specific NOX4 KO mice compared with wild-type littermates (Radermacher, Kleinschnitz and Schmidt, unpublished observations).
It was recently suggested that NOX4-derived hydrogen peroxide might also have a beneficial role in the cerebral circulation by mediating cerebral vasodilatation and increasing blood flow. However, this effect should be selective for the ischemic zone, as a general cerebral vasodilation would rather be harmful because of the “steal effect,” a phenomenon that refers to the natural principle of the flow collateralization of arterial occlusions. To weigh the degree of potential harm versus the benefit of inhibiting NOX4 and to approve or reject this hypothesis, the effects of different NOX isoform-specific inhibitors at different time points after stroke and also in different species will be of interest. At present, all available data clearly point toward an essential pathophysiological role of NOX4 in ischemia-induced brain damage.
An accessory role for NOX2?
NOX2, a key enzyme of the innate immune and inflammatory response (48), is thought to be involved in almost every cardiovascular disease setting (58), including I/R injury (39). NOX2 was the first NOX homologue that was discovered. Several studies have been performed to study the role of NOX2 in stroke (8, 12, 13, 29, 32 –34, 63, 66). These studies used the same commercially available NOX2 KO mouse strain (strain No. 002365 from Jackson), which had already been generated in 1995 (47). Two of the studies (34, 66) reported the use of wild-type littermates, while the remaining groups used the recommended control animals from Jackson (strain No. 000664).
The purpose of the first published NOX2 stroke study was to elucidate the role of NADPH oxidases in neutrophil-derived ROS in stroke injury. The authors investigated whether deleterious NOX2 originates from influx of peripheral leucocytes and/or from activation of on-site microglia. Although brain lesions were reduced by 46% in NOX2 KO mice when compared with wild-type littermates, the amount of neutrophils migrating into the brain was the same in NOX2 KO and wild-type mice. This raised the question whether NOX2 implicated in brain damage originates from brain tissue (neuronal or vascular origin) or from inflammatory cells. Therefore, the authors transplanted bone marrow from wild-type into NOX2 KO mice and vice versa, thus eliminating the influence from circulating neutrophil-derived NOX2. Either way, they observed no difference in infarct size, concluding that the detrimental role of NOX2 involves the presence of peripheral neutrophils in the infarcted brain (66). Fourteen years later, Tang et al. repeated these experiments (63). Unlike the procedure used in the first study of Walder et al. (66), Tang et al. assessed different time points after bone marrow transplantation and shielded the animal's head to avoid a confounding induction of neuronal damage and microglia activation by irradiation. They concluded that NOX2 from peripheral immune cells contributes more to neurodegeneration than does microglia-derived NOX2 (63).
In NOX2 KO mice, the peak levels of ROS formation (72 h poststroke) were attenuated and so were infarct volumes (34). However, ROS were still detectable at that time, and other sources of ROS (such as other NOX homologues) may still play a role. With regard to infarct size observed in NOX2 KO animals, the published data are conflicting. Most studies suggest a protective effect of NOX2 deletion on stroke outcomes. Initially, we could not reproduce these findings, although we used a high number of animals and followed the STAIR guidelines (33). More recently, we performed further stroke experiments in an older cohort of NOX2 KO mice (20 weeks) in two different labs, amounting to a total of n=21 wild-type and n=14 KO mice. Pooling data from both centers, we observed a small reduction in infarct size (Radermacher et al., submitted for publication). Thus, the NOX2 isoform seems to play a role in stroke pathology in elderly mice, probably via its role in inflammatory processes. However, the role of NOX2 appears to be more modest when compared with that of NOX4. This and a possible publication bias against studies with no or smaller effect sizes (45) may explain some of the conflicting literature. Based on these divergent results, a meta-analysis including all stroke studies performed in NOX2 KO mice would be of interest. It may be that there are other studies that did not find any protection by deleting NOX2, which have not been published. Significantly, such a potential publication bias should be revealed by a funnel plot. This graphical method plots the result of each study (positive or negative) against the sample size of the same study, which—in the ideal case—results in an inverted funnel shape peaking at the real effect size. An asymmetry of this funnel indicates that studies are missing, either because they were not found by the search strategy of the systematic review or as they have not been published. This is important, because a meta-analysis will only pool published data, thereby making a null effect seem real. This analysis should not only be applied for the case of NOX2 KO mice, but would also be interesting for NOX1 and NOX4 KO mice, once more stroke studies have been published.
One important confounder that may explain the qualitatively different results could be inflammation. In human stroke, the inflammatory response is thought to be at least partly initiated to remove dead tissue. Factors that could potentially influence the extent of inflammation and contribution of NOX2 are the degree of aseptic surgery or a housing effect (NOX2 KO mice are immune compromised and develop granulomas). The more prominent the inflammation is, the more prominent the role of NOX2 may become. Possibly, the inflammation may also have the opposite effect by distracting neutrophils that are removed from dead tissue to fight bacteria. Interestingly, a paradoxical anti-inflammatory effect of NOX2 was observed in autoimmune diseases (27), which further complicates the considerations just mentioned.
