
Abstract
Introduction
Sources and Biochemical Properties of ROS
There are many processes and reactions that result in the production of ROS within the cell. A major source of intracellular ROS in the aging process is superoxide anion radical O2 •− produced by mitochondria (Fig. 1). During the process of cellular respiration, O2 •− is continuously generated as a result of electron leak from the electron-transport chain. O2 •− is produced mainly at complex I and complex III due to transfer of electrons to molecular oxygen. Superoxide radical that is produced by complex I is released into the mitochondrial matrix, whereas complex III forms O2 •− both in the matrix and the inner mitochondrial space (9, 68). Another major source of superoxide is NADPH oxidases, a family of membrane-bound enzymes that catalyze controlled production of O2 •− by coupling NADPH-derived electrons to oxygen. Superoxide can be also produced in the cell through one-electron transfer reactions catalyzed by a number of enzymes, including monoamine oxidase, cyclooxygenases, lipoxygenases, and components of the cytochrome P450 system.

Superoxide radical is very unstable (half-life 10−6 s) and is rapidly dismutated to H2O2 and molecular oxygen by superoxide dismutases (SODs) (25). In mammalian cells, manganese superoxide dismutase (MnSOD) is present within mitochondrial matrix, whereas cytosolic and extracellular forms of SOD contain copper and zinc in their active site (CuZnSOD) (24). SOD accelerates the rate of spontaneous dismutation more than 1000-fold. If superoxide is not removed by SOD, it can react with iron–sulfur ([Fe–S]) clusters, releasing iron and compromising the functions of [Fe–S] cluster-containing proteins (34). Due to its high reactivity and inability to diffuse through membranes because of the negative charge, O2 •− is considered as a poor signaling molecule (12).
In contrast to O2
•−, H2O2 is less toxic and is better suited for signaling purposes, in part due to its longer half-life (cellular half-life
If not removed, H2O2 can be homolytically cleaved in the presence of redox-active Fe(II) iron (Fenton reaction) to form highly reactive hydroxyl radicals (HO•). Hydroxyl radicals are toxic and cause damage to different macromolecules, including nucleic acids, proteins, and lipids. They are also short-lived (half-life 10−9 s), which limits their use as signaling molecules (25).
Depending on biochemical properties of ROS signaling molecules, each of these species (O2 •−, H2O2, and HO•) has a distinct set of cellular targets. As discussed above, O2 •− targets mainly [Fe–S] cluster-containing proteins. One such example in Escherichia coli is a redox-sensitive transcription factor SoxR, which regulates transcription of genes involved in degradation of O2 •− (e.g., mitochondrial MnSOD) and regeneration of [Fe–S] clusters in proteins (ferredoxin:NADPH oxidoreductase) (33). HO• has a very broad reactivity and can interact with a large number of biological molecules.
The major targets of H2O2 are two sulfur-containing amino acids, cysteine (Cys) and methionine (Met), which are the most susceptible amino acids to oxidation by this compound (43). Oxidation of both free Met and Met residues by H2O2 forms methionine sulfoxide (MetSO). As this modification can alter the structure and function of target proteins, organisms evolved a family of enzymes, referred to as methionine sulfoxide reductases (Msr), which can repair oxidatively-damaged Met. However, H2O2 reacts with Met residues very slowly (rate constant of 10−2 M −1•s−1), which makes Met an unfavorable target for the H2O2-receptor function (34).
Although oxidation of free Cys by H2O2 is also slow (2–20 M −1•s−1, at neutral pH) (79), the reactivity of Cys residues in proteins can be significantly increased by the protein environment. High reactivity of Cys and its ability to be reversibly modified by H2O2 make it ideally suited for redox signaling. The following unique features of Cys are often employed by major regulatory protein targets of H2O2. First, Cys thiols can covalently interact with other thiol groups and form intra- and intermolecular disulfide bonds, a reaction promoted by H2O2. Disulfide bond formation is reversible as disulfides can be reduced by either thioredoxin (TRX) or glutaredoxin (GRX) systems. Second, Cys (along with His) is often employed in metal-binding sites in proteins to coordinate metals such as Zn. Upon oxidation of Cys residues in metal-containing redox-active proteins, the metal ion is released, resulting in conformational changes that can alter protein function. Finally, the side chain of reactive Cys can be directly oxidized by H2O2 to form cysteine sulfenic acid (R-SOH) (Fig. 2). The sulfenic acid form is unstable and is either further oxidized to cysteine sulfinic acid (R-SO2H) or can react with an adjacent Cys-SH group, leading to disulfide bond formation. Alternatively, R-SOH can be stabilized by formation of sulfenyl amide in some proteins (e.g., in protein tyrosine phosphatase PTP1B) (63). Reversible oxidation of Cys thiols is known to play an important role in redox regulation via the formation of sulfenic acid intermediates.

