
Abstract
Introduction
P
After phagocytosis and activation of NADPH oxidase (NOX), neutrophils and macrophages produce reactive oxygen species (ROS) that kill invading microbes in the phagosomal compartment (2, 21). Several reports suggested that Ras-related C3 botulinum toxin substrate 2 (Rac2) protein is a key regulator of phagocytosis and formation of ROS (5, 8, 68). Malawista et al. (36) demonstrated killing of Staphylococci by non-ROS-producing neutrophil cytoplasts that was sensitive to NOS inhibitor, implicating the importance of NO and RNS. NO was also implicated in intracellular killing of microbes in macrophages (60). Kuncewiczk et al. (28) suggested physical interaction between iNOS and Rac2 (a NOX component) in LPS-stimulated murine macrophages; however, functional importance of complex was not studied. Activation of Rac2 and phagocytosis that followed IFN-γ priming was found to be associated with activation of iNOS in chronic granulomatous disease (CGD) patients (15). Role of NO in augmenting ROS and RNS in PMNs has been documented by us and others (32, 42, 44, 55, 56). The present study was undertaken to assess the functional importance of iNOS–Rac2 interaction in localization of iNOS and generating NO and oxygen/nitrogen radicals, protein-tyrosine nitration in human and mice PMNs.
The present study defines important features and actions of inducible nitric oxide synthase (iNOS) after phagocytosis of coated beads in human polymorphonuclear neutrophils (PMNs), which were confirmed by using PMNs from iNOS-knockout mice. Though reported for macrophages, the present study demonstrates interaction of iNOS and Rac2 in human and mice neutrophils, its functional role in iNOS translocation to phagosomal membrane, their functions in phagocytosis, reactive oxygen species/reactive nitrogen species generation, nitration, and elimination of phagocytosed pathogen for the first time. The information provided in this report will hopefully assist future studies to assess iNOS/Rac2 interaction in various physiological and pathological conditions.
We have demonstrated that iNOS was present constitutively in both human and mice neutrophils. After phagocytosis of bovine serum albumin (BSA) coated beads, increase in NO, superoxide radicals and ROS/RNS, and consequent BSA nitration was identified as iNOS dependent by using iNOS null neutrophils and iNOS specific inhibitor. Role of Rac2 in killing in vivo was demonstrated by pretreating mice with Rac inhibitor, NSC23766. Furthermore, our data suggested key role of Rac2 interaction in iNOS translocation to phagosomes. We also demonstrated the role of Rac2 in phagocytosis mediated elevation of NO and ROS/RNS, as it was decreased in presence of Rac inhibitor, NSC23766 or after knockdown of Rac2 in human neutrophils. Together, our study suggests importance of iNOS in ROS/RNS generation and microbial killing by neutrophils in Rac2 dependent manner.
Results
Expression of transcripts and detection of iNOS protein in human and mice PMNs
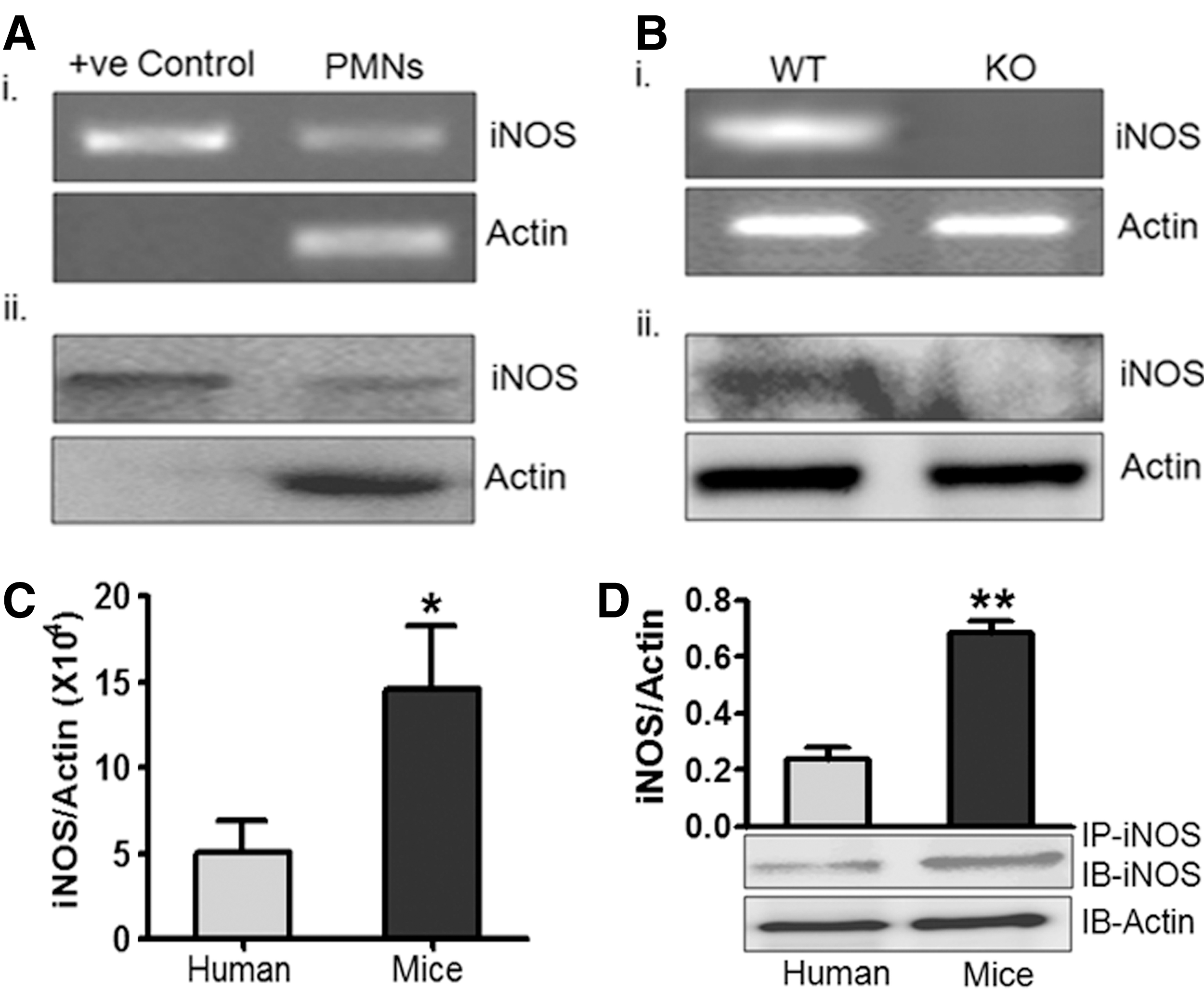
Constitutive expression of iNOS mRNA was found in healthy human peripheral PMNs with human iNOS clone as the positive control (Fig. 1A.i). Real time reverse transcription–polymerase chain reaction (RT-PCR) showed iNOS copy number 160–230/ng of total mRNA in human PMNs. Presence of iNOS protein in human PMNs total cell lysates was also confirmed by immunoprecipitation followed by immunoblotting using iNOS antibody (Fig. 1A.ii). Expression of iNOS mRNA and protein in wild type (WT) mice and iNOS-knockout (iNOS KO or iNOS−/−) was also measured and as expected mRNA and iNOS protein were present in WT mice PMNs but absent in PMNs from iNOS−/− mice (Fig. 1B). Moreover, iNOS mRNA and protein levels in mice PMNs were approximately three times higher than human PMNs (Fig. 1C, D). These results have clearly demonstrated constitutive presence of both transcript and protein of iNOS in human, as well as in WT mice PMNs, as reported earlier (6, 45).
