
Abstract
Introduction
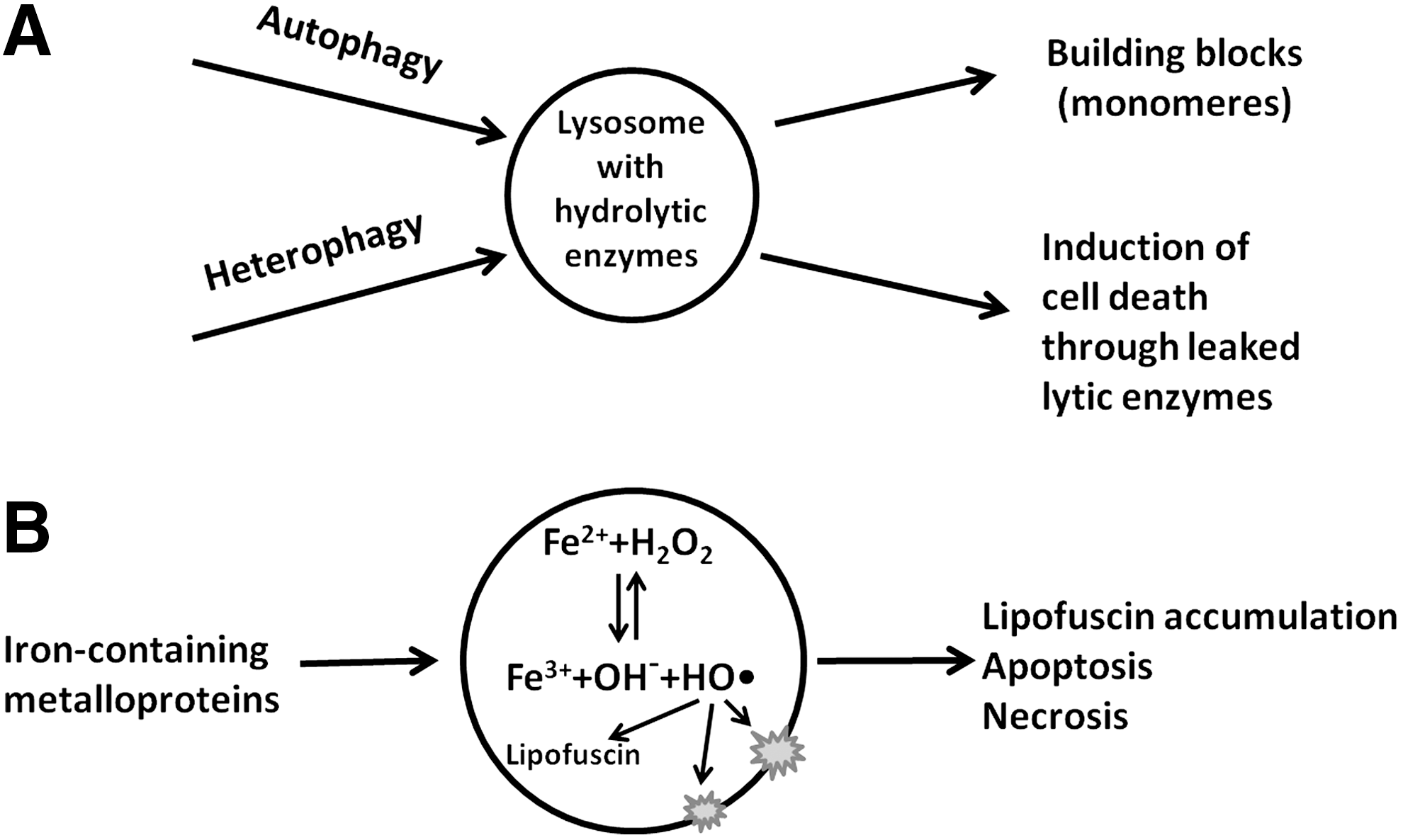
Apart from this, lysosomes function in repair after, for example, ionizing irradiation or oxidative stress. This so-called reparative autophagocytosis removes damaged cell components and organelles (8, 36). Under starvation conditions, lysosomes may help cells survive by digesting some of the cell's own components to provide building blocks for the synthesis of important macromolecules (19). In some cell types, lysosomes, or their content, can be exocytosed (49). It has been described that tumor cells use this mechanism to secrete lysosomal proteases in order to infiltrate surrounding tissue (62).
When metalloproteins are turned over, metals (such as iron, zinc, and copper) are released within lysosomes (42, 58). Iron is liberated, in particular, due to the degradation of cytochromes, hemoglobin and myoglobin, catalase and other enzymes. Moreover, lysosomal degradation is believed to be the principal mechanism of iron release from ferritin, the main cellular iron-binding protein (4, 56). As a result, lysosomes contain much higher quantities of iron than other organelles and are probably the only place where iron exists in a low-mass redox-active form (42) (Fig. 2). The amount of intralysosomal iron has a tendency to increase with age due to the incapability of cells to release iron (24).
When cells divide, many new biological structures are synthesized, while damaged cellular components are distributed to the daughter cells and thereby diluted. This decreases the demand on autophagy and is consistent with the low degree of autophagy found in mitotically active cells (26). In contrast, nondividing cells, such as differentiated neurons and cardiac myocytes, require intensive autophagy for their homeostasis (79).
There is abundant evidence that insufficient autophagic activity may contribute to malignant transformation by promoting increased oxidative stress, genomic instability, and accumulation of damaged organelles (67). Despite active proliferation, transformed cells are usually characterized by increased autophagy and generally up-regulated lysosomal activity. This allows them to cope with degenerative changes due to altered metabolism and disturbed circulation (38, 50). In addition, they secrete increased amounts of lysosomal proteases, promoting angiogenesis as well as tumor growth and invasion (38). Based on these properties of tumor cells, autophagic inhibition can be used as an anticancer strategy (72). The destruction of cancer cells can, for instance, be stimulated by the induction of lysosome-mediated cell death mechanisms through agents that promote metal toxicity (32, 48). This review is focused on the role of lysosomal iron and, to some degree, on copper in cell death and on possible anticancer strategies.
Lysosomal Degradation Function
Lysosomes contain numerous acid hydrolases that are capable of degrading a variety of biological molecules, including proteins, lipids, carbohydrates, and nucleic acids (55). The acidic pH of the lysosomes (between 4.5 and 5.0) is provided by the function of vacuolar ATPase, an enzyme that uses ATP for pumping protons into the lysosomal lumen (51). Extracellular material enters cells for heterophagy by invagination of the plasma membrane, resulting in the formation of early endosomes (endocytosis). Intracellular substances can be delivered to lysosomes in three different ways, corresponding to three types of autophagy: macroautophagy, microautophagy, and chaperone-mediated autophagy (CMA) (72). The principal lysosomal degradation pathways in mammalian cells are presented in Figure 3.