Specificity of ROS Signaling
ROS-mediated signaling has been shown to regulate a number of different cellular processes, ranging from activation of stress response pathways to regulation of normal growth and development. Specific examples include regulation of protein tyrosine phosphatases (63), PTEN (40), and hypoxia-inducible factor (HIF-1a) (4, 30, 45), circadian rhythms (18), as well as regulation of autophagy (64) and inflammatory responses (5, 52). Moreover, ROS have been implicated in activation of the ataxia-telangiectasia (ATM) protein kinase, which is involved in stem cell self-renewal, in response to DNA double-strand breaks (29), among other ROS signaling targets. Although a role for intracellular ROS in signal transduction has been established, it is still controversial how the specificity of redox signaling by ROS is achieved. ROS have distinct chemical properties, which distinguish them from other signaling molecules. In contrast to other signaling molecules, which regulate functions of proteins through non-covalent binding of ligands to their cognate receptors, ROS often operate through chemical interactions with specific amino acid residues (e.g., Cys) in target proteins that lead to covalent modifications (53). Given a large number of Cys residues in proteins and high reactivity of ROS in general, a wide range of promiscuous targets could be affected by these highly reactive molecules. Nevertheless, experimental evidence from studies of known redox signaling pathways shows a high level of specificity in ROS signaling.
The signaling function of ROS is generally thought to result from direct oxidation of ROS-specific receptor proteins involved in various cellular signaling pathways (19, 46) and that high specificity and selectivity of this interaction is achieved by different reactivity of various Cys residues (82). A second mode of regulation involves a cascade of thiol oxidoreductases that transfer oxidizing equivalents from H2O2 to target proteins. One example of such system is the regulation by thiol-dependent peroxidases, which are a group of peroxide scavenging enzymes that have evolved high reactivity toward H2O2. In addition to degrading peroxides, these proteins can also oxidize protein thiols and form disulfides in other substrates. This capability is employed by the Gpx3-Yap1 redox relay in Saccharomyces cerevisiae to regulate transcriptional response to oxidative stress. Gpx3 serves as a sensor of H2O2 (15) and can activate Yap1 transcription factor by forming multiple (up to three) intramolecular disulfide bonds, which promote accumulation of Yap1 in the nucleus and Yap1-dependent gene activation (55). An additional thiol peroxidase, Tsa1, has also been reported to sense H2O2 and activate Yap1-mediated transcriptional response to oxidative stress in S. cerevisiae by forming disulfides in Yap1 (54, 62, 69). Moreover, Schizosaccharomyces pombe peroxiredoxin Tpx1 has been shown to activate Pap1 transcription factor and Sty1 stress-activated protein kinase in a similar fashion (3, 76, 77). These observations suggest a possibility that H2O2 can specifically oxidize a narrow group of proteins that have high reactivity to H2O2 (e.g., thiol peroxidases), which in turn transfer the signal to specific transcription activators (and other proteins) by forming disulfide bonds in these targets. Therefore, uncoupling H2O2 sensing function from transcriptional regulators in such redox relays can dramatically increase specificity of H2O2 signaling. How widespread this mechanism is in living organisms remains unclear. Recently, a study of a yeast strain lacking all thiol peroxidases demonstrated that, at least in this organism, thiol peroxidases are major targets of H2O2 signaling and mediate genome-wide regulation of gene expression in response to H2O2 (21). Yeast cells lacking all 8 thiol peroxidases were unable to sense and transcriptionally respond to H2O2, whereas other oxidant/stress signaling pathways were not affected. These data suggested that thiol peroxidases have dual function in redox regulation: they (i) protect cells from oxidative stress by scavenging toxic H2O2 molecules, and (ii) specifically transmit oxidative signals to activate transcription factors and regulatory proteins in response to H2O2 (Fig. 3). This model also suggests that thiol peroxidases are the main regulators of H2O2 response in yeast. Although direct oxidation of signaling and regulatory proteins by H2O2 also likely exists, it plays a secondary role (21). Whether thiol peroxidases regulate redox signaling by sensing H2O2 and activating cellular target proteins in other organisms, including mammals, awaits further investigation. In line with a possible involvement of thiol peroxidase-mediated signaling in regulation of metazoan aging are recent reports that show a dual role of Prxs in modulating stress resistance and longevity in C. elegans (56) as well as in stimulating survival of murine macrophages exposed to oxidized low density lipoprotein (oxLDL) (11). In addition to protecting cells from oxidative stress by degrading H2O2, in both studies Prxs have been shown to activate the p38 MAPK stress-related signaling pathway. Prx activity has been also implicated in regulating PTEN signaling by protecting this protein from oxidation-induced inactivation (8). However, it should be noted that it is often difficult to differentiate between the contribution that the signaling and antioxidant effects of thiol peroxidases make to their role in longevity.