Subcellular distribution of iNOS in human PMNs
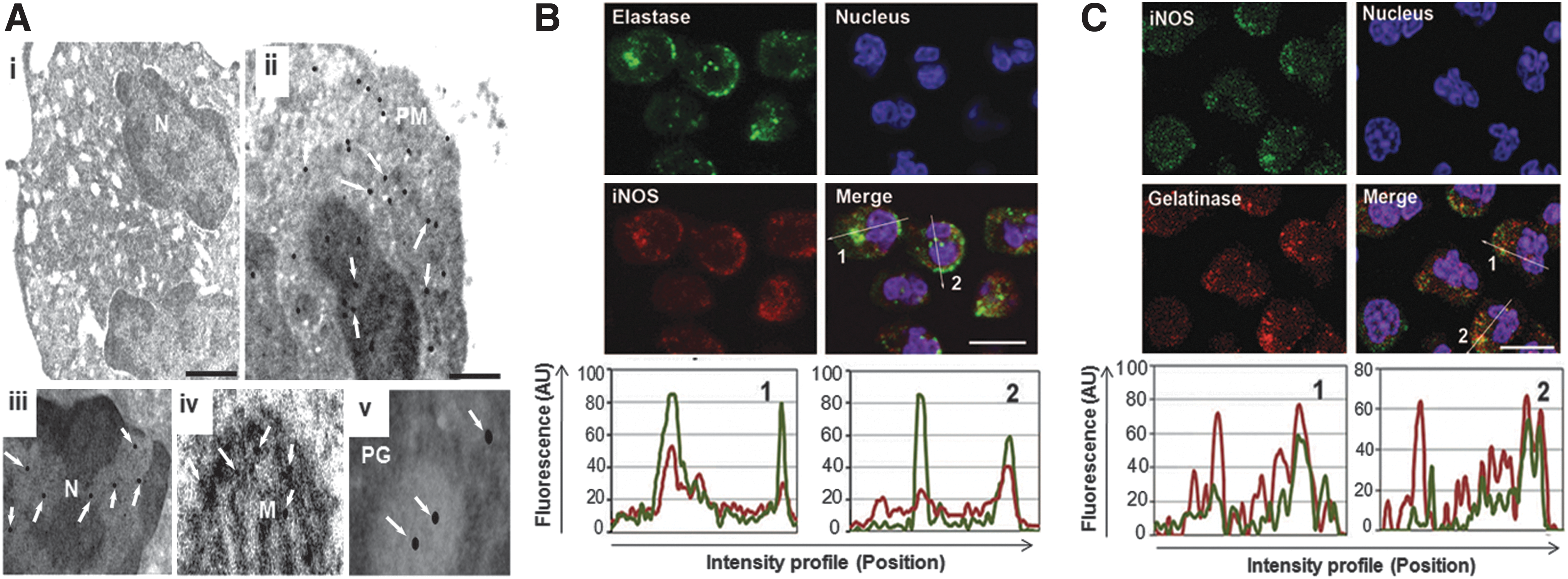
Intracellular distribution of iNOS protein in the human PMNs was explored as its activity is regulated temporally and spatially through its cell localization (49). Immunoelectron microscopy conducted with iNOS antibody and a secondary antibody conjugated with 10 nm gold particles showed distribution of iNOS protein in cytoplasm (Fig. 2A.ii), nucleus (Fig. 2A.iii) and other organelles. Higher magnification clearly showed gold particles corresponding to iNOS in mitochondria (Fig. 2A.iv) and granules (Fig. 2A.v). No significant binding was observed with isotype-matched control antibody or secondary antibody alone under similar experimental conditions (Fig. 2A.i). By using confocal microscopy, iNOS protein was also found to be colocalised with elastase and gelatinase; thus, confirming its presence in the neutrophils granules (Fig. 2B, C) and cytoplasm. In these experiments secondary antibodies were used to ensure the specific labeling and Z-stacks were used at each plane in similar conditions and no labeling was observed in the negative controls (data not shown). The results obtained so far demonstrate constitutive presence of iNOS in human PMNs and iNOS protein was localized in various subcellular compartments.
