
Abstract
Introduction
CYB561 members are implicated in physiological processes, including plant ‘front-line’ (apoplastic) defense; iron uptake; ASC regeneration in neuroendocrine tissues; and tumor growth. CYB561s are also of interest because of their particular biochemical (structure-function) properties. They are composed of six TM α-helices, the central four of which make up the ‘CYB561-core,’ coordinating two heme b molecules with four conserved His. The core domain harbors the intra-molecular electron transfer, and, intriguingly, is a building block in other proteins, often in association with other redox domains such as the N-terminal domain of dopamine β-monooxygenase (DOMON). Together, the CYB561 protein family expands our horizon on the role of ASC in cell metabolism.
Physiological Functions and Biological Relevance
ASC recycling
Some CYB561s support ASC regeneration, through one-electron reduction of partially oxidized ASC, monodehydroascorbate (MDHA) to ASC, using electrons provided by ASC, on ‘the other’ side of the membrane. In animals, such TMET activity is catalyzed by the chromaffin granule CYB561 (CGCytb/CYB561A1, for systematic nomenclature, see Supplementary Table S1; Supplementary Data are available online at

Work on plant membranes revealed the presence of ASC-reducible, di-heme-like b-type cytochromes, with similar biophysical properties as bovine CGCytb/CYB561A1, in several species (4, 5, 22). A Phaseolus vulgaris CYB561, presumably plasma membrane (PM)-located, mediated TMET in purified, ASC-loaded, vesicles (2). Extracellular ASC provides antioxidant defense against gaseous oxidants (ozone, NOx), and CYB561 could be involved in apoplastic ASC regeneration (Fig. 1E). The Arabidopsis genome reveals four genes encoding six-TM CYB561s with an average of around 30% sequence identity to CGCytb/CYB561A1 (3), but it is not yet clear which of the gene products is involved in PM TMET.
Fe3+-reductase activity and iron homeostasis
A new CYB561 member was identified in the cell membrane of mouse duodenum epithelial cells, in response to hypoxia and iron deficiency (39). Mouse duodenal DCytb/CYB561A2 reduces intestinal Fe3+ before uptake (Fig. 1B). ASC is an electron donor to mouse DCytb/CYB561A2, at least in vitro (54). The elevated expression of DCytb/CYB561A2 in mice with systemic iron overload (hemochromatosis) provides additional support for a role in iron uptake (23).
ASC-mediated TM Fe3+-reductase activity was confirmed by complementation of Δfre1Δfre2 Fe3+-reductase-deficient yeast lines. Reduction of extracellular Fe3+-chelates in transformed yeast cells, expressing genes for three mouse and one Arabidopsis CYB561 protein (CGCytb/CYB561A1, DCytb/CYB561A2; LCytb/CYB561A3; TCytb/CYB561B1, respectively), was demonstrated (10, 54). For CGCytb/CYB561A1, the TM Fe3+-reductase activity was also demonstrated in ASC-loaded proteo-liposomes (11). These experiments suggest that, at least in vitro, all CYB561s may have Fe3+-reductase activity. Therefore, the actual in vivo activity of CYB561s, that is, Fe3+ reduction versus MDHA reduction, is probably dependent on substrate availability and biological context.
Free Fe3+ would readily be reduced by ASC, if occurring in the same compartment, and Fe2+ catalyzes free radical generation through Fenton-type reactions. Separation of Fe3+ and ASC, along with CYB561 catalyzing TMET, thus provides efficient control over ASC-mediated Fe3+ reduction.
Arabidopsis TCytb/CYB561B1, used in the yeast Fe3+-reduction assays, is localized in the tonoplast (20, 50) (Fig. 1D). Given that the Arabidopsis TCytb/CYB561B1 ASC-binding (electron acceptor) site is predicted on the cytoplasmic protein side, this suggests that this CYB561 could reduce Fe3+ in the vacuolar lumen, supporting transport to the cytoplasm. Vacuolar Fe2+-transporters (NRAMP3, NRAMP4), which may act in conjunction with TCytb/CYB561B1, have been identified (27). Possibly plant vacuoles store iron for remobilization when metabolic needs exceed supply. ASC-dependent CYB561-mediated Fe3+-reductase activity in plants may complement the well-characterized NADH-dependent activity of the FRO proteins (27).
