
Abstract
Introduction
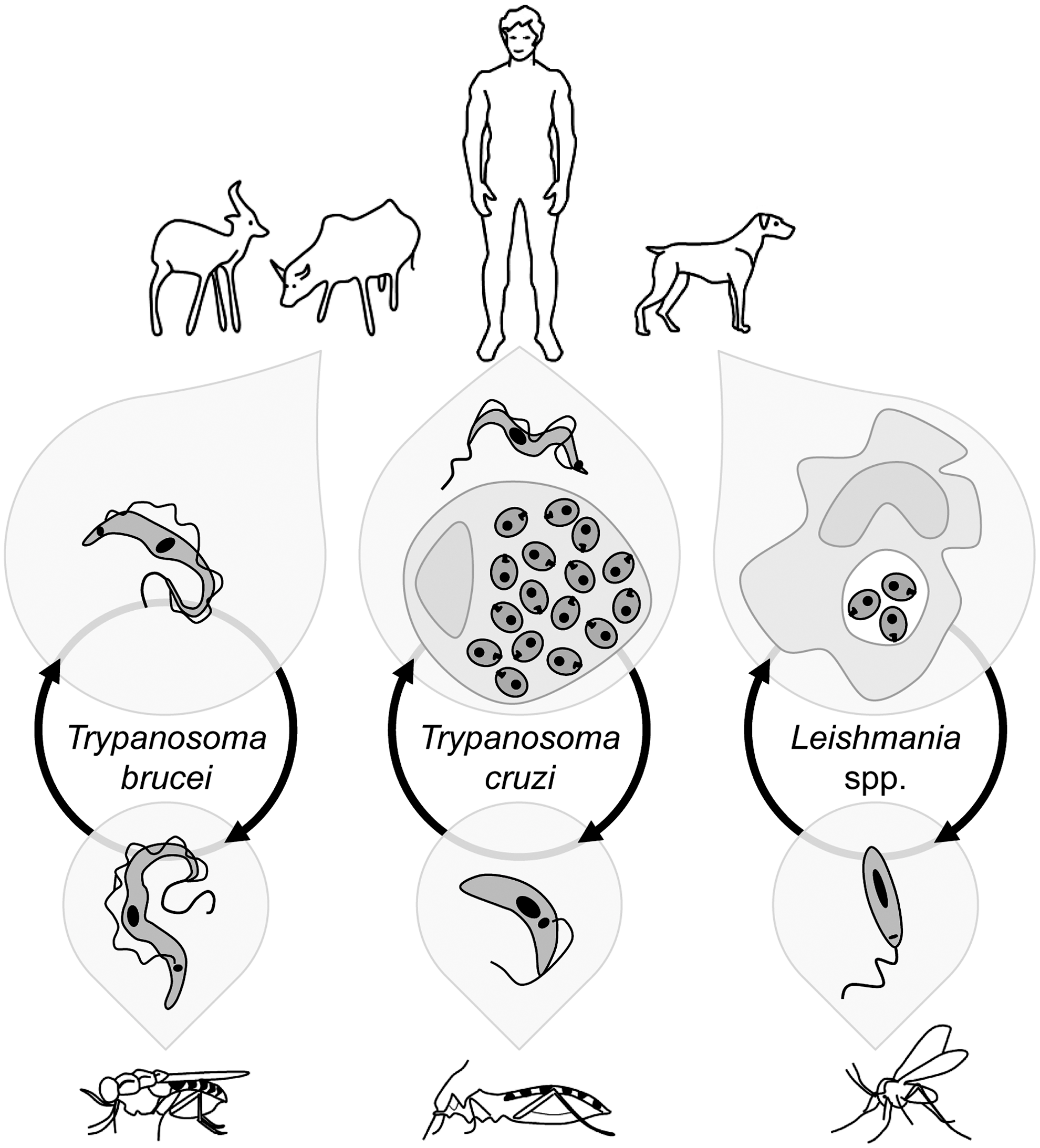
Trypanosomatids encompass a vast group of organisms included in the order Kinetoplastida, many of which are parasites of humans, animals, and plants. For simplicity, this review is restricted to the medically relevant Leishmania spp., the agents of human and canine leishmaniasis, to the Trypanosoma brucei complex, which causes sleeping sickness in humans and Nagana in cattle, and to Trypanosoma cruzi, responsible for Chagas' disease. The main life cycle stages of these parasites are depicted in Figure 1.