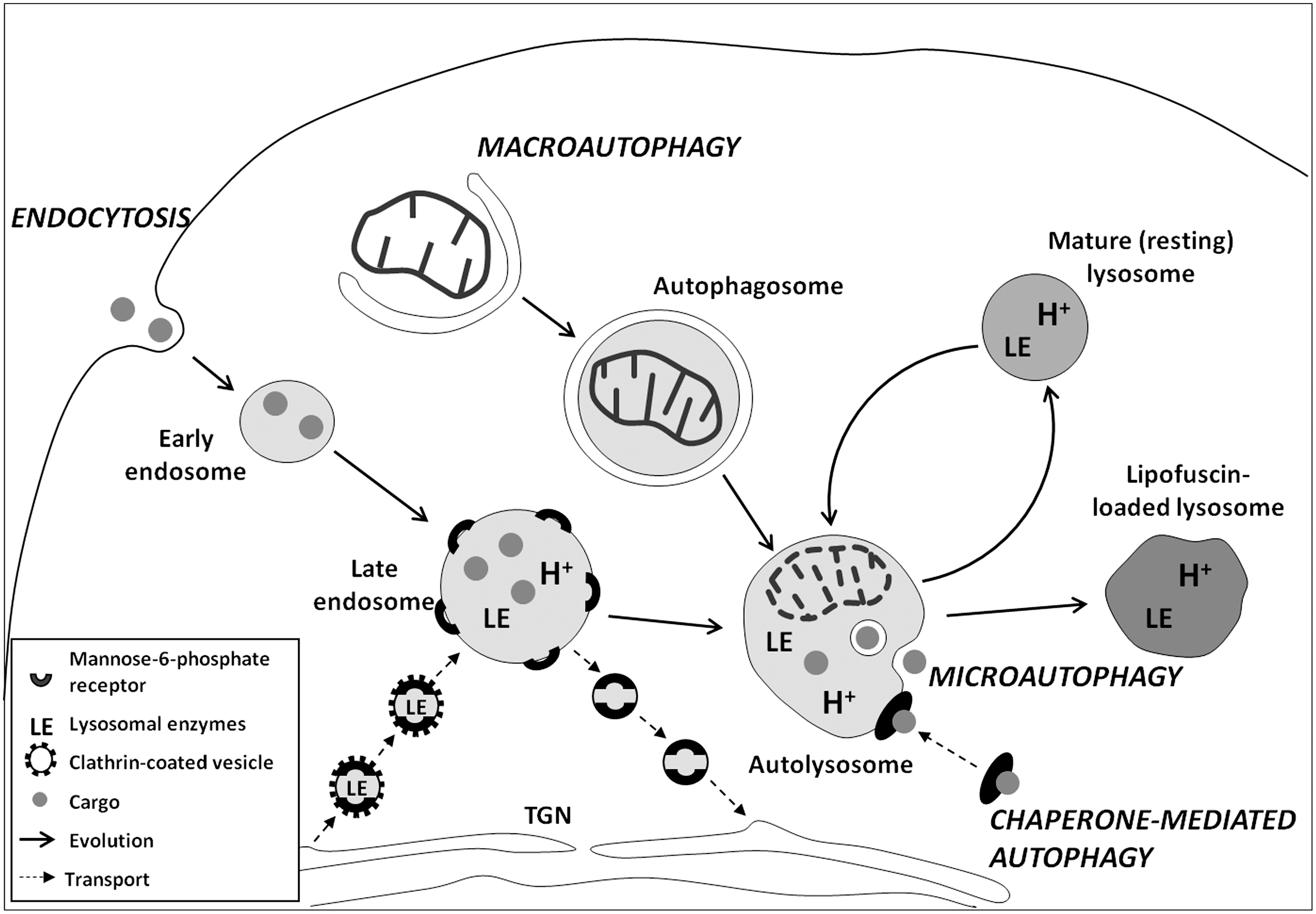
In macroautophagy, portions of cytoplasm that can contain organelles, such as mitochondria, are sequestered by membrane cisterns (phagophores) that form autophagosomes, which are double-membrane-bounded vacuoles. Neither early endosomes nor autophagosomes are acidic. They are also devoid of lysosomal enzymes. If they fuse with each other, amphisomes are formed (86). Early endosomes gradually progress to late endosomes and mature lysosomes. This process is associated with increasing endosome acidification and the transport of lysosomal enzymes by small vesicles that pinch off from the trans-Golgi network (TGN). Lysosomal enzymes are delivered in mannose-6-phosphate receptor (MPR)-bound form, suggesting that late endosomes as well as Golgi-derived vesicles and TGN contain MPR. The acidic pH triggers the release of acid hydrolases into the endosomes, while MPR are recycled back to the TGN (49). Mature lysosomes differ from late endosomes by a lower pH, a higher concentration of acid hydrolases, and the absence of MPR, indicating that the transport of the lysosomal enzymes is completed. It is suggested that autophagosomes and amphisomes can also receive acid hydrolases from Golgi-derived vesicles (82). Alternatively, autophagosomes and amphisomes receive lysosomal enzymes by fusion with late endosomes or mature lysosomes, suggesting that autophagic and heterophagic degradation pathways work together (86).
Molecular mechanisms of macroautophagy have been extensively investigated through analyses of yeast mutants with affected vacuolar degradation (53). More than thirty proteins (Atg proteins) and corresponding genes (ATG-genes) involved in macroautophagy in yeast, as well as their mammalian orthologs, have been identified (87). These proteins are responsible for the initiation of autophagy, formation and expansion of the phagophore, closure and completion of the autophagosome, and its docking and fusion with endosomes and/or lysosomes. A detailed description of molecular processes involved in macroautophagy is reviewed elsewhere (52, 72, 87). In some cases, macroautophagy can occur in the absence of certain Atg proteins (so-called noncanonical autophagy) (18).
In microautophagy, portions of the cytoplasm enter endosomes or lysosomes through invagination of the limiting membrane, often resulting in the formation of multiple intralysosomal vesicles. This pathway provides for the degradation of soluble macromolecules and small organelles (72). Although a number of proteins involved in microautophagy have been identified in yeast (46), the molecular machinery of this process in mammals remains unknown. It may be an automatic process, as lysosomes in a percoll solution spontaneously show the invagination of a microautophagocytotic type (1).
CMA, described in higher eukaryotes, is a selective process. Specific proteins with a particular pentapeptide motif KFERQ (for lysine-phenylalanine-glutamate-arginine-glutamine) are recognized by a heat shock cognate protein of 70 kDa (HSC70). The complex then binds at the lysosomal outer surface to the lysosome-associated membrane protein type 2A (LAMP-2A) (72). On binding, the substrate is transported across the lysosomal membrane.
