
Abstract
General Introduction
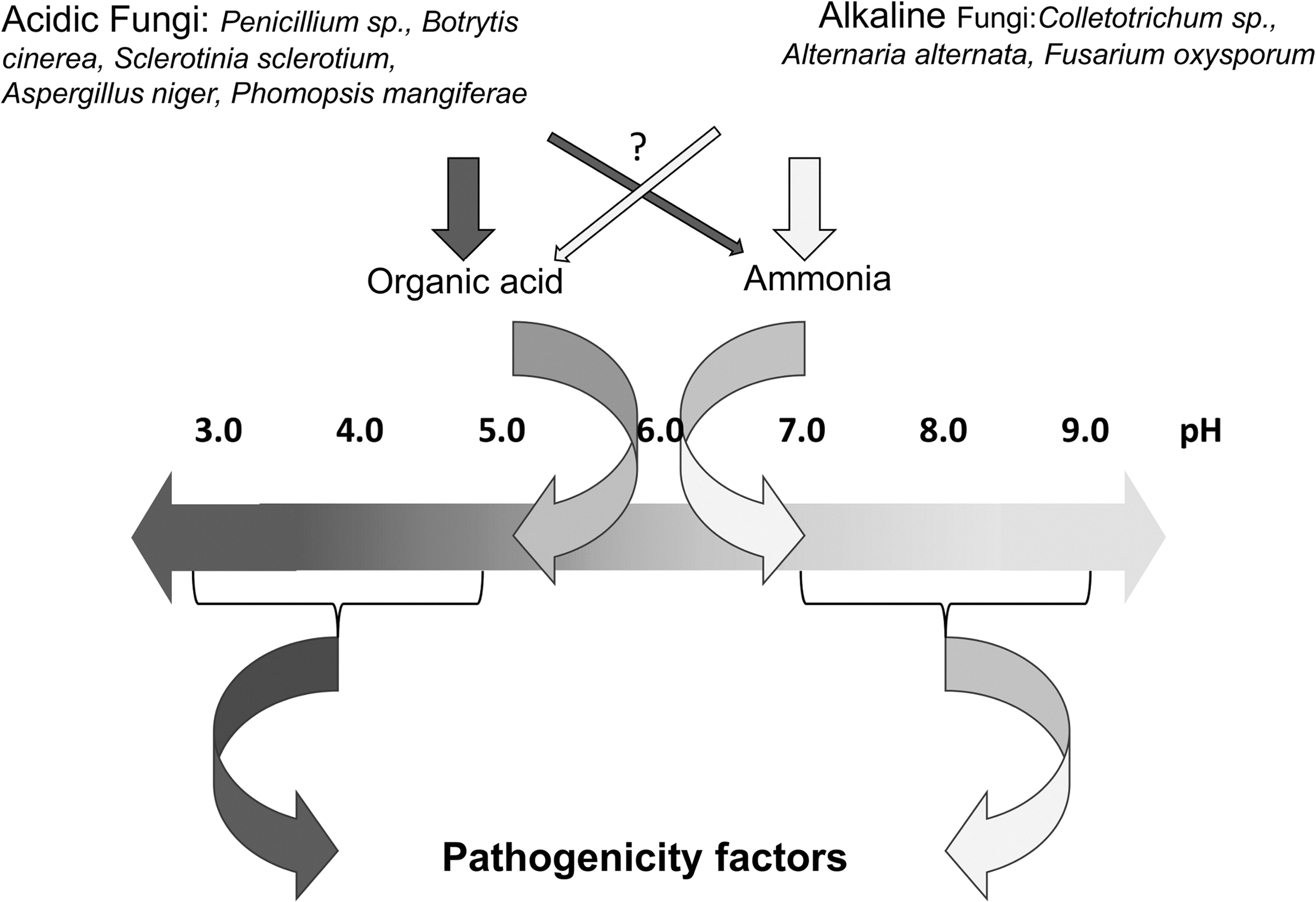
Ambient pH levels or the factors that determine host pH are important in that they determine the capability of the pathogen to successfully colonize, invade, and kill the targeted host, with the aid of secreted pathogenicity factors (Fig. 1). Since pH is a critical consideration in fungal attack strategy, the pathogen cannot simply adapt to any pH determined by its host; therefore, it has developed environmental sensing mechanisms that enable it to modify the ambient conditions to best suit its offensive arsenal and, at the same time, to compromise the host defense. Environmental pH is also one of the most important physiochemical factors that affect cell growth and development. In the case of multicellular organisms that have the facility to regulate the conditions of their internal environment, it has been shown that important cellular processes, such as protein synthesis, and growth and proliferation may be negatively impacted by even mild acidification (19, 29, 32, 106, 120). This alteration reflects the inability of cells to function normally at suboptimal pH. To survive under the varied pH conditions encountered, these organisms have developed mechanisms to monitor external pH, and they respond by regulating the entry and exit of appropriate ions. With regard to fungi, several mechanisms involved in this process have been described (25, 26, 80, 81,115, 127), but apparently the most widely distributed mechanism, with some variants, is a signal transduction pathway designated by various authors as RIM or PAL for yeasts or filamentous fungi, respectively (6, 9, 108, 115, 147), and that have received the general designation, PAL/RIM (5, 27, 59) (Fig. 2).