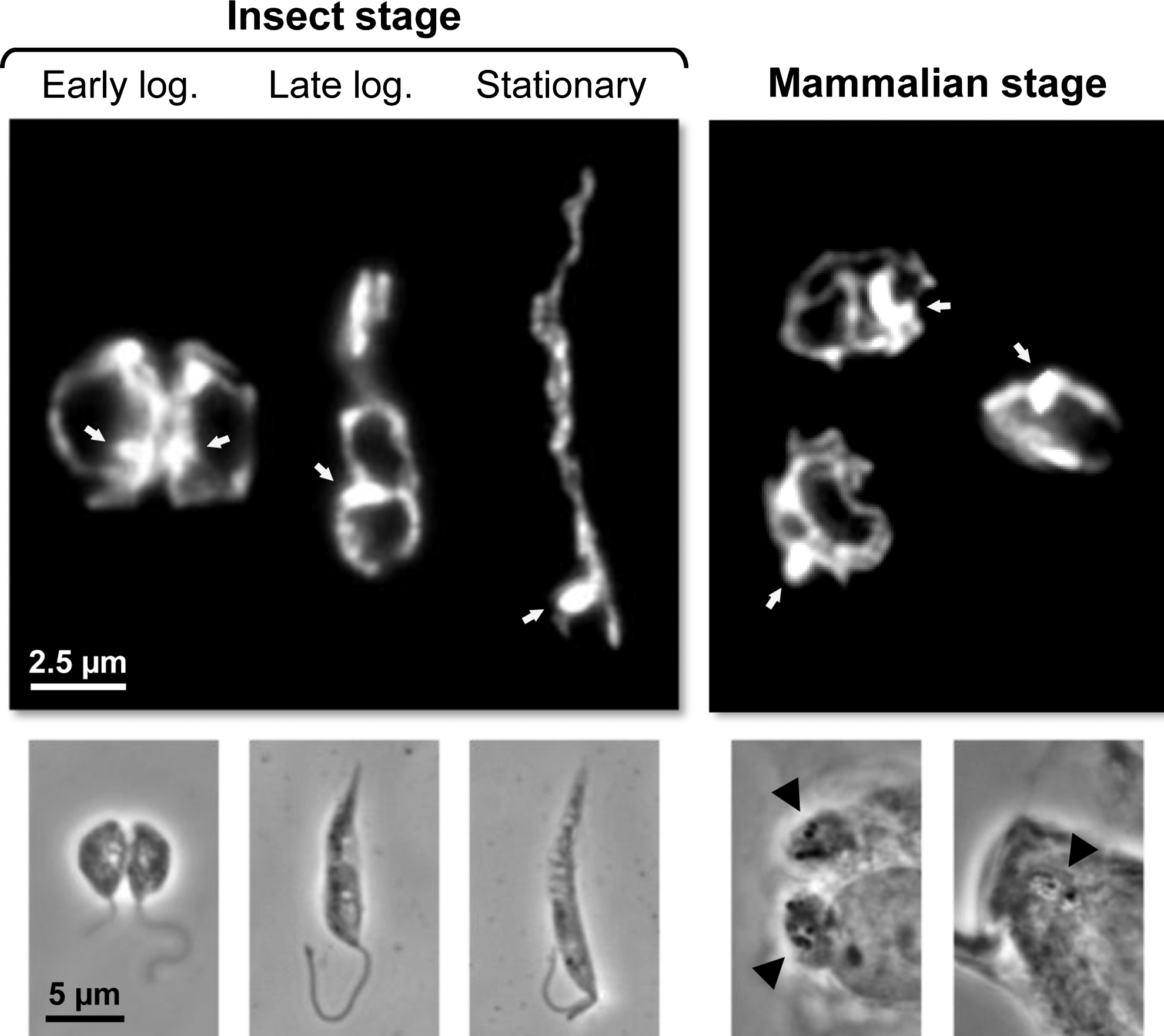
Several pieces of evidence, including recent data on the characterization of the mitochondria protein-import machinery (75), indicate that trypanosomatids are among the earliest diverging eukaryotes to have mitochondria (43). In these organisms, this organelle is a single structure (68) with a tubular form which extends throughout the cell. It contains an outer membrane, a dense matrix, and an inner membrane that can fold into thin and irregularly distributed cristae whose number varies widely (25). In fact, one characteristic of trypanosomatid mitochondria is their plasticity, with the exact morphology, volume, and activity depending on factors such as the cell cycle phase, the stage, and the species, or the nutritional sources available (25). Figure 2 illustrates the alterations in the morphology of Leishmania infantum mitochondria along parasite development. The variability in trypanosomatid mitochondria is even more striking in T. brucei. While in procyclic parasites thriving at the expenses of amino acids such as proline, mitochondria are large and highly branched; in bloodstream forms, which obtain their energy solely from glycolysis, they are small and repressed. Within the mitochondrial matrix and located perpendicular to the axis of the flagellum, lies the kinetoplast, a disk-shaped structure that contains the mitochondrial DNA (the kinetoplast DNA or kDNA) which in trypanosomatids is composed of a dense network of two types of circular molecules, maxicircles and minicircles. The former encode some of the mitochondrial proteins while the second specify unique RNA molecules required for RNA editing, a distinctive process occurring within trypanosomatid mitochondria which converts aberrant RNA sequences into functional messages (7). Despite singularities such as the ones mentioned above, many of the physiologic processes taking place in mitochondria of eukaryotes are also present in trypanosomatids. Still, some of these may exhibit unique features as is the case of the mechanism for cytochrome c (cyt c) biogenesis (3).