Autophagy is an ongoing self-renewal process that occurs at a basic level under normal conditions but increases dramatically due to starvation, hypoxia, oxidative stress, ionizing radiation, and other forms of damage (40, 76). The regulation of macroautophagy has been more extensively studied and is better understood than that of microautophagy and CMA. Transcription factor EB (TFEB) and mammalian target of rapamycin complex 1 (mTORC1) are believed to play a central role in the regulation of macroautophagy and lysosomal biogenesis in mammals (69, 71). Under normal conditions, mTORC1 is located at the lysosomal surface and phosphorylates TFEB, preventing its translocation to the nucleus. Under starvation and other stress conditions, mTORC1 is detached from the lysosomes and inactivated. As a result, TFEB is not phosphorylated and, therefore, translocates to the nucleus, triggering the expression of multiple genes that are responsible for lysosomal biogenesis and autophagy [described in detail in (71)].
The bulk of cellular degradative activity is performed by lysosomes. However, many proteins, mainly short-lived ones, are degraded by multicatalytic proteinase complexes, proteasomes (57), and calpains. The latter are calcium-dependent neutral proteases that are active in the cytosol and mitochondria (35, 73). Mitochondrial proteins can also be degraded by matrix Lon and membrane-bound AAA proteases (3, 6).
Lysosomes and Iron Metabolism
Iron circulating in the bloodstream is mostly transferrin-bound Fe(III) that enters cells via receptor-mediated endocytosis. Due to acidification of iron-containing endosomes (called siderosomes), iron is released, while transferrin and its receptor are recycled back to the plasma membrane. Fe(III) is then reduced to Fe(II) by ferrireductase and transported to the cytosol by the divalent metal transporter 1 (DMT1) (56). There are also indications that such endosomal iron can be directly transferred to mitochondria through a “kiss and run” mechanism (65).
Cytosolic iron is mainly stored in nonredox-active form as a component of ferritin (i.e., iron combined with apoferritin), which possesses ferroxidase activity, transforming Fe(II) to Fe(III) (10, 56). When iron is needed for synthetic processes, it is released from ferritin. There is evidence that this occurs intralysosomally through the autophagic degradation of ferritin (29, 37, 45). Due to the acidic pH of lysosomes and the presence of reducing agents such as cysteine and glutathione, Fe(III) is converted to the redox-active Fe(II) (42, 70). Another reducing agent inside lysosomes contributing to the formation of Fe(II) may be the superoxide anion radical (O2 •–) that can form in lysosomes from mitochondria undergoing autophagic degradation. Conceivably, the production of superoxide can even increase in such mitochondria due to the disabling of the electron transport complexes by lysosomal proteases (78). Fe(II) can then be relocated to the cytoplasm, probably by DMT1 or mucolipin 1 (23, 42). Some iron may be released from ferritin after its degradation by proteasomes (20, 89). It has also been suggested that iron can be directly released through pores in ferritin molecules (47, 74), although the evidence for this mechanism is weak (Fig. 4).
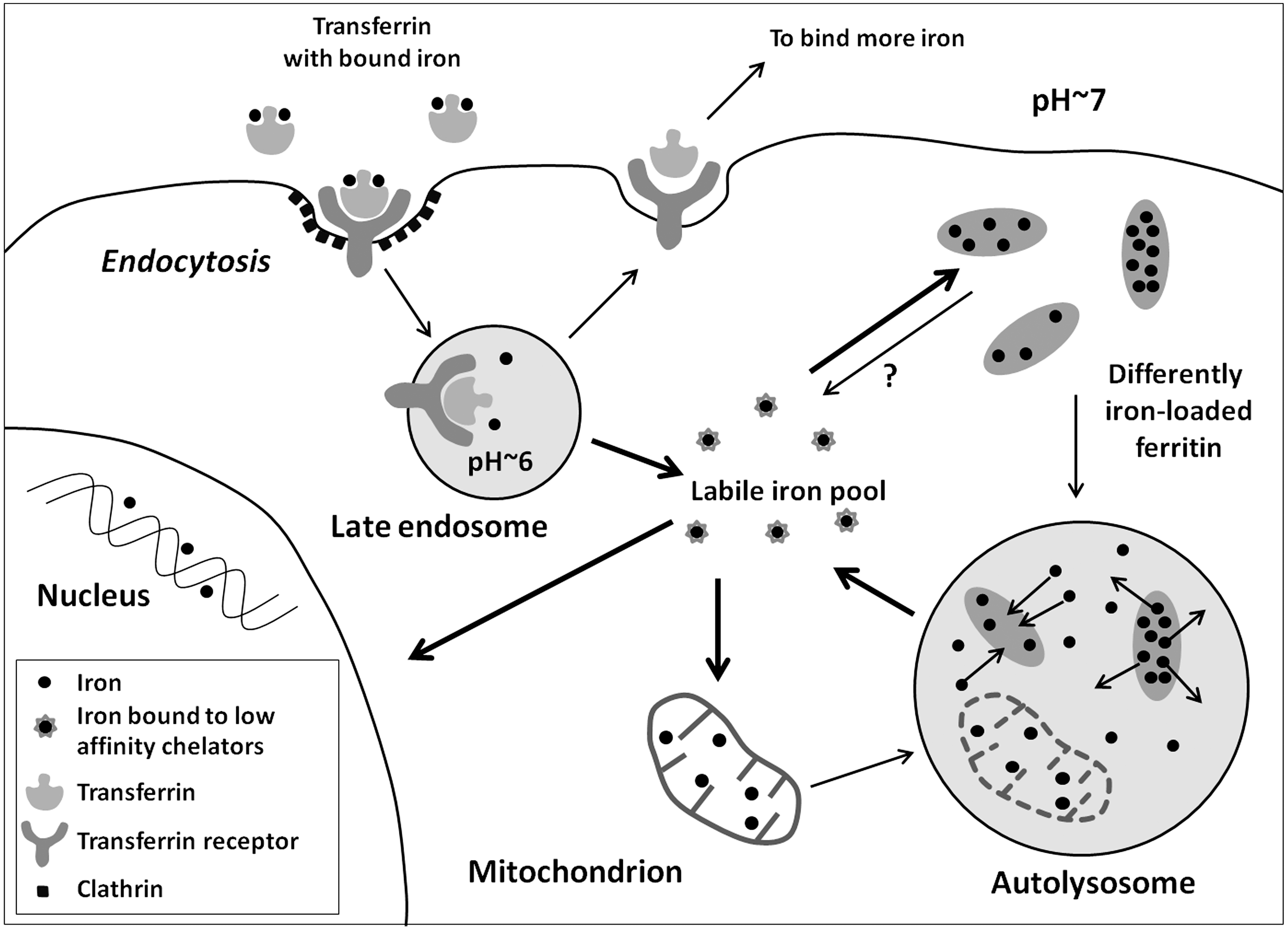
Apart from ferritin, lysosomes degrade a number of other iron-containing macromolecules, including cytochromes and several enzymes (42). In cardiac myocytes and skeletal muscle fibers, a large part of lysosomal iron is derived from autophagocytosed myoglobin, while macrophages receive iron from heterophagocytosed erythrocytes (56). Due to the continuous degradation of ferruginous materials, lysosomes are particularly rich in low mass iron, which is mainly present in its redox-active Fe(II) form (see above). This makes lysosomes particularly sensitive to oxidative stress as compared with other organelles (42). In iron overload, such as in hemochromatosis, lysosomes accumulate considerable amounts of iron, mainly in the form of hemosiderin. This brown insoluble pigment is basically derived from denatured, incompletely degraded ferritin and contains FeO in its core (63).