Phagocytosis in human PMNs generates ROS/RNS and NO
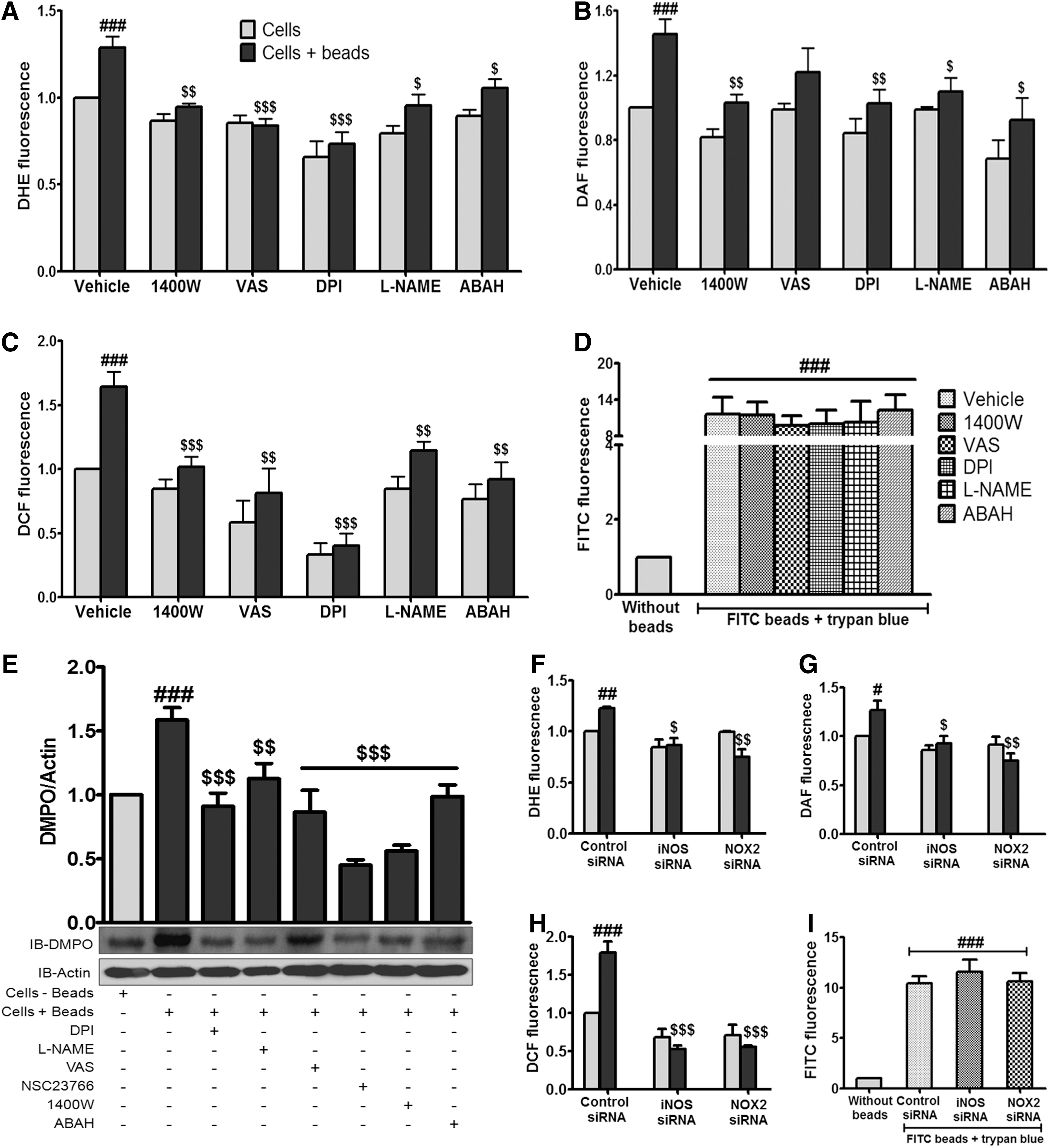
Experiments were directed to assess the functional significance of iNOS in the PMNs. Human PMNs were exposed to latex beads and generation of superoxide radical, NO and collective ROS/RNS was monitored. We detected ROS/RNS using multiple fluorescent probes and complementary methods, including 5,5-dimethyl-1-pyrroline-N-oxide (DMPO) adduct and nitrite formation after phagocytosis of BSA-coated beads. Enhanced fluorescence yields of preloaded dihydroethidium (DHE), 4,5-diaminofluorescein (DAF) and 2,7-dichlorofluorescein (DCF) in human PMNs after phagocytosis of beads, confirmed increased generation of superoxide radicals, NO and collective ROS/RNS, respectively (Fig. 3A–C). Further to identify the mechanistic details we pretreated human PMNs with 1400 W, a specific iNOS inhibitor; L-NG-nitroarginine methyl ester (L-NAME), a general NOS inhibitor; diphenyleneiodonium chloride (DPI), a NOS and NOX inhibitor; 1,3-Benzoxazol-2-yl-3-benzyl-3H-[1,2,3]triazolo[4,5-d]pyrimidin-7-yl sulfide (VAS), the NOX inhibitor or 4-aminobenzoic acid hydrazide (ABAH), a myeloperoxidase (MPO) inhibitor where DHE, DAF and DCF fluorescence yields in the control cells decreased or remain unaltered but the overall availability of the reactive species in the beads phagocytosed PMNs was considerably low notwithstanding the perceptible enhancement effect (Fig. 3A–C). It was found that the interventions used had no significant effect on phagocytosis of coated beads (Fig. 3D) using Trypan blue to differentiate between adherent and phagocytosed beads. As specificity of fluorescent probes is often in doubt, experiments were performed therefore, using alternative and specific methods, such as DMPO nitrone adduct antibody to capture collective ROS/RNS generation and Griess reagent for NO formation to confirm these observations. Enhanced ROS/RNS generation after phagocytosis of beads was evident by the formation of DMPO nitrone adducts (Fig. 3E) and pretreatment of PMNs with DPI, L-NAME, VAS, 1400 W, or ABAH reduced the collective formation of ROS/RNS (Fig. 3E). Augmentation in NO formation after phagocytosis was also reflected in the nitrite content (Supplementary Fig. S1A; Supplementary Data are available online at
Involvement of iNOS in phagocytosis induced NO and ROS/RNS generation in mouse PMNs
To verify our hypothesis that an increase in ROS/RNS generation was associated with NO and ROS/RNS generation from iNOS, experiments were performed in WT and iNOS−/− mice. Like human PMNs, mice PMNs also generated NO, superoxide radicals and ROS/RNS after phagocytosis of beads and it was significantly attenuated in the presence of 1400 W, VAS, DPI, L-NAME, or ABAH (Fig. 4A–C). These interventions also had no effect on phagocytosis of beads (Fig. 4D). Similar to human PMNs, inhibition of NOS by L-NAME or 1400 W had similar effect on ROS/RNS generation under basal and stimulated condition in mice PMNs (Figs. 3 and 4). PMNs from iNOS−/− mice generated significantly less amount of superoxide radicals, NO and ROS/RNS as compared to WT mice PMNs (Fig. 4A–C). Pretreatment of cells with 1400 W or DPI attenuated NO, superoxide radicals and ROS/RNS generation from WT mice PMNs, while 1400 W had no effect on NO, superoxide radicals and ROS/RNS generation from iNOS−/− mice PMNs, unlike DPI, a dual inhibitor of iNOS and NOX (Fig. 4A–C). Moreover, phagocytosis was comparable in PMNs from WT and iNOS−/− mice (Fig. 4D) suggesting that enhanced generation of ROS/RNS and NO after phagocytosis in WT neutrophils depended on iNOS, a significant contributor of NO in PMNs, particularly in inducing/demanding conditions.

