
Abstract
Introduction
The focus of this review is on the redox reactions that occur in the phagosome and how they contribute to antimicrobial activity. It covers what reactive oxygen species are generated, mechanisms of production, and likely targets within the phagosome, as well as assessing the importance of different oxidative reactions in microbial killing. The main enzymatic players are the NADPH oxidase responsible for generating superoxide and hydrogen peroxide (H2O2), and myeloperoxidase (MPO), which is released from azurophilic granules and uses the products of the NADPH oxidase to generate other oxidant species, including hypochlorous acid (HOCl). Much is known about the properties of these enzymatic systems. However, understanding how they function in the phagosome has been more challenging. Phagosomal conditions are very different from those in the external environment of the neutrophil and often impossible to replicate experimentally. Therefore, we have assembled a picture of the conditions that prevail within the phagosome and used information from simulations of redox reactions that are likely to occur under these conditions, as well as findings using oxidant probes and oxidative biomarkers, to assess the fate of superoxide and how oxidative killing occurs.
While our focus is on redox reactions and oxidative killing, it is well established that neutrophils do not rely on a single mechanism to combat the range of pathogens that they encounter. As reviewed elsewhere (25, 56, 118) various granule proteins are released into the phagosome that can kill microorganisms nonoxidatively. They may also act synergistically with oxidants (69, 85). There is also another recently identified neutrophil process that may augment phagosomal antimicrobial activity by killing extracellularly. It involves the release of nuclear chromatin along with associated, predominantly granule, proteins from the neutrophils (29). The process is activated by a range of stimuli and creates a mesh-like structure, termed neutrophil extracellular traps (NETs). NETs can trap bacteria, fungi, and protozoa and it has been proposed that the presence of antimicrobial proteins enables them to kill these microorganisms (29, 191, 197). MPO acting in concert with added H2O2 appears to be particularly effective (141). However, while NETs are an exciting discovery with potential relevance to infection and inflammation, their contribution to in vivo antimicrobial activity is yet to be determined.
The widely held view is that the diversity of killing mechanisms is required because effective antimicrobial defense is so important for host survival (133). However, there are uncertainties about how effectively nonoxidative processes function in the phagosome and how oxidants kill in this environment (77, 133, 152, 169). It is clear that NADPH oxidase activity is critical for neutrophil function, as demonstrated by the susceptibility to infections of individuals with chronic granulomatous disease (CGD), a genetic disease in which the NADPH oxidase is inactive (172, 178, 202), and of mice that lack a component of the oxidase (8, 128, 194). HOCl is the strongest microbicidal oxidant produced by the neutrophil and would appear to provide the ideal antimicrobial defense. Yet MPO deficiency is relatively common but only occasionally associated with infection (113, 132, 142). We discuss this conundrum and make a case for MPO-derived reactive oxidants for frontline killing of most organisms.
Source of Reactive Oxidants—NADPH Oxidase Activity
Redox reactions in the neutrophil phagosome are all initiated by the activation of the NADPH oxidase complex. Oxygen is reduced to superoxide radicals (13) with the reducing equivalents supplied by NADPH (Reaction 1). The process is activated by the ingestion of opsonized particles, as well as a range of immune stimuli or bacterial components that interact with neutrophil receptors [reviewed in Ref. (45)].
Structure and assembly
The neutrophil NADPH oxidase is a multicomponent complex that belongs to the NOX family of proteins (21, 115, 135). Now classified as NOX2, it was the first member to be identified and for a long time thought to exist only in phagocytic cells. However, NOXs occur widely in animal and plant species and expression of at least one family member has been reported for almost every tissue examined. The role of NOXs thus extends beyond host defense to a wide range of redox-regulated cell signaling responses (31).
NOX2 is a complex comprised of two integral membrane protein subunits, gp91phox and p22phox, plus several regulatory subunits (Fig. 2) (119, 135). The catalytic center is in gp91phox (also referred to as flavocytochrome b558) and involves FAD plus two cytochrome b hemes. The flavocytochrome and p22phox are located predominantly (85%) on the membrane of specific granules, with the remainder on secretory vesicles and the plasma membrane. Three other proteins (p47phox, p67phox, and rac2) are essential for activity and reside in the cytosol of resting neutrophils. Other cytosolic proteins with as yet poorly characterized regulatory functions also associate with the complex. Superoxide generation requires assembly of the NOX2 components into an active complex. Activating stimuli cause phosphorylation on multiple sites on p47, changing its conformation and causing p47, p40, and p67 to translocate and associate with gp91phox (Fig. 2). The specific granule and plasma membranes become the phagosomal membrane due to invagination of the cell surface and granule fusion, so the majority of NOX activity following particle ingestion occurs at the phagosomal membrane (46). As shown using cytochemical methods, superoxide and H2O2 are generated (27, 28, 158) and the system is set up to direct oxidants at the ingested microbe.

When considering the role of NADPH oxidase activity in neutrophil function, it is important to note that some intracellular activity occurs at nonphagosomal sites (33, 158, 192). These include the membranes of specific granules, perhaps before they fuse with the phagosome. However, intracellular superoxide generation is also seen with stimuli that do not induce phagosomal formation (121), and as reviewed by Bylund et al. (33), likely sites include vesicles or endosomes. The nonphagosomal intracellular oxidase activity is apparently not involved in antimicrobial activity and it is possible that, as in other cell types, it has a signaling role that contributes to the inflammatory response (24, 33).
Enzymology, directionality, and electrogenic effect of NADPH oxidase
A key feature of the NOXs is that the reaction they catalyze is directional across the membrane (111, 119). The complex is oriented such that NADPH is consumed in the cytosol and the electrons are transferred to FAD, then sequentially via the two hemes to oxygen (Fig. 2). As the external surface becomes internalized during particle ingestion, superoxide is released into the phagosome.
The Km for oxygen is only ∼10 μM, which enables the oxidase to function at the low oxygen tensions present in tissue (67). At maximum capacity the isolated enzyme can transfer ∼160 electrons/(heme·s−1) (111). It can be calculated that the fully activated oxidase would consume the basal concentration of NADPH in the neutrophil (∼50 μM) in less than a second (45). Thus, NADPH must be regenerated continuously from NADP+ to maintain the oxidative burst. This occurs via the hexose monophosphate shunt pathway (26, 214), with glycogen breakdown providing most of the glucose required (181). Glucose-6-phosphate dehydrogenase (G6PD) is the first enzyme in the hexose monophosphate shunt. G6PD deficiency affects millions of the world's population and occurs with differing severity depending on the specific mutation in the protein (35). Although neutrophil function is in most cases unaffected, the most severe cases (with <1% normal G6PD activity) have CGD-like symptoms due to the inability of their neutrophils to mount an effective oxidative burst (15, 72).
Because it is directional, the oxidase is electrogenic and transfers electrons out of the cytoplasm into the phagosome (or surroundings if it is on the external surface). This creates a charge imbalance that needs to be counteracted to prevent depolarization of the membrane and shutdown of the oxidase. Charge compensation is provided primarily by hydrogen ions transported by voltage-gated proton channels (44, 47). The VSOP/HV1 channel appears to be primarily responsible (55, 148, 155). With the balancing flow of protons, the negative charge in the phagosome is dissipated by superoxide dismutation (Reaction 2) and cytoplasmic acidification due to NADPH oxidation is overcome.