ROS Formation Within Trypanosomatid Mitochondria
Physiologic generation of ROS is an unquestionable phenomenon in aerobic organisms, the mitochondrion being an important site for its occurrence. Although technical limitations have up to now precluded the accurate determination of the levels generated in vivo, studies with isolated mitochondria estimate that monoelectronic reduction of molecular oxygen (O2) to superoxide anion (O2 •−) accounts for up to 0.1% of the total O2 consumed by resting cells (37). In trypanosomatids, production of ROS within mitochondria is also documented. Evidence collected from a number of studies have established that generation of these molecules can be triggered by many drug compounds (14, 38), including some used in clinics as is the case of miltefosine (52), as well as by classic respiratory chain inhibitors (33, 56, 77). An important question, however, is whether ROS formation does occur along the parasites life cycle. In this regard, studies reporting production of mitochondrial ROS in response to physiologic stimuli, such as serum complement (72) and heat shock (4), and linking the formation of these molecules to induction of apoptotic-like death, are particularly important as they suggest that ROS generation is a phenomenon that can occur in vivo and have functional significance for trypanosomatids.
Although there are solid data associating ROS with trypanosomatid mitochondria, the exact site for their production has not been as thoroughly addressed as in other systems. Of relevance, the isolation of the single mitochondrion of trypanosomatids in an intact form is difficult. Such analyses are, thus, usually carried out either using mitochondrial enriched fractions (vesicles) displaying membrane potential or, more frequently, whole parasites selectively permeabilized with digitonin at concentrations that preserve the integrity of the organelle (85).
In most eukaryotes, the respiratory chain is the main site for ROS production within mitochondria. During transference of reducing equivalents along the several intermediates of the chain, some electrons may escape, allowing for the monovalent reduction of molecular oxygen to superoxide anion (O2 •−). This radical ion is the primary ROS formed in cells and the precursor for hydrogen peroxide (H2O2) and other species (48, 51). With the possible exception of T. brucei bloodstream forms, the respiratory chain might as well constitute a source of reactive oxygen species to trypanosomatids. In fact, in spite of differences relative to other eukaryotes, the metabolism of all these organisms also entails electron flow along the chain (11, 59, 62, 83).
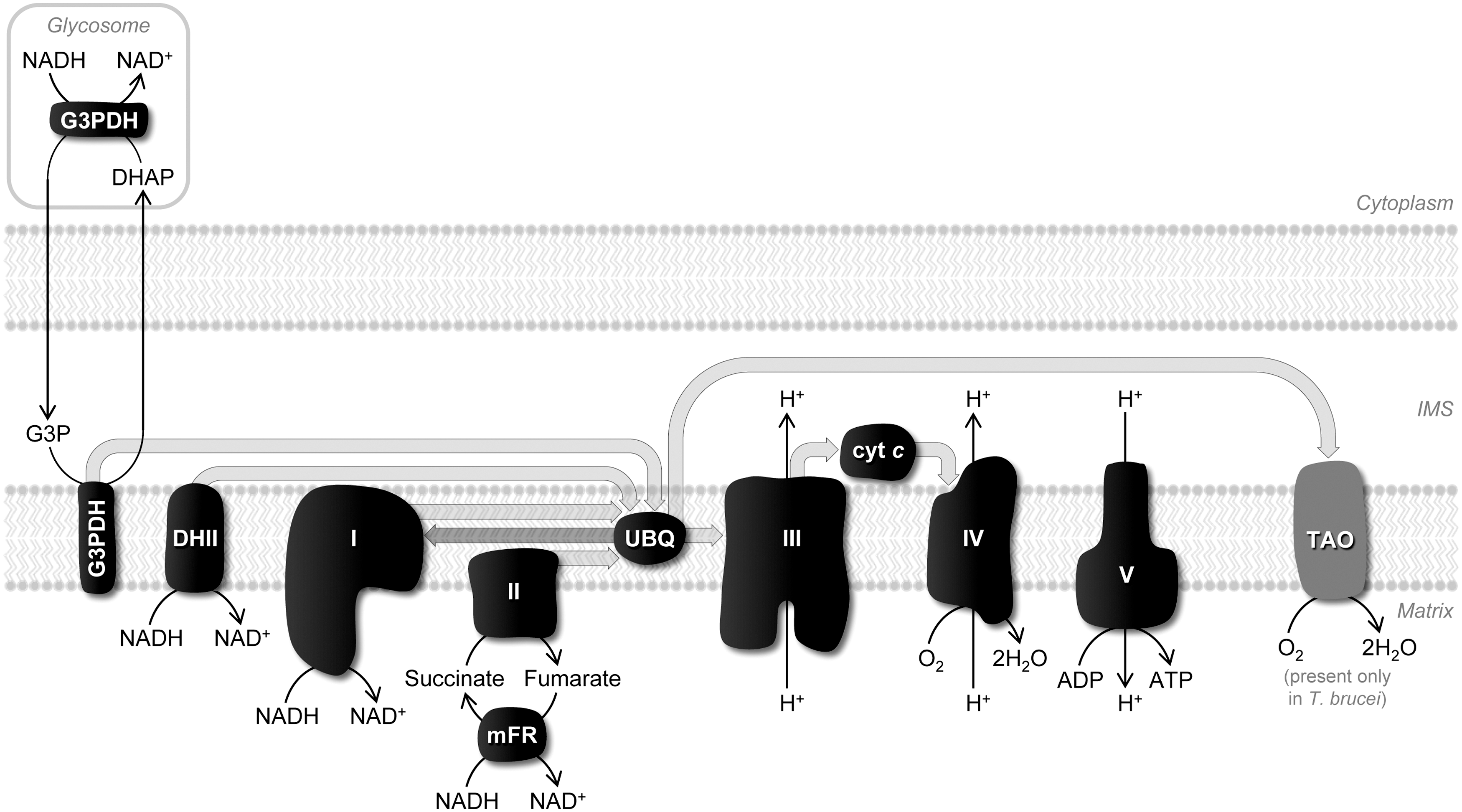
The main features of the respiratory chain of trypanosomatids are depicted in Figure 3. Although there are species and stage differences in the chain, in general terms, electrons from NADH and succinate enter the chain at different points via the mobile carriers ubiquinone (UBQ) and cyt c and are transferred to a terminal oxidase (complex IV). The latter uses this reducing power to produce water from molecular oxygen. Electron transfer through complexes III and IV is coupled to proton translocation promoting the formation of an electrochemical gradient between both sides of the inner mitochondrial membrane that sustains ATP synthesis.