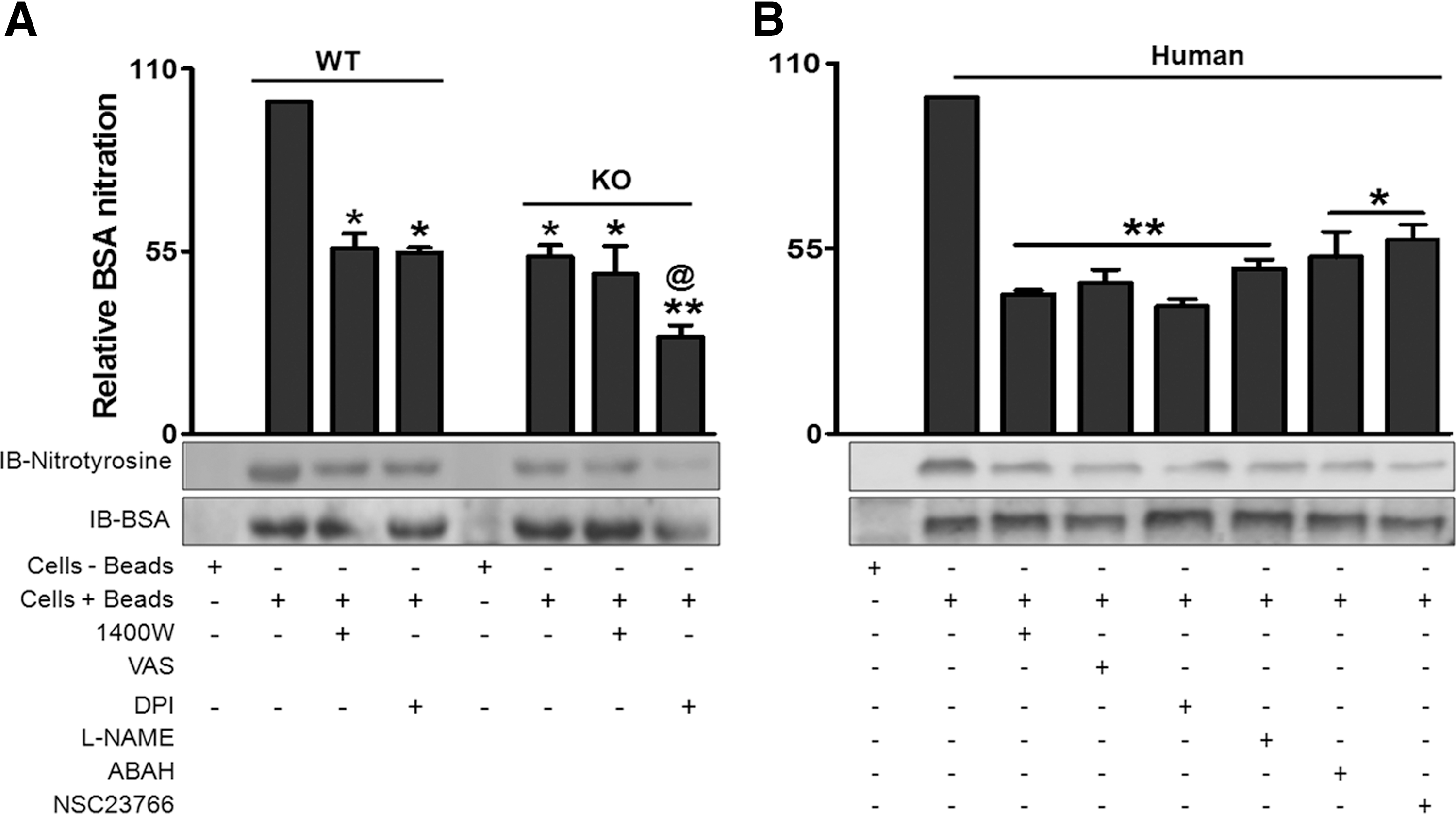
The increased NOS-dependent ROS/RNS generation after phagocytosis of beads led to a question as to whether the amount of nitrotyrosine truly reflects the production of ONOO− in these cells. Further by using iNOS−/− PMNs that displayed significantly less tyrosine nitration of BSA after phagocytosis than WT counterparts, we confirmed participation of iNOS derived reactive species in protein nitration (Fig. 5A). We also observed that the activity of tyrosine nitration was comparable in control and 1400 W pretreated iNOS−/− mice PMNs. In contrast WT PMNs exhibited high activity of tyrosine nitration and was sensitive to iNOS inhibitor, 1400 W (Fig. 5A). These data thus, specified involvement of iNOS and NOX-derived species in the protein nitration process in mice PMNs.
Tyrosine nitration of BSA-protein during phagocytosis
Experiments conducted so far confirm our hypothesis that NO enhances collective generation of ROS/RNS, as well as peroxynitrite formation. Monitoring by antibodies of nitrotyrosine we also found significant tyrosine nitration of BSA protein after phagocytosis of coated beads suggesting formation of peroxynitrite in the human PMNs too (Fig. 5B). Sensitivity of this activity to inhibitors, 1400 W (iNOS inhibitor), VAS (NOX inhibitor), DPI (NOS/NOX inhibitor), L-NAME (NOS inhibitor) and ABAH (MPO inhibitor) (Fig 5B) implicate actions of iNOS, NOX and MPO. Moreover, inhibition of phagocytosis by NSC23766, a Rac inhibitor led to the reduction of BSA tyrosine nitration. These findings clearly indicated that along with NOX and MPO, iNOS also play an important role in protein tyrosine nitration. We subsequently concentrated our efforts to find out the possible role of Rac in iNOS mediated NO generation (a least explored area of research), rather than divulging in to the chemistry of reactive species generated by modulating catalytic activity of NOX, NOS and/or MPO.