The neutrophil NADPH oxidase is viewed predominantly as a source of toxic oxidants. However, an alternative function has been proposed (156, 157, 169) in which some of the negative charge is counterbalanced by transport of potassium ions into the phagosome thus increasing the ionic strength and aiding solubilization of antimicrobial granule proteins. While the proposed potassium transport mechanism has been discounted (59, 129, 133), other consequences of the electrogenic effect are theoretically possible (152). For example, there could be electron receptors other than oxygen, or it could act as a driving force for secondary transport into negatively charged vesicles (114). Rapid consumption of NADPH should also be considered as a mechanism whereby NOX activity could regulate cellular NADPH levels. This has the potential to influence the wide variety of cellular redox processes that depend on NADPH. It may be particularly relevant for the nonphagosomal activity in stimulated neutrophils, and for NOX activity in other cells.
Regulation and termination of NADPH oxidase activity
Superoxide generation by the neutrophil is a finite, regulated process that can be primed, activated, and terminated (45). Priming, which is instigated by a number of proinflammatory cytokines and lipopolysaccharide, does not in itself induce superoxide production, but prepares the cells to mount an augmented oxidative burst when subsequently exposed to a full stimulus [reviewed by Sheppard et al. (174)]. At a molecular level, priming alters the structural organization of the NADPH oxidase by causing partial phosphorylation and translocation of subunits to the specific granule or plasma membranes, as well as cytoskeletal changes.
The duration of the respiratory burst varies depending on stimulus [as summarized by DeCoursey and Ligeti (45)]. For phagocytosis, time courses ranging from a minute measured as oxygen uptake with opsonized latex (170) to 90 min with Nesseria meningitides (46) have been reported. Obtaining accurate estimates is difficult because measurements are confounded by the rate at which ingestion occurs, by variability between cells, and by cells taking up multiple particles. With yeast particles under conditions designed to synchronize phagocytosis, a peak of superoxide production at 10–15 min (a period that would include ingestion as well as assembly of the oxidase) and then a gradual decline over a similar period was observed (71). Studies using a fluorescent probe for HOCl suggest that oxidant production in most phagosomes lasts a few minutes but some show more extended activity (189). Although there is variability, it would appear that most of the oxidase activity typically occurs over several minutes following particle ingestion.
How the neutrophil terminates its oxidase activity is not well understood. Observations that MPO-deficient neutrophils exhibit an extended burst following particle ingestion (106) point to oxidative inactivation as a possible contributor. However, further study indicated that this phenomenon reflects greater general deterioration of normal compared with MPO-deficient cells (49) and oxidation is probably not a major mechanism. Evidence to date implicates changes in kinase/phosphatase activity, and decreased phosphorylation of p47 and the species bound to Rac, as dominant factors (45).
Neutrophil Oxidants—Nature and Production
Once formed in the phagosome, superoxide has the potential to spawn an array of reactive oxygen species. However, when the chemistry of their formation is considered, the probable yields of particular oxidants vary enormously. For example, there is good evidence that superoxide dismutates to H2O2, which then reacts with MPO to generate HOCl and chloramines. Support for phagosomal production of other oxidants, including hydroxyl radical, singlet oxygen, ozone, and hypohalous acids besides HOCl, is less convincing. It should be born in mind that oxidants observed in the extracellular environment of neutrophils may not necessarily be produced under phagosomal conditions. In this section we outline the relevant chemistry for the oxidants that are likely to be formed in the phagosome and act as antimicrobial agents. More extensive coverage of the chemistry of reactive oxygen species is described elsewhere (74, 204, 205).
Superoxide and hydroperoxyl radical
Superoxide's name evokes extreme reactivity. But in aqueous solution, it is neither a strong oxidant nor a powerful nucleophile. Its major chemistry is that of a moderate one-electron reductant; a ligand for metal complexes, such as ferric MPO; and a strong Bronsted base that removes protons from weakly acidic substrates (110, 167). Its fastest reactions are with other radicals, such as nitric oxide (NO), nitrogen dioxide, and organic radicals. Superoxide also oxidizes iron–sulfur clusters of dehydratases, such as aconitase (87). This inactivates the enzymes and releases iron, which can react with H2O2 to form hydroxyl radicals. This sequence of reactions is proposed as a major contributor to the toxicity of superoxide (86).
Superoxide is in equilibrium with the hydroperoxyl radical (HO2 •). With a pKa of 4.8, there will be only ∼0.1% hydroperoxyl radical at neutral pH. However, the hydroperoxyl radical may still be relevant to bacterial killing at the high micromolar concentrations of superoxide likely to be present in phagosomes. It is much more oxidizing than superoxide and its higher reactivity with some biomolecules should be enough to overcome their difference in concentration. Also, its neutral charge will enable it to pass through membranes that exclude superoxide (43).
Hydrogen peroxide
H2O2 is formed from the dismutation of superoxide. In the phagosome, dismutation occurs either spontaneously or via reactions catalyzed by MPO (207). Although H2O2 can permeate bacteria, it is unlikely to be directly bactericidal at the concentrations achieved in the phagosome (88). Its relatively benign nature is explicable in terms of its chemistry. Although it has a high two-electron reduction potential (H2O2/H2O; 1.77 V) and is therefore a strong oxidant, a high activation energy makes it a kinetically sluggish oxidant of most biomolecules. However, rates of reaction with iron–sulfur clusters are sufficiently fast (k=104 M −1·s−1) for H2O2 to damage dehydratases and kill bacteria by this mechanism (86). H2O2 reacts rapidly with heme proteins (204). Thus, MPO is likely to be its main target within phagosomes. If H2O2 does diffuse into bacteria containing catalases and peroxiredoxins, they should be protective by degrading it to harmless products (149).
Reactions catalyzed by MPO
MPO is discharged into the phagosome as bacteria are phagocytosed and accounts for ∼25% of the neutrophil proteins in this vacuole (107). It is the only enzyme in the phagosome known to react with superoxide and H2O2 at meaningful rates (97, 103) and it therefore dictates phagosomal redox chemistry. MPO is a 150 kDa highly cationic protein (pI ∼ 10). It is a dimer of two identical dimers, each containing heavy and light polypeptide chains. A heme prosthetic group connects each heavy and light chain via two ester bonds with Asp94 and Glu242 as well as a unique sulfonium linkage with Met243 (66). The latter linkage imparts high reduction potentials to the redox intermediates of MPO and is responsible for the enzyme's green color. The two hemes are located on the same face of the protein in crevices about 20 Å in depth (61). Access is via a narrow channel on the distal side that limits the entry of bulky substrates (Fig. 3). All studies to date demonstrate that the two hemes have the same reactivity and do not interact (66).

MPO is an intriguing enzyme because it has several activities and multiple substrates (103). Its complex chemistry makes disentangling its role in bacterial killing both challenging and fascinating. While commonly viewed as a pro-oxidant enzyme that catalyzes the production of hypohalous acids and free radicals, it also consumes superoxide and H2O2 via three different enzymatic cycles that correspond to antioxidant activities. The activities and substrates relevant to the redox chemistry within phagosomes are described in the following sections.