Even under normal conditions, proteins (including iron-containing ones) are not completely degraded. This is the result of a continuously ongoing oxidative damage to macromolecules under autophagic degradation. As a consequence, long-lived postmitotic cells, such as cardiac myocytes and neurons, progressively accumulate lipofuscin (age pigment), an intralysosomal indigestible, autofluorescent material (13, 81). Lipofuscin has been shown to contain high amounts of iron and consists of protein residues that are linked by aldehyde bridges (34). Lipofuscin-rich cells show increased sensitivity to oxidative stress, probably due to the high content of iron within lipofuscin-loaded lysosomes (75). Most importantly, progressive lipofuscin accumulation decreases the lysosomal degradative capacity of aged cells (77). As a result, aged postmitotic cells cannot effectively recycle their damaged mitochondria and, consequently, die because of lack of ATP. This deteriorative mechanism is described in detail in the mitochondrial-lysosomal axis theory of aging (14, 81).
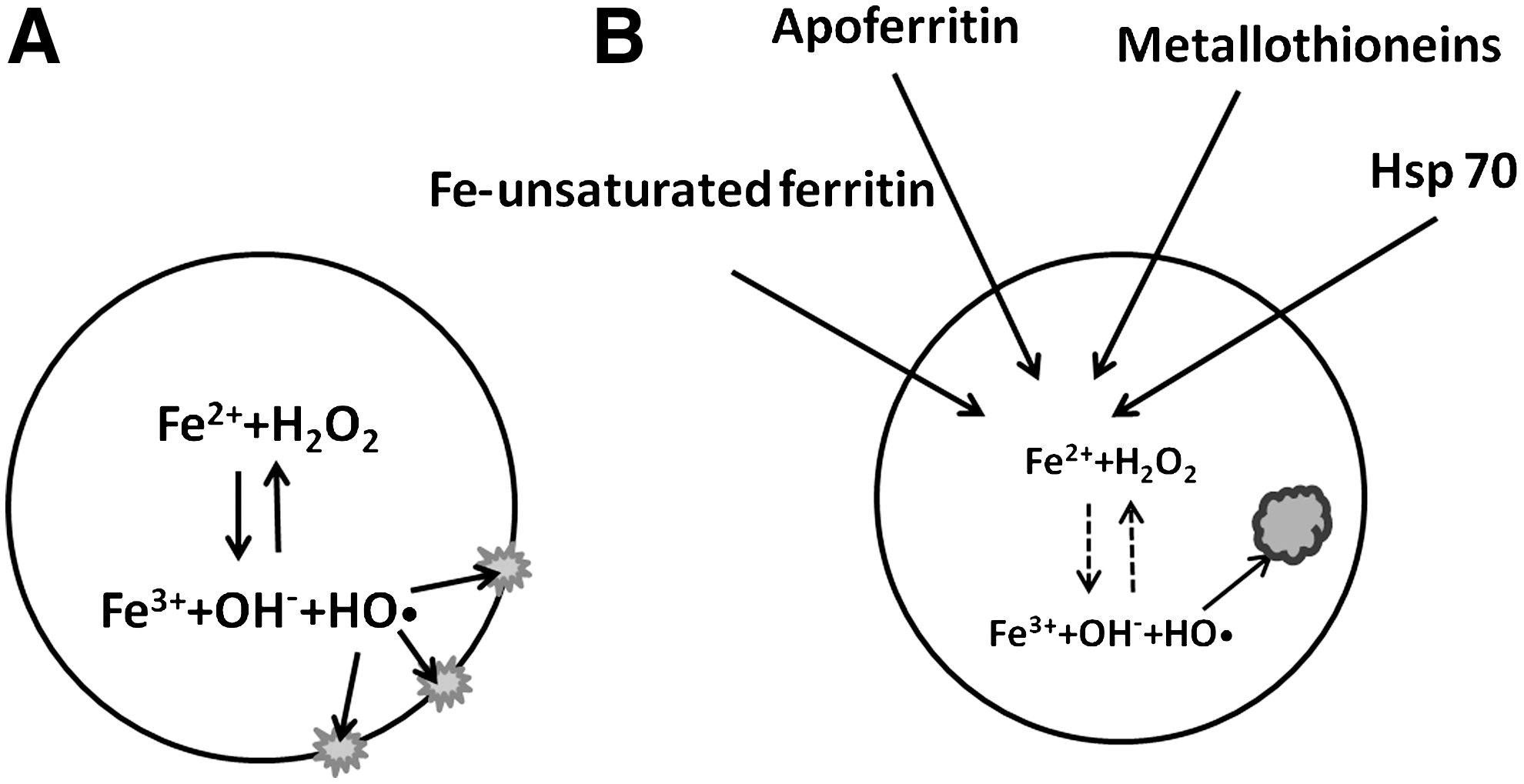
When iron-binding proteins, such as apoferritin, ferritin with remaining iron-binding capacity, metallothioneins, and heat shock protein of 70 kDa (HSP70), enter lysosomes for recycling, they can temporarily bind redox-active iron before being degraded. Thereby, they decrease oxidant-induced lysosomal membrane permeabilization (LMP) and consequent apoptosis (42) (Fig. 5). This may explain why certain malignant cells, being rich in iron-binding stress proteins, show resistance to ionizing radiation and antineoplastic drugs (9, 60). In contrast, when ferritin is highly saturated (e.g., in hemochromatosis), the lysosomal compartment is enriched with iron, and cells are highly sensitive to oxidative injury (42).
Lysosomal Iron and Oxidative Stress
Normal biologic respiration is associated with continuous electron leak from the mitochondrial electron transport chain, resulting in the continuous formation of hydrogen peroxide that easily passes biological membranes. In addition, hydrogen peroxide is produced by various oxidoreductases that operate by one-electron transfers. Although catalase and glutathione peroxidase effectively degrade most hydrogen peroxide, some of it may diffuse into lysosomes before being degraded [reviewed in (80)].