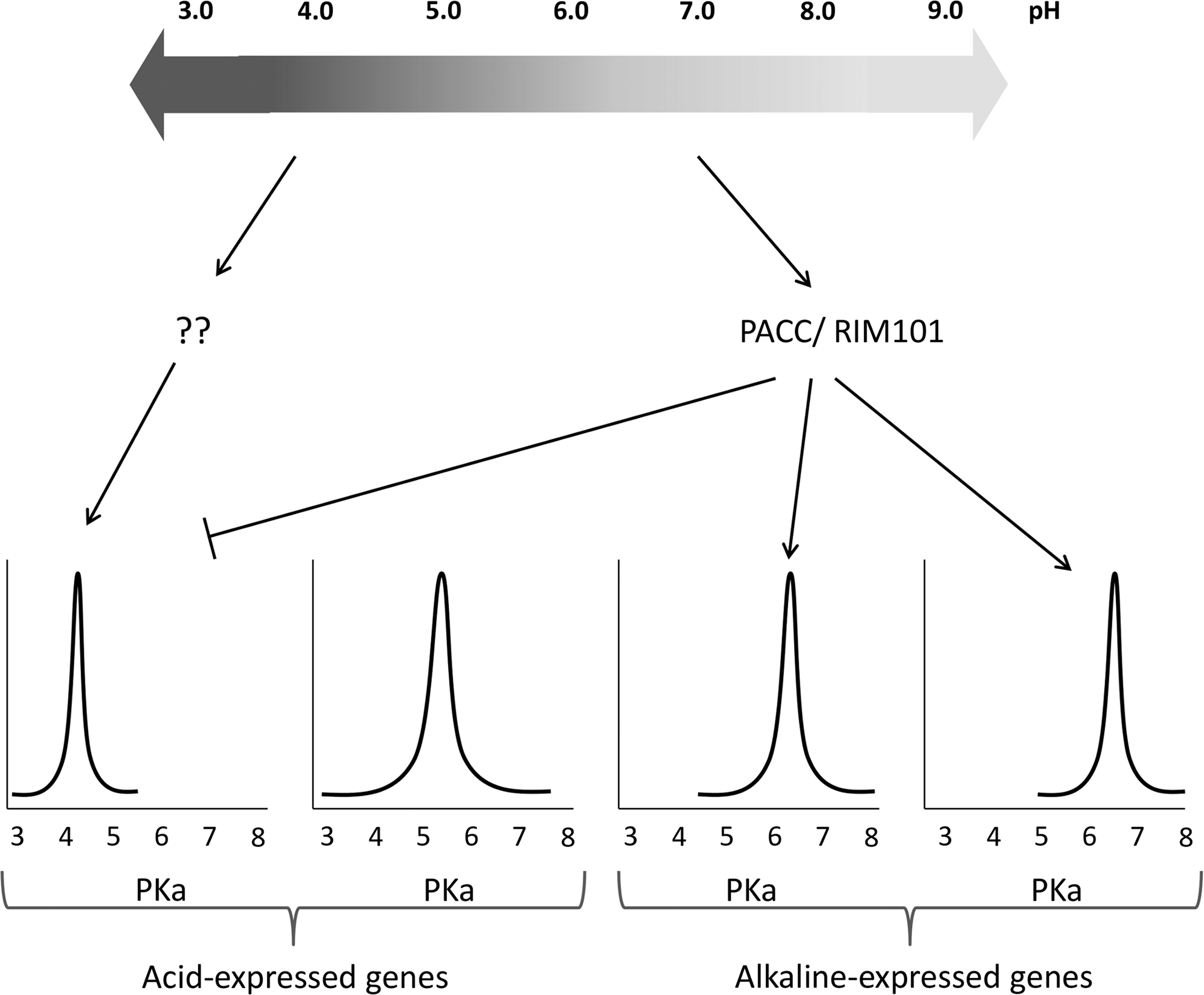
The Pal/Rim signaling pathway promotes, as a final signaling step, the proteolytic activation of a zinc finger transcription factor named PACC in filamentous fungi or RIM101 in yeasts (115). This factor activates the transcription of some genes at alkaline pH and represses others at acidic pH (51, 54, 100, 143). Regulation of those genes has been assumed to be correlated to their optimal protein activity (Fig. 3). The pathogen's ability to change the environment and activate signaling pathways enables it to control not only cell growth and development, but colonization as well. This pathogenicity depends on the pathogen's interaction with the host. Therefore, a given pathogen will behave differently toward different hosts, and vice versa.
Fungal factors that modulate the host environmental pH promote secretion of the arsenal of pH-specific pathogenicity factors (Fig. 1), which includes toxins, expression-regulating factors, genes that modulate host defense, specific transporters, redox-protective substances, cell wall-degrading enzymes and proteinases (24, 55, 138). Simultaneously, the secreted pH-modulating factors affect the host in a direct way as well (Fig. 2). This review discusses the effects of pH changes and of the factors that modulate pH on various processes in host–pathogen interaction.
Fungal pH Modulators Affecting Ambient pH and Pathogenicity
The extent of pH change in the plant tissue depends on host buffer capacity and initial pH (1, 76, 121, 123). The ability of the pathogen to alter the pH locally has been described initially for Sclerotinia sclerotiorum, but similar findings been have extended to some other pathogens, such as Colletotrichum spp., Alternaria alternata, Botrytis cinerea, Penicillium expansum, P. digitatum, P. italicum, Phomopsis mangiferae, and Fusarium oxysporum (39, 52, 53, 89, 100, 101, 123, 124, 129). To simplify the terminology in the following discussion, fungi that alkalinize the environment will be referred to as “alkaline fungi” and those that acidify it will be called “acidic fungi” (Fig. 1).