Oxidative activities of MPO
Chlorination activity
The ferric enzyme harnesses the high oxidation potential of H2O2 to form an array of powerful and toxic oxidants. Reaction with H2O2 (k=2×107 M −1·s−1) converts the enzyme to an Fe(IV) cation radical called Compound I (65, 124) that in turn oxidizes a multitude of substrates by removing either one or two electrons. The two-electron reduction potential of Compound I is 1.16 V (10). Substrates that donate two electrons to Compound I include chloride, bromide, and the pseudohalide thiocyanate (65) as well as H2O2. The (pseudo)halides are oxidized to the corresponding hypohalous acids and the enzyme is reduced back to its native ferric state (shown for chloride oxidation to HOCl in Fig. 4A). Within phagosomes, chloride will be the predominant halide that is oxidized because the concentrations of the others are too low for them to be appreciable substrates over the course of the respiratory burst (37, 193).

One-electron substrates, such as H2O2, nitrite, and tryptophan, that react well with Compound I but reduce Compound II slowly cause accumulation of Compound II and inhibit HOCl formation (103). Superoxide can reverse this and should therefore be viewed as a cosubstrate in the chlorination activity of MPO (Fig. 4A). Mycobacterium tuberculosis, Nocardia asteroids, and Actinobacillus pleuropneumoniae discharge SOD (20, 112, 117) and may use the enzyme to protect themselves from oxidative killing by preventing superoxide from recycling Compound II.
It has been proposed that chloride reacts with Compound I to form a chlorinating intermediate that chlorinates taurine at the enzyme's active site (123). This contrasts with earlier observations that the oxidant generated by MPO has similar reactivity to reagent HOCl (203). More recent studies suggest that MPO may either release free HOCl, or if the substrate is accessible to the active site, chlorinate it directly (154). Further investigation is required to establish the functional relevance of the latter mechanism.
Peroxidase activity
MPO functions as a classical peroxidase where Compound I oxidizes substrates by removing a single electron to produce a substrate free radical and Compound II. This redox form of the enzyme exists as an Fe (IV) intermediate. Compound II reacts with a second substrate molecule to produce another radical and regenerate the ferric enzyme (Fig. 4B). Compound I has a one-electron reduction potential of 1.35 V (64), which makes it one of the strongest physiological one-electron oxidants known and capable of oxidizing a wide variety of reductants. Compound II has a more restricted substrate preference due to its lower reduction potential (0.97 V) and possibly a narrower substrate channel (2). As a consequence its reduction limits the rate at which the enzyme turns over.
Physiological substrates for the peroxidase activity include superoxide, tyrosine, ascorbate, urate (126), serotonin (211), and nitrite [reviewed in Refs. (42, 103)]. Neutrophil stimulation in the presence of these substrates initiates radical-mediated processes, including lipid peroxidation, tyrosine nitration, and protein crosslinking (12, 81, 126, 210). The radicals produced by tyrosine, urate, and serotonin react at almost diffusion-controlled rates with superoxide to form organic hydroperoxides (shown for tyrosine in Fig. 5) (130, 209). These hydroperoxides could be of particular relevance to the redox chemistry in the phagosome because they are more reactive than H2O2 and are potentially bactericidal (126, 211). Thus, MPO has the capacity to catalyze the generation of a range of radical-derived species that may contribute to oxidative killing.

Antioxidant activities of MPO
Superoxidase activity
Superoxide reacts rapidly with both Compound I and Compound II (k=5.6×106 M −1·s−1 and 1.1×106 M −1·s−1, respectively, at pH 7.4) (97, 99). Consequently, MPO can be considered as a superoxidase that uses H2O2 to oxidize superoxide to dioxygen (Fig. 4C). As chloride competes with superoxide for Compound I (104), this reaction would be relevant inside phagosomes only when the concentration of chloride is low.
Superoxide dismutase activity
Ferric MPO reacts with superoxide [k=2×106 M −1·s−1; (100)] to form Compound III [an Fe (VI) intermediate]. This reaction is fast enough to compete with H2O2 reacting with ferric MPO. Compound III is the major form of MPO when the enzyme is exposed to a superoxide-generating system in the presence of chloride (206). It is not a dead-end complex because under these conditions the enzyme still produces HOCl. Compound III is reduced by ascorbate and serotonin (84, 211), and is involved in the hydroxylation of salicylate (102) and oxidation of melatonin (212). Inside the phagosome, however, superoxide is its most likely reductant. This reaction completes a catalytic cycle whereby superoxide is dismutated to H2O2 and oxygen (Fig. 4D). Experimental evidence for the cycle comes from the finding that a superoxide-generating system is able to convert MPO to a maximum of 90% Compound III (97). This indicates that superoxide must reduce Compound III back to the ferric enzyme and provides an estimated rate constant of ∼2×105 M −1·s−1. The superoxide dismutase activity of MPO is considerably less than that of SOD where the analogous rate constants are four orders of magnitude greater.
Catalase activity
Compound I oxidizes H2O2 by removing two electrons to produce oxygen. This reaction is thermodynamically facile [E0’(O2/H2O2)=0.28 V] and fast (k=2×106 M −1·s−1) (104). Although not without debate (51), MPO has the potential to act as a true catalase (Fig. 4E). It has been proposed (169) that the prime function of MPO is to act as a catalase, thereby protecting other antimicrobial granule proteins from oxidative damage. However, above 40 mM chloride, the chlorination activity dominates (104) and little catalase activity would be expected under phagosomal conditions.
Oxidants produced by MPO
Hypohalous acids
Stimulated neutrophils generate hypohalous acids (37). Around 25%–30% of the oxygen consumed by the cells can be detected by extracellular HOCl traps (101, 198). HOCl [E0’(HOCl/Cl−)=1.28 V] is a strong two-electron oxidant (42). It also undergoes chlorination reactions in which chloride is incorporated into an organic molecule. HOCl has a pKa of 7.4 so it will exist as an equal mixture with hypochlorite (−OCl). HOCl chiefly oxidizes methionine and cysteine residues (k ∼ 3×107 M −1·s−1) (144). Methionine is oxidized to methionine sulfoxide and also dehydromethionine if it is N-terminal (19, 146). Cysteine residues are oxidized initially to sulfenyl chlorides which then form disulfides, sulfenic acids, and higher oxidation states (42, 204), and may crosslink with lysine residues to form sulfinamides and sulfonamides (63). The next preferred targets are amine groups (k=103–105 M −1·s−1) (144), disulfides, and tryptophan residues. Tyrosine reacts slowly with HOCl to produce 3-chlorotyrosine (k=44 M −1·s−1). A more likely route to this chlorine footprint in proteins is via transchlorination from a juxtaposed chloramine (23, 48).
HOCl is highly toxic to bacteria and other microorganisms (107). Therefore, it appears ideally suited to act as a broad spectrum antibiotic against all pathogens that are ingested by neutrophils. An MPO system generating HOCl is much more lethal than other cytotoxic neutrophil proteins (107). Bactericidal mechanisms are discussed in greater detail by Hurst (85) and include disruption of ATP production due to destruction of cellular electron transport chains and the adenine nucleotide pool (3, 18). Oxidation of methionine residues in cytosolic and inner membrane proteins of bacteria has also been implicated (162). HOCl can also cause delayed toxicity when treated bacteria undergo aerobic growth (50).