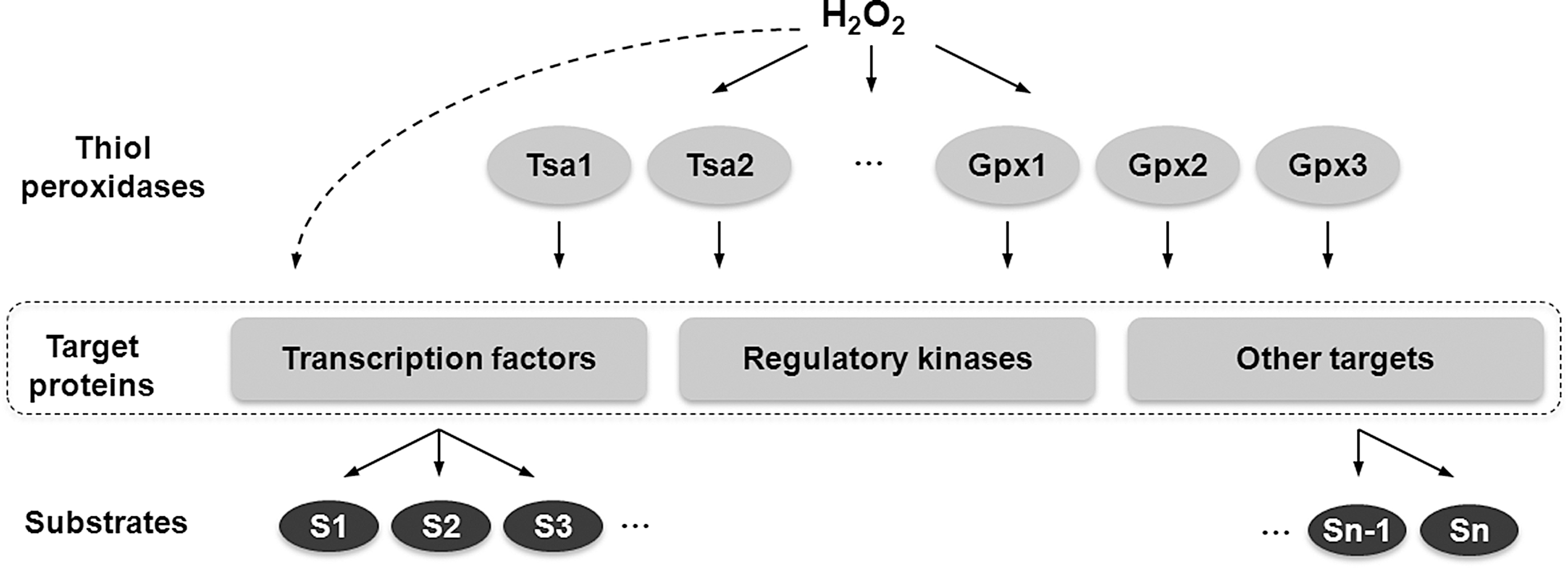
Another possible mechanism by which H2O2 specificity may be achieved is by controlling the concentration of H2O2 and co-localization of the target protein in vicinity of the source of H2O2 production. Local H2O2 concentration can be modulated by peroxiredoxins (Prxs), which are themselves subject to H2O2-mediated inactivation. In eukaryotic cells, catalytic Cys in Prx proteins can be overoxidized by H2O2 to a sulfinic acid (-SO2H) form, inactivating Prx (60, 82). This inhibition is reversible by ATP-dependent reduction of the sulfinic acid by the tightly regulated enzyme sulfiredoxin (Srx) (2, 80). According to the “floodgate hypothesis”, active Prxs normally keep H2O2 concentration low, thereby preventing cellular damage (82). However, during signal transduction, rapid production of H2O2 results in high local H2O2 concentrations, which could inactivate Prxs. Prx inactivation leads to further accumulation of H2O2 for signaling purposes, which could oxidize downstream target proteins in the specific region of the cell (31). This model is consistent with the recent observation of a transient tyrosine phosphorylation on membrane-associated peroxiredoxin 1 (Prx1) upon stimulation of cells by a growth factor, which leads to inactivation of Prx1 (81). This transient phosphorylation only occurs on membrane-associated, but not cytosolic Prx1 leading to localized accumulation of H2O2 needed for signaling. In addition, recent studies suggest that H2O2 not only can diffuse across plasma membrane, but may also be transported through membrane aquaporin channels (1, 48). Therefore, H2O2 channeling to specific cellular sites may provide additional mechanisms, by which high specificity of ROS signaling can be achieved. Finally, it is also possible that overall specificity of H2O2 signaling may result from a combination of the different mechanisms described above and other unidentified factors that may modulate signaling functions of ROS.