Other physiological functions
In addition to a role in ASC and Fe metabolism, CYB561 members may be involved in other physiological processes. A watermelon PM CYB561B is up-regulated under drought and high-light stress and was proposed to constitute a route for thermal dissipation of excess light energy by functional interaction with apoplastic ASC oxidase (43) (Fig. 1E). In humans, a CYB561 (TSCytb/CYB561D2/101F6) is encoded in chromosomal region 3p21.3 and is related to suppression of tumor cell growth. Treating lung cancer cells with TSCytb/CYB561D2 plasmid-DNA nanoparticles and ASC, synergistically suppressed lung cancer cell development (46). For yet other CYB561s, limited functional information is still available. For example, mouse LCytb/CYB561A3 is located in lysosomal membranes in various tissues and is possibly associated specifically with macrophages (64) (Fig. 1C). However, its physiological impact is not yet clear. Lysosomal iron pools play a role in oxidative stress control (30), which could involve LCytb/CYB561A3-Fe3+-reductase activity. Finally, one should consider that other electron donors and acceptors (than ASC and MDHA/Fe3+) may still be identified, linking CYB561s to yet other physiological processes.
Tissue- and subcellular-distribution
CGCytb/CYB561A1 was first identified in catecholamine-synthesizing (chromaffin) vesicles, and was, consistently, demonstrated in various neuroendocrine tissues (53). It is now clear that plant and animal CYB561s occur in a wide range of organs, tissues, and cell types. Remarkably specific cell-type localizations are sometimes observed, but not yet understood, such as testicular Sertoli cells for mouse LCytb/CYB561A3 (64). Some animal organs (e.g., lung, spleen, brain, and duodenum) contain multiple CYB561 isoforms. However, in these cases, the proteins are apparently not located in identical cell types or membrane types. In Arabidopsis, CYB561 isoforms show some, but not complete, overlapping organ distribution patterns, and CYB561s occur in different membrane types (52, 62). Therefore, CYB561 isoform expression does not appear to be redundant. In a recently isolated Arabidopsis homozygous KO of TCytb/CYB561B1, transcription of at least three other CYB561 isoforms was not enhanced, supporting this notion (unpublished).
The subcellular membranes containing CYB561 consistently are membranes separating the cytoplasm and an acidic compartment, including acidic extracellular environments in plants and animals (Fig. 1). This pH gradient is most likely maintained by the activity of H+-transporting ATPases, and is probably important in CYB561 TMET function.
CYB561 Structure and Structure-Function Relationship
