
Abstract
Introduction
Of particular importance is the redox (or electrophilic) activity of 8-nitroguanosine, which suggested to us that guanine nitration may have a biological function (3, 47). In this context, our discovery of a nitrated cyclic nucleotide, 8-nitroguanosine 3′,5′-cyclic monophosphate (8-nitro-cGMP), led to many findings; for example, 8-nitro-cGMP, in addition to having the potential to activate cGMP-dependent protein kinase (PKG), evidenced the strongest electrophilic property among all nitrated guanine derivatives that we tested (48, 50). 8-Nitro-cGMP thus acts as an electrophilic signaling molecule, so that it can react with particularly reactive sulfhydryl groups of Cys residues to form a protein–(8-S-Cys)-cGMP adduct, and we identified this reaction as a post-translational modification (PTM) of proteins that we called protein S-guanylation (50).
An emerging aspect of redox signaling is mediation of this signaling by electrophilic byproducts of NO and ROS, which include the electrophilic nucleotide as well as various unsaturated fatty acid-derived species generated by means of reactions of ROS, NO, and their products (RNOS) (45, 58). The regulatory mechanisms and metabolism of these electrophilic signaling molecules remain undefined however. Our recent study revealed that the enzymatically generated hydrogen sulfide anion (HS−) regulates the metabolism and signaling actions of endogenous electrophiles (37). In this review article, therefore, we describe electrophilic signaling mediated by 8-nitro-cGMP, with a specific focus on the signaling function via protein S-guanylation and its regulation by hydrogen sulfide (H2S), which we recently discovered.
Formation of 8-Nitro-cGMP
Guanine nitration
NO was first discovered as a signaling molecule that regulates vascular tone and neuronal systems (6, 36). Although its signaling functions are mainly mediated via a cGMP-dependent mechanism, other pathways that are not directly related to cGMP may be responsible for diverse aspects of NO signaling (7, 33, 43, 51). Such a cGMP-independent pathway may be controlled by nitrosylation and nitration of amino acids, proteins, lipids, and nucleotides, with these biological processes being induced by RNOS (which are derived from NO and ROS) such as peroxynitrite (ONOO−) and nitrogen dioxide (NO2) (Fig. 1) (5, 14, 43, 50, 51, 59). For several years, nitration of nucleic acids was thought to be caused by RNOS, as our in vitro study showed (39). Also, we and other groups reported the presence of nitrated guanine derivatives, including 8-nitroguanine and 8-nitroguanosine, and their formation in various cultured cells in vitro and in tissues in vivo from animals with viral pneumonia and humans with lung disease, and even in human urine (3, 39, 49, 56, 62).
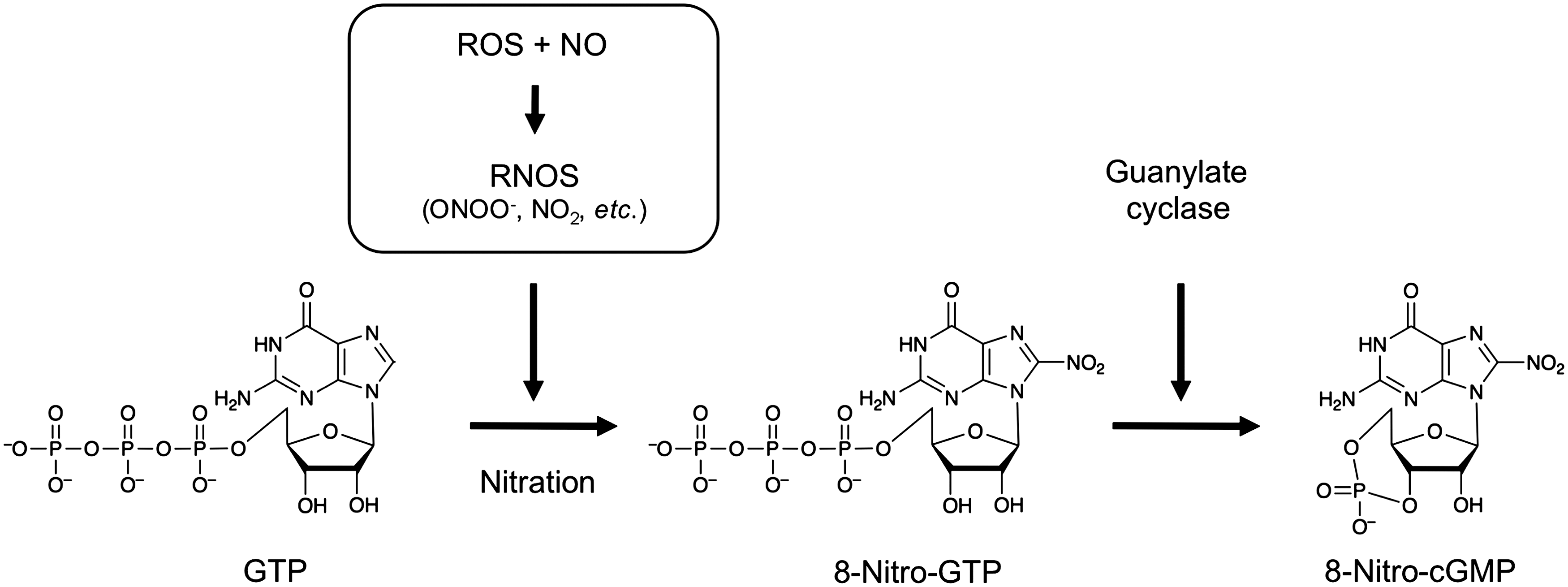