Genome-Wide Identification of Reactive Cys Residues
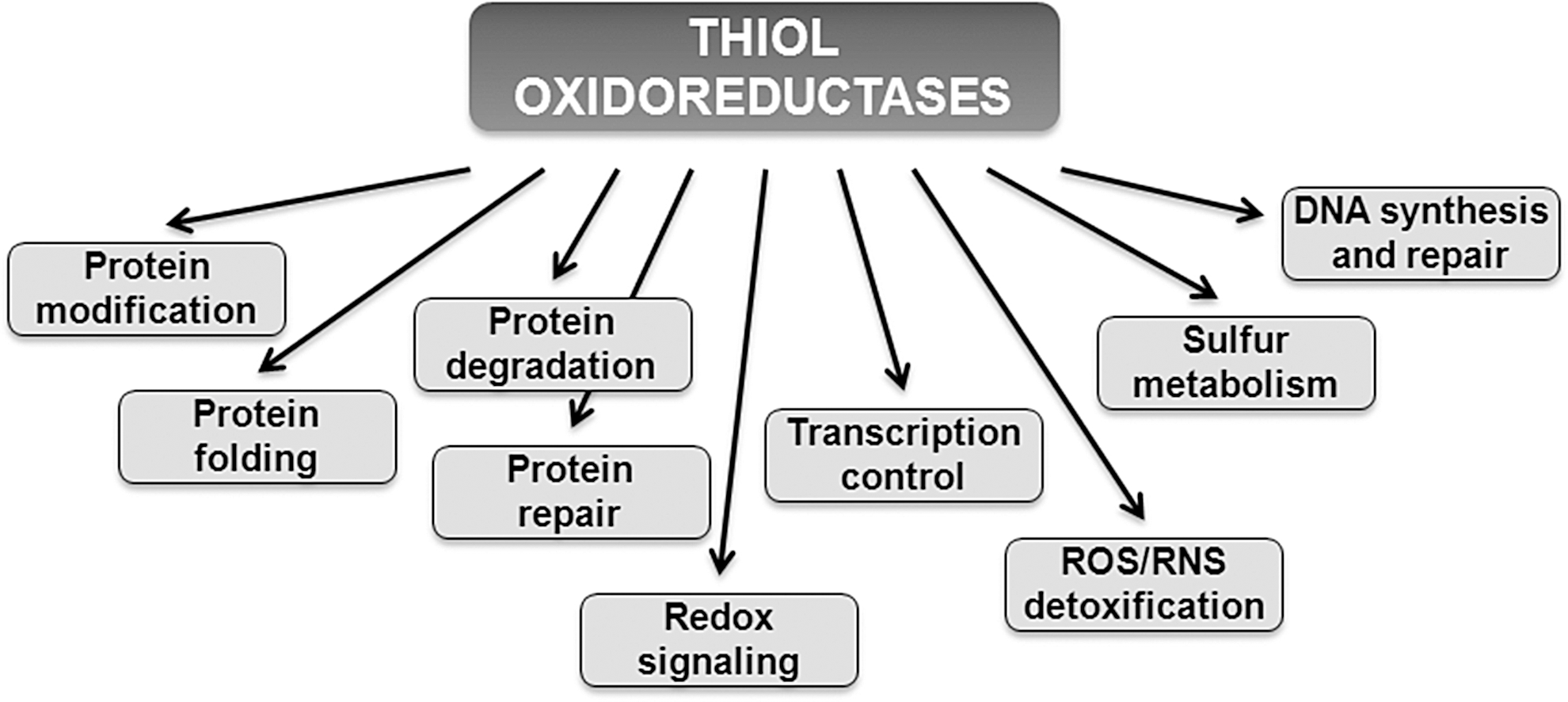
Reactive Cys residues in redox signaling proteins serve as major regulatory targets of H2O2. Many of these regulatory proteins belong to a class of enzymes known as thiol oxidoreductases, which control a variety of cellular functions (Fig. 4). These proteins utilize catalytic Cys residues for reduction or oxidation of their substrates and are ideally suited for the reaction with H2O2 (22). Although many thiol oxidoreductases have been characterized, identification of reactive Cys in proteins has been limited to experimental approaches that utilized individual proteins. Recent advances in the field were made by employing computational approaches (20, 23), which were used to identify reactive Cys residues on a genome-wide level (Fig. 5), as well as large-scale proteomic (mass spectrometry-based) approaches, including two-dimensional electrophoresis gel (2DE)-based fluorescent methods (10), OxICAT (42), and isoTOP-ABPP (isotopic Tandem Orthogonal Proteolysis—Activity-Based Protein Profiling) (78), which were used to identify Cys-containing proteins oxidized in vivo. In particular, computational studies revealed that thiol oxidoreductases are present in all living organisms and generally account for around 0.5%–1% of all proteins encoded in the genome. The abundance of thiol oxidoreductases in cells demonstrates a previously not appreciated complexity of redox regulation and suggests that thiol-based redox signaling is a widespread process across different domains of life. Rapid accumulation of publicly available genome sequences combined with comparative genomics analyses should allow detection of new thiol oxidoreductases and identification of redox-active Cys in these proteins. It would be also important to further characterize the function of thiol oxidoreductases and their contribution to redox regulation of cellular processes, including aging.

Role of Redox Signaling in Aging
The “free radical theory” of aging postulates that aging results from accumulation of oxidative damage to macromolecules (32). However, organisms evolved specific stress response pathways to protect cells from oxidative stress. These pathways sense ROS and activate transcription factors that regulate expression of genes involved in oxidative stress defence. Although the sources of ROS and the mechanisms of ROS-inducible oxidative stress-protective responses are well defined, little is known about the role of redox signaling in aging.