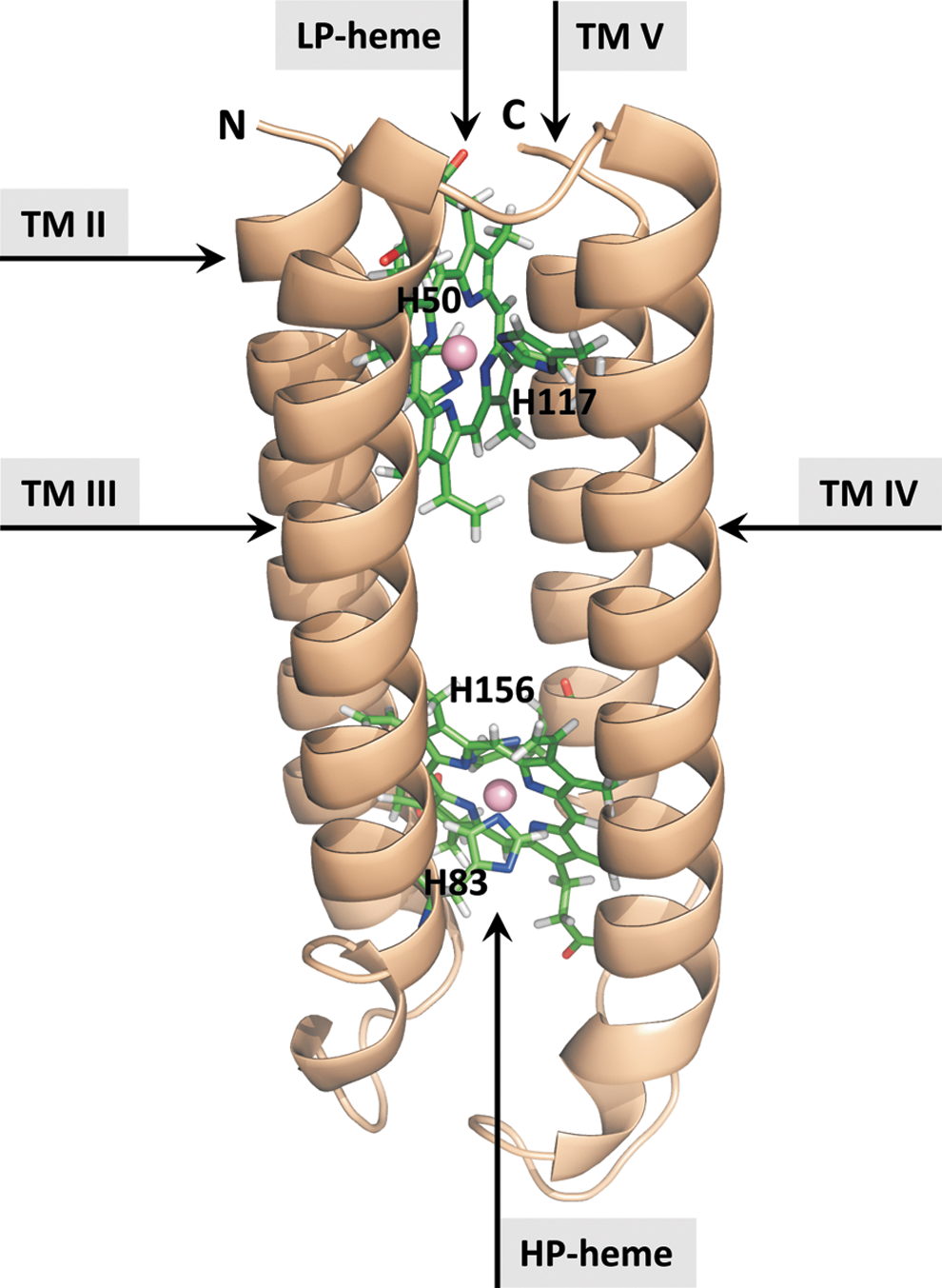
The CYB561 protein family, in plants and animals, was identified on the basis of sequence comparison to the amino acid sequence of the bovine chromaffin granule CGCytb/CYB561A1 (59) (Fig. 2). Until today, this is also the only native CYB561 protein for which biochemical and biophysical measurements are available. Biochemical and structural information of other plant and animal CYB561s is derived from recombinant proteins, expressed in yeast (Saccharomyces cerevisiae, Pichia pastoris), insect (Sf9) cells, and Escherichia coli (8, 11, 15, 20, 33, 35 –37, 41, 45, 54). CYB561s are membrane-spanning proteins that are generally accepted to contain six TM α-helix domains and two pairs of His residues, arranged on four consecutive TM helices, coordinating two b-type hemes, located on each side of the membrane and accepting electrons from ASC (Figs. 2 and 3). Crystal-structure-level 3D atomic information is not (yet) available for any CYB561. However, 3D atomic resolution models were calculated for human CGCytb/CYB561A1, and Arabidopsis TCytb/CYB561B1 to support data analysis (6) (Fig. 4).


Midpoint redox potentials
Redox titration results consistently reveal the presence of two one-electron redox centers (b-type hemes), with midpoint redox potential (E0′) values ranging from +80 to +190 mV for the ‘high potential’ (HP) heme, and −20 to +60 mV for the ‘low potential’ (LP) heme, (15, 35, 56). Among CYB561 isoforms, there is a relatively consistent difference of about 100 mV between the E0′ of the heme centers, supporting intra-molecular electron transfer mechanisms. Recently, it was demonstrated by electron paramagnetic resonance (EPR) and site-directed mutagenesis that the HP-heme is located on the cytoplasmic side of the TM proteins (16).
ASC reducibility
Reducibility by ASC is recognized as a particular CYB561 feature, generally reaching reduction levels of 50% to 90% (relative to Na-dithionite [DTH]). Why 100% reduction levels are not reached, even at high ASC concentrations, is not yet fully understood, in part because the ASC and/or DTH reduction mechanism is not yet fully unraveled. The midpoint redox potentials of the electron transfer reactions involving ASC are around +320 mV (MDHA/ASC) and −200 mV [dehydroascorbate (DHA)/MDHA], and +60 mV for the DHA/ASC redox couple (25). Since the MDHA concentration in the cell is orders of magnitude lower than that of ASC (due to disproportionation), the electron transfer from ASC to CYB561 is favored under physiological conditions. Thermodynamic evidence supports the idea that CYB561-mediated TMET may be coupled to a local protonation/de-protonation cycle (42, 44).