After the publication of updated STAIR criteria, some research groups started re-investigating the role of NOX2 under influence of gender or comorbidities. Brait et al. (8) showed that NOX2-dependent ROS production from T-lymphocytes after stroke is increased during reperfusion, and that NOX2 only affects lesion size in male but not in female mice, possibly because of estrogen having anti-inflammatory effects. Since hypercholesterolemia is a frequent comorbidity in stroke patients, another group tested the neuroprotective drug betulinic acid in hypercholesterolemia-developing ApoE KO mice, which had larger infarcts than wild-type mice. Betulinic acid seemed to protect against stroke, probably via NOX2, neuronal NOS, and inflammatory NOS down-regulation (40).
In conclusion, it is likely that both isoforms, NOX2 and NOX4, are, to some extent, implicated in cerebral I/R injury, depending on as yet unclear parameters. However, due to higher neuroprotection that we have observed in NOX4 KO mice and due to potential immunosuppressive effects of NOX2 inhibition, we believe that NOX4 is the more promising target for stroke therapy.
Pharmacological Approaches to Inhibit NOX Activity
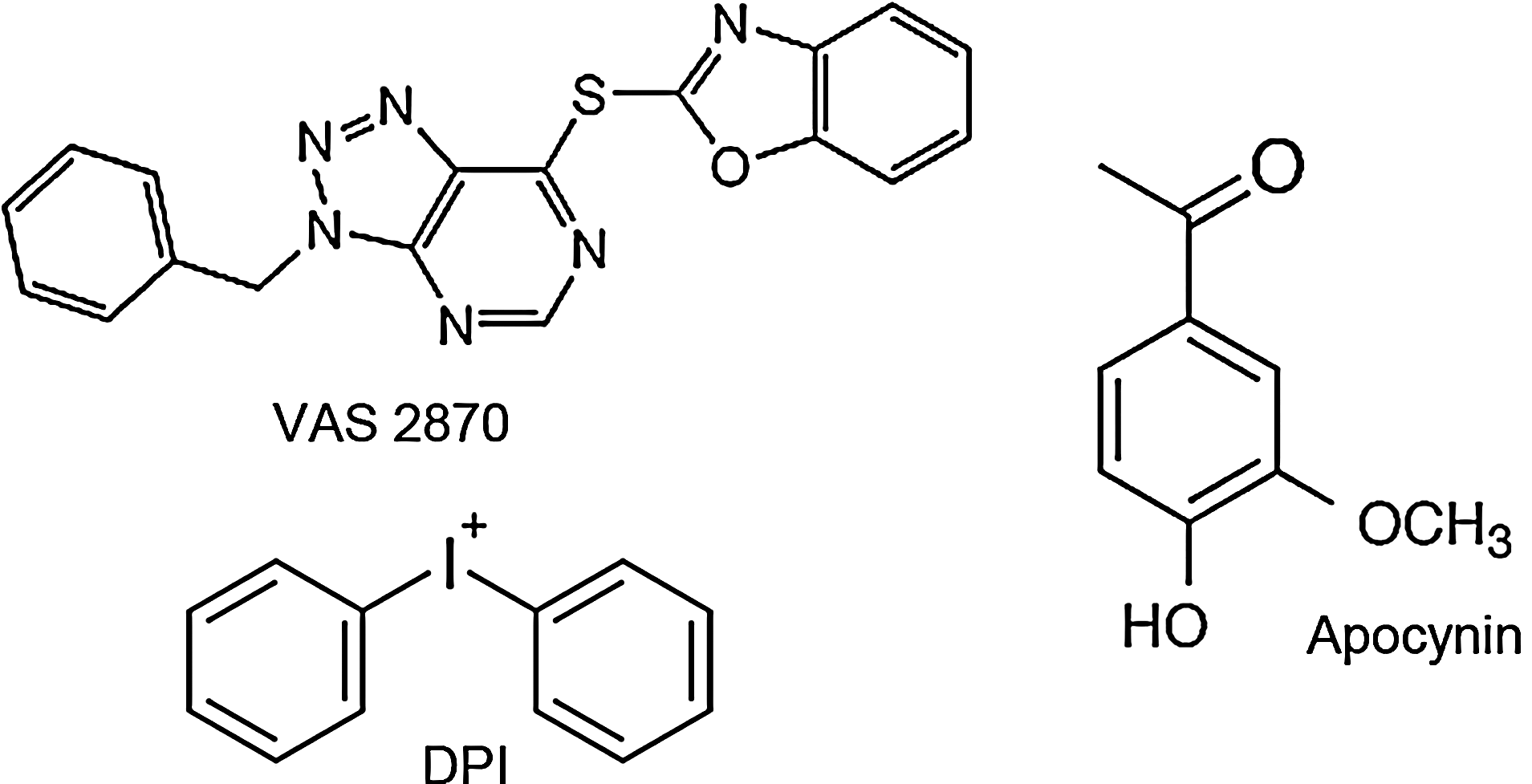
In addition to using genetic animal models, another and clinically more relevant option for validating NOX enzymes as therapeutic targets is the use of pharmacological compounds that selectively suppress NADPH oxidase activity. The most widely used substances that inhibit NOX have been apocynin and diphenylene iodonium (DPI) (Fig. 5). DPI already showed neuroprotection in vivo when administered poststroke (43). However, its usefulness, initially demonstrated as an inhibitor of phagocyte-related NOX, is limited, as it also interacts with NOS and other flavoproteins (69). Apocynin, a polyphenolic molecule with anti-inflammatory properties, is thought to block the assembly of p47phox and NOX2 and has to be activated by myeloperoxidases, which are not present in all cells. The interpretation of studies using apocynin is further complicated by its antioxidant capacity (25) and inhibition of Rho kinase (52). Thus, classical NOX inhibitors are not specific for NADPH oxidases, and, therefore, any results on the role of NADPH oxidase obtained by using apocynin or