Whereas young organisms are capable of effectively responding to environmental and internal stresses, such as hypoxia, altered metabolism, and oxidative stress, there is compelling evidence that these regulatory processes are altered during aging. It is thought that the inability of aged cells to respond effectively to cellular damage leads to decreased stress resistance and accumulation of “senescence factors,” such as DNA mutations, by-products of metabolism, oxidatively damaged proteins, and protein aggregates (13, 59, 66). Consistent with this view, genetic mutations that extend lifespan are associated with increased cellular stress resistance in variety of species, including yeast, worms, fruit flies, and mice (44).
Role of ROS-mediated signaling in regulating yeast stress resistance and lifespan
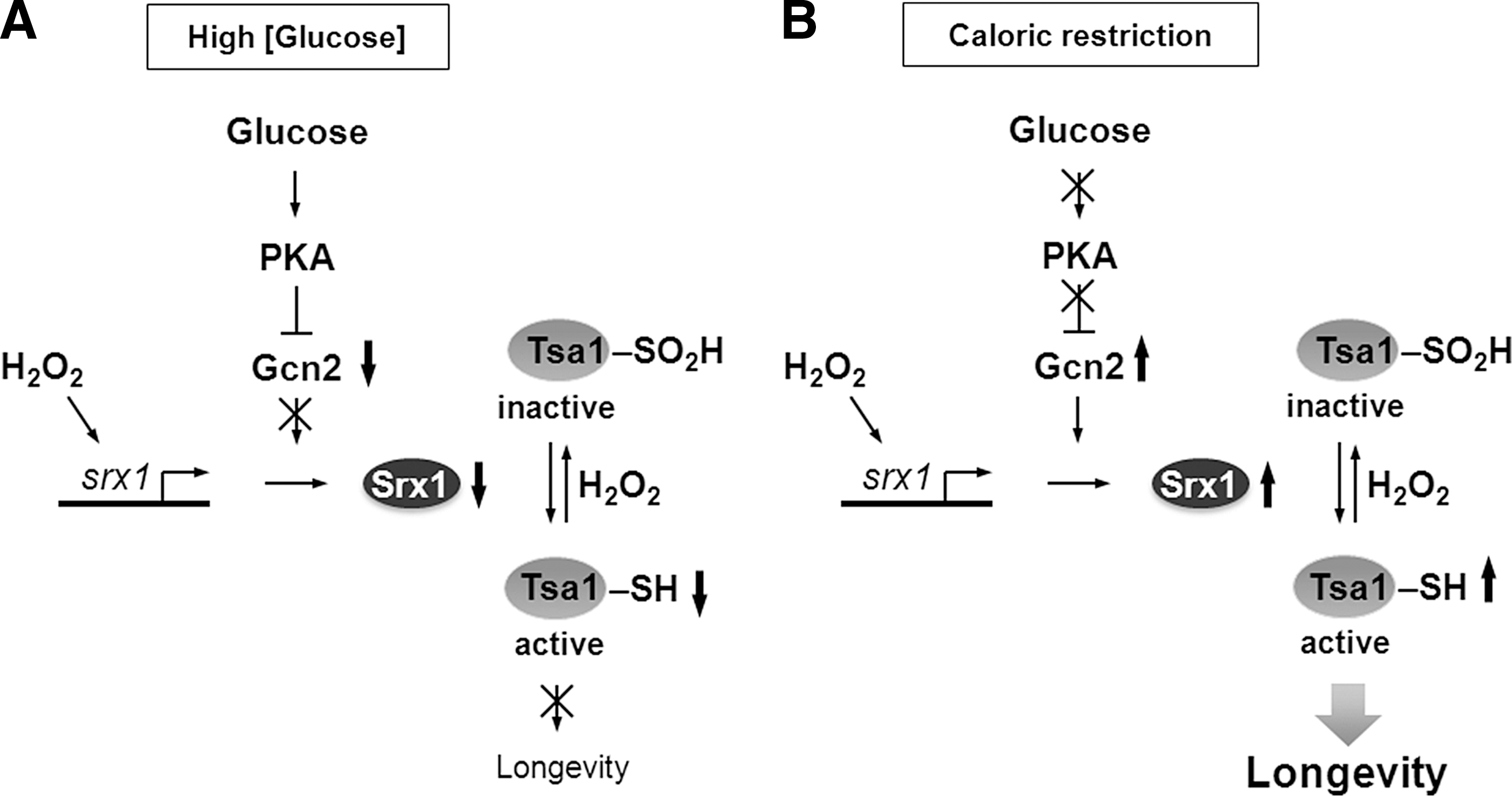
Emerging evidence from studies in model organisms revealed that ROS-mediated signaling plays an important and previously unappreciated role in regulating cellular stress resistance and organism's lifespan. One of the examples that illustrate importance of redox signaling in aging comes from the study of lifespan extension by caloric restriction (CR) in S. cerevisiae. Peroxiredoxin Tsa1 and its partner sulfiredoxin Srx1 have been shown to mediate extension of replicative lifespan and increased H2O2 resistance elicited by CR in yeast (49). During aging, Tsa1 is subject to H2O2-mediated inactivation by hyperoxidation, and CR enhances expression of Srx1, which is required for reactivation of Tsa1 contributing to increased resistance to oxidative stress and longevity (Fig. 6). These findings suggest that stress resistance associated with CR depends on ROS-mediated signaling pathways and that, during aging, hyperoxidation of Tsa1 can inhibit the ability of cells to activate its downstream regulatory targets and respond to oxidative stress. Considering that CR can extend lifespan in a wide variety of species and that peroxiredoxins (Prxs) have been conserved during evolution, it is possible that Prxs can also modulate longevity and contribute to the lifespan extension effect of CR in higher eukaryotes. Consistent with this possibility, a neuronal Prx was identified in D. melanogaster as a downstream target of the insulin signaling-regulated transcription factor FOXO, which is involved in the increased resistance to H2O2 and the lifespan extension effect of CR (41). It should be noted, however, that some organisms (e.g., C. elegans), lack any Srx gene or Srx-like enzyme activity and do not show an increase in Prx hyperoxidation with age (70). Thus, the induction of Srx by CR observed in yeast is unlikely to mediate CR‐induced increases in lifespan in these organisms. A detailed mechanism by which Prxs may contribute to longevity and the identity of transcription factors and regulatory proteins responsible for the Prx-dependent lifespan extension during CR remains to be established. Moreover, the exact signaling role of Prxs in the normal aging process, as