Direct molecular-binding interactions between ASC and CYB561 have not been measured, but steady-state and fast kinetic measurements on plant and animal CYB561s suggest the existence of two distinct ASC-‘affinity’ sites, with approximate values of 0.01 mM and 1 mM, for CGCytb/CYB561A1 (7, 11, 28). These parameters are not real binding constants but rather characterize the interaction between ASC and CYB561, resulting in the reduction of the HP and LP-hemes. The effect of diethylpyrocarbonate on ASC reduction kinetics also supports this hypothesis (58). A comparison of conserved sequence regions in the CYB561 core led to the tentative assignment of two substrate binding sites located on the cytoplasmic and luminal side of the protein (47, 59) (Fig. 2).
Spectrum analysis of mouse and bovine CGCytb/CYB561A1 and of TSCytb/CYB561D2 revealed the presence of two distinct, split α-bands in the spectrum of ASC-reduced proteins, which is consistent with the presence of two heme pockets (9, 12, 28). In Arabidopsis TCytb/CYB561B1 and mouse TSCytb/CYB561D2, singular value decomposition analysis identified two distinct b-type heme spectra that could be assigned to the two CYB561 hemes (12, 16). The property of split α-bands points to the fact that each heme is located in an anisotropic electrostatic field, originating from the distribution of charged amino acid residues around the heme pockets. Given the limited primary structure similarity between CYB561 isoforms, it is not surprising that the fine structure of the split α-band slightly differs, not only for the two hemes but also between isoforms (9, 12, 15, 16).
Other reducing agents
Common reductants such as NAD(P)H and reduced glutathione (GSH) are poor electron donors to CYB561s. Reduction by NAD(P)H of various CYB561 isoforms never exceeded 5% (relative to DTH) (12, 57). GSH does not reduce CYB561, but other thiol reagents, in particular dihydrolipoic acid (DHLA), are almost as efficient as ASC in reducing mouse CGCytb/CYB561A1 and TSCytb/CYB561D2; however, with only one apparent, high affinity, binding site (12). In addition, the Arabidopsis TCytb/CYB561B1 was reduced by DHLA, although with much lower sensitivity and biphasic concentration dependence (12). Thus, it is possible that DHLA plays a different role in CYB561 reduction in plants and animals. In cells, DHLA is present mostly in its lipoamine form, tethered to conserved Lys residues on lipoyl-accepting protein domains (13). It may, therefore, be worth to investigate lipoyl-domain-containing proteins as potential CYB561 redox partners. In addition, dithiothreitol, and its stereoisomer, dithioerythritol, reduce Arabidopsis TCytb/CYB561B and mouse TSCytb/CYB561E (12, and unpublished).
Site-directed mutagenesis
Important structure-function information is derived from targeted mutations in the highly conserved heme-coordinating His residues of the CYB561-core (Fig. 2 and 3). For example, detailed EPR studies of Arabidopsis TCytb/CYB561B1 mutant proteins revealed that the HP-heme center is located on the cytosolic side of the protein (16), opposite to that previously suggested (56, 58). Western blot analysis and spectroscopy demonstrated that mutation of His residues coordinating the LP-heme (intra-vesicular-side) of Arabidopsis TCytb/CYB561B1, and mouse CGCytb/CYB561A1, resulted in nearly undetectable protein levels. In contrast, mutations in the HP-heme-coordinating residues hardly affected CYB561 expression, but resulted in altered ASC-reduction kinetics and reduced heme content (11, 28, 34). Somewhat contradicting results were obtained with His mutations in human DCytb/CYB561A2, and need further investigation (37, 45).
Positively charged residues near the hemes may facilitate the interaction with the ASC and MDHA anions, as is, for example, the case in the ASC-mediated reduction of soybean APX (14, 31). Lys and Arg residues are frequently found in the loop regions between the TM helices (Fig. 2), and an Arg residue (R72 in mouse CGCytb/CYB561A1), located near the HP-heme, is fully conserved among CYB561A members (59). Mutagenesis of R72 resulted in the loss of the ‘high affinity’ ASC reduction, supporting its role in substrate binding and suggesting that the high-affinity ASC-binding site was close to the HP heme (11). Arabidopsis TCytb/CYB561B1 contains no Arg in the vicinity of the heme, but still displays two distinct affinity sites (7). Its role is possibly replaced by a conserved Lys (K80) residue. This idea is supported by mutagenesis of K83 in maize CYB561B1, which resulted in altered midpoint redox potentials and ASC-reduction kinetics (41).