Alkaline Fungi
Ambient alkalization by fungi is achieved by active secretion of ammonia. Ammonia production by fungi seems to result from activation of proteases, followed by deamination of amino acids, which leads to ambient alkalization (69) (Fig. 3). Ammonium accumulation has been detected in association with pathogenicity of many Colletotrichum species, including C. gloeosporioides, C. acutatum, C. higgisianum, C. graminicola, and C. coccodes (1, 46, 100, 110); Alternaria alternata (52, 53); and Fusarium oxysporum (101). The ammonium secreted by Colletotrichum spp., F. oxysporum and A. alternata result in alkalization of the host tissue, and its concentration can reach approximately 5 mM, as found in decayed avocado, tomato, and persimmon fruits (1, 52, 53, 100). In every case, increased ammonium accumulation has been related to enhanced pathogenicity of Colletotrichum spp. (1, 2, 100). Recent findings that F. oxysporum, C. graminicola, and C. higginsianum secrete ammonia (101, 110) suggest that various fungal pathogens modulate the surrounding pH to create an optimal pH for expression and activity of their pathogenicity factors.
In the case of A. alternata, ammonia accumulation lead to pH increase by 2.4 pH units in several hosts, such as tomato, pepper, melon, and cherry (52, 53). Interestingly, ammonia accumulation and pH increase were not correlated across host species, suggesting that pH increase in each host depended on a complex interaction that involved the buffer capacity of the tissue, nitrogen availability, and fruit initial pH (53). Indeed, fruits differ in their buffer capacity and pH (52, 53), and low pH was found to activate ammonia production and secretion in Colletotrichum spp. (1, 76).
Acidic Fungi
Other pathogenic fungi, such as P. expansum, P. digitatum, P. italicum (124), P. mangiferae (39), Aspergillus niger (130), B. cinerea (89), and S. sclerotiorum (13) use tissue acidification in their attack. Tissue acidification is enhanced by secretion of organic acids and/or H+ excretion. S. sclerotiorum and Botrytis decrease the host pH by secreting significant amounts of oxalic acid (OA) (89, 129), gluconic acid is secreted by Phomopsis mangiferae (39), and combinations of gluconic and citric acids are mainly secreted by Penicillium (124) and Aspergillus (130). Penicillium expansum acidifies the tissue to pH levels of 3.5 to 4.0, at which polygalacturonase (pg1) transcription was found to be significantly enhanced (124). Acidic pH-specific expression of other members of the PG family was found in S. sclerotiorum (129) and in B. cinerea (157).
In Penicillium expansum, the gene encoding glucose oxidase (GOX2) is responsible for accumulation of gluconic acid; and functional analysis of the goxRNAi mutants showed that the differential level of gluconic acid production affected the pathogen's interaction with the host (11). Barad and coworkers (11) showed that downregulation of GOX2 impaired the pathogen's ability to produce gluconic acid and its capability to infect apple fruit. A second approach to functional analysis of the contribution of GOX2 to pathogenicity used the heterologous overexpression of GOX2 in the nondeciduous-fruit pathogen, P. chrysogenum. In this case, overexpression of GOX2 in P. chrysogenum enhanced gluconic acid production, sporulation, and surface colonization of pear fruit, but fruit colonization was still limited. These results indicate that although the observed increase in gluconic acid production is a factor that promotes surface colonization and sporulation, complete pathogenicity still requires the expression of other pathogenicity factors. Similarly, pathogenicity of oxalate-deficient mutants of S. sclerotiorum was lost on all host plants, whereas revertant strains that regained their oxalate biosynthesis capacity exhibited normal virulence (62). Further, in Botrytis cinerea pathogenicity was correlated with the amounts of OA produced by various strains infecting leaves of bean and grapevine (70, 78).
The ability of F. oxysporum sp. phaseoli to alkalinize the environment by ammonia secretion (101) or to acidify the extracellular pH by secretion of fusaric acid (18, 25, 26, 40) indicates that a given pathogen can deploy various tools, and it may activate any of them, depending on the host conditions.
Ambient pH and Mechanism of Genes Activation
Expression of pH-regulated genes is mediated by the integrated functions of seven genes: PACC, PALA, PALB, PALC, PALF, PALH, and PALI (86, 115, 147) (Fig. 4). Loss of function in any of these seven genes leads to phenotypes that express genes whose expressions are predominantly detected or elevated under the acidic ambient growth conditions of the wild type (113, 147). There is evidence that all PAL proteins act in a concerted manner upstream to the transcription factor PACC, thereby providing both transduction and proteolytic processing (47, 113,115, 116). Under alkaline conditions, two proteolytic steps activate PACC72: first, PACC72 is cut to PACC53—this proteolysis is pH dependent, based on a primary interaction with PALA through two specific motifs found at the C-terminal region of PACC (153) and is presumably carried out by the protease PALB (43); second, independently of pH, PACC53 is converted to PACC27, which is transcriptionally active and is responsible for repression of acidic genes at alkaline pH (54) and for the activation of genes alkaline, including PACC itself (25, 26, 95, 113, 144) (Fig. 4). Under acidic ambient conditions, PACC remains cytosolic and inaccessible for proteolysis, but in response to pH signaling, the Pal apparatus activates PACC. In its active form PACC27 is translocated to the nucleus, where the zinc-finger region recognizes the consensus DNA site 5′-GCCARG (96, 147). PACC regulation has been most extensively studied in the genus Aspergillus, but it was also found in many other fungi (6, 117). However, few publications have described the whole amount of genes that might be regulated by PACC specifically during fungal pathogenicity (53, 159).