well as whether Prx inactivation can be beneficial for signaling (see the “floodgate hypothesis” above) remain puzzling (31). In a recent report, Day et al. provided evidence that in S. pombe Prx inactivation by high H2O2 may enhance cell survival during oxidative stress by preserving the pool of reduced Trx (14). S. pombe peroxiredoxin Tpx1 and transcription factor Pap1 form a redox relay that regulates the transcriptional response to H2O2. In the absence of stress, Tpx1 is maintained in cells in the reduced state by the NADPH-dependent Trx system, with Trx1 serving as a primary reductant. During exposure to H2O2, Tpx1 activates Pap1 by forming disulfide bonds; however, high levels of H2O2 lead to overoxidation and inhibition of Tpx1 activity. Surprisingly, the authors found that hyperoxidation of Tpx1 increases the pool of reduced Trx1 and promotes survival of cells exposed to toxic levels of H2O2. They hypothesized that the beneficial effect of Tpx1 inactivation may result from an increased level of reduced Trx1, which can be used for repair of oxidatively damaged proteins by methionine sulfoxide reductases (Msr). Although this study proves that sensitivity of Tpx1 to hyperoxidation may provide a benefit during acute exposure to H2O2, it does not offer a clear explanation why sensitivity of Tpx1 to hyperoxidation has evolved (36) and whether preserving the pool of reduced Trx1 by Tpx1 inactivation may affect the lifespan of yeast cells.
In addition, ROS signaling has been implicated in extending chronological lifespan caused by inactivation of the nutrient-responsive target of rapamycin (TOR) kinase in S. cerevisiae (57), which has been shown to be involved in the lifespan extension by CR. It was hypothesized that these effects can be attributed to “mitochondrial hormesis” or “mitohormesis” (61, 65), a process whereby elevated mitochondrial ROS generation leads to constitutive activation of antioxidant defense mechanisms and expression of survival genes. However, specific signaling pathways and downstream effectors that are activated by CR-induced ROS production remain obscure. More recently, both low-glucose media and reduced signaling of the nutrient-sensitive TOR and protein kinase A (PKA) pathways have been shown to extend the chronological lifespan of S. pombe through increased ROS production and activation of the stress-dependent Sty1 mitogen-activated protein (MAP) kinase (87). Sty1 kinase is involved in upregulation of stress-related genes and can be activated by multiple stresses, including H2O2. During stress, Sty1 is phosphorylated by Wis1 and translocates to the nucleus, where it activates its downstream transcription factor Atf1. It should be noted that among other stress-related genes, Atf1 regulates expression of Srx1 that is necessary for reactivation of Tpx1 in S. pombe. In turn, Tpx1 may also regulate Sty1 signaling by enhancing H2O2-induced Sty1 phosphorylation (76). Whether Tpx1 and Srx1 can mediate the CR-induced lifespan extension in S. pombe remains unknown.