DPI should be questioned (43, 57, 69, 71). A more specific NOX inhibitor is the triazolopyrimidine VAS2870 (Fig. 5), which already proved its protective effects in preclinical stroke experiments (33). VAS2870 and its derivate VAS3947 likely inhibit all NOX isoforms and are neither general flavoprotein inhibitors nor ROS scavengers (64, 69). Mechanistically, VAS2870 seems to be a pan-NOX inhibitor (70), as it inhibits assembly or conformational changes to active NOX complexes (3). When a complex assembly of NOX2 with its cytosolic binding proteins was induced before the addition of VAS2870, no inhibition of NOX activity was observed (3, 21). When VAS2870 was added before complex assembly induction, inhibition of NOX2 activity was observed (3). Therefore, the order by which a NOX complex assembly activator and a NOX inhibitor are added in cell-free assays is crucial. Very recently, potential off-target effects of VAS2870 have been published, suggesting that the compound alkylates cysteine thiol residues (61). The authors suggest that thio-alkylation of cysteine residues may play a role in NOX inhibition. Nevertheless, the importance of this off-target alkylation in the presence of physiological glutathione concentrations and the occurrence of other off-target effects of VAS2870 need to be further analyzed in a more extended off-target screen.
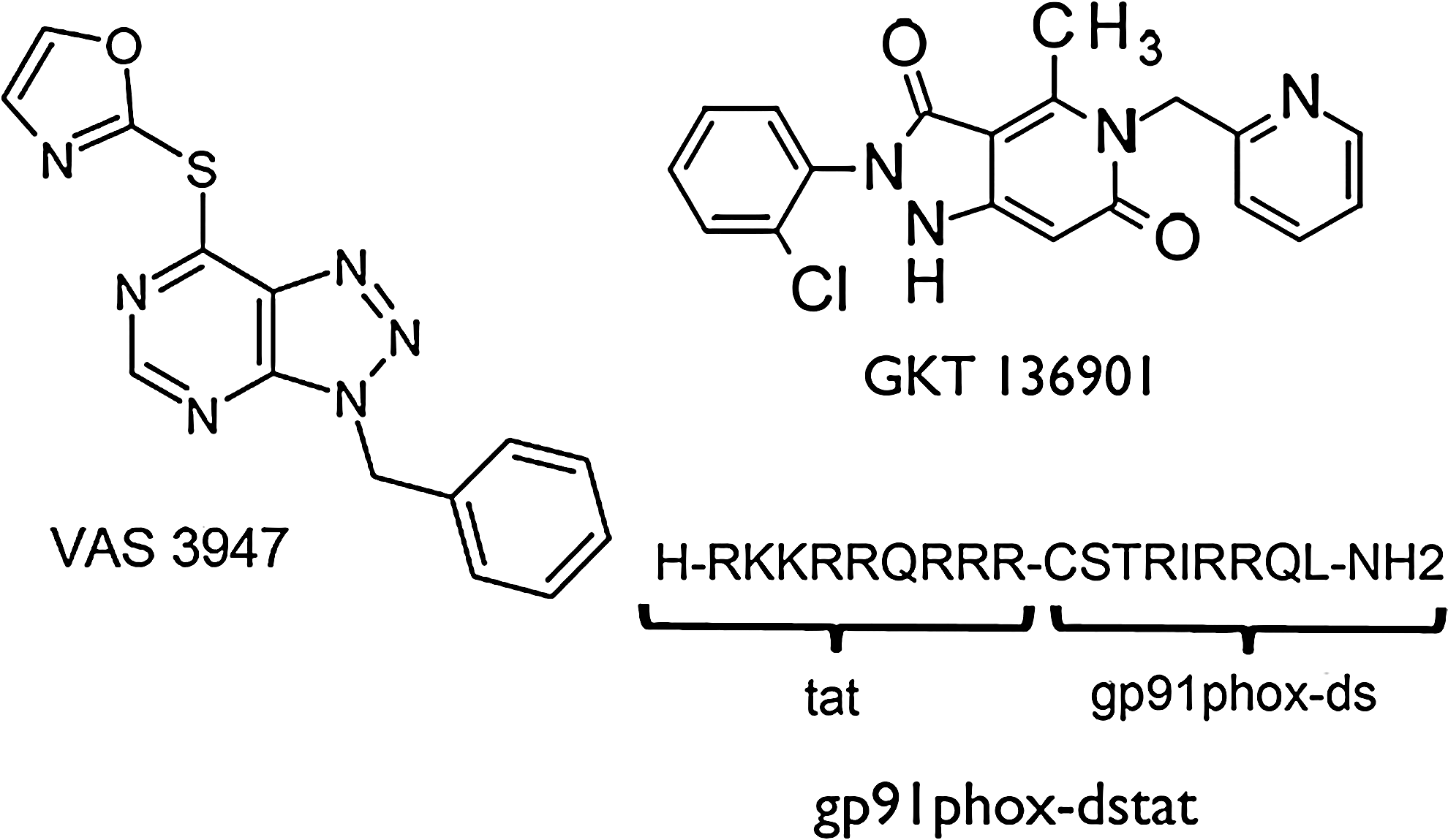
Beside VAS compounds, which are general NOX inhibitors, NOX isoform-specific inhibitors may represent a better tool for assessing the role of individual NOX isoforms and also for developing experimental therapeutics. One approach was to use peptides, such as the chimeric peptide, gp91ds-tat (50), which combines the p47phox docking sequence of NOX2 with the cell penetrating tat peptide (Fig. 6). This peptide not only prevents NOX assembly and is relatively specific for NOX2, but also potentially inhibits NOX1 (50). However, off-target effects of gp91ds-tat have been described in vivo (2). More recently, a selective NOX1/4 inhibitor, the pyrazolopyridine derivate GKT136901, was introduced (Fig. 6) (36).