Effect of Rac inhibition/silencing on microbial killing and ROS/RNS generation
To test the role of Rac in microbial killing, regulation of NO generation and its interaction with iNOS, various sets of experiments were performed in mice treated with NSC23766 (IP-2 mg/kg, before 15 min of Escherichia coli/beads challenge) or vehicle. In the first set of experiments, mice that were pretreated with 2% starch to augment peritoneal PMNs population, were challenged intraperitoneally with E. coli (1×106 cells) after the protocols used in mice peritonitis model (40). In control mice without NSC23766, colony-forming units (CFU) in the peritoneal fluid were reduced significantly 1 h after the challenge reflecting the elimination after phagocytosis of E. coli by peritoneal PMNs, while CFU numbers in the peritoneal fluid of NSC23766 treated mice were significantly more suggesting reduction in phagocytosis (Fig. 6A, B). This suggested the inhibitory effect of Rac inhibition on phagocytosis in vivo (Fig. 6C). To validate in other set of experiments, mice were injected with FITC coated latex beads in the peritoneal cavity and assessed for phagocytosis by gating LY6G positive PMNs, which confirmed the inhibitory effect of Rac inhibition on phagocytosis (Fig. 6D–G). In yet another set of experiments, peritoneal cells collected from the vehicle and NSC23766 treated mice were challenged in vitro with latex beads to assess DHE, DAF and DCF fluorescence (Fig. 6H). PMNs from NSC23766 treated mice thus, exhibited significant reduction in phagocytosis and subsequent superoxide radicals, NO and ROS/RNS generation (Fig. 6). These experiments suggested a role of Rac in NO/ROS/RNS generation and microbial killing. Subsequent experiments were performed in vitro using human and mice PMNs to confirm the role of Rac in NO generation. As was seen in mice pretreated with NSC23766, incubation of human/mice PMNs with NSC23766 also significantly reduced phagocytosis (Fig 7A and Supplementary Fig. S2A), superoxide radical, NO and ROS/RNS generation (Fig. 7B–D and Supplementary Fig. S2B–D). To confirm the role of Rac in NO, superoxide radicals and ROS/RNS generation, Rac2 was silenced by using siRNA leading to ∼70% reduction of Rac2 expression in comparison to nontargeted control siRNA (Supplementary Fig. S1E). Rac2 knockdown in human PMNs led to inhibited NO and ROS/RNS augmentation particularly after bead phagocytosis (Fig. 7F–H). Moreover, Rac2 silencing also reduced phagocytosis of beads by 30%–50% in PMNs (Fig. 7E) confirming thereby an additional role of Rac2 in the modulation of iNOS activity. Experiments conducted so far by targeting Rac using various strategies demonstrated reduction in iNOS derived NO, ROS/RNS generation, tyrosine nitration, as well as microbial killing, and led us to explore the possibility of iNOS and Rac2 interaction if any in PMNs as was reported in macrophages (28).


Rac2 regulates iNOS translocation to the phagosomes
Experiments were performed to systematically assess iNOS interaction with Rac2. Presence of iNOS and Rac2 protein was initially checked in the resting and bead activated human PMNs by confocal microscopy after immunolabelling. The data obtained demonstrated presence of iNOS and Rac2 in the cytosol of the resting cells (Fig. 8A), while iNOS with Rac2 migrated to the phagosomal membranes after phagocytosis (Fig. 8A and Supplementary Fig. S3). Presence of iNOS in phagosomal membrane was also confirmed by colocalization of iNOS with phagosomal marker LAMP-1 (Supplementary Fig. S4E). Cells incubated alone with the secondary antibodies under similar equipment settings did not exhibit fluorescent signals (data not shown). These experiments suggest interaction of iNOS with Rac2 and also their redistribution after phagocytosis in the human PMNs, as evidenced by the overlapping signals in the histograms of these proteins (Fig 8A, lowest panel).

Interaction of iNOS with Rac2 and their translocation to the phagosomes was now examined in human PMNs by immunoprecipitation of the PMNs lysates using Rac2 antibody. The immunoprecipitate obtained from the resting, as well as activated human PMNs revealed presence of p67phox, a NOX subunit, which is well reported to interact with Rac2 in the cytosol of resting cells, its migration to the phagosomal fraction in the bead activated PMNs and reduction of its translocation in the NSC23766 treated cells (Fig. 8B). Probing of these membranes with anti-Rac2 confirmed presence of Rac2 in the cytosol of the resting cells, phagosomal fraction of the activated cells and inhibition of its migration in NSC23766 pretreated PMNs (Fig. 8C). These membranes when blotted with iNOS antibody, showed iNOS presence in the cytosol of resting cells, in the phagosomal fraction of activated cells and inhibition of its migration in NSC23766 pretreated PMNs (Fig. 8D). Same was observed in the lysates of resting PMNs after IP with iNOS (Supplementary Fig. S4A, B) and rabbit IgG (negative control) under similar conditions showed no response in these blots. Moreover, expression of iNOS and Rac2 mRNA and protein was not altered after phagocytosis of beads in the presence or absence of NSC23766 (Supplementary Fig. S5A–D); thus, indicating that the changes observed were not due to the augmented protein levels. Rac2 mediated iNOS translocation was now assessed by silencing of Rac2 in human PMNs. Silencing of Rac2 inhibited the translocation of iNOS from cytosol to the membrane fraction (Fig. 8E, F). Interaction of iNOS and Rac2 was also assessed in NOX2 silenced human PMNs. Equal expression of iNOS and Rac2 in control and NOX2 silenced human PMNs confirms that iNOS/Rac2 interaction is independent to NOX2 (Supplementary Fig. S4C, D). Moreover, Rac2 and iNOS interaction was also apparent in WT mice PMNs, while in iNOS−/− only Rac2 was found (data not shown). These findings suggest critical role for the distinctive phagosomal translocation of the interacting partners, Rac2 and iNOS, in their cellular responses.
The experiments conducted in the present study thus, demonstrate importance of iNOS in the collective ROS/RNS generation, tyrosine nitration and microbial killing, after phagocytosis due to its redistribution along with Rac2 in the phagosomal compartment.