Ambient pH and Pathogenicity Factors
Host pH is an important parameter in aqueous environments, since it enhances or limits the expression and activities of the pathogenic arsenal of enzymes, and these in turn may determine the virulence. This pH regulation modulates the arsenal of proteins secreted by the attacking organisms (42, 107, 121). A change in the ambient pH during fungal attack has been suggested to be a critical factor in the expression of pathogenicity factors such as pectate lyase (PL) and endoglucanases in several host/pathogen interactions (53, 123, 124, 159). This finding has raised the question of how great is the importance of environmental pH regulation for fungal pathogenicity genes.
The expression of genes encoding enzymes that are secreted only at alkaline pH has been suggested to be a regulator of pathogenicity under alkaline environments. Analysis of the endoglucanase1 from A. alternata indicated maximal gene expression at pH levels higher than 6.0, that is, at values similar to those found in the decayed tissues in which maximal virulence is observed (53). In C. gloeosporioides, PELB (encoded PL) is expressed and secreted at pH levels higher than 5.7, also similar to those present in decaying tissue (122, 159, 160). In C. gloeosporioides, PELB expression during pH alkalization followed a pattern similar to that of PAC1, suggesting the presence of a regulatory mechanism for control of secreted enzymes (48). Miyara et al. (99) used a ΔpacC mutant to demonstrate that genes encoding for pectolytic enzymes under alkaline conditions were activated by PACC in C. gloeosporioides. Similarly, for acidic environments, both pg1 and pg5 expression of F. oxysporum were controlled by acidic pH (25, 26), and the sequence corresponding to the PACC consensus recognition site was found in pg1; moreover, in F. oxysporum a ΔpacC mutant exhibited significantly decreased virulence at acidic pH, suggesting that PACC is a negative regulator of genes important for fungal attack under acidic conditions. The endoPG family of B. cinerea was differentially expressed under differing pH values, but although evidence for a PACC homolog has been found, there is no report of PACC in B. cinerea as a regulator for genes that affect fungal pathogenicity. In another acidifying pathogen, P. expansum, analysis of the transcript levels of pepg1 showed the highest accumulation at pH 4.0 and only minor expression at pH values higher than 5 (124). Also in S. sclerotiorum, the endoPg, pg1, displayed acidic-pH-specific expression. Transcription of the PACC homolog, pac1, declined during acidification concomitantly with an increase in pg1 expression, which suggests the existence of a complex virulence mechanism that controls PG expression under acidic conditions.
Other putative virulence factors, such as laccase, and protease responded in a pH-regulatory manner and were mainly expressed at acidic pH values of 3.1–6.0, which are close to those at the infection site of Botrytis (89, 102, 103). Presence of protease inhibitors may modulate pathogenicity by inhibition of ammonia production; it was shown that the presence of proteases inhibited Mycosphaerella, Fusarium, Botrytis, and Alternaria pathogenicity (151, 161, 162), which implies that these proteases may indirectly modulate virulence by modulation of ambient pH (25, 26, 89, 118, 119, 123).
In an attempt to genetically rationalize the existence of such large gene families of endoPGs in B. cinerea (149, 156) and in S. sclerotiorum (88, 129), and of glucanases in A. alternata (52), it was postulated that large gene families of cell wall-degrading enzymes could be involved in adapting the pathogens to their wide host range and to a wide range of pHs and environmental conditions. Further, increasing evidence for the differential expression of each family member under varied pH conditions illustrates the ability of a fungus to fine-tune its pathogenicity to the prevailing environmental conditions (52, 53, 89). During the study of differential expression of endoPG by B. cinerea in various plant tissues, Have et al. (146) selected two fruits—apple and courgette—with low and neutral pH, respectively. The regulation of Bcpg2 is negatively modulated by low ambient pH (157), which might explain its lack of expression in apples, whereas Bcpg3 expression is induced at low pH in liquid cultures (157), as in apples. In addition, Manteau et al. (89), in examining the resolution between two B. cinerea isolates—630 from grapevine (approx. pH 3.5) and T8 from tomato (approx. pH 6.0)—for expression of putative virulence factors found that isolate T8 displayed a more pronounced PG synthesis at the tomato pH, whereas 630 secreted more laccase. This fine-tuning of enzyme induction and secretion in response to the ambient pH, not only between hosts but even between isolates, further demonstrate the importance of the specific regulatory system controlled by environmental pH.
Ambient pH and Protein Secretion, Activity and Pathogenicity
Ambient pH may modulate fungal pathogenicity by activating transcriptional and post-transcriptional processes. PL expression of C. gloeosporioides began at pH 5.0 and increased up to pH 5.7, but secretion in culture was detected only when the pH reached 5.8, and the level of secretion increased with alkalization up to pH 6.5. This shows that host pH differentially regulates the expression and secretion of PL (48). Metarhizium anisopliae alkalinized the pH through ammonia accumulation and enhanced protease activity, demonstrating that proteases also are pathogenicity factors induced at alkaline pH during colonization of insects (140, 141).
PACC transcription factor modulated a group of pathogenicity factors expressed under alkaline condition (100). Development of PAL mutants that prevented the activation of PACC in Ustigalo maydis resulted in incapacity to secrete extracellular proteases (5, 27) and a triacylglycerol lipase (59). In C. gloeosporioides, development of a Δpac1 loss-of-function mutant attenuated virulence by 72% (99), whereas the ΔpelB mutant elicited 40% decreases in virulence (160), which suggests that deletion of PAC1 adds about 32% more to the downregulation of the expression of other gene(s) that contribute to fungal colonization.