Mutations in other conserved and partially conserved residues of mouse CYB561s had variable effects on TMET activity and ASC reduction (34, 41, 45, 54). Consistent with the putative location of the electron-accepting and -donating sides, mutations in loop regions on the intra-vesicular side severely decreased LCytb/CYB561A3 TM Fe3+-reductase activity, but mutations on the cytoplasmic side had comparatively less effect (54).
A key function of CYB561s is TMET, through sequential reduction and oxidation of the HP and LP-hemes. Conserved residues presumably located in TM domains (e.g., Q131 in mouse LCytb/CYB561A3/) have been identified, and mutation resulted in strongly decreased Fe3+-reductase activity (54). This suggests that electron transfer my involve oxidation/reduction of intramolecular amino acids. On the other hand, electron transfer along chains of prosthetic groups is well established, as long as the “edge-to-edge” distance between groups is smaller than 14 Å (15, 48).
CYB561-Domain-Containing Proteins
CYBDOM
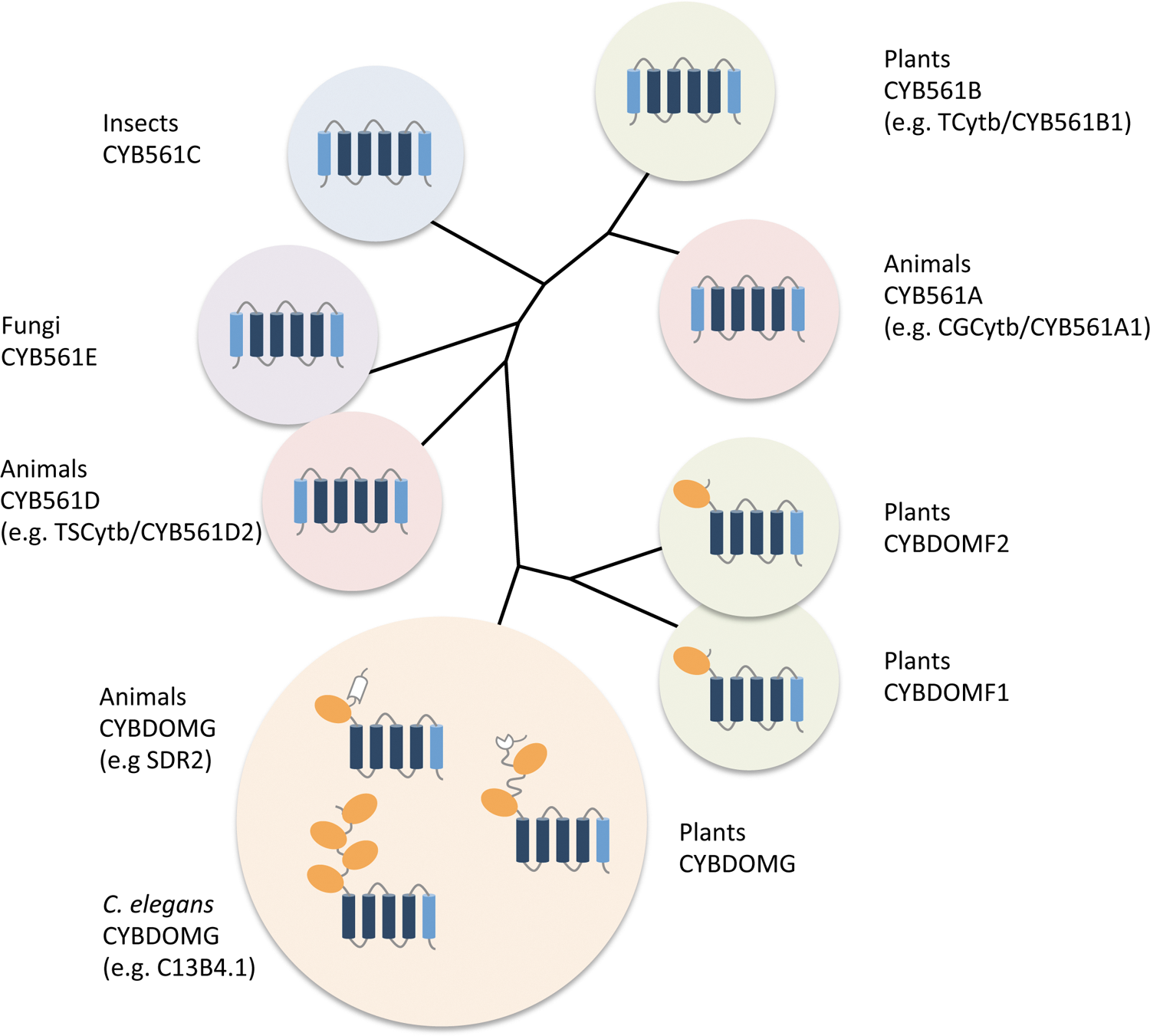
Identification of a CYB561-core domain at the C-terminus of a protein named stromal cell-derived receptor 2 (SDR2) provided evidence that CYB561s could be part of multi-domain proteins (49) (Fig. 5). Bio-informatic analyses demonstrated that most commonly a single CYB561-core follows a DOMON-domain (CYBDOM), but some proteins contain up to four DOMONs in a row (e.g., C13B4.1 of C. elegans; Fig. 5) (1, 26, 49, 51).