Discussion
Characterization of the role of iNOS in generation of ROS/RNS and identification of interacting protein partners in human PMNs is critical for gaining insight into the complexity of infective and inflammatory pathologies associated with many human disorders attributed to oxidative/nitrosative stress (14, 46, 54, 57, 58). Circulating NO and its metabolites derived from PMNs (53) are needed for systemic blood pressure regulation (37), aortic rings relaxation (47), inhibition of platelet aggregation (11) and microbicidal action (13, 22, 36).
The effect of NO depends on its steady-state levels and on the sub-cellular organelle from where it is being produced. Moreover, NO production and the subcellular distribution of NOS isoforms seem to play a major role in health and in disease, which have been associated with alterations in NO production (61). This study also demonstrated the presence of iNOS in elastase containing primary and gelatinase containing tertiary granules, intracellular compartments, plasma membrane and most prominently in the cytoplasm of human PMNs (Fig. 2). Tyrosine nitration of ingested bacteria (13) by this stored iNOS is likely to be part of the implied role of iNOS in killing the phagocytosed bacteria. NO production in the subcellular compartments, its cellular targets and downstream effects thus, need to be understood in PMNs in the context of various pathological conditions.
The key isoform of NOS enzyme, iNOS, is known to produce sustained amount of NO during immune and inflammatory responses. Here, we assessed its role in the generation of NO, superoxide radicals and ROS/RNS after phagocytosis of coated beads by human and mice PMNs. We used the technique of flow cytometry that gives precise information pertaining to the cell population and particle uptake (12), DMPO method, a reliable method of collective ROS/RNS generation (41) and Griess reagent (55) to measure nitrite, the metabolic end product of NO formation. Our data demonstrated that, in addition to NOX (9) and MPO (41), iNOS also has a role in generating superoxide radical and ROS/RNS (Figs. 3 and 4) and the consequent protein-tyrosine nitration (Fig. 5). Strong evidence for the participation of iNOS in these activities was provided by their inhibition on addition of the iNOS inhibitor, 1400 W (Ki=7 nM for iNOS, Ki=2 μM for nNOS) (19) and by using iNOS siRNA (Figs. 3 and 4). PMNs treated with L-NAME, a non specific NOS inhibitor, also exhibited similar results suggesting importance of iNOS (Figs. 3 and 4). Human and mice PMNs constitutively expressed iNOS though human cells possess less amount of iNOS protein (Fig. 1C, D) (6). Indeed, we also found constitutive expression of iNOS in PMNs isolated from German subjects. Previous studies have proposed a role of iNOS in the modulation of PMNs functions in cytoplasts (36), CGD patients (15) and in iNOS−/− mice (35). The detection of very low responses in PMNs of iNOS−/− mice in the present study is in agreement with these reports (Fig. 4).
Bimodal action of NO both as a facilitator (3, 4) and as a protector of oxygen free radicals (62, 64) is reported. Interaction of NO with superoxide anion, both formed in stimulated PMN's, generates peroxynitrite, which nitrates tyrosine residues (3, 4). Peroxynitrite dependent protein nitration of BSA was shown by Kuo et al. (29) and Galinanes et al. (18). Earlier reports also suggested the role of NOX and MPO on protein tyrosine nitration (17, 20, 24). Nitration of BSA was also observed after in vitro treatment of BSA with a NO donor and superoxide radical generating system and it was inhibited in presence of iNOS, NOX and MPO inhibitors (data not shown), similar to nitration of BSA after phagocytosis of the coated beads (Fig. 5). Results obtained in human (Figs. 3 and 5B) and WT mice PMNs (Figs. 4 and 5A) indicate the importance of MPO in augmenting ROS/RNS formation and BSA nitration. Moreover, significant amount of BSA nitration in iNOS−/− (Fig. 5A) further support the role of MPO in ROS and RNS generation.
Xia et al. (66) demonstrated that purified iNOS is capable of generating superoxide radicals in addition to NO and was significantly reduced by
The present study implies a role of iNOS, NOX and MPO derived species in nitration of phagocytosed particles, and the role of iNOS was confirmed by using PMNs from iNOS−/− mice. NOX also potentiated NO mediated protein tyrosine nitration in phagosomal compartment, which was reduced in presence of VAS, a NOX inhibitor (Fig. 5B) with no significant change in DAF fluorescence (Figs. 3B and 4B). Surprisingly, we also observed less NO in Nox2 knockdown PMNs but find it difficult to explain these observations with current understanding of NOS and NOX regulation. Since both iNOS and NOX subunit (p67phox) translocate to phagosome during phagocytosis with Rac2, it could be possible that they move as a complex and their activity is regulated by Rac2. We also observed iNOS and Rac2 interaction in Nox2 silenced human PMNs (Supplementary Fig. S4C, D). Role of Rac2 (68), NOS (36) and interactive effect of RNS and ROS (22) in microbial killing has been documented. Decreased bacterial killing and azurophilic granule secretion was observed in patients harboring Rac2D57N dominant negative mutation (1, 30, 63). Our ex vivo and in vivo studies showed that Rac2-inhibition by NSC23766 reduced bacterial killing (Figs. 6 and 7). Moreover, studies with human/mice PMNs treated with NSC23766 or Rac2 silenced human PMNs also showed inhibition of superoxide, NO and ROS/RNS production after phagocytosis of particles (Figs. 6H and 7B–D, F–H and Supplementary Fig. S2B–D). Our findings are not only in tune with clinical observations (1, 30, 63) suggesting critical role of Rac2 in various pathological conditions, but also suggest involvement of iNOS.