Complex I
In a typical eukaryote, one of the main sites for ROS generation within the respiratory chain is complex I, which releases O2 •− on the matrix side (48). In trypanosomatids, this complex shows some singularities, including the absence of proton extrusion activity, but it appears to conserve all subunits containing the redox centers required for UBQ reduction (1, 63, 86). Nevertheless, it is relatively consensual that in all these organisms complex I displays low NADH dehydrogenase activity (1, 13, 77), which could explain why O2 •− originating from this complex was not detected neither in T. brucei procyclics (33, 86) nor in T. cruzi epimastigotes (13, 38). Superoxide formation from this locus was, however, reported in Leishmania donovani promastigotes upon stimulation with 100 μM rotenone (56).
Type II NADH dehydrogenase
In addition to complex I, a second inner membrane NADH dehydrogenase (DHII), with flavin as cofactor and also devoid of proton translocation activity, was characterized in T. brucei (35). Owing to the presence of orthologous sequences in the genomes of T. cruzi and Leishmania spp. (Tc00.1047053508717.20 and LinJ.36.5620;
Mitochondrial glycerol 3-phosphate dehydrogenase
Another dehydrogenase present in the inner mitochondrial membrane of all trypanosomatids, but particularly important in bloodstream forms of T. brucei, is glycerol-3-phosphate dehydrogenase (G3PDH) (64). This FAD-dependent enzyme oxidizes glycolysis-derived glycerol-3-phosphate (G3P) forming dihydroxyacetone phosphate (DHAP), as part of a shuttle between the glycosome (the organelle where glycolysis takes place) and the mitochondrion that helps keeping redox balance within the glycosome (Fig. 3). Mitochondrial G3PDH also feeds electrons in the respiratory chain. Although not recognized as a source of ROS in trypanosomatids, analogous enzymes in other organisms generate O2 •− to the intermembrane space (24).
Complex II
Apart from NADH, electrons can be supplied to the electron transport chain from succinate to UBQ via complex II, or succinate dehydrogenase. In other eukaryotes, complex II is only seldom referred to as producing O2 •− (90), perhaps because structural features of this complex limit accessibility of O2 to FAD (48). ROS production from trypanosomatid complex II was reported in T. cruzi epimastigotes (38, 80) and in L. donovani promastigotes (56) stimulated with the complex II inhibitor thenoyltrifluoroacetone (TTFA).
Reverse electron transfer
Electrons derived from succinate are also often referred to give rise to O2 •− through a different mechanism, reverse electron transfer (Fig. 3). In this case, reducing equivalents supplied via complex II to UBQ are transported back to complex I (and in trypanosomatids perhaps to type II NADH dehydrogenase) inducing ROS formation (48, 58). Reverse electron transfer, which is stimulated by a high inner membrane potential and a reduced UBQ pool, was credited as originating O2 •− in T. brucei (33) and in T. cruzi (13), but it is debatable whether this mechanism occurs in vivo.