Because of the abundant guanine nucleotides stored in cells, their derivatives would be expected to become good targets of nitration by RNOS. To verify the biological formation of nitroguanine and its derivatives, we initially synthesized various derivatives, including 8-nitroxanthine, 8-nitroguanine, 8-nitroguanosine, and the monophosphate, triphosphate, and cyclic monophosphate of 8-nitroguanosine, as standard authentic compounds to be used for chemical and immunochemical determination of guanine nitration occurring in vivo (50). We then prepared polyclonal and monoclonal antibodies for these nitrated nucleotides. These antibodies were indeed useful not only for immunochemical analyses but also for chemical identification of 8-nitroguanine-related compounds that were to be separated from crude cell and tissue extracts and even from biological samples such as urine. For example, with high-performance liquid chromatography (HPLC) with electrochemical detection (ECD), we elucidated the chemical identity of 8-nitro-cGMP (50). We applied the lysate of RAW 264.7 cells (a murine macrophage cell line), which had been stimulated with lipopolysaccharide (LPS) and cytokines to express iNOS, to an immunoaffinity column containing antinitroguanine antibodies (clones 1G6 and NO2-52) to separate and enrich 8-nitroguanine derivatives, followed by HPLC-ECD analysis. We thereby established that 8-nitro-cGMP was a major nitration product as well as found minor components such as 8-nitroxanthine and 8-nitroguanine. Liquid chromatography–tandem mass spectrometry (LC-MS/MS) confirmed this finding of 8-nitro-cGMP as a major nitration product among various nitrated guanine derivatives, as we previously reported (50).
Mechanism of guanine nucleotide nitration in cells
Another issue that we sought to clarify was how nitration of guanine nucleotides could occur in cells. As described above, to achieve biologically relevant nitration, cells must express both superoxide anion (O2 −) and NO to produce the potent nitrating agent ONOO−, or nitrite plus hydrogen peroxide (H2O2)–heme proteins such as myeloperoxidase to generate another nitrating agent, NO2 (43). Chemical and cellular analyses conducted in our laboratory verified that ONOO− was the most likely molecular species responsible for nitration of guanine nucleotides. In addition to ONOO−, however, nitrite plus H2O2 and heme proteins such as myeloperoxidase may nitrate guanine nucleotides in a particular molecular environment in cells. We used rat C6 glioma cells to study cell formation of 8-nitro-cGMP, because these cells are well known to produce superoxide and NO in response to stimulation with LPS plus proinflammatory cytokines via activation of NADPH oxidase 2 (Nox2) and expression of iNOS (1, 18). In these stimulated cells, mitochondrion-derived superoxide was identified as a direct determinant of 8-nitro-cGMP formation. Immunocyte/histochemical analyses of cultured macrophages and human lung tissue also revealed that the mitochondrion is one of a major organelles where 8-nitro-cGMP is formed (50, 56). These findings were the first demonstration that mitochondrion-derived superoxide with concomitant NO generation plays an important role in biological nitration of guanine nucleotides. Mitochondrion-derived superoxide production was itself regulated by H2O2 generated from Nox2, which suggests a critical link between Nox2-dependent H2O2 production and mitochondrial superoxide production (1). This finding may lend credence to our proposal that 8-nitro-cGMP may serve as a unique second messenger formed downstream of NO and ROS generation.