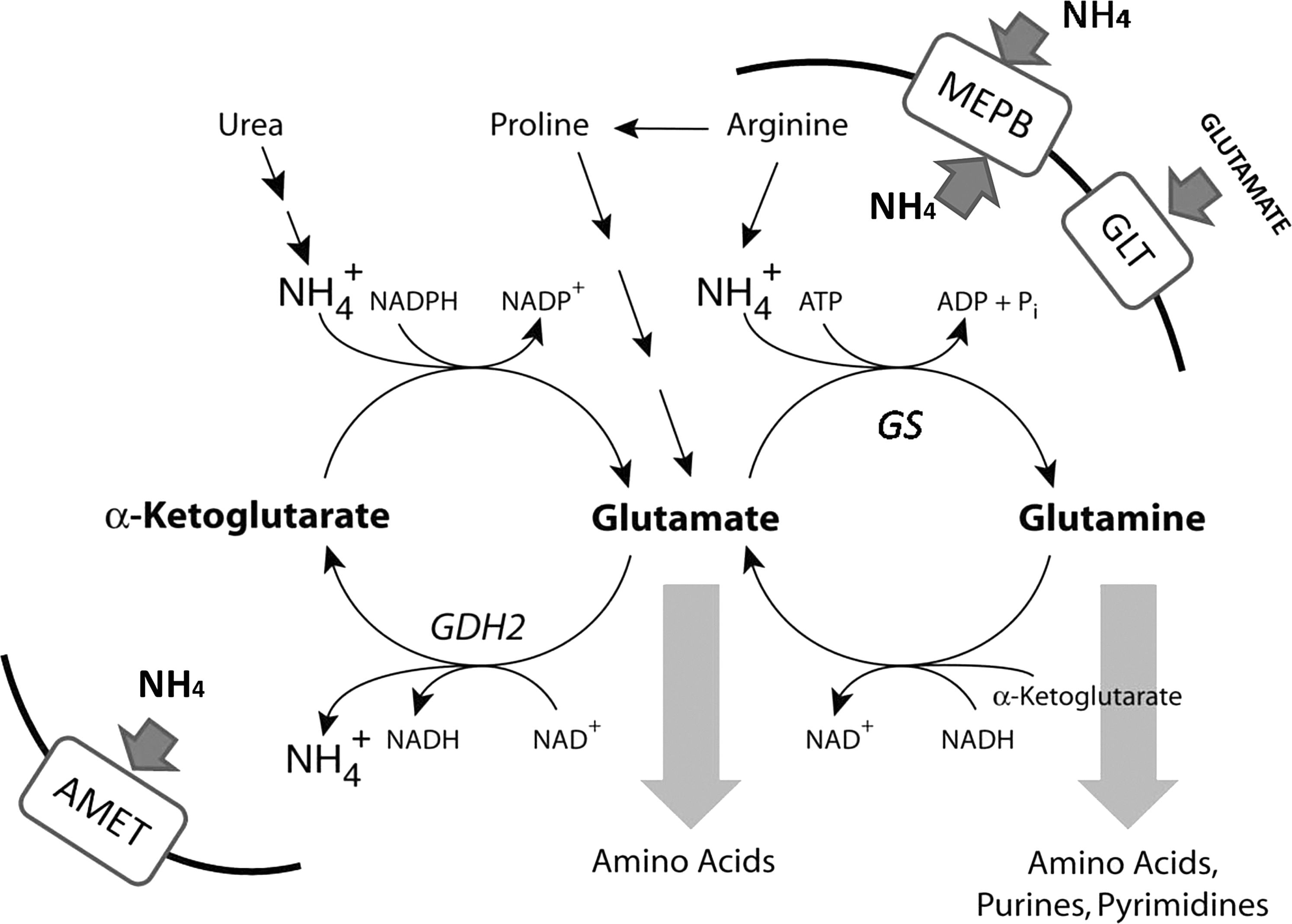
Analysis of genes regulated under alkaline conditions, by means of C. gloeosporioides cDNA macroarrays, showed regulation of eight genes related to nitrogen and ammonia metabolism, seven genes related to carbon metabolism, two genes related to calcium signaling, the glycosyl hydrolase DFG5 gene that is a sensor for alkaline pH damage in yeast (139), and others (100). Analysis of the transcript activation of nitrogen-metabolism genes showed differential regulation in the Δpac1 loss-of-function mutant: the Δpac1 strain exhibited transcriptional upregulation of glutamate dehydrogenase GDH2 (the acid-expressed gene) and downregulation of glutamine synthase GS1, glutamate transporter GLT, and ammonia permease MEPB (the alkaline-expressed genes) (100) (Fig. 5), which confirmed that ammonia accumulation during C. gloeosporioides pathogenicity is pH dependent, as previously observed for Aspergillus nidulans (21, 117). The full range of genes and processes regulated by PAC1 in alkalinizing pathogens is still unknown and will be the basis for future studies.