Chloramines
Chloramines are formed when HOCl reacts with primary and secondary amines (Reaction 3) (186). The major targets in the neutrophil will be amine groups on proteins. Chloramines break down slowly to reactive aldehydes and ammonia (Reaction 4) (78, 79), which reacts with further HOCl to form monochloramine [ammonium chloride (NH2Cl)] (Reaction 5).
Chloramines react slowly with HOCl to produce dichloramines (186). When they contain a substituent on their α-carbon (as in N-terminal amines), these species break down rapidly to liberate either NH2Cl or ammonia dichloramine (Reaction 6) (41). Chloramine formation on amino acids, proteins, and ammonia has been detected in the surroundings of stimulated neutrophils (182, 184). Further, neutrophils produce dichloramines because it was shown that they were responsible for the MPO-catalyzed incorporation of amines into granule proteins (187).
Chloramines are generally much less-reactive oxidants than HOCl (144). They react predominantly with sulfur-containing amino acids (147, 185). Products are similar to those with HOCl except that chloramines cause little formation of higher oxidation products and they preferentially target thiolate anions. This makes them more selective than HOCl at inactivating low-pKa thiol enzymes. Chloramines are poor chlorinating agents but as noted above can cause intramolecular tyrosine chlorination.
Chloramines kill a wide range of microorganisms. Their toxicity depends very much on their charge and ability to diffuse into bacteria (185). For example, neutral NH2Cl and NHCl2 are at least as toxic as HOCl but negatively charged taurine chloramine is a weak antimicrobial agent (41). Charged chloramines can still be toxic by undergoing exchange reactions with ammonia to form ammonia chloramine (70, 145).
Reactive nitrogen species
Neutrophils have the potential to produce a range of reactive nitrogen species (131). However, human neutrophils are much less capable at producing NO than their murine counterparts and an inflammatory environment is critical for inducing iNOS expression (136). Early studies gave contrasting and controversial results as to whether human neutrophils produce NO (131). Later work, however, revealed the presence of inducible NO synthase activity in neutrophils isolated from the urine of patients with urinary tract infections and in peripheral blood neutrophils after exposure to inflammatory cytokines (57, 201).
Generation of NO by neutrophils opens up a rich panoply of chemistry that could be used to kill bacteria (58, 60). The reaction of NO with superoxide (k=1.6×1010 M −1·s−1) is extremely rapid and would be expected to occur if NO enters the phagosome. Peroxynitrite, the product of this reaction, is bactericidal in its own right but is more likely to react with carbon dioxide to produce nitrogen dioxide (NO2 ●) and the carbonate radical (CO3 −●). Peroxynitrite, as well as MPO/nitrite, can give rise to nitrotyrosine, a signature for reactive nitrogen species production (89). Both mechanisms involve nitrogen dioxide reacting with tyrosyl radicals (116), but the reaction responsible for the production of nitrotyrosine by neutrophils is still a matter for debate (54).
NO modulates the activity of MPO by reversibly binding to its ferric form (1). It is also consumed by MPO when it reacts directly either with Compound I or with radicals derived from other peroxidase substrates (52). Peroxynitrite oxidizes ferric MPO (k=6×106 M −1·s−1) (62) to form nitrogen dioxide and Compound II in a concerted reaction via nitrite and Compound I. Although HOCl reacts with nitrite to give nitryl chloride (53), this should be limited by more favored reactions of HOCl with other targets.
Hydroxyl radical and singlet oxygen
Hydroxyl radicals are the most reactive of all biologically derived chemicals and oxidize myriad biomolecules. As discussed elsewhere (85), they kill bacteria but paradoxically their high reactivity limits their efficiency. They are more likely to react with other targets within the phagosome before encountering the bacterium. Hydroxyl radical formation by neutrophils has been widely studied, but after critical examination of probe specificity, the consensus is that this is not an efficient process and accounts for very little of the oxygen consumed by the cells (77, 161). Superoxide-driven Fenton chemistry was initially invoked as the source of hydroxyl radicals, but a more likely route is the reaction of superoxide with HOCl (k=7.6×106 M −1·s−1) (34, 153).
Although singlet oxygen was initially proposed to be the source of the chemiluminescence of stimulated neutrophils (6), this was discounted by the use of more specific probes. Although there is one report of high yields (180), the prevailing view is that singlet oxygen generation by neutrophils is very low (85, 94). Possible routes to singlet oxygen include the reaction between HOCl and H2O2 (105) and the decay of lipid and other organic hydroperoxides generated by MPO (150). It is thermodynamically possible that oxidation of superoxide by MPO could produce singlet oxygen as in Figure 4C, but this has not been investigated (110).
At the turn of the millennium several high-profile articles proposed that antibodies could catalyze the production of ozone from singlet oxygen and water (199) and that neutrophils generated ozone by this mechanism (14, 200). However, subsequent studies have shown that the signature reactions used to implicate ozone (conversion of indigo carmine to isatin sulfinic acid and formation of particular oxysterols) could also be carried out respectively by superoxide (98) and alternative well-established oxidative pathways (177, 190). More compelling evidence for production of ozone by inflammatory cells has not been forthcoming.
Likely phagosomal oxidants
Given all the above permutations, MPO-derived products stand out as the major species likely to be formed from the superoxide and H2O2 produced by neutrophils. Reactive nitrogen species also deserve attention, and exploration of the interplay between the NADPH oxidase, NO synthase, and MPO is bound to be fruitful in understanding how reactive species conspire to kill pathogens. On the other hand, roles for hydroxyl radicals and singlet oxygen appear to be limited. However, the enzymatic and chemical reactions described previously have been observed mainly in cell-free systems or in the environment of neutrophils and will not necessarily occur under the conditions that exist inside the phagosome. We therefore consider the phagosomal environment and the fate of superoxide in this situation.