Complex III
Within the respiratory chain of different organisms, complex III (ubiquinol:cytochrome c oxidoreductase) is usually a major source of ROS. Interestingly, Fang and Beattie (33) did not rank this site as a potential local for O2 •− generation in T. brucei. On the contrary, in L. donovani promastigotes treated with antimycin (56) or with the primaquine analogue tafenoquine (14), as well as in T. cruzi epimastigotes cultured with Serratia marcescens prodigiosin or stimulated with antimycin (38), O2 •− formation was readily triggered at this complex. None of these studies has, however, addressed the submitochondrial compartmentalization of the produced ROS. In other systems, O2 •− may be directed to the matrix or to the intermembrane space, depending on which of the two sites within mammalian complex III (designated as Qi and Qo) generates this radical species.
Complex IV and the alternative terminal oxidase of T. brucei
Complex IV, or cytochrome c oxidase, is not regarded as a source of ROS, neither in trypanosomatids nor in other eukaryotes. Nevertheless, a diminished oxidase function of complex IV, either inherent to the parasite or caused by drugs, obviously induces mitochondrial ROS production as any constraint to electron flow favors their escape to O2. A lower activity of complex IV, together with an increased activity of complexes II and III, was referred to as the reason why T. cruzi trypomastigotes generated higher levels of ROS than epimastigotes (41).
When discussing ROS production in trypanosomatid mitochondria, it is important to mention trypanosomatid alternative oxidase (TAO) (Fig. 3). This protein, which is restricted to the inner mitochondrial membrane of both bloodstream and procyclic forms of T. brucei, does not exhibit proton translocation capacity and, therefore, cannot contribute to the proton gradient that drives ATP formation (23), but functions as an electron acceptor. In the parasite mammalian forms, TAO completely replaces complex IV, its function being to oxidize UBQ formed in consequence of G3PDH activity. In the insect stage, TAO coexists with complex IV and, albeit less active than in bloodstream forms (23), it is also functional. In fact, TAO could substitute for complex IV when this was silenced by RNAi (46). Interestingly, it was suggested that, by assisting complex IV in removing excess reducing equivalents and using these to reduce O2 to water, TAO could function in procyclics to decrease mitochondrial O2 •− (34).
Fumarate reductase as a potential site for ROS generation
In addition to the classic respiratory chain components, other mitochondrial NADH-dehydrogenases may leak electrons directly to O2 and, in consequence, give rise to O2 •− (48). In trypanosomatids one of these is mitochondrial fumarate reductase (mFR), a Krebs cycle enzyme that regenerates succinate from fumarate at the expenses of NADH in what constitutes a peculiarity of these parasites (Fig. 3). Mitochondrial fumarate reductase was demonstrated to be a potential site for O2 •− formation in T. brucei and T. cruzi insect stages (13, 26, 33, 84).
ROS Elimination in Trypanosomatid Mitochondria
Superoxide anion formed in mitochondria may give rise to other ROS (Fig. 4). Among these, H2O2 may cross biological membranes (8) and exert its effects in other subcellular compartments. Apart from participating in physiologic processes (e.g., by regulating signaling pathways) (10), H2O2 may have deleterious consequences on cells owing to its capacity to react with biological molecules as well as to originate the highly reactive hydroxyl radical (•OH). The fine regulation of ROS levels in mitochondria is thus required, so that the cytotoxic impact of these molecules is circumvented without compromising their physiologic functions. Enzymes with ROS-eliminating activity as well as nonenzymatic mechanisms are implicated in such control. Next, we will review the available information on the antioxidant enzymes present in trypanosomatid mitochondria (Fig. 5).


Superoxide dismutases
Elimination of O2
•− is an adequate means to regulate the formation of other ROS. Even though this may occur by the spontaneous dismutation of superoxide to H2O2 and O2, this reaction is accelerated by the activity of superoxide dismutases (SODs) (55). These enzymes dismutate O2
•− following a mechanism that involves a metal (M) co-factor, as detailed next:
Trypanosomatid mitochondria are equipped with two SODs, FeSODA and FeSODC (30, 89) (Fig. 4). Noticeably, these two enzymes utilize iron as metal co-factor and not manganese as mitochondrial SODs of other eukaryotes. This dependence on iron is reminiscent of SODs of prokaryotes and plant chloroplasts. The submitochondrial compartmentalization of FeSODA and FeSODC has never been experimentally addressed, but, at least in the case of the former enzyme, bioinformatic analysis supports its residence in the matrix (9). As for FeSODC, its unusually large N-terminal signal peptide was suggested by Dufernez et al. to target the enzyme to the intermembrane space (30) and by Bodył et al. (9) to the matrix, its final destination remaining doubtful. As a note, one alternative means to remove O2 •− in this specific compartment is via its cyt c-driven oxidation to O2 (Fig. 4), in analogy to what is described for other systems (70). Additionally, it is conceivable that, as reported for other organisms, O2 •− moves through anion channels into the cytosol (44) where it can be eliminated by local antioxidants.