As mentioned earlier, lysosomal iron is present mainly in its Fe(II) form, being able to catalyze the Fenton reaction, that is, a homolytic splitting of hydrogen peroxide yielding the extremely reactive hydroxyl radical:
Hydroxyl radicals (with the half life of around 10−9 s) attack biomolecules undergoing autophagic degradation, causing their oxidative modification. Under normal conditions, with a relatively low, steady-state autophagic activity and low production of hydrogen peroxide, this results in a gradual accumulation of lipofuscin pigment within long-lived postmitotic cells (see above and Fig. 5). In enhanced autophagy (e.g., associated with the repair after cellular injury by hypoxia, ionizing radiation or various toxic agents, or when lysosomal degradation is inhibited), a more rapid formation of the pigment occurs. This pigment, which is similar to lipofuscin by the composition and mode of origin, is often called ceroid (13).
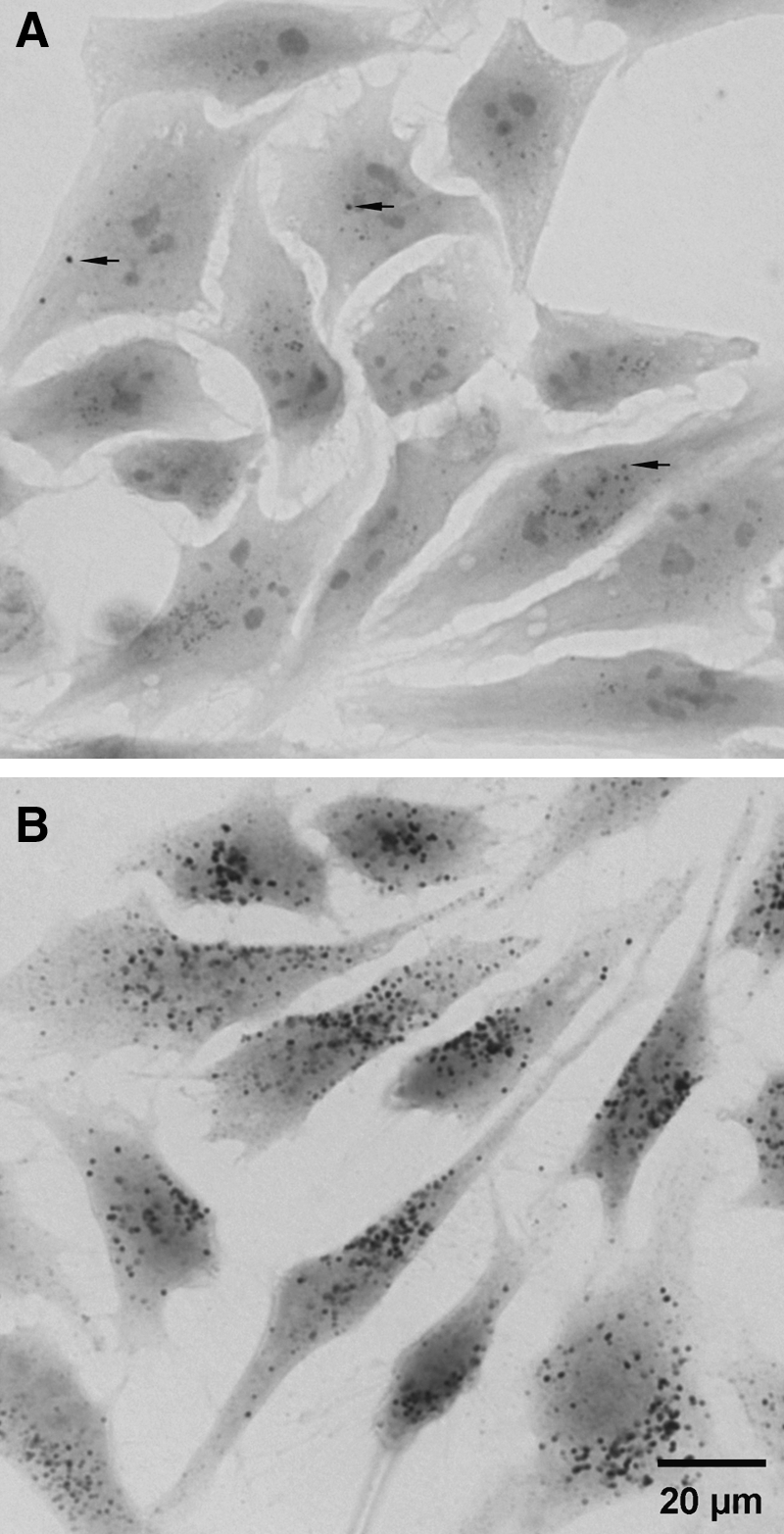
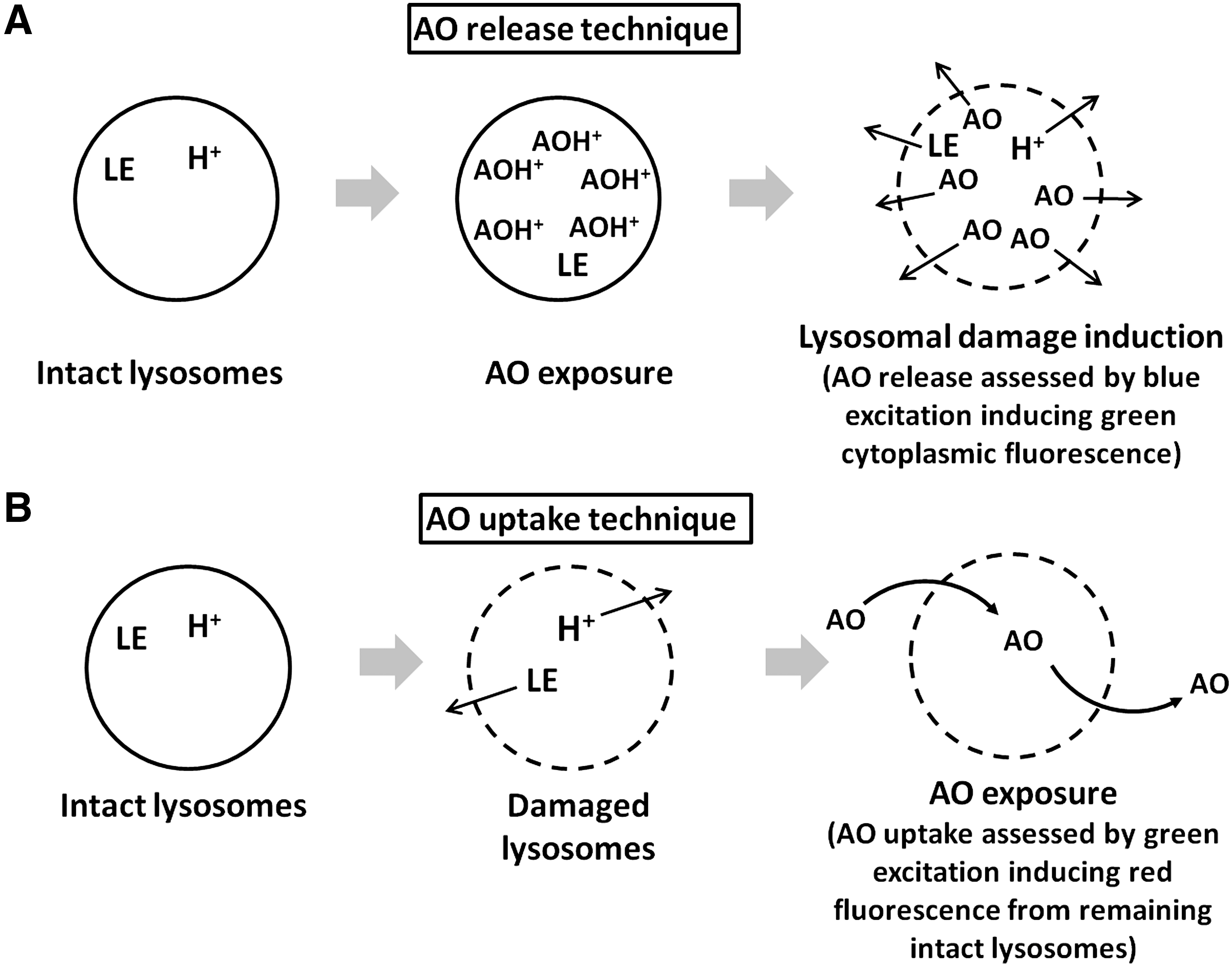
Lysosomes experience pronounced oxidative injury during enhanced oxidative stress, when cells are exposed to relatively high doses of oxidants, or in iron overload. This results in damage to their membranes and increased LMP. As a consequence, lysosomal enzymes are released to the cytosol, triggering apoptosis (see below). Lysosomes within the same cell show a wide variation in their sensitivity to oxidative stress. In cell culture models, the stability of lysosomes can be assessed by their uptake of acridine orange or other acidotropic fluorochromes (80) (Fig. 6). It was demonstrated that while some lysosomes can be damaged by a relatively slight oxidative stress, others remain stable even after exposure to severe oxidative stress (58). Such a variation in lysosomal vulnerability by oxidative stress can depend, in particular, on the different iron content of lysosomes (Fig. 2). Apparently, cells survive when only a few lysosomes are damaged, while the apoptotic machinery is activated when the injury is more pronounced (2).
Lysosomes and PCD
Apoptosis, or PCD, is an evolutionarily conserved mechanism that is used for the removal of redundant, damaged, or diseased cells (28, 30). Apoptosis is essential for the elimination of cells undergoing malignant transformation, as well as for tissue remodeling during development, that is associated with the periodic removal of certain cell groups (28, 30). During apoptosis, cells undergo a controlled enzymatic digestion and are phagocytosed by their neighbors or macrophages. This prevents excessive damage of surrounding tissues and consequent inflammation. A release of toxic substances from dying cells results in necrosis (85).
It is now well established that both caspases and autophagy can be responsible for the degradation of cellular constituents during apoptosis (28, 30). Morphologically, apoptosis is characterized by chromatin condensation and fragmentation of the nucleus and cytoplasm into apoptotic bodies. In some cases, multiple autophagic vacuoles form as well. This mode of PCD is sometimes called autophagic cell death (22). The distinction between classical (caspase-dependent) apoptosis and autophagic cell death is, however, obscure, as the processes involved in the two types of cell death are often combined and even work in concert (28, 30). Moreover, even in classical apoptosis, lysosomal enzymes significantly contribute to cell death (see below), suggesting the indirect involvement of autophagy in the process.