The Phagosomal Environment
Phagosome formation and degranulation
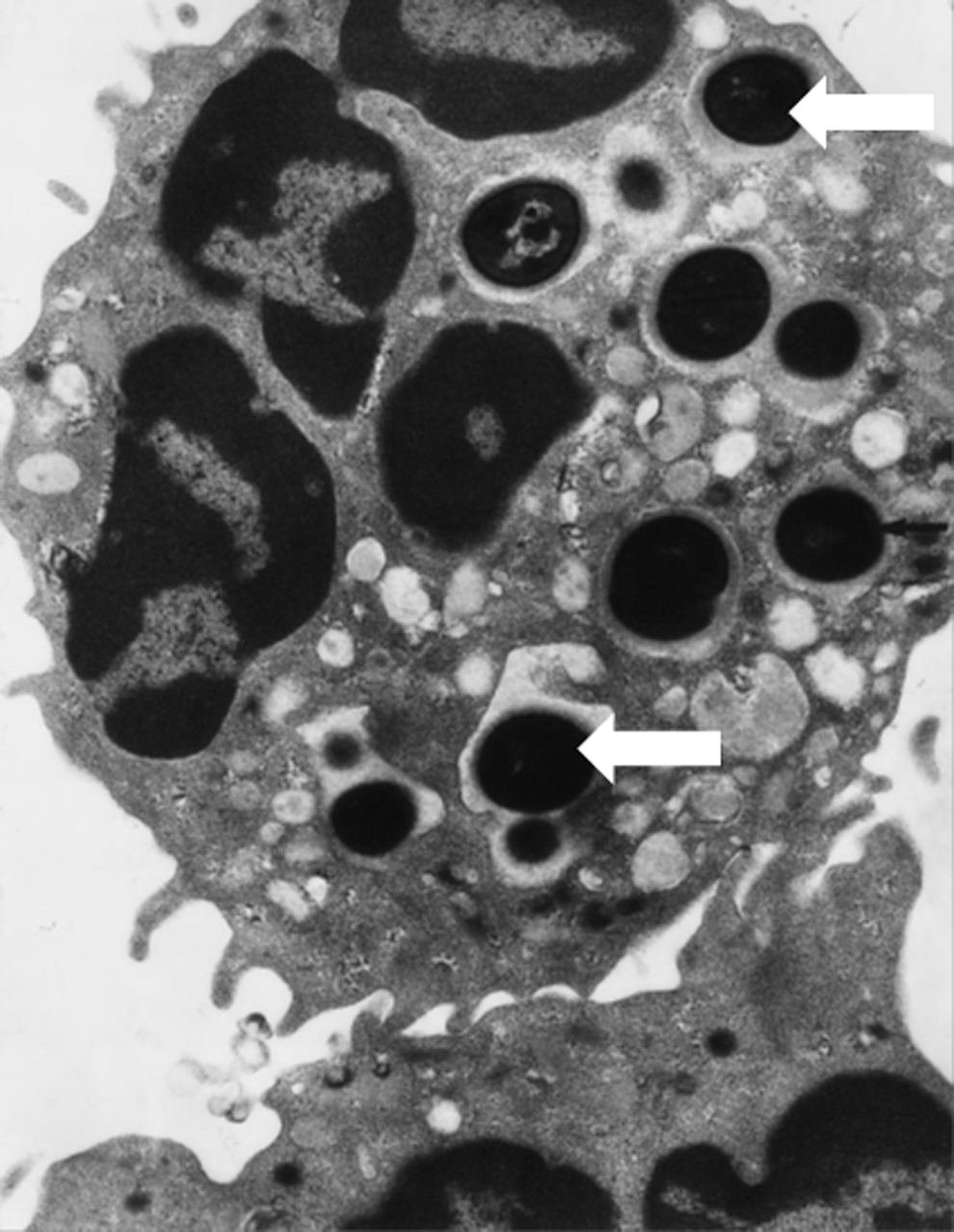
When the neutrophil ingests a bacterium, it spreads over its surface, creating a very narrow space where oxidants and granule proteins are released and killing occurs (16, 159, 164) (Fig. 1). Little extracellular medium is taken up (22), degranulation begins within seconds of ingestion, and the NADPH oxidase becomes activated on the fused plasma and specific granule membranes (83, 164). Based on electron microscopic and biochemical data [analyzed in Ref. (207)], it can be deduced that the phagosomal volume is very small, and that granule contents make up almost half the total volume. As granules are packed with protein, the concentrations of MPO and other proteins are very high (Table 1). Sequential fusion of individual granules will expand the size of the phagosome over time, as well as replenish protein constituents that may have been expended. Swelling will also occur due to osmotic influx of water as the bacterium is digested.
Period when most of killing occurs.
Equivalent to 1.5 μm diameter surrounding 1 μm bacterium. Volume will increase over time.
Maximum assumes 80% degranulation with a maximum number of 20 particles ingested (92). Total MPO ∼2.5 μg/106 neutrophils (17).
During initial stages; drops to ∼pH 6 over an hour (91).
Measured value of 3–4 nmol/min per 106 cells for neutrophils ingesting maximum number (15 –20) of particles (92, 170).
Information was taken from Winterbourn et al. (207) and was used in kinetic simulations.
Superoxide
Oxygen is converted to superoxide within the phagosomal space. Phagocytosing neutrophils at maximum capacity (15–20 particles ingested per cell) consumes 3–4 nmol/min oxygen per 106 cells (92, 170). From these measurements, which take into account any limitations due to oxygen diffusion and the approximate volume of the phagosomes associated with this number of particles (Table 1), a consumption rate in each phagosome of ∼2.5 mM/s can be calculated. This corresponds to an extremely high superoxide flux.
Phagosomal pH
Most of the bacterial killing takes place in the early stages after ingestion when the pH is neutral to slightly alkaline. This was first demonstrated by Segal and coworkers (171) who reported an initial rise from 7.4 to 7.8. Others have confirmed this finding (36, 91) although the initial rise was not observed in the latter study. Only over an hour, as the phagosome matures and NADPH oxidase activity declines, does the pH drop to 6–6.5.
MPO substrates
The redox chemistry that takes place will depend on what substrates are available for the enzymatic and chemical reactions described previously. Oxygen is not generally limiting for NADPH oxidase activity, and influx of protons maintains the respiratory burst and allows superoxide to dismutate. Thus, H2O2 will be available to react with MPO. As already described, MPO catalyzes numerous peroxidase reactions. Other than chloride, little is known about the availability of other potential MPO substrates. In a study using a chloride-sensitive probe attached to beads, a phagosomal chloride concentration of ∼70 mM was estimated for neutrophils in physiological medium (139). This could have arisen from the medium, cytoplasm, or granules. Little is known about chloride in granules although indirect evidence (32) implies that the concentration is not high. Neutrophil cytoplasm contains 80 mM chloride (176), and a variety of chloride channels could provide access to the phagosome. Soluble agonists, as well as particle ingestion, can cause rapid chloride efflux into the surroundings through specific channels (32, 73, 125, 175). These studies did not consider influx into phagosomes, but measurements by Painter and coworkers (137 –139) indicate that the cystic fibrosis transmembrane conductance regulator (CFTR) transporter enables chloride entry from the cytoplasm. This observation was made with MPO inhibited so the impact of HOCl formation was not assessed.
Confining reactions to the phagosome
Neutrophils face the conundrum of directing reactive oxidants toward microbes while minimizing damage to themselves or host tissue. Phagocytosis partly achieves this but to be effective, toxins should remain in the phagosome. A surrounding membrane is not necessarily sufficient as some species are membrane permeable. Superoxide should be largely retained as it has low permeability and, even though it can pass through anion channels, this transport is inefficient (122). However, uncharged H2O2 is able to diffuse through membranes (either neutrophil or bacterial) and will only be trapped if there are highly reactive constituent(s) in the phagosome. Although HOCl can pass through membranes in its protonated form, a membrane barrier partially restricts its passage (195) and less would be expected to escape.
From the broad picture of what conditions are likely in the phagosome, it is clear that both the MPO concentration and flux of superoxide are higher than can be studied experimentally, chloride availability could be an issue for HOCl production, and protein targets for oxidants such as HOCl are highly prevalent. In the following section we consider how the oxidant produced by the NADPH oxidase and MPO should react under these conditions.
Modeling Redox Reactions in the Phagosome
Using available information on the phagosomal environment plus the enzymology of MPO, it is possible to model the redox reactions that are likely to occur when superoxide is generated. The general principle is to construct rate equations using rate constants and reactant concentrations for all the reactions under consideration, allow superoxide to be generated in the system, and simulate outcomes using a kinetics program such as the Simulink function of Matlab (207). As well as known quantitative data, estimated values for concentrations can be included and varied to assess how critical each parameter is to the outcome. These analyses have been described in detail (207) and the main conclusions including any updates since the earlier publication are summarized here.