The role of both FeSODA and FeSODC as antioxidants has been evidenced by forward and reverse genetics. Illustrating this, overexpression of FeSODA was reported to protect Leishmania from the free radical generating agents paraquat, nitroprusside (67), and antimycin (39), whereas downregulation of FeSODC by antisense RNA rendered Leishmania tropica promastigotes more sensitive to menadione (40), a known O2 •− inducer. Likewise, knocking down of FeSODA by RNAi had a negative impact on the resistance of T. brucei bloodstream form parasites to paraquat, a phenotype that was not reproduced by FeSODC-deficient parasites (89). Even though FeSODA and FeSODC are apparently active as antioxidants in the parasite context, neither enzyme is crucial for survival of bloodstream T. brucei parasites (89). This fact may result from a mutual compensation of function or it may reflect an insufficient generation of O2 •− by the rudimentary mitochondrion of T. brucei at this life stage. In Leishmania the impact of mitochondrial SODs on parasite survival appears more prominent, as lack of FeSODC resulted in reduced survival of L. donovani amastigotes in mouse macrophages (40). In the case of T. cruzi, the report that FeSODA expression is upregulated in infective metacyclic trypomastigotes relative to noninfective epimastigotes (6) suggests that this enzyme may also play a role in parasite adaptation to the mammalian host. This observation may, nevertheless, apply only to some parasite strains, as it was reported that FeSODA expression may vary with the T. cruzi lineage (69). One final note to mention that the antioxidant activity of FeSODA may also account for its involvement in the regulation of apoptotic-like death, as suggested for both T. cruzi (72) and L. donovani (39).
Cytochrome c peroxidase
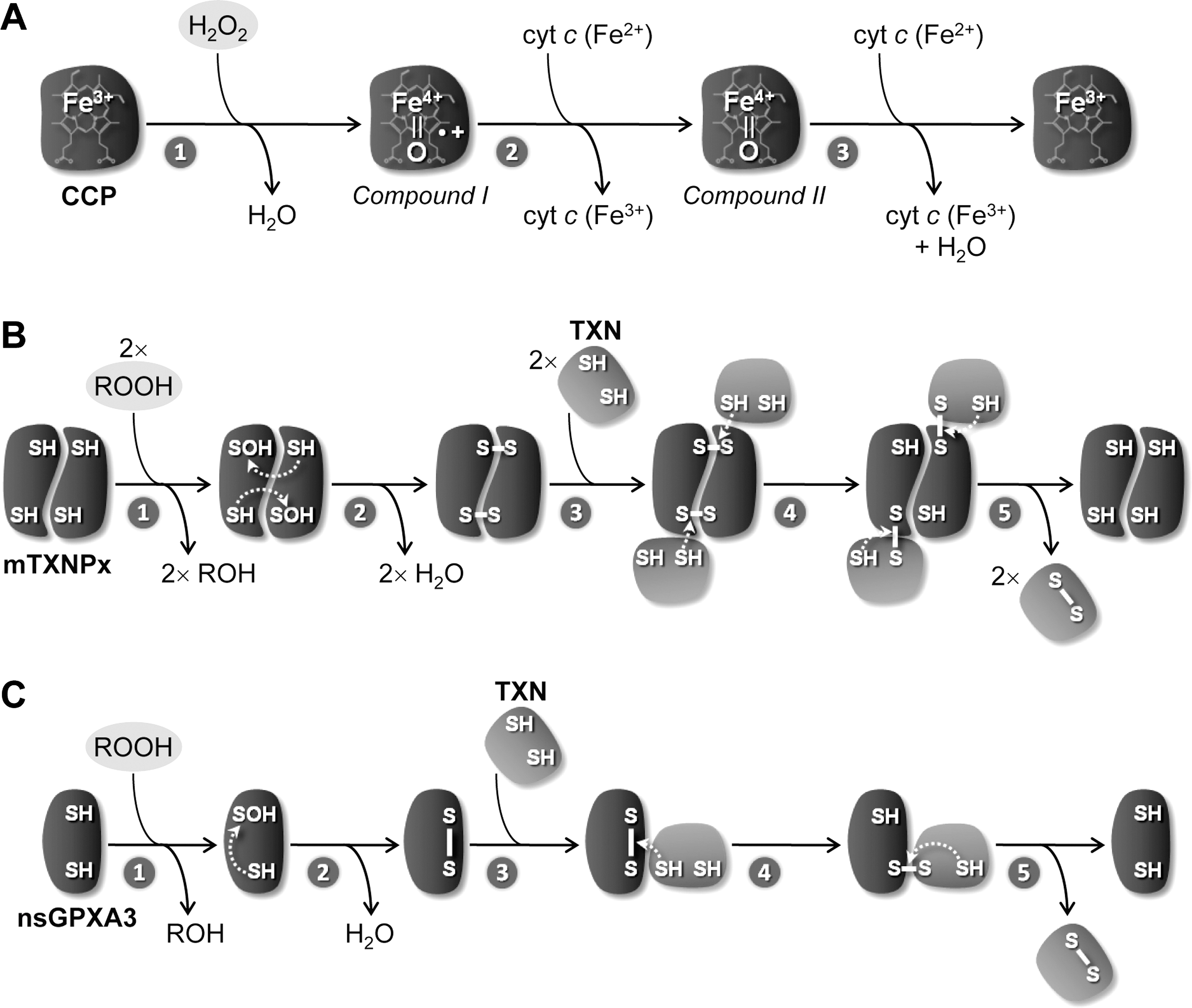
Within trypanosomatid mitochondria H2O2, the product of O2 •− dismutation, can be removed by different peroxidases. One of these is cytochrome c peroxidase (CCP), a heme-containing enzyme that is present in Leishmania spp. Originally described in Leishmania major (LmCCP) as an ascorbate-dependent peroxidase (2), this enzyme is now known to be preferentially reduced by ferrous cyt c (47) following the mechanism depicted in Figure 6A. LmCCP is localized in the inner mitochondrial membrane with its catalytic domain facing the intermembrane space (28). The enzyme plays an antioxidant role (28, 29, 66), but it is redundant throughout the L. major life cycle (66). This observation is not entirely surprising if one takes into consideration that LmCCP has no obvious representative in T. cruzi and T. brucei (Fig. 5). A specific review on LmCCP is presented in this Forum Issue by Adak and Pal.