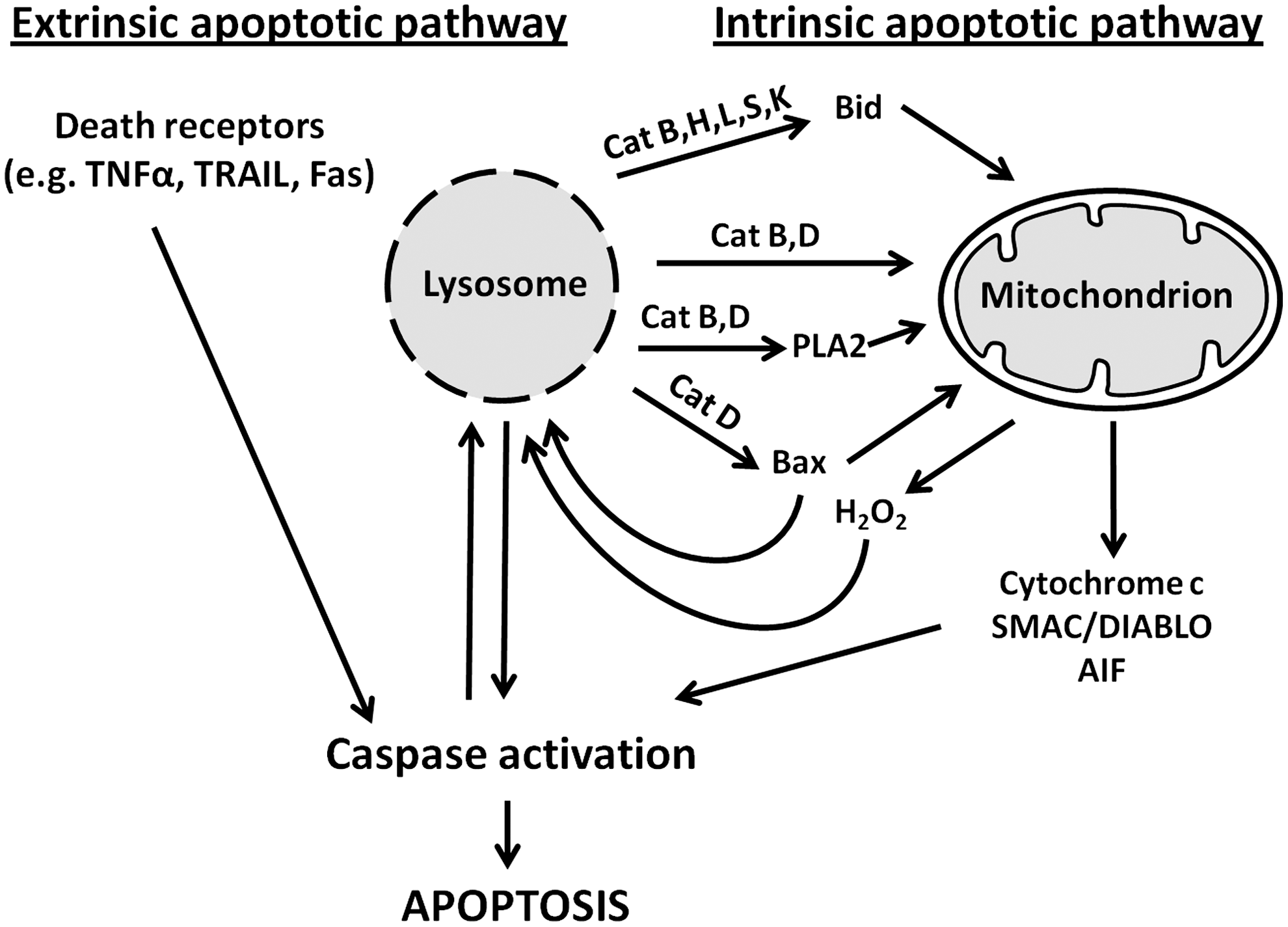
Caspases are a family of cytosolic cysteine endopeptidases that are activated by two pathways (33). The extrinsic apoptotic pathway is initiated after the stimulation of death receptors, such as tumor necrosis factor receptors, on the plasma membrane. The intrinsic apoptotic pathway is triggered in response to internal cell injury, which is mediated by mitochondrial depolarization and the release of cytochrome c, second mitochondria-derived activator of caspases/direct IAP binding protein with low pI (SMAC/DIABLO), and some other regulatory proteins. Signaling from death receptors leads to caspase-8 activation, while the mitochondrial pathway results in the activation of caspase-9. Caspases 8 and 9, called initiation caspases, activate execution caspases such as 3 and 7, which are responsible for the final cleavage of cellular constituents [extensively reviewed in (28, 30, 38)]. Both receptor-mediated and mitochondrial pathways may be interdependent and converge at the execution stage (28, 30) (Fig. 7).
When caspases are inhibited, injured cells are still able to undergo PCD due to mitochondria-to-nucleus translocation of apoptosis-inducing factor (AIF), causing DNA fragmentation and chromatin condensation (28, 30). In some cases, the progress of PCD can be disabled soon after its induction, for example, when viruses that infect cells encode anti-apoptotic proteins. Such cells develop necrosis-like death, which is called programmed (regulated) necrosis (27).
LMP, resulting in the release of acid hydrolases into the cytosol, was observed in various normal and malignant cells at early stages of apoptosis, often before any mitochondrial alterations (11, 38). Apparently, lysosomal enzymes, such as cathepsins B and D, induce mitochondrial permeability transition either directly or by activating phospholipase A2, promoting cytoplasmic translocation of cytochrome c, AIF, and other pro-apoptotic agents (90). Recent studies suggest that lysosomal enzymes can induce mitochondrial permeability transition more specifically, through the cleavage of BH3 interacting-domain death agonist (Bid) (cathepsins B, H, L, S, and K) or the activation of Bcl-2-associated X protein (Bax) (cathepsin D) (17, 38). In addition, cathepsins have been reported to directly activate caspases (7, 88). Lysosomal enzymes can also be involved in cell death that is secondary to other pro-apoptotic factors. In particular, the pro-apoptotic protein Bax, activated through death receptor signaling, has been shown to induce LMP, along with its previously known effect on the mitochondrial permeability transition (83).
Lysosome-Mediated Cell Death and Metal Chelators
There is extensive evidence that LMP often occurs secondary to oxidative stress, the degree of which greatly depends on the lysosomal content of low-mass iron (25, 44, 68). Moreover, due to preferential accumulation of excessive iron within lysosomes, LMP is apparently the main mechanism underlying cell death in situations of iron overload (42).