Additional data unequivocally proved the biological relevance of endogenous 8-nitro-cGMP formation. We performed extensive LC-MS/MS analyses by utilizing the stable isotope dilution technique with C6 cells expressing iNOS, so that we could measure the exact amount of 8-nitro-cGMP produced in cells (18). We analyzed not only 8-nitro-cGMP but also cGMP, because clarification of their distinctive production kinetics (quantity- and time-dependent profiles), and thus differentiation between the two compounds, may lead to better understanding of their physiological and even pathological significance. In fact, we could assess the exact amount of endogenous 8-nitro-cGMP by using our stable isotope dilution technique and LC-MS/MS analysis in a spike-and-recovery study with control nucleotides synthesized with stable 15N isotopes. By means of this technique, we determined that quite high levels (>40 μM) of 8-nitro-cGMP were formed in cells, and that these levels were much higher than cGMP levels formed in the same cells (18). This result indicates that 8-nitro-cGMP is one of the major cyclic nucleotides formed in cells. Also, our interpretation of the 8-nitro-cGMP formation mechanism obtained from our quantitative LC-MS/MS analysis is that 8-nitro-cGMP is not formed by direct nitration of cGMP; rather, 8-nitro-cGMP is eventually generated after nitration of the abundant GTP, to produce 8-nitro-GTP, which then reacts with soluble guanylate cyclase (sGC) to generate 8-nitro-cGMP (Fig. 1) (18). Indeed, studies with in vitro systems revealed that among various guanine nucleotides, including cGMP, GTP was most susceptible to ONOO−-mediated nitration (1). Furthermore, we found that the cells that little express sGC (such as mouse macrophages) produced significant levels of 8-nitro-cGMP, and also found from our preliminary in vitro system that not only sGC but also a particulate form of GC can catalyze the 8-nitro-cGMP formation (18, 23, 50, our unpublished data). These findings suggest that nitration of GTP is a main pathway for 8-nitro-cGMP formation in cells.
Metabolism of 8-Nitro-cGMP
Reaction with glutathione
Oxidative stress reportedly leads to production of various endogenous electrophiles, including 8-nitro-cGMP and nitrofatty acids. Understanding of their biological functions necessitates comparison of their chemical properties such as reactivity with sulfhydryls, reversibility of electrophilic adduction (S-alkylation), compartmentalization, and biological stability. In general, electrophiles react readily with the sulfhydryls of Cys residues of different sulfhydryl-containing peptides and proteins, because they have high reaction rate constants with sulfhydryls. Most highly reactive electrophiles seem to undergo reversible electrophilic adduction almost nonselectively with many sulfhydryls, and thus the stability of such adductions in cells may be greatly affected by glutathione (GSH), which is a major cellular low-molecular-weight sulfhydryl-containing compound (Fig. 2). Because of the redox-recycling potential of GSH that is present in cells at greater than millimolar concentrations, the C–S bonds of the S-alkylation protein adducts will effectively dissociate, and the electrophiles released will again bind covalently with GSH to form low-molecular-weight S-alkylation adducts. GSH is therefore a determinant of the metabolic fate of different electrophiles formed either endogenously or exogenously (Fig. 2).