Ambient pH and Its Involvement in Intra- and Extracellular Transport
In yeast, Saccharomyces cerevisiae, RIM101 (the homolog of fungal PACC) promotes alkaline growth by repressing the expression of the NRG1 gene (Negative Regulator of Glucose). This NRG1 is a transcription factor that inhibits the expression of ion transporters, such as the Na+-ATPase transporter, that are important for maintaining ion homeostasis during alkalization (81, 82). Along with promoting growth in alkaline pH, RIM101 activity is involved in invasive growth, sporulation, and ion homeostasis (81, 82). It is likely that, in response to altered metal ion availability at alkaline pH, RIM101 also induces the expression of genes encoding an iron transporter, iron permease (SIT1 and CFT1), and a copper transporter (CTR4) (112). In Aspergillus spp. and other filamentous fungi, PACC family members also have been related to cation stress response. PACC in Aspergillus is involved in regulation of iron uptake and intracellular homeostasis, by directly binding to the promoters of siderophores (50), zinc uptake under various ambient pH levels, by altering the regulation of transporter proteins (4, 152). Salt-stress response in Fusarium oxysporum was shown to be mediated by PACC, and a direct role in the regulation of a sodium ATPase (ENA1) transporter was demonstrated (26). In A. nidulans loss of function mutations in PACC and PAL genes cause sensitivity to elevated concentrations of molybdate anion, which is a characteristic phenotype used to classify mutant alleles (22, 147). Molybdate acts as a cofactor for various intracellular enzymes such as the nitrate reductase and other pathways involved in phosphate and nitrogen metabolism (7, 8). Isolation of molybdate-resistant suppressors of a pacC- allele enabled the identification of HALA, a serine/threonine kinase homolog to yeast HAL4/5 kinases involved in saline tolerance and regulation of potassium uptake across the plasma membrane (56, 58, 104), suggesting their importance for maintain ion homeostasis.
Moreover, not only transport across the plasma membrane, but also intracellular trafficking might be regulated by ambient pH. Endocytic machinery is fundamental to correct PACC processing; elements of the Pal pathway interact with or are closely related to vacuolar protein sorting (VPS), and members of endosomal sorting complex required for transport (ESCRT) complexes in either Aspergillus or yeasts such as Yarrowia lipolytica and Candida albicans; in the latter, these pathways are important for growth and virulence at alkaline pH (14, 35, 60, 126, 141). In addition to PACC, tolerance to alkaline ambient pH requires the activity of two major transcriptional factors: CRZA and SLTA (138). CRZA is the homolog of S. cerevisiae CRZ1p and mediates response to alkaline pH. Salt stress is mediated, in turn, by the activity of calcineurin protein phosphatase (94, 142). CRZ homologs are found in diverse fungi, (33, 138), which are required to induce tolerance to both monovalent and divalent cations, (23, 58, 138). SLTA modulates the content, distribution, and size of vacuoles in A. nidulans (23, 58). In summary, a concerted action of PACC, CRZA, and SLTA homologs in fungi contributes to the intracellular organization, and thereby elaborates the appropriate response to environmental alkalinization, which contributes, most probably, to endocytosis of membrane receptors and secretion of the extracellular metabolites and enzymes needed for proper growth or virulence.
Modulators of Ambient pH and Their Effect on Morphogenesis
OA and sclerotia formation
OA is essential for S. sclerotiorum pathogenicity; mutants deficient in OA production were nonpathogenic (44, 62). Interestingly, such OA-deficient mutants also were unable to develop sclerotia, and this phenotype was mitigated by OA addition (62, 128). Culture pH has been shown to significantly influence OA biosynthesis and sclerotial development (129) (Fig. 2). Neutral or alkaline pH rapidly increased OA accumulation but suppressed sclerotial development, but OA accumulation reduced the ambient pH and created conditions that favored sclerotial development (129). Further, use of the Δpac1 mutant confirmed the pH-mediated regulation of OA production, sclerotial development, and pathogenesis (127).
What are the mechanisms that modulate differentiation, sexual development, pathogenicity, and monitoring of environmental stresses in S. sclerotiorum? Rollins and Dickman (128) found that cyclic adenosine monophosphate (cAMP) played a role in regulating the transition between mycelial growth and sclerotial development: increases in endogenous or exogenous cAMP levels inhibited sclerotial development while substantially elevating OA accumulation. Several mitogen-activated protein kinases (MAPK) signaling cascades were found to function in coordination with other pathways, such as cAMP/PKA and pH-dependent pathways in fungi (e.g., S. cerevisiae, Schizosaccharomyces pombe, Candida albicans, and F. oxysporum) during development and during response to environmental stresses (26, 41, 77, 84) (Fig. 2). In Sclerotinia, it was found that cAMP-mediated signaling interacted with or functioned in parallel with pH signaling, to regulate OA levels and sclerotial development. This conserved MAPK homolog regulated sclerotial development through interconnections with pH-dependent and cAMP-dependent pathways (30).
Ammonia and appressoria formation
C. gloeosporioides secretes large amounts of ammonia at the initial stage of spore germination (100) originating by degradation of nitrogen sources such as glutamate (64, 135). Ammonia secretion in germinating spores is very important for the fungus; usually few soluble nutrients are available on the host surface (66, 145). Ammonia accumulation also was found to activate fungal appressorium formation and differentiation: (i) appressorium formation was accompanied by a 200-fold increase in GDH2 expression and ammonia accumulation (200 mM per gram of dry weight); (ii) reduction of ammonia levels by spore dilution reduced appressorium formation, and exogenous addition of ammonia restored it; (iii) Δghd2 mutant strains showed reduced appressorium formation compared with the WT, and exogenous addition of ammonia restored appressorium formation; and (iv) C. gloeosporioides WT and Δghd2 strains showed no differential colonization of the fruit mesocarp, whose penetration does not require appressoria and where nutrients are readily available in the fruit flesh, but they strongly differed in pathogenicity on the fruit peel, where appressoria were induced for fruit penetration (100). This indicates that the effect of ammonia on germ-tube differentiation to appressoria is primarily the result of a nutrient signal that activates the nitrogen metabolism. In cases in which readily available nutrients, such as glutamine, are present the fungus will colonize the host by using mycelia, without appressoria (12, 66). Δghd2 strains of Magnaporthe grisea also were reported to affect appressorium formation, but in that case, ammonia was not considered a key factor in their formation (111). In S. cerevisiae, ammonia release also affected morphological development, and controlled the transition from budding colonies to the filamentous phase (114, 165), which suggests that, with regard to fungal structural differentiation, ammonia production is important in itself, and not only as a pH modulator (Fig. 2).
pH Modulators and Host Response
Ambient pH and each of the secreted pH modulators (ammonia and OA) have multiple effects on the plant host and thereby affect fungal pathogenicity.