Thiol peroxidases
The other hydroperoxide-reducing enzymes operating within trypanosomatid mitochondria are two thiol peroxidases, namely a 2-Cys-peroxiredoxin enzyme, designated mTXNPx (16, 82, 87) [or, according to the recent nomenclature proposed by Gretes et al. (42), Prx1m], and a nonselenium glutathione peroxidase, referred to as nsGPXA3 (45, 79). The activity of these molecules does not require any cofactor or prosthetic group and, instead, depends on the reactivity of two redox active cysteines. The mechanisms of reaction of these two classes of enzymes, originally dissected by Rhee (22), Poole (31), and Maiorino (53), are depicted in Figure 6B and 6C.
Both mTXNPx and nsGPXA3 are assumed to be present in the mitochondrial matrix, where they can exert their activity as H2O2-reducing enzymes (16, 74, 78) (Fig. 4). In addition to H2O2, these two enzymes accept small organic hydroperoxides (18, 27, 78) and, in the case of nsGPXA3, lipid hydroperoxides (27, 78) as oxidizing substrates.
Consistent with the in vitro activity of mTXNPx as a peroxidase, its ectopic overexpression was shown to confer parasites resistance towards H2O2 of exogenous origins (16, 49, 87). Based on these observations, the main physiologic function of mTXNPx was for a long time regarded to be that of an antioxidant. This concept was, nevertheless, challenged by a recent report demonstrating that the peroxidase function of mTXNPx is not crucial, even though the protein itself is essential during the mammalian stage of L. infantum (20). Instead, it is possibly the chaperone-like activity of mTXNPx that is required for amastigote replication (20). In T. cruzi, mTXNPx is also expected to be relevant for parasite survival in the vertebrate host, as suggested by the observations that this enzyme is upregulated in virulent T. cruzi strains (71) or when these parasites differentiate into metacyclic forms (6). Moreover, mTXNPx overexpression was found to enhance T. cruzi survival in mammalian cells (73). For the T. cruzi mTXNPx molecule, however, the actual contribution of its peroxidase activity for parasite survival remains to be investigated, following on the abovementioned report that its L. infantum counterpart displays an alternative function (20). In what concerns the mitochondrial nsGPXA3, functional studies in T. brucei revealed that this enzyme is not vital, its depletion having resulted in cardiolipin peroxidation and in a transient retardation of growth (27). That finding agrees with the fact that other trypanosomatids, namely T. cruzi and Leishmania braziliensis, can thrive in the absence of a nsGPXA3 orthologue (Fig. 5). Altogether, these observations suggest that when eliminated individually the mitochondrial thiol peroxidases, or more precisely their peroxidase activity, are not vital to trypanosomatids. One possible explanation for this redundancy could be that one molecule might overcome the absence of the other, even though they have different preferences for hydroperoxides. A definitive conclusion about the relevance of both mitochondrial thiol peroxidases as antioxidant devices would, hence, require their simultaneous down regulation. Another unsolved issue regarding mitochondrial thiol peroxidases is their physiologic source of reducing equivalents, as explained next.