The role of lysosomal low-mass iron in apoptosis is consistent with numerous reports that show the protection of cells by iron chelators. Endocytosis of the potent iron chelator desferrioxamine (DFO) before the induction of apoptosis significantly increased the viability of various cell types (2, 12, 59, 91). DFO and other iron chelators have also been shown to have a radioprotective effect, suggesting that iron-catalyzed oxidation of lysosomal membranes and consequent LMP contribute to radiation-induced cell damage (43, 60). This is probably due to the fact that ionizing radiation triggers reparative autophagy (see above), resulting in increased lysosomal uptake of ferruginous materials and consequent accumulation of low-mass iron within lysosomes.
Apart from DFO, other iron chelators, such as the water-soluble 1-propyl-2-methyl-3-hydroxypyrid-4-one (CP22) and the lipid-soluble salicylaldehyde isonicotinoyl hydrazone (SIH), have been shown to protect cells from oxidative and radiation-induced damage. Unlike DFO, which accumulates within lysosomes leading to iron depletion, these chelators can be washed away without further negative consequences for iron metabolism (43, 54).
Neoplastic cells are characterized by increased uptake of iron, which is needed to facilitate their high proliferation rate (39). As noted earlier, they also show enhanced autophagic rate, resulting in the accumulation of large amounts of iron within the lysosomal compartment. Based on these properties of malignant cells, several anticancer strategies have been proposed. First, the inhibition of macroautophagy has been found to be effective in enhancing the toxicity of several antineoplastic drugs [reviewed in (72)]. Second, high intralysosomal iron content makes neoplastic cells increasingly sensitive to various oxidants and ionizing radiation (see above). Consistent with this, it has been found that radiation-resistant tumors contain high levels of metal-binding proteins, such as ferritin, metallothioneins, and heat shock proteins, all of which reduce the amounts of intralysosomal redox-active iron (5, 29, 41). Third, large doses of metal chelators can induce the iron starvation of malignant cells (39, 64).
Recently, a number of tridentate chelators with antineoplastic activity have been developed (64). Some of them have very low LD50 values and, interestingly, are also active when applied in the form of iron complexes (sic!), creating some doubt about their activity being exclusively a result of iron chelation. The structural formulas and pKa values of several of these metal chelators indicate that they are lysosomotropic and will undergo “proton trapping” in an acidic compartment (48, 64, 66). If these tridentate chelators bind copper as well as iron, the tridentate copper chelates should be very redox active at a 1:1 chelator/copper ratio (only three out of the four copper coordinates are bound) and be able to react with a variety of reducing agents, including cysteine that is abundant in the lysosomal compartment (61). In contrast, iron is bound at a 2:1 chelator/iron ratio with all six iron coordinates bound, thereby preventing redox activity. Copper is a much more potent catalyst of Fenton-type reactions than iron. Consequently, the small amounts of hydrogen peroxide that usually reach the lysosomal compartment by diffusion might be enough to induce substantial production of hydroxyl radicals in lysosomes, especially in malignant cells that may be already under increased oxidant stress.