The DOMON-domain refers to the N-terminal domain of dopamine β-monooxygenase (or dopamine[beta]-hydroxylase, DBH); (1, 49), catalyzing the reduction of dopamine to norepinephrine in chromaffin granules. In the process, ASC is oxidized to MDHA, which may be regenerated by CYB561 (Fig. 1A). The DOMON-domain is widespread in nature, often in association with redox domains such as the Cu2+-monoxygenase domain in DBH or CYB561 in CYBDOMs, and/or with adhesion modules (e.g., reelin-domain in SDR2) (Fig. 5) (26). In DBH, this module is probably involved in tetramerisation (29), but not in catalysis or catecholamine binding (61).
CYBDOM sequences have been identified in the genome of animals, fungi, plants, and prokaryotes (26). Eukaryotic CYBDOMs were classified in two major groups. The ‘F’-group, including plant proteins with single CYB561- and DOMON-domains, and the ‘G’-group, including SDR2-related sequences (from animals or plants), sometimes with multiple DOMON domains (59) (Fig. 5). G-group CYBDOMs often also contain the reelin domain (animals) or DM13 domain (plants) (26, 51).
It is now clear that plants contain larger CYBDOM gene families than do other organisms (51, 59). For example, Arabidopsis contains at least ten CYBDOM-containing genes: eight belonging to the F-group, one to the G-group, and a divergent gene that could not easily be classified (51). In addition, Arabidopsis and other plant genomes contain CYBDOM-related genes with either no DOMON or no CYB561-domain (e.g., see section on heme-b binding). On the contrary, SDR2 is the only CYBDOM of the human genome, and single SDR2 orthologous genes are found in mouse and fly genomes, while C. elegans contains four G-group CYBDOMs (59).
All of these CYBDOMs contain a canonical CYB561-core, with four fully conserved His, at the N-terminus of four consecutive TM α-helices, plus an additional C-terminal α-helix, corresponding to the sixth α-helix of CYB561s (Fig. 5). In spite of these strictly conserved features, CYB561s and CYBDOMs have highly divergent sequences (59).
A common feature of CYBDOM sequences, at least in eukaryotes, is the presence of a signal sequence directing these proteins to the secretory pathway. Consistently, proteomic studies in plants have identified CYBDOM proteins on the PM (55), in some cases in association with lipid rafts (32). In tobacco roots expressing a chimeric CYBDOM-GFP from soybean, fluorescence was clearly associated with the PM (A. Costa and P. Trost, unpublished). In mammals, SDR2 is also believed to reside in the PM (61).
Topology predictions show that the C-terminus of CYBDOMs (corresponding to the C-terminus of CYB561s) is cytosolic and, consequently, the N-terminus of the five-α-helices CYB561-domain, connecting to the DOMON domain(s), is extracellular. Indeed, DOMON domains are commonly extracellular and either heme- or sugar-binding domains (26). The heme-binding DOMON domain of cellobiose dehydrogenase (CDH) of ligninolytic fungi plays a role in the attack to the lignocellulosic extracellular matrix of plants (63). Sugar-binding DOMON domains are also found in bacterial extracellular cellulases (26), overall suggesting a role of DOMON-containing proteins in modifying extracellular polymers (see section on Why CYBDOMs?).