Links between ROS-induced damage/ROS-signaling responses and aging in higher eukaryotes
Although a role for ROS in aging in higher eukaryotes remains controversial (50, 74), recent studies manipulating the expression of SOD and catalase, antioxidant genes involved in control of ROS levels, in nematodes provide additional support for the importance of ROS-mediated signaling in regulating aging. SOD and catalase are the major antioxidant enzymes that detoxify O2 •− and H2O2, respectively. Whereas yeast and fruit flies have two SOD genes encoding cytoplasmic and mitochondrial enzymes, there are five SOD isozymes in C. elegans (two mitochondrial SODs, encoded by sod-2 and sod-3; two cytosolic SODs, encoded by sod-1 and sod-5; and one extracellular SOD, encoded by sod-4). There are also three different catalases, encoded by ctl-1, ctl-2, and ctl-3 genes. Several groups examined the consequences of eliminating expression of these enzymes or their overexpression in worms (reviewed in Refs. 26 and 74). According to the free radical theory of aging, which postulates that aging results from accumulation of molecular damage caused by ROS, a decreased lifespan would be expected, when ROS levels are elevated by deletion of genes such as SOD or catalase. On the other hand, overexpression of these ROS-detoxifying genes would be expected to extend lifespan. Indeed, previous studies in yeast and fruit flies have demonstrated that deletion of either mitochondrial or cytosolic SOD genes shortens lifespan, which could be interpreted as consistent with this theory. However, deletion of individual sod genes had little effect on lifespan in C. elegans, and in some cases (e.g., deletion of sod-2) it could even extend lifespan (16, 73). Furthermore, extended lifespan in sod-2 mutant worms was associated with increased levels of oxidative damage, suggesting that ROS toxicity does not play a major role in lifespan regulation in these animals (73). One possible explanation of why deletion of individual sod genes failed to shorten lifespan is compensation by additional sod genes. However, a recent report from the Hekimi laboratory demonstrates that worms lacking all five sod genes are viable and have normal lifespans, despite significantly increased sensitivity to multiple stresses (75). These observations indicate that oxidative damage caused by superoxide radical does not contribute to worm aging in all experimental conditions. It appears that SOD activity is dispensable and is required only when animals are exposed to acute stresses.
Consistent with these findings, lifespan extension from overexpression of sod-1, the major cytosolic Cu/ZnSOD, and sod-2, the major mitochondrial MnSOD, in C. elegans was not caused by reduced oxidative damage (6). Instead, SOD overexpression was associated with elevated levels of H2O2 and increased protein oxidation, suggesting a possible role of H2O2-mediated signaling in regulating lifespan in worms.
Further support for the role of ROS-mediated signaling in aging is provided by experiments in which worms were treated with increasing concentrations of paraquat, a superoxide generator, or vitamin C, which has a superoxide scavenging activity (75). This study demonstrated that increased superoxide levels, due to addition of low levels of paraquat, resulted in lifespan extension in wild-type worms, whereas at high doses of paraquat the lifespan was decreased. In contrast, treatment of wild-type worms with antioxidants had little effect on their lifespan. This biphasic pattern suggested a model in which C. elegans longevity is dependent on a balance between the prosurvival ROS-mediated signaling and ROS toxicity (Fig. 7). At low levels of superoxide (caused by a decrease in mitochondrial superoxide detoxification through the deletion of sod-2 or produced by the addition of low levels of paraquat), the prosurvival effect of ROS signaling on lifespan is much stronger than the ROS toxic effect leading to lifespan extension. But, at high concentrations, superoxide leads to a dose-dependent decrease in lifespan due to its toxicity. Consistent with this model, treatment of sod quintuple mutant with antioxidants increased lifespan by 15%–20%, whereas the addition of low levels of paraquat decreased lifespan, presumably because the basal superoxide level in this mutant exceeds optimum concentration required for ROS signaling (75). Together, these observations suggest that both lack of antioxidant genes and their overexpression can lead to complex effects due to a dual function of ROS in redox control. Thus, researchers have to be cautious when interpreting such studies (51).

Lifespan extending effects of ROS in long-lived C. elegans, in which mitochondrial function is genetically disrupted
Impaired mitochondrial function has been implicated in the aging process in a number of different ways. Although mutations of some genes that encode subunits of mitochondrial electron transport chain, for example, gas-1 (37) and mev-1 (35), have been found to accelerate aging in worms due to an increase in ROS-induced oxidative stress, some mutations that affect mitochondrial function lead to lifespan extension (e.g., isp-1 and nuo-6) (85). It has been proposed that both isp-1 and nuo-6 increase C. elegans lifespan by lowering levels of ROS and oxidative damage to proteins. However, recent analysis of superoxide generation in these mutants revealed that isp-1 and nuo-6 worms have in fact increased rather than decreased ROS levels, and that superoxide is necessary and sufficient for the lifespan extension (84). These observations support a model wherein superoxide produced by mitochondria can act as a signaling molecule triggering specific, yet unidentified, signaling pathways that induce protective and repair mechanisms. The nature of the signaling pathways that may contribute to the ROS signaling-mediated lifespan extension remains largely unknown, and considering the large number of potential ROS targets (see above) it would not be surprising if multiple signaling pathways are involved. One potential mechanism that has been uncovered recently links mitochondrial ROS production to the induction of the mitochondrial unfolded protein response (UPRmt) (17). In this study, inhibition of mitochondrial activity in the intestine or neuronal cells, by tissue specific knockdown of a cytochrome C oxidase 1 subunit (cco-1), increased ROS levels and extended C. elegans lifespan. The authors also demonstrated that inhibition of mitochondrial function can promote longevity in a cell-nonautonomous manner, as knockdown of mitochondrial electron transport chain components in neurons was able to induce the UPRmt signaling pathway in the intestine and extend lifespan. Although the chemical nature of the signal that couples mitochondrial inhibition in neuronal cells to activation of the UPRmt in the intestine remains unknown, it is a possibility that ROS produced in mitochondria may act as a diffusible signaling molecule to activate the UPRmt-mediated transcriptional response. In this regard, it would be important to test whether tissue-specific upregulation of ROS levels in worms by manipulating the expression of antioxidant genes may also extend lifespan in a cell-nonautonomous manner.