Fate of superoxide and interaction with MPO
Superoxide is generated at the high rate of ∼5 mM/s. Almost all dismutates to H2O2. A steady state is rapidly reached as consumption equals production and superoxide stabilizes at the high concentration of ∼20 μM (Fig. 6). Interestingly, superoxide is largely consumed by MPO. Lowering the MPO concentration below the 1 mM value used in the simulation (Table 1) increases the superoxide concentration and if MPO is absent, over 100 μM is reached. MPO stabilizes as predominantly as Compound III under phagosomal conditions [as observed experimentally (206)] and the MPO reactions depicted in Figure 4D are responsible for dismutating the superoxide.

Generation and consumption of H2O2
Even though H2O2 is generated from superoxide dismutation at a high rate, sufficient MPO is present to prevent it accumulating. Contrary to the common assumption that a high concentration is reached, H2O2 stabilizes in the low micromolar range (Fig. 6). With less MPO, the concentration does increase, but only modestly due to diffusion out of the phagosome. For example, with 0.1 mM MPO, ∼30% escapes and even with no MPO, H2O2 reaches only about 30 μM. Some will penetrate the bacterium but more will be lost into the neutrophil cytoplasm. Therefore, an advantage of the MPO concentration being high is that it restricts the H2O2 to within the phagosome.
Peroxidase activity and HOCl production
The fate of the H2O2 in the phagosome depends on what MPO substrates are present. As a starting point in our simulations, we considered only chloride and kept it constant at the measured concentration of 70 mM (139). Under these conditions almost 90% of the oxygen consumed is converted to HOCl (Fig. 6). Less HOCl is formed at lower MPO concentrations due to more H2O2 escaping from the phagosome (50% efficiency with 10-fold less MPO). Lowering the chloride concentration also decreases HOCl formation but, even at 20 mM chloride, 75% efficiency is maintained. The decrease is mainly due to superoxide competing with chloride for Compound I (see Fig. 4A, C). Thus, if the phagosomal chloride decreases, MPO functions more as a superoxidase. Even though MPO can exert catalase activity (Fig. 4E), under a broad range of phagosomal conditions this makes little impact on H2O2 consumption.
A requirement for chloride becomes apparent when chloride consumption is included in the simulations. Chloride is then depleted in less than a minute and production of HOCl ceases. Ongoing production of HOCl therefore requires replenishment of chloride. It will be regenerated when HOCl oxidizes its most favored substrates, such as thiols or methionine. However, once oxidizable substrates are consumed, formation of chloramines or other chlorinated molecules will result in net chloride loss. Chloride will also be lost through diffusion of HOCl or chloramines into the bacterium or neutrophil cytoplasm. Chloride influx is then required to maintain efficient HOCl production in the phagosome. However, there is still uncertainty about whether it involves CFTR or other chloride channels and whether chloride transport limits HOCl production.
Although there are many other halogenation and peroxidase substrates for MPO, these are likely to have a minor impact on the redox chemistry of the phagosome. With the high rate of H2O2 formation, they would be rapidly oxidized, but at physiological concentrations well below millimolar, these substrates would be consumed within seconds. Without an efficient mechanism for replenishment, they would account for very little of the H2O2 consumed by the MPO. The products would have to have potent effects to contribute to killing or other oxidative reactions in the phagosome. Even though substrates such as tryptophan or nitrite can inhibit MPO by trapping it as Compound II, turnover by superoxide overcomes this.
To summarize, modeling studies indicate that MPO is responsible for catalyzing dismutation of phagosomal superoxide regardless of chloride concentration. Provided the cell can provide adequate chloride, it is an efficient generator of HOCl. If chloride is limited, it functions more as a catalyst for removal of superoxide and H2O2 (Fig. 7).

Phagosomal targets for HOCl
The presumption is that when HOCl is generated in the phagosome, it is directed at the ingested pathogen. However, with the high concentration of granule proteins present, these are also likely to be targets. Modeling shows that methionine and cysteine residues in phagosomal proteins will be rapidly oxidized, followed by amines, disulfides, and tryptophan (143). Tyrosine is a much less-favored target. Of other possible reactions, singlet oxygen formation from HOCl plus H2O2 is slow (82) and should not be significant at the low H2O2 concentration in the phagosome. Hydroxyl radical formation from the reaction of HOCl with superoxide is more favorable but would only occur when amine groups were depleted. An amino group concentration of 1% of that in Table 1 is sufficient to suppress this reaction and could easily be achieved through ongoing release of granules or bacterial constituents.
With the high concentration of protein targets in the phagosome, an important consideration is the extent to which they restrict HOCl from reaching the ingested bacterium. The distance that HOCl can travel before it is consumed depends on how concentrated the targets are and how fast they react, and can be estimated by modeling (207). If the phagosome is assumed to be homogeneous and to contain 200 mg/ml protein, this distance would be initially be ∼0.03 μm, increasing to 0.1 μm as the most reactive protein targets become oxidized. Only after most of the amine groups are modified should HOCl diffuse across the width of the phagosome. Therefore, it should react close to its site of generation, and at least initially, only a fraction would travel far enough to reach the encapsulated bacterium.
An important caveat is that the phagosome is not homogeneous. Granule fusion occurs at discrete sites and superoxide is generated on the phagosomal membrane. Further, MPO binds to the surface of some (but not all) microbes (5, 127, 173), and this could provide a mechanism for bypassing, scavenging, and directing the microbicidal action of HOCl (4). However, for organisms that show high affinity binding, the number of binding sites measured per bacterium was in the range 4000–17,000 (5, 127) whereas a phagosome containing 1 mM MPO will contain over 100,000 molecules. Therefore, only a few percent of HOCl generated should come from bound MPO.
This analysis leads to the conclusion that in the initial period after phagocytosis, much of the HOCl generated should react with protein targets and phagosomal protein should be highly oxidized and possibly inactivated. To some extent this has been observed (40, 196). Some HOCl should reach the bacterium, either from bound MPO or from the phagosomal lumen as protein targets are depleted. However, the model implies that a substantial excess of HOCl would need to be generated in order for the bacterium to receive a cytotoxic dose. As amines are the most prevalent oxidizable groups, protein chloramines should be formed. These could break down to give more diffusible and cytotoxic ammonia chloramines that could contribute indirectly to HOCl-dependent killing. In subsequent sections we assess how well these predictions fit with experimental observations.
Evidence for Oxidants in the Neutrophil Phagosome
While modeling leads to predictions of what redox reactions should occur in the phagosome, hard evidence for many of the reactions still needs to be sought. Many of the probes and biomarkers used to detect reactive oxygen species lack specificity, which makes it difficult to attribute effects to a particular oxidant (208). There is convincing cytochemical data for intraphagosomal generation of superoxide and H2O2 (158, 160, 183) and the identification of 3-chlorotyrosine as a specific biomarker (48) soon led to irrefutable evidence that HOCl is formed during phagocytosis. Tyrosine was loaded into sheep red blood cells and converted to 3-chlorotyrosine when the cells were ingested by neutrophils (80). Incorporation of chlorine into fluorescein bound to phagocytosable particles reinforced this finding (93), and the high yield of chlorofluorescein derivatives suggested that the amount was sufficient to kill entrapped bacteria. In a recent study, dichlorodihydrofluorescein conjugated to yeast was oxidized and chlorinated inside the phagosomes of a myeloid cell line (189). In most phagosomes, this process ceased after ∼10 min. In others, it continued for longer but at a slower rate. In all of these studies, chlorination required NADPH oxidase activity and MPO.