Fe3+-reductase activity
The CYB561-core in CYBDOMs contains all necessary elements to catalyze TMET. Recombinant fly and mouse SDR2 demonstrate Fe3+-reductase activity in Xenopus oocytes (61). SDR2 is, therefore, considered a TM reductase and was renamed ferric chelate reductase 1 (e.g., in UniProt). Whether Fe3+-reductase activity can be extended to other CYBDOMs is unknown. Since the CYB561-domain can catalyze by itself the TM reduction of Fe3+-chelates, the role of the DOMON domain in SDR2 redox activity is uncertain. However, DOMON domains are most likely b-type cytochromes and may form a redox chain with CYB561 domains.
Heme-b binding in the DOMON domain
The DOMON-containing CDH of wood-degrading and phytopathogenic fungi is an extracellular flavo-cytochrome that participates in early events of lignocellulose degradation (38). The 3D structure of the DOMON-domain of CDH shows heme binding through an uncommon His-Met coordination (21) (Fig. 6). The heme is tightly fitted into a pocket of the concave inner β-sheet of the overall β-sandwich structure that is typical of the DOMON domain (26). Interestingly, both heme-axial ligands of Phanerochaete chrysosporium CDH (M65, H163) are conserved in CYBDOM sequences, strongly suggesting that DOMON-domains of CYBDOMs may also bind a heme b. These residues are conserved neither in the DOMON of DBH (that bind no hemes) nor in DOMON proteins that bind a sugar in place of the heme (26).

A close relative to CYBDOMs is the plant protein AIR12, which is composed of a single DOMON domain, including the conserved Met-His pair that in P. chrysosporium is involved in heme ligation (21, 51). AIR12 is attached to the external leaflet of plant PMs via a GPI anchor and was experimentally demonstrated to bind a single heme b (51). The peculiar electron paramagnetic resonance spectrum of AIR12 is consistent with the Met-His heme-coordination displayed by CDH. AIR12 is the only CYBDOM-like DOMON-domain for which heme binding was experimentally demonstrated. Arabidopsis contains a single AIR12 gene, and AIR12-knock out mutants are resistant to the necrotrophic pathogen Botrytis cinerea (R. Barbaro, A. Costa, F. Sicilia, P. Trost, unpublished). Whether the function of AIR12 depends on the interaction with a CYB561 isoform (reminiscent of CYBDOM proteins) remains to be demonstrated.
Why CYBDOMs?
The architecture of CYBDOM proteins, with a heme b-binding DOMON-domain linked to a five-α-helix CYB561-domain, suggests an ‘electron wire,’ extending well beyond the PM surface. Asking how the function of CYBDOMs may be related to their redox activity and architecture is particularly challenging in plants, where CYBDOMs are often coded by ten genes or more.
P. chrysosporium lines, defective in the DOMON-enzyme CDH, fail to colonize wood. It has been proposed that CDH may participate in lignin degradation (38, 63). In this view, CDH, extending into the cell wall, reduces an Fe3+-chelate (e.g., Fe3+-oxalate) with cellobiose being the reductant. In the presence of hydrogen peroxide (possibly produced by the flavo-domain of CDH itself), the reduced iron could generate hydroxyl radicals through Fenton chemistry, which may attack cell wall polymers, including lignin. CYBDOMs may well be involved in a similar reaction. The CYB561-core could provide cytoplasmic reducing equivalents to the extracellular heme(s), supporting hydroxyl radical-based modifications of cell wall polymers at a safe distance from the PM. As cell wall-loosening agents in plants, hydroxyl radicals are involved in fundamental processes such as for example, seed germination and cell elongation (40). In this context, CYBDOMs may constitute a redox bridge between cytosolic ASC and cell wall sites where hydroxyl radicals may be locally produced by DOMON-catalyzed Fenton chemistry, similar to the DOMON-containing CDH of basidiomycetes.
Footnotes
Acknowledgments
This work was supported by the Flemish Fund for Scientific Research, FWO (grant G.0118.07N to H.A.), by the Hungary-Romania Cross Border Cooperation Program of the EU (HURO/0901/219; to A.B.), and by the Italian Ministery of University and Research (PRIN2009 to PT and RB). The help of Dr. Tibor Pali (Biological Research Center, Szeged, Hungary) with the composition of ![]() , and of the HUGO Gene Nomenclature Committee, are gratefully acknowledged.
, and of the HUGO Gene Nomenclature Committee, are gratefully acknowledged.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.