Recruitment of NOX subunits and their assembly on the newly formed phagosome is necessary for oxidative burst (7). As proposed earlier (9, 48), our finding also supported translocation of NOX component p67phox along with Rac2 to phagosomal compartment after phagocytosis of beads/microbes (Fig. 8B). In the present study Rac2 was identified as an iNOS interacting protein in human (Fig. 8 and Supplementary Fig. S4A, B) and mice PMNs, as evidenced from the coimmunoprecipitation and immunocytochemical experiments. NSC23766, a Rac inhibitor or silencing with Rac2 siRNA inhibited phagocytosis and generation of ROS/RNS (Figs. 6 and 7 and Supplementary Fig. S2), nitration of phagocytosed particle (Fig. 5B) and iNOS translocation to phagosome/membrane fraction (Fig. 8D, F); thus, marking the functional importance of Rac2–iNOS interaction. Rac activity, that is, Rac-GTP has been reported to increase during phagocytosis (16). NSC23766 inhibits Rac-GTP form and blocks most of Rac functions among which iNOS translocation is to be included. These results indicated that iNOS and Rac2 are functionally coupled through their effects on both iNOS-mediated NO generation and Rac2-driven NOX oxidase activity. Association of Rac2 with iNOS and NOX-2 in bead activated PMNs will enable spatially regulated NO and superoxide radical generation in the close proximity and will also facilitate production and formation of peroxynitrite.
Summarizing the findings, we demonstrate iNOS–Rac2 interaction in neutrophils and also the key role of iNOS in ROS/RNS and nitration using various methodologies to deplete iNOS. Furthermore, the study recognized Rac2 mediated regulation of iNOS translocation to phagosomes, ROS/RNS generation; iNOS mediated nitration and microbial killing in a model of peritonitis. Thus, our study suggests an important role of Rac2 in iNOS dependent ROS/RNS generation and microbial killing in neutrophils.
Materials and Methods
Isolation of PMNs from blood
Blood was collected from the anticubital vein in citrate-phosphate-dextrose (1:7) from healthy volunteers after prior consent. The donors were free from any major disease and any prior medication for last 72 h. PMNs were isolated as described previously (41) and purity was ascertained by using CD15 antibody using FACS Calibur (Becton Dickinson) and it was always more than 90%. Viability of the isolated PMNs, as assessed by Trypan blue exclusion was more than 98%. The study was performed in accordance with the guidelines of the ethical committees of SGPGI, KGMU and CSIR-CDRI (Lucknow, India).
Isolation of PMNs from mice bone marrow
Studies with C57BL/6 wild-type (WT) and iNOS−/− mice were performed according to the Institutional Animal Ethics Committee in Goethe University, Frankfurt, Germany and CSIR-CDRI, India. Mice were killed by cervical dislocation and femurs and tibiae were collected into Hank's balanced salt solution (HBSS) containing 0.1% BSA. After cleaning soft tissue and muscle from bone, cells were flushed in HBSS containing 1% BSA and 15 mM EDTA. The cell suspension was centrifuged at 400 g, the pellet was resuspended in 45% (v/v) Percoll (GE Healthcare) containing 0.1% BSA. The suspension was laid over three-layers of Percoll gradient (81%, 62% and 55%) and centrifuged at 1000 g for 30 min at 4°C (Sigma 3K30). The neutrophils between 81% and 62% layer were harvested and washed with HBSS (33). The purity of isolated PMNs was assessed by LY6G and CD11b antibodies using flow cytometry (FACS Calibur; Becton Dickinson). To eliminate monocyte contamination, the cells were gated on anti-Ly-6Cmed/lo (31, 59).
Generation of superoxide radicals and ROS/RNS
PMNs (1–2×106 cells/ml) pretreated with various interventions [vehicle, 1400 W (100 μM), VAS (25 μM), DPI (10 μM), L-NAME (1 mM), ABAH (100 μM) or NSC23766 (100 μM)] for 15 min at 37°C or after silencing with siRNA, were loaded with 2,7-dichlorofluorescein diacetate (DCF-DA, 10 μM) or dihydroethidine (DHE, 10 μM) for 10 min. Subsequently, latex beads were added to PMNs suspension and incubated for 30 min at 37°C, superoxide radicals (DHE) and ROS/RNS (DCF) generation was assessed by acquiring 10,000 events using FACS Calibur and analysed by Cell Quest programme as described earlier (27).
Collective, ROS/RNS generation was also determined by using DMPO as described earlier (10, 41). PMNs (2×106cells/ml) pretreated with vehicle, 1400 W (100 μM), VAS (25 μM), DPI (10 μM), L-NAME (1 mM), ABAH (100 μM) or NSC23766 (100 μM) for 15 min were subsequently incubated with beads and DMPO (50 mM) at 37°C (10, 41). Cells were lysed after 30 min of incubation by adding ice cold lysis buffer [10 mM Tris, 0.9% NaCl, 1 μg/ml Aprotinin, 100 μg/ml PMSF, 20 μg/ml Pepstatin A, 1 mM EDTA pH 7.4 and 5 mM diisopropyl fluorophosphates (DFP)] and the protein (30 μg) from each sample was loaded on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membrane, blocked with 5% BSA in TBST and then incubated with rabbit anti-DMPO-nitrone antibody (1:3000 in 1.5% BSA TBST) for 2 h at room temperature.
Generation of NO
PMNs (1–2×106 cells/ml) pretreated with vehicle, 1400 W (100 μM), VAS (25 μM), DPI (10 μM), L-NAME (1 mM), ABAH (100 μM) or NSC23766 (100 μM) for 15 min at 37°C or after silencing with siRNA were loaded with 4,5-diaminofluorescein diacetate (DAF-2DA, 10 μM) for 10 min. Latex beads were added to the samples and incubated for 30 min at 37°C. Fluorescence of at least 10,000 cells was acquired by FACS Calibur and subsequently analysed using Cell Quest programme (27).
Phagocytosis assay
FITC labeled latex beads were added to PMNs (1:50) suspension containing Ca2+ and Mg2+ and incubated for 30 min at 37°C to assess phagocytosis (25). Phagocytosis was also determined in 1400 W (100 μM), VAS (25 μM), DPI (10 μM), L-NAME (1 mM), ABAH (100 μM) or NSC23766 (100 μM) pretreated PMNs or in PMNs after silencing with siRNA, by incubating cells with FITC-coated beads. Trypan blue (200 μg/ml) was used to quench the fluorescence and to differentiate between phagocytosed and adherent beads. Samples (10 –20,000) were acquired using FACS Calibur and were analysed by Cell Quest programme.