Formation of 3-chlorotyrosine in bacterial proteins after ingestion of Staphylococcus aureus established that HOCl produced in phagosomes reacts with ingested bacteria (38, 162). On the basis that protein chloramines are intermediaries in the chlorination of tyrosine residues, it can also be implied from this result that chloramines must be formed on neutrophil and bacterial proteins. Others showed that the extent of chlorination of bacterial proteins after phagocytosis of Pseudomonas aeruginosa was decreased by 75% when the neutrophils were from individuals with cystic fibrosis (138). This result provides support for the proposal that the CFTR delivers chloride to phagosomes and enables MPO to maintain production of HOCl.
When S. aureus were ingested by neutrophils, 3-chlorotyrosine measurements indicated that most of the HOCl reacted with neutrophil rather than bacterial proteins (38). In this study it was also uncertain whether the bacteria were exposed to sufficient HOCl to kill them. However, this biomarker is not ideal for quantifying production of HOCl as it accounts for a small percentage of the oxidant. Nevertheless, similar doubts were raised when iodination of bacterial and host cell proteins was compared (157). These findings, plus observations that most of the host chlorination occurs in the phagosome (73), fit with the modeling data that suggest that proteins bathing the bacterium are major targets for HOCl.
Expression of green fluorescent protein (GFP) in the cytosol of bacteria was a clever way of detecting HOCl during killing of ingested microorganisms. GFP is highly sensitive to HOCl but not to chloramines or other oxidants (140, 168). In both studies bleaching of GFP in phagocytosed bacteria was reported and the degree of bleaching indicated that sufficient HOCl had reacted with the bacteria to kill them. The only conundrum is that viability was lost much more rapidly than the GFP was bleached. Chlorination of the fluorescent tyrosyl residue in GFP is expected to be slow, so it may not occur until the more reactive reductants inside the bacteria are consumed. Measurement of methionine sulfoxide in bacterial proteins after killing of ingested microorganisms also demonstrated that enough oxidant reacted with them to be responsible for their demise (162). NADPH oxidase, MPO, and chloride were required, implicating HOCl or a species derived from it such as a chloramine.
In recent years, specific and sensitive probes for HOCl have been developed (39, 96, 109, 213). Fluorescence measurements with rhodamine-based probes have confirmed production of HOCl in phagosomes in real time (Fig. 8) and HOCl-specific cadmium-selenide quantum dots have also demonstrated phagosomal HOCl production. Further work with these probes should enable a superior kinetic appreciation of the production and modulation of HOCl inside phagosomes than is currently available.
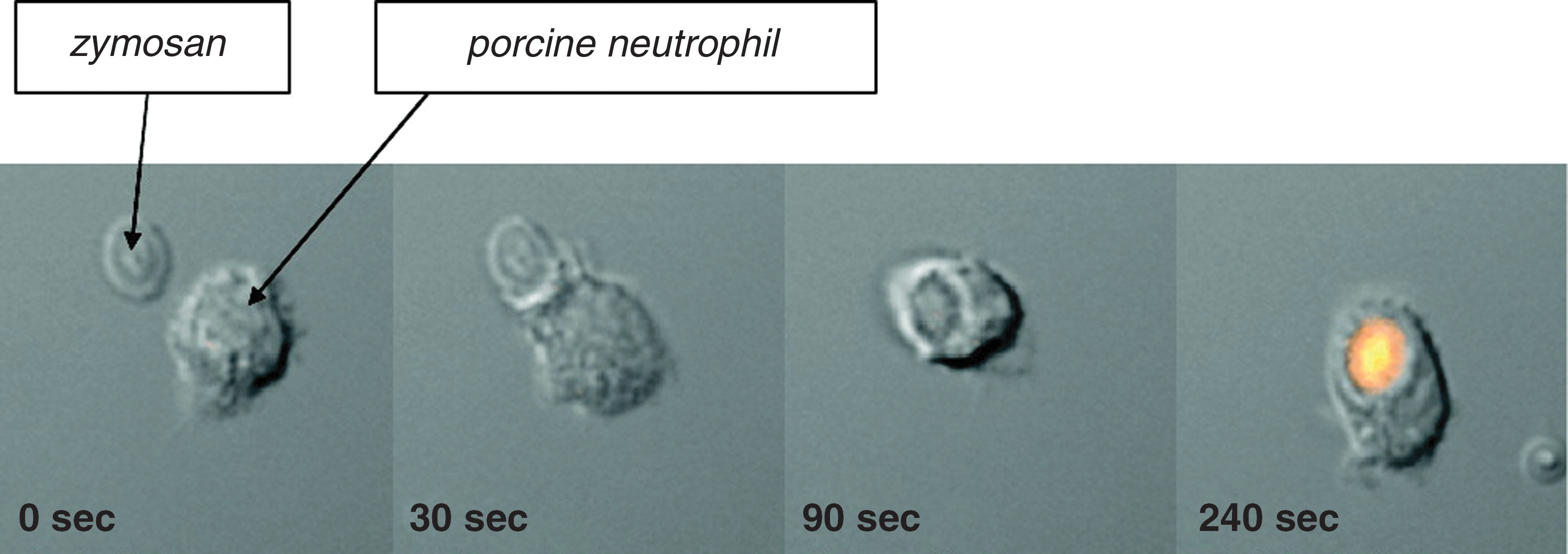
Collectively, these studies provide overwhelming evidence that HOCl is produced in the phagosome. Most studies suggest that the amount formed is adequate to kill the ingested bacteria. What remains to be determined is exactly how much of the oxygen consumed by neutrophils is converted to HOCl, how much reacts with bacteria compared with neutrophil proteins, and whether chloramines formed on neutrophil proteins contribute to killing.
Other oxidants must also be formed inside phagosomes. There is good evidence that 3-nitrotyrosine (57) and oxidized lipids (151) are produced. Both involve radical mechanisms. Oxidation of nitrite by MPO is unlikely to occur because even at high millimolar extracellular nitrite concentrations no nitration of a fluorescein probe was seen (93). Nitrated tyrosine is more likely to originate from peroxynitrite through the NADPH oxidase and NO synthase working in tandem. Formation of cytotoxic amounts of peroxynitrite has been detected in phagosomes of mouse macrophages (7). However, these do not contain MPO and whether superoxide reacts with NO in neutrophils has yet to be determined.
Oxidants and Microbial Killing
From the evidence presented, it is apparent that phagocytosed bacteria are exposed to a high flux of superoxide. H2O2 and HOCl are produced and there appears to be sufficient HOCl produced to be microbicidal. There is therefore a credible argument for HOCl being responsible for oxidative killing, either directly or via chloramines, and acting as the frontline defense against most microorganisms. It is not the only defense, as a range of antimicrobial peptides and proteins, including defensins, cathelicidins, serine proteases, and lactoferrin, are also released into phagosomes and are capable of killing by nonoxidative mechanisms (25, 56, 118). Nauseef has described these as “a coalition of antimicrobial elements, some transient and others long lived, that collaborate to create an inhospitable environment optimized to damage or kill most ingested microbes.” Within this context, is it possible to define specific roles for oxidative killing, MPO, and HOCl?