Similar to yeast, glucose restriction extends lifespan in C. elegans through increased mitochondrial ROS generation and ROS-dependent induction of long-term stress resistance (65). Moreover, treatment of calorically restricted nematodes with different antioxidants prevented both stress resistance and the lifespan extension effect, suggesting that induction of stress resistance during CR is dependent on generation of ROS.
Together, the above examples in yeast and nematodes revealed that, while ROS can contribute to aging by damaging macromolecular structures, the generation of ROS at low levels can promote longevity by activating redox-regulated signaling pathways. It remains an open question whether increased ROS signaling can promote lifespan extension in mammals. In multicellular organisms, ROS-mediated signaling has been adapted for purposes other than oxidative stress defence (i.e., transcriptional activation of antioxidant response genes). Such nonstress-related ROS-mediated signaling is unique to eukaryotes and appears to regulate normal growth and development. The naked mole rat (Heterocephalus glaber), an exceptionally long-lived rodent (maximum lifespan exceeds 30 years), provides an example of how elevated ROS signaling may contribute to the increased longevity in mammals. In addition to delayed aging, naked mole rats show increased oxidative stress and ROS production at young age, yet their proteome is resistant to oxidative damage (58). Whether elevated ROS production contributes to the exceptional longevity of this organism or is just a correlative event remains unknown. Further elucidation of mechanisms underlying unique stress resistance and longevity of the naked mole rat should improve our knowledge of the role played by ROS-mediated signaling in aging, and availability of this organism's genome, which has been recently sequenced, should accelerate these studies (38).
Role of ROS-inducible pathways in regulating longevity in mammals
Several signaling pathways that have been implicated in regulating lifespan in mammals are subject to regulation by ROS, but the specific upstream ROS sensor proteins remain largely unknown. One of the pathways activated by ROS that might link ROS-mediated signaling with the aging process is the Keap1-Nrf2 pathway. Nrf2 transcription factor regulates the expression of phase 2 detoxification and antioxidant genes. In turn, activation of Nrf2 is repressed by an inhibitor protein named Keap1, which targets Nrf2 for proteasomal degradation through ubiquitination of its specific Lys residues (39). However, H2O2 and other oxidative stress and environmental cues inactivate Keap1 so that it can no longer promote Nrf2 degradation, leading to its nuclear accumulation. The ROS-sensing mechanism involves modification of specific cysteine residues in Keap1 and conformational changes that inactivate Keap1 activity (86). Recently, an Nrf2 homolog in worms, SKN-1, has been shown to mediate the lifespan extension effect of reduced insulin/IGF-1-like signaling in worms (72) linking ROS signaling with the aging process.
Future Directions
ROS, which have been long considered as toxic compounds causing oxidative stress, are now recognized as unique signaling messengers that are produced in a regulated manner and control a wide range of cellular processes, including but not limited to, cell proliferation, circadian rhythms, immune response, and aging. Intracellular levels of ROS are sensed by various cellular transducers, which relay the signal to activate downstream targets and signaling pathways. Dysregulation of ROS homeostasis and ROS signaling have been linked to the development of various age-related diseases, including diabetes (27), cancer (47), and neurodegeneration (7), but their exact roles in the aging process still remain puzzling.
In recent years, many of the mechanisms regulating the production of ROS and basic principles of ROS-based redox signaling have been identified. Despite this progress, studies on the role of ROS-mediated signaling in aging lag behind, in part due to insufficient information about the specific regulatory targets of ROS. Large-scale identification of reactive Cys residues and studies on thiol oxidoreductases, enzymes that utilize catalytic redox-active Cys residues, found that an increasing number of proteins are subject to redox regulation and revealed complexity of redox-mediated signaling pathways. It would be important to identify functions of these proteins under both physiological and pathophysiological conditions. Moreover, further characterization of these pathways and mechanisms that coordinate redox signaling may provide better understanding on how dysregulation of ROS-mediated redox signaling contributes to aging and allow development of new strategies to delay the aging process in humans.
Footnotes
Acknowledgments
This work was supported by National Institutes of Health grants AG021518 and AG038004 (to V.N.G.), and AG040191 (to V.M.L.). This research was conducted while V.M.L. was an Ellison Medical Foundation/AFAR Postdoctoral Fellow.
Author Disclosure Statement
No competing financial interests exist.