BSA nitration
PMNs (5×106 cells) pretreated with vehicle, 1400 W (100 μM), VAS (25 μM), DPI (10 μM), L-NAME (1 mM), ABAH (100 μM) or NSC23766 (100 μM) for 15 min were subsequently incubated with BSA-coated beads for 30 min at 37°C. After phagocytosis, PMNs were subjected to hypotonic lysis, and centrifuged at 13,000 g for 5 min, supernatant was discarded and the beads were mixed with 2×Laemmli's sample buffer (26). After SDS-PAGE, the amount of BSA loaded was assessed by western blot using BSA antibody (1:3000) and its nitration by probing with nitrotyrosine antibody (1:3000). Relative BSA nitration in each sample was taking a ratio of loaded BSA and nitrated BSA after densitometry analysis of respective bands using image J software (National Institute of Health).
iNOS transcripts by RT-PCR
Total RNA (5 μg), isolated from human or WT/iNOS−/− mice PMNs using Tri reagent (Invitrogen), was reverse transcribed with RevertAid™ H Minus first strand cDNA synthesis kit using oligo (dT) primer as per the manufacturer's instructions. RT-PCR with iNOS primer (Table 1) was performed as described previously (52). Human iNOS clone (pcDNA3-iNOS) was used as positive control for iNOS. β-Actin was used as reference gene for normalization. Copy number of iNOS gene in human PMNs was determined by real-time PCR by comparison with a standard curve using known amounts of pcDNA3-iNOS plasmid (1 ng of DNA represents 108 copies of iNOS gene).
Neutrophil transfection
Human neutrophils (2×106 cells) were resuspended in 100 μl Nucleofector® solution V (Lonza) containing 5 μg of iNOS, Rac2, Nox2, or Control siRNA. Cells were transfected using Nucleofactor II electroporation device (Amaxa Biosystems) and the Nucleofector™ program Y-019. After transfection, cells were removed carefully and immediately transferred to 12-well plates containing RPMI medium supplemented with 10% FBS and 100 ng/ml GM-CSF to prevent PMNs apoptosis. Neutrophils were maintained for 24 h at 37°C in 5% CO2 incubator. After 24 h incubation, viability of PMNs was found to be around 80% as assessed by PI staining using FACS Calibur.
Assessment of iNOS protein in PMNs
Human and WT/iNOS−/− mice PMNs (total protein 750 μg) were lysed in ice-cold radio immunoprecipitation assay (RIPA) buffer [phosphate-buffered saline (PBS) containing 1 mM EDTA, 1 mM sodium orthovanadate, 1 mM sodium fluoride, 1 μg/ml aprotinin, 100 μg/ml PMSF, 20 μg/ml pepstatin, 5 mM DFP, 1% Triton X-100, and 0.1% SDS] for 30 min at 4°C. Immunoprecipitation and western blotting with iNOS antibody were performed as described previously (52). iNOS recombinant protein (20 ng; Cayman chemicals) was used as a positive control.
Subcellular distribution of iNOS by immunolabelling
Human resting PMNs (5×104 cells) and PMNs after phagocytosis of beads were fixed in 4% (w/v) paraformaldehyde in PBS (pH 7.4) at 25°C for 30 min and washed twice for 5 min each with PBS containing 0.5% (w/v) glycine. The washed cells were allowed to immobilize on 0.01% (w/v) poly-
Preparation of cytosolic, membrane and phagosomal fractions
Human PMNs (1×108 cells) pretreated with vehicle or NSC23766 (100 μM) for 15 min were incubated with latex beads at 37°C for 30 min. After phagocytosis PMNs were split in equal numbers in two tubes, one tube was used to isolate phagosomes as described earlier (43), while the other tube was used to isolate cytosolic fraction (16). Cytosol and membrane fraction were also prepared from resting human PMNs (5×107 cells) and from Rac2 siRNA transfected human PMNs (1×107 cells) (16). Detailed Supplementary Materials and Methods section are given in the online Supplementary Data.
Immunoprecipitation of Rac2 and iNOS
All samples (750 μg proteins) were immunoprecipitated with 1 μg of Rac2/iNOS antibody overnight at 4°C followed by incubation with 40 μl Protein A Sepharose for 2 h at 4°C. The immune-precipitates were collected after centrifugation followed by washing with RIPA buffer (PBS containing 1 mM EDTA, 1 mM sodium orthovanadate, 1 mM sodium fluoride, 1 μg/ml aprotinin, 100 μg/ml PMSF, 20 μg/ml pepstatin, 5 mM DFP, 1% Triton X-100 and 0.1% SDS) and wash buffer (50 mM Tris, pH 8.0). The immune complexes were run on 10% SDS-PAGE and transferred to PVDF membrane followed by immune-blotting with p67phox, Rac2, or iNOS antibody to observe the relative changes of p67phox/Rac2/iNOS in the cytosol, membrane and phagosome fractions. β-Actin was used as loading control in each supernatant/sample.
In vivo phagocytosis in peritoneal neutrophils
To augment peritoneal PMNs populations, mice were injected intraperitoneally (i.p.) with 1 ml of 2% starch in PBS and after 48 h mice were administered vehicle (PBS) or NSC23766 (2 mg/kg, i.p.) and were used to monitor the effect of Rac inhibition on phagocytosis and ROS/RNS formation (40, 51). Detailed Supplementary Materials and Methods section are given in the online Supplementary Data.
Statistical analysis
Data have been represented as mean±SEM from three to six independent experiments and were analyzed by one-way ANOVA test followed by Newman–Keul's post analysis. Student's t-test analysis was also used to analyze the samples and the data were considered significant at p<0.05.
Footnotes
Acknowledgments
The study was supported by a financial grant to M.D. from Department of Biotechnology-INDIGO and from CSIR network project BSC0102. Award of research fellowships to A.J., A.K.S., M.D., S.K., R.S., R.S.K. from the Council of Scientific and Industrial Research, India is gratefully acknowledged. We wish to profusely thank Prof. B. Bruene, Goethe University, Germany for providing iNOS KO mice and also the support provided to Mr. Abhishek Kumar Singh during his stay in his lab. Authors acknowledge Prof. S. Lamas, CBMSO, Madrid and Prof. Ramasarma, IISc Bangalore for the critical suggestions and their help during the preparation of the manuscript. The excellent technical help of Mr. A.L. Vishwakarma and Mrs. M. Chaturvedi for the Flow Cytometry study, Mr. Manish Singh and Ms. Kavita Singh of Confocal Imaging Facility, Ms. Abha Tamta and Madhuli in EM studies from Sophisticated Analytical Instrument Facility; CSIR-Central Drug Research Institute, Lucknow is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.