Host Response to Ambient pH
Changes in the pH of the host ambient medium have been shown to strongly affect the apoplastic pH, but not the cytosolic pH (61). The pH changes in various plant-cell compartments that result from ion effluxes seem to play important roles in the regulation of reactive oxygen species (ROS) production and plant defense responses. For example, an oxidative burst was rapidly elicited simply by transferring bean cells into a medium buffered at higher pH than the former one (16, 155). Medium (or apoplast) alkalization induced respiratory burst oxidase homolog (RBOH) activation, probably as a result of induced depolarization of the plasma membrane and subsequent K+/H+ exchange, followed by Ca2+ influx/Cl2 efflux (109, 137, 163). Transient extracellular alkalinization is an essential factor in induction of the signal transduction pathway during induction of the defense response and plant programmed cell death (PCD) (34, 63, 132, 134). In this connection, changes in external pH rapidly alter plant gene expression and modulate host responses, similarly to elicitors (79). Studies of plant responses to bacterial and fungal pathogens that impose those variations in external pH could cause substantial changes in host gene expression and defense response (6, 85). Also in mammalian systems, pH changes have been hypothesized to modulate the oxidative burst of phagocytotic cells (136), which suggests that there is a global ROS response to ambient pH changes that could activate several physiological and defense response processes in hosts.
Host Response to Ammonia
Ammonia, as a pH modulator, could itself affect host responses: (i) direct toxic effects can spring from plant nutrition with ammonium instead of nitrate; and (ii) accumulation of ammonia secreted by various necrotrophic fungi during ambient alkalinization could elicit a host response.
Ammonium has toxic pleiotropic effects on plants when it is used as a sole source of nitrogen. The main phenotypes are: leaf chlorosis, growth suppression, yield depression, decrease of root-to-shoot ratio, and inhibition of seed germination (20). The mechanisms of induced ammonia toxicity include activation of plant-membrane transport, activation of plasma membrane ATPase, elevation of membrane flux, change in the concentration of cytosolic protons, disruption of biochemical pH-stat, and induction of ethylene synthesis (20, 92, 164)
Environmental alkalization during the colonization by Colletotrichum sp. secreted a significant amount of ammonia (1, 110, 158, 159) and led to ROS activation in a calcium- and pH-dependent manner, with a maximum activation at pH 7 to 8 (2). Plant ROS are known to play an important role in eliciting a wide range of defense mechanisms (10, 38, 57, 65). In plants, the NADPH oxidase homologs have been termed RBOH; they are the main factor in modulating ROS production in response to pathogens (2, 3, 131, 148, 165). This indicates that ammonia accumulation by the pathogen modulates the host response during necrotrophic colonization. Ammonia accumulation also induced ion leakage, DNA laddering, deformation of nuclei, and terminal dutp nick end labeling (TUNEL) stain, all of which indicated its involvement in plant PCD (2, 3) (Fig. 6). Consequently, the tomato rboh mutant showed significantly diminished production of ROS in response to either Colletotrichum colonization or ammonia application, and showed increased resistance to pathogen attack (2).
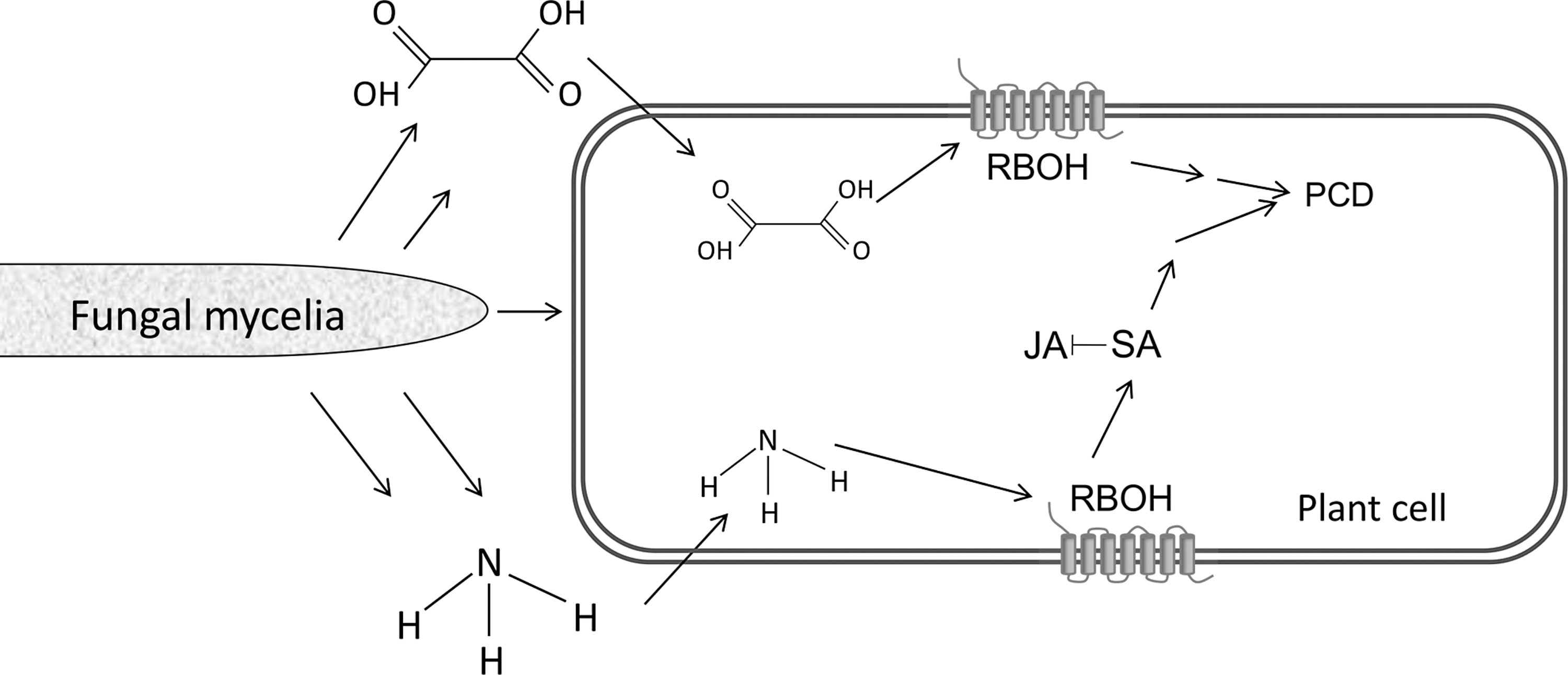
The RBOH activation and ROS accumulation at the infection site also led to activation of host salicylic acid-dependent genes (2, 3). These findings suggest that ammonification at the edge of the infection site deregulates host responses and enhances PCD, which, in turn, would facilitate activation of fungal necrotrophic pathogenicity (Fig. 6). Indeed, C. coccodes mutants with reduced ammonia production exhibited diminished pathogenicity (1, 2, 100), and tomato rboh mutants, which displayed less PCD, were less susceptible than the WT tomato to Colletotrichum necrotrophic attack (2). Effects of ammonia were observed not only in plant–fungal interactions: Coccidioides immitis, a fungal pathogen of humans that causes respiratory disease, is known to alkalinize its environment with ammonia (36, 73, 97); mutants with compromised ammonia secretion were less pathogenic (98), and ammonia was shown to compromise the rat-host immune response and cause inflammation (67). Further, in rat astrocytes and brain edema ammonia was shown to activate NADPH oxidase, which was followed by autophagy (17, 31, 68, 71, 74, 75, 105, 125, 133), which suggests that ammonia possesses a global role of action on NADPH oxidase and cell death.