To validate NADPH oxidases as a target for stroke therapy, apocynin has mostly been used as NOX inhibitor. Jackman et al. (29) showed that apocynin given prestroke caused a reduction in infarct size, but not when administered poststroke. Indeed, in most studies, apocynin was given before stroke onset (32, 62, 67), a procedure that does not allow statements on therapeutic relevance in a clinical setting, where any therapy has to work poststroke. Tang et al. did not find any additional neuroprotection in apocynin-treated mice on top of NOX2 gene deletion (63). Does this exclude a role for another NOX isoform in ischemia-induced brain damage? This is probably not the case, as apocynin interferes with p47phox binding to NOX and NOX activation. However, NOX4 does not seem to depend on p47phox (41). Neuroprotective effects of the NOX inhibitor VAS2870 have been demonstrated when injected intrathecally into the spinal liquor space of wild-type mice, but did not mediate any further protection in NOX4 KO mice, suggesting that NOX4 is the major detrimental player (33). Nevertheless, stroke experiments with other, more isoform-specific NOX inhibitors would be of great interest. Alternatively, NOX activity can also be repressed by using small interfering RNAs (3), but the interference of siRNA with other than the intended NOX isoform has to be excluded (53).
Conclusion
In conclusion, NOX4 and, possibly to a lesser extent, NOX2 appear to play a role in ischemic stroke. Both are up-regulated in human stroke, and experiments with experimental compounds suggest that NOX inhibition may be relevant for a highly innovative neuroprotective poststroke therapy. Clearly, further studies using different KO models and inhibitors in different species are needed to fully validate the NOX isoform that is the most promising target and a possible time window for further development.
Footnotes
Acknowledgments
The authors wish to acknowledge national and international competitive grant support by the NHMRC, EU and ERC (HHHW), the Brain Foundation of the Netherlands (KR), and DFG (CK). Work performed at the German Mouse Clinic was funded by grants from the German Federal Ministry of Education and Research (BMBF, NGFN-Plus grants No. 01GS0850, 01GS0851, and 01GS0854).
Author Disclosure Statement
H.H.H.W.S. is a cofounder of Vasopharm GmbH, which is developing NOX inhibitors. C.K. has received funding from Novartis Pharma GmbH, Nürnberg, Germany, to conduct research on NADPH oxidase.