Many malignancies have been found to be significantly richer in copper than normal cells [reviewed in (15, 31)], and we may, thus, assume that tumor lysosomes would be rich in low-mass copper. Neocuproine and diethyl dithiocarbamate are examples of such lysosomotropic copper chelators that may promote the redox activity of copper by binding some but not all coordinates leading to LMP. The well-known tendency of tumor cells to generate excess acid may further promote the accumulation of chelators that are susceptible to proton trapping. We expect to see in the near future the initiation of clinical trials on selected copper chelators, because they have already proved to be effective in malignant cells in culture (15, 16, 31). Recently, the group of Des Richardson demonstrated a particularly efficient antineoplastic activity of the lysosomotropic copper chelator Dp44mT (48, 84).
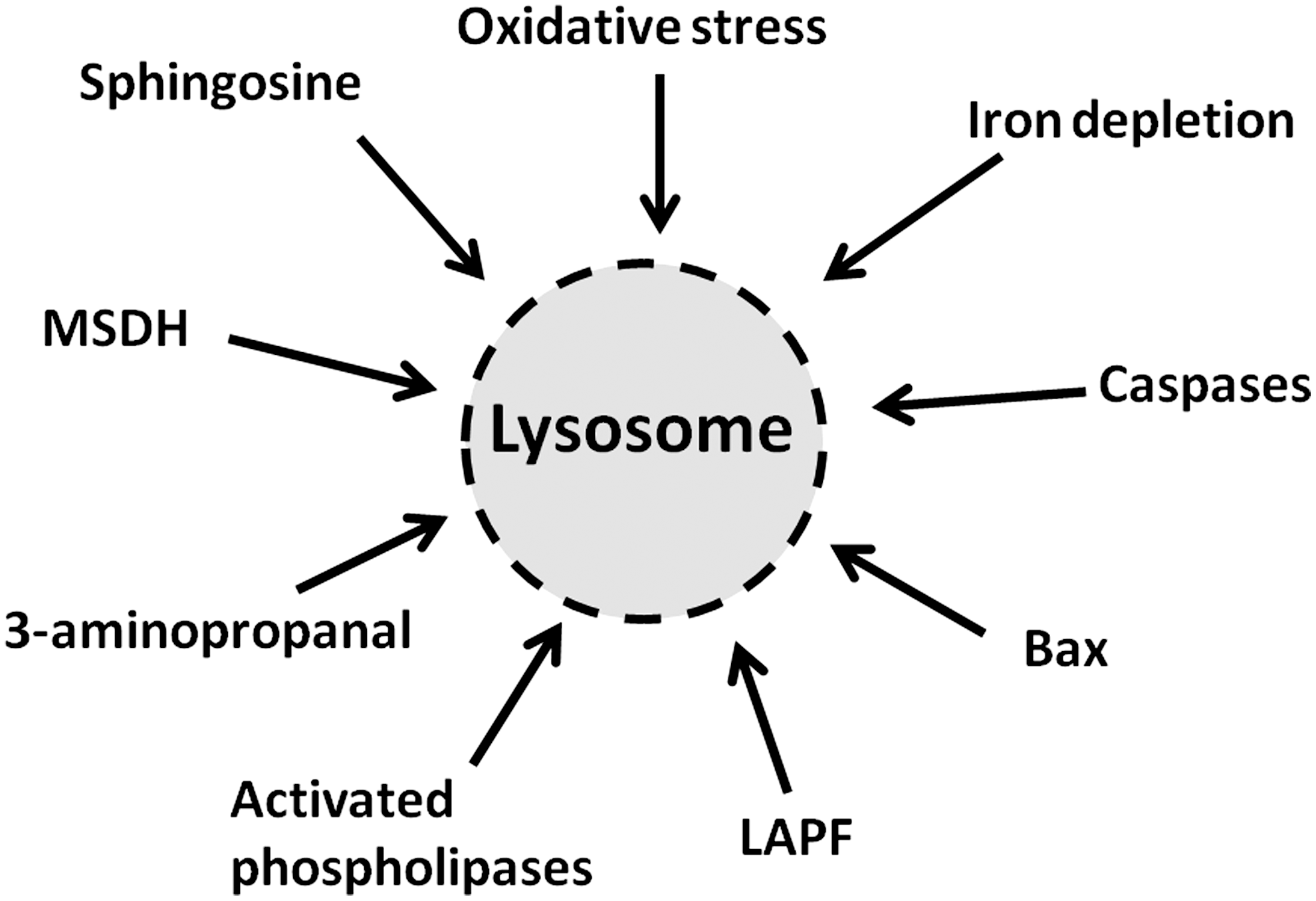
Surprisingly, apoptosis induced by iron depletion after exposure to SIH is preceded by lysosomal destabilization, indicating that lysosomal damage can be induced not only by iron-dependent oxidative stress but also by iron depletion (44). Oxidative stress occurring simultaneously with iron starvation due to general iron chelation should clearly not induce lysosomal labilization. Therefore, the destabilizing effect on lysosomes of iron starvation should be a consequence of other apoptogenic stimuli that are yet to be identified. The finding, nevertheless, provides additional support to the emerging view that LMP is a much more common phenomenon in apoptosis than has so far been recognized. A selection of molecules and processes that damage lysosomes is depicted in Figure 8.
Conclusion
Autophagy is a nonstop life-sustaining mechanism that provides for the efficient removal of damaged or worn-out cellular constituents, including various macromolecules, biological membranes, and organelles. Lysosomes degrade a variety of iron-containing proteins, which are delivered to them through autophagy or endocytosis, resulting in lysosomal enrichment with iron. As a potent pro-oxidant, iron promotes oxidative damage to macromolecules, making them undegradable and resulting in the progressive accumulation of intralysosomal waste material, lipofuscin, within long-lived postmitotic cells. Intralysosomal iron damages lysosomal membranes, resulting in their permeabilization and apoptosis induction. Based on this knowledge, antioxidant defense, in particular by using iron chelators, could be considered a possible anti-aging strategy.
Tumor cells are characterized by active autophagy and increased iron uptake that are needed to maintain their high proliferation rate. The use of iron chelators, leading to iron depletion, has been found to be effective in killing certain tumor cells. Similarly, copper chelators, resulting in copper chelation in a redox-active complex, have been found useful as well. Other lysosomal-targeted anti-cancer strategies involve the damage of iron-rich tumor cells by oxidants or ionizing radiation, as well as the inhibition of autophagy.
Footnotes
Acknowledgment
The authors would like to thank Professor John W. Eaton for helpful discussions and linguistic corrections.