Host Response to OA
OA secretion by S. sclerotiorum is essential for pathogenicity and is remarkably multifunctional (15, 28, 49, 62, 72, 90, 93, 150, 154) (Fig. 2). OA affects host responses through numerous physiological processes, such as ambient acidification; strong Ca2+ chelation, which weakens the plant cell wall and compromises the host defense responses (28, 37, 91); guard cell regulation; vascular plugging with oxalate crystals (49); and inhibiting the host callose-deposition response to the necrotrophic infection (154) (Fig. 2).
OA also indirectly aids Sclerotinia pathogenicity by acting as a signaling molecule, via complex manipulation of host ROS (28, 72). Oxalate lowers the pH and inhibits the oxidative burst of the host plant at pH 3 to 4, and thereby compromises plant defense responses (28). Recently, OA was shown to generate reducing conditions in the cell, in correlation with the inhibition of the host oxidative burst and other defense responses (154); OA-deficient mutants were unable to induce reducing conditions and thereby to inhibit the oxidative burst, and therefore were unable to cause disease. However, addition of dithiothreitol reducing agent elicited transient reducing conditions that suppressed the oxidative burst, caused reversion to the OA-deficient mutant phenotype, and restored pathogenicity (154). The mechanisms by which OA triggers redox reduction and modulates plant defense pathways probably affect both host and pathogen. In contrast, OA also induced plant ROS production that correlated with a pH increase of 5 to 6, that later induced ethylene production, anion channel activation, DNA laddering, and a TUNEL reaction, all of which indicated host PCD in response to OA (51, 72) (Fig. 6). DPI inhibited ROS production and cell death (72), and NADPH oxidase activity was elevated in response to OA (87), which suggests involvement of an NADPH oxidase-mediated ROS during the fungally induced PCD. This host PCD is necessary for necrotrophic colonization. When the host PCD was inhibited by expression of mammalian apoptosis inhibitors in tobacco plants the transformed plants were protected from infection by the necrotrophic Sclerotinia (30, 45). Thus, organic acids may have a dual role in ROS regulation, because such regulation may downregulate and also activate the plant oxidative burst, possibly in a pH-dependent fashion, to fit its necrotrophic development (28, 72, 154).
Conclusions
During fungal interactions with plant hosts, several fungi acidify or alkalinize their ambient pH by secreting organic acids or ammonia, respectively. These fungi regulate an arsenal of enzymes, each in accordance with the environmental pH and the enzyme's requirements for optimal activity, to increase fungal pathogenicity (Fig. 1). This arsenal includes genes and processes that compromise host defenses, provide metabolites that affect cytoskeletal structure, contribute to intracellular signaling, produce cell wall-degrading enzymes, regulate specific transporters, induce redox protectant systems, and generate factors needed by the pathogen to effectively cope with the hostile environment found within the host. Further, evidence is accumulating that OA and ammonia are multifunctional (Fig. 2) in that these molecules change the ambient pH, activate virulence factors, and simultaneously hijack the plant response and induce PCD (Fig. 6) to further enhance their necrotrophic attack.
Footnotes
Acknowledgment
We acknowledge the support of the US-Israel Binational Agricultural Research and Development Fund (BARD) during several stages of our work.