Thiol peroxidases are typically reduced by CxxC-containing oxidoreductases of the thioredoxin family (which includes thioredoxins and glutaredoxins) or by proteins possessing thioredoxin-like active sites (Fig. 6B and 6C). Trypanosomatids harbor a specific thioredoxin-like subfamily of proteins, known as tryparedoxins (TXNs) (60), which are the preferred reducing substrates for thiol peroxidases in these organisms. Tryparedoxins are nonenzymatically reduced by trypanothione [T(SH)2] (32), a conjugate of glutathione and spermidine that largely replaces glutathione functions in trypanosomatids and that is itself maintained in the reduced state by the flavoenzyme trypanothione reductase (TR) at the expenses of NADPH. Whereas in biochemical assays, both mTXNPx and nsGPXA3 are reduced by the TR/T(SH)2/TXN system (18, 74, 78), it is not clear how this reaction proceeds in the parasite context. This doubt stems from three different reasons. First, the evidence for the presence of TR in mitochondria is conflicting. In an immunolocalization study in T. cruzi (45), TR was reported to partially localize to the mitochondrion (57), but this observation was not corroborated by subsequent studies (15, 78, 81, 88) based on subcellular fractionations of T. brucei, T. cruzi, and L. infantum (13, 58, 61, 67). Second, it is not known whether T(SH)2 is present in mitochondria. Since trypanothione synthetase appears to be restricted to the cytosol (65), the thiol would have to be imported into the organelle by means of carriers either specific or not. Up to now, no such transporters have been identified. Third, the requirement for a mitochondrial TXN is also questionable as trypanosomatids can thrive without a TXN molecule in this cell compartment (17). These organisms do possess a mitochondrial TXN-like enzyme (TXN3 in Leishmania spp. or TXN2 in T. cruzi and T. brucei), but this is a mitochondrial outer membrane protein that has no possibility to interact directly with, hence reduce, mTXNPx and nsGPXA3 (Fig. 5). Leishmania spp., but not T. cruzi and T. brucei, harbor an additional mitochondrial TXN molecule (the Leishmania TXN2) (19) that is, nevertheless, redundant (17). In this context, it is worth mentioning that the nonessentiality of a mitochondrial TXN also impacts on other proposed TXN-dependent mechanisms taking place in this organelle, namely the reduction of the universal minicircle sequence binding protein (UMSBP) that is required for kDNA replication (61).
In the absence of a mitochondrial TXN, reduction of thiol peroxidases might be carried out by other oxidoreductases containing both a CxxC motif and a thioredoxin domain. Among these, thioredoxins and glutaredoxins are unlikely to exhibit such activity. The latter, despite having representatives in the mitochondrion of these parasites (21, 36), are unable to replace TXN as reductants of thiol peroxidases (21, 54). As for thioredoxin, although the T. brucei enzyme was shown to reduce TXNPx (79) and nsGPXA3 (45) in vitro, its presence in the mitochondrion was never demonstrated. In what concerns other putative dithiol-containing oxidoreductases predicted in the genome of trypanosomatids, it is difficult to infer about their role as reductants for thiol peroxidases without experimental evidence.
Nonenzymatic scavengers of hydroperoxides in trypanosomatid mitochondria
In trypanosomatid mitochondria hydroperoxides may also be eliminated nonenzymatically. Since T(SH)2 is capable of scavenging H2O2 directly (12, 45), this thiol comes out as an attractive candidate to perform such a role, providing its presence in the organelle is ascertained. Apart from trypanothione, other low molecular mass thiols [e.g., glutathione, ovothiol (5), or free cysteine] as well as thiols exposed on protein surfaces (76) and other antioxidants (e.g., ascorbate) might contribute to mitochondrial antioxidant function. Determination of the concentration of trypanothione and of other thiols in the mitochondria of the several trypanosomatids will help elucidating the importance of nonenzymatic mechanisms on hydroperoxide removal in this organelle.
Conclusion
In spite of species and life cycle stage differences, mitochondria of trypanosomatids have the potential to generate ROS, either in the respiratory chain or in other sites. These organelles are equipped with several antioxidant enzymes, two of which dismutate O2 •− (FeSODA and FeSODC) and three that reduce H2O2 and other hydroperoxides (CCP, mTXNPx and nsGPXA3). Interestingly, all five enzymes are only preserved in Leishmania spp., with T. brucei and T. cruzi missing one or two of these molecules (Fig. 5). This fact, added to the findings that the antioxidant activity of these enzymes is not crucial for parasite survival, suggests that their functions may overlap and/or may be efficiently replaced by nonenzymatic scavengers of ROS. The redundant role of mTXNPx and nsGPXA3 as peroxidases is in agreement with the nonessential character of their preferred reducing agent (TXN) within mitochondria. In the future, the simultaneous depletion of several redundant antioxidant enzymes will certainly shed light on the relative importance of catalytic elimination of ROS in trypanosomatids.
Footnotes
Acknowledgments
The authors acknowledge Fundação para a Ciência e a Tecnologia, Portugal (grants SFRH/BPD/80836/2011, PTDC/CVT/70275/2006, PTDC/BIA-MIC/100910/2008 and PTDC/CVT/100090/2008) and Fundação Calouste Gulbenkian, Portugal (grant P-105335) for continuous financial support.
Disclosure Statement
No competing financial interests exist.