It is evident from the problems with infection seen in CGD, that NADPH oxidase activity is essential for broad spectrum antimicrobial defense. The most straightforward explanation is that if superoxide is not generated in the phagosome, oxidative killing is defective. There is strong evidence to support this. It has long been recognized that neutrophils isolated from patients with CGD have impaired killing of a wide range of microorganisms (107, 172). They cope as well as normal cells with some bacteria but struggle with most. Supplying CGD neutrophils with H2O2, although not fully recapitulating superoxide production by the NADPH oxidase, often rectifies their killing defect (68, 90), and strains of Streptococcus pyogenes that generate their own H2O2 are killed by CGD cells whereas nonproducers remain viable (165).
Neutrophils isolated from individuals with MPO deficiency are also defective in killing many microorganisms (107). This is mirrored by treatment of neutrophils with azide, or a specific MPO inhibitor (188). In one study, the time to kill 50% of ingested S. aureus was 10 min for normal cells, 38 min when the NADPH oxidase was inhibited, and 24 min for MPO-deficient cells (76). Thus, the majority of killing was reliant on the NADPH oxidase and strongly dependent on MPO. The finding that SOD coupled to S. aureus inhibited killing by 30% reinforces the concept that the functions of superoxide and MPO are linked. The experimental evidence discussed previously strongly suggests that phagocytosed bacteria are exposed to HOCl in sufficient amounts to kill. In particular, the extent of MPO-dependent oxidation of methionine-containing peptides in phagocytosed Escherichia coli indicated that they had received a lethal dose (162). This recent study clinched the argument that HOCl or chloramines are bactericidal within phagosomes.
The main conundrum is the difference in phenotype between MPO deficiency and CGD. Individuals with MPO deficiency rarely suffer from serious infection (113, 132, 142) yet most cases of CGD present with recurrent and life-threatening illnesses (172, 178, 202). At first glance this suggests that MPO plays a minor role in microbial killing. However, this conclusion may be simplistic and mask the importance of HOCl. It is not surprising that CGD has greater consequences than MPO deficiency when it is considered that NOX2 is present in other cells of the immune system (135). Consequently, cells that work with neutrophils to fight infection will also be impaired. In addition, there are aspects of CGD that cannot be attributed to defective killing. The formation of granuloma (from which the disease got its name) can be induced experimentally in gp91-deficient mice by administration of dead bacteria (128). There is also evidence of prolonged inflammation and other hyper-inflammatory conditions associated with CGD (11). One contributor may be impaired clearance of neutrophils from sites of inflammation. The normal process involves apoptosis; exposure of surface markers, including phosphatidyl serine; and removal of the apoptotic cells by macrophages (166). NADPH oxidase (but not MPO) products contribute to the mechanism (95), and CGD neutrophils, or those treated with an oxidase inhibitor, have altered expression of proteins in the apoptotic pathway (108) and show defective phosphatidyl serine exposure and uptake by macrophages (75).
Although microbial killing by cells deficient in MPO is impaired, it is not absent. Nonoxidative mechanisms will still function and other oxidative processes may become more efficient. For example, the rise in concentration of superoxide to ∼100 μM could make it toxic, either directly or through the hydroperoxyl radical that would exist at ∼1 μM under these conditions. Although H2O2 would not reach a high concentration, more would diffuse into bacteria. A third possibility is that the high phagosomal concentration of superoxide should favor peroxynitrite production. This may be combined with increased inducible NO synthase expression and NO production, as observed in MPO knockout mice (30). These animals were not compromised by lack of HOCl production and in fact had decreased mortality compared with normal mice.
In some instances oxidative and nonoxidative killing mechanisms may be redundant or the prevalent mechanism may vary depending on the surroundings of the neutrophil. This is discussed in more detail elsewhere (85). It can be illustrated by a study with E. coli (163) in which DNA synthesis was readily inhibited by reagent HOCl when they were killed by normal neutrophils. However, these bacteria were killed equally well by CGD and MPO-deficient neutrophils and DNA synthesis was unaffected. This result implies that even though HOCl was produced, nonoxidative mechanisms were just as efficient at eradicating the bacteria. Some microorganisms may evade oxidative killing entirely. One example is the killing of Streptococcus pneumoniae by neutrophils, which was shown to require serine proteases but not NADPH oxidase activity and oxidants (179). Other organisms may resist toxic oxidants with virulence factors, such as release of superoxide dismutase (20, 112, 117) or production of the carotenoid staphyloxanthin (120), or upregulate protective mechanisms in response to an oxidant stress (85). These results illustrate the point that while oxidative processes are important, they may not kill all bacteria and/or are a reliable back up if other mechanisms fail.
A key observation from experimental studies is that whereas MPO-dependent killing by HOCl tends to be a quick process, antimicrobial peptides and other oxidative processes act over a longer period. Therefore, it could be that the slower processes that operate in MPO deficiency are able to cope with low-level exposure to pathogens and mild infections. However, they could be overwhelmed if an MPO-deficient individual is exposed to a high bacterial insult. Indeed, MPO knockout mice are more resistant to infection than CGD mice but rapidly succumb when challenged with a large inoculum of several bacterial types (8, 9). This could explain why few complications of MPO deficiency are seen.
Conclusions and Outstanding Issues
The tiny space of the phagosome is an inhospitable environment in which neutrophils use a number of mechanisms to destroy most microorganisms. Although there are areas of uncertainty, most of the available information can be accommodated by the following working hypothesis for how killing occurs. We propose that the joint action of the NADPH oxidase and MPO provides the frontline defense in subjecting the ingested organism to an initial high flux of HOCl. In most cases HOCl or products derived from it are lethal, but organisms that evade this onslaught are killed by longer acting antimicrobial proteases and peptides. With some organisms, nonoxidative processes will be fast enough to provide early killing and dominate the process whereas others will evade one or more mechanisms. In MPO deficiency, nonoxidative processes and oxidative processes dependent on superoxide and H2O2 come into play. These alternative mechanisms are generally much slower than MPO-dependent killing, and may be adequate for handling low-level infection but not a massive insult.
Admittedly, this simplified model is not all encompassing, and there are a number of key questions to be answered. First, we need to know whether entry of chloride into the phagosome can keep pace with the enormous flux of superoxide and maintain HOCl production, and the mechanism by which this occurs. HOCl reacting with phagosomal proteins seems paradoxical because it should decrease bactericidal efficiency and inactivate granule proteins. The consequences of protein chlorination and the involvement of chloramines in killing need further investigation. More information is needed on whether the electrogenic action of the NADPH oxidase has a purpose other than generating superoxide, whether superoxide has a function in its own right in the phagosome, whether peroxynitrite is formed, and what oxidative killing mechanisms might operate in the absence of MPO.
Footnotes
Acknowledgments
The authors' work on neutrophil oxidants has been supported by grants from the Health Research Council of New Zealand.