The best-characterized chemical and pharmacological feature of 8-nitro-cGMP is its relative stability in cells, such that it maintains its signaling functions. In fact, the electrophilicity of 8-nitro-cGMP is much lower than that of other endogenous electrophiles. The second-order rate constant for the reaction of 8-nitro-cGMP with the GSH sulfhydryl is 0.03 M −1 s−1 at pH 7.4 and 37°C (Table 1) (50). 8-Nitro-cGMP is thus believed to be much less reactive compared with certain other electrophiles such as α,β-unsaturated aldehydes, ω-6 and ω-3 unsaturated fatty acids, and nitroalkene fatty acids, which include 4-hydroxy-2-nonenal (HNE), 15-deoxy-Δ12,14-prostaglandin J2, and nitrolinoleic and nitro-oleic acids. The second-order rate constants for the reaction of those electrophiles with GSH are 1.3 M −1 s−1 (HNE), 0.7 M −1 s−1 (15-deoxy-Δ12,14-prostaglandin J2), 355 M −1 s−1 (nitrolinoleic acid), and 183 M −1 s−1 (nitro-oleic acid) at pH 7.4 and 37°C (4, 13, 48). Therefore, these reactive electrophiles can react with GSH sulfhydryls more than 20–10,000 times more rapidly than does 8-nitro-cGMP (48). In other words, 8-nitro-cGMP is stable enough to remain at appreciable levels in cells even with excessive amounts of GSH.
HNE, 4-hydroxy-2-nonenal.
The reaction of some electrophiles with GSH is facilitated by enzymes such as glutathione S-transferase (GST). Reactive electrophiles produced in cells during oxidative stress are detoxified by the actions of GST (21). Although we did investigate whether 8-nitro-cGMP could serve as a substrate for GST, we observed no obvious degradation of 8-nitro-cGMP by the catalytic reaction with GST (unpublished data). Indeed, even with the use of several isoforms of GST, 8-nitro-cGMP was not metabolized by GSH. This notable biochemical characteristic of this compound thus supports its remarkable stability and thus its availability for cellular redox signaling.
The relatively stable nature of the endogenous electrophile 8-nitro-cGMP compared with other endogenously produced electrophiles may validate the major biological relevance of 8-nitro-cGMP formed in cells. As discussed above, in our recent isotope dilution–LC-MS/MS analysis, we carefully analyzed and precisely quantified the 8-nitro-cGMP formed in cells. That a large amount of 8-nitro-cGMP persisted in cells may be consistent with its poor electrophilic nature, because electrophilic compounds are believed to react readily with sulfhydryl compounds such as GSH and be degraded, as just mentioned. During its long reaction with sulfhydryls in vitro, 8-nitro-cGMP indeed slowly undergoes denitration and releases its nitro-moiety, so that it loses its unique electrophilicity. However, because of the reaction rate constants for the sulfhydryl of GSH noted earlier, we now believe that 8-nitro-cGMP, compared with other biological electrophiles, is an inert electrophile. We again found support for this electrophilic property in our recent finding that generation of 8-nitro-cGMP was not greatly affected after elimination of GSH by administering buthionine sulfoximine to NO-treated cells or to LPS plus cytokine-treated cells (18).
Reaction with H2S
Endogenous formation of H2S has clearly been observed in recent years in mammalian cells and tissues (26, 32). The chemical nature and physiological functions of H2S are mostly undefined, however. Although H2S is proposed to have anti-inflammatory and antioxidant effects, in view of the modest rate constants for the reaction of H2S with ROS and RNOS such as H2O2 and ONOO− (9), H2S is not expected to directly scavenge ROS and RNOS unless they are strong electrophiles.
Because GSH does not seem to be a major determinant of 8-nitro-cGMP metabolism, as mentioned above, we investigated, by utilizing RNAi screening with a focus on Cys metabolism and its redox-related metabolic pathways, other as-yet unidentified factors that may contribute to the regulation and metabolic pathways of 8-nitro-cGMP signaling (37). Our screening revealed a significant impact of endogenous H2S generation on 8-nitro-cGMP metabolism. Two key enzymes of H2S biosynthesis, cystathionine β-synthase (CBS) and cystathionine γ-lyase, appeared to mediate the metabolism of cellular 8-nitro-cGMP in various cultured mammalian cells. This metabolism occurred concomitantly with the release of nitrite, which suggests a mechanism that depends on the nucleophilic qualities of H2S (pK a 6.7), especially the anionic HS− that predominates in neutral biological solutions (Fig. 3). To clarify how H2S could undergo an addition reaction (i.e., electrophile sulfhydration), we used HPLC-MS/MS to analyze the reaction of 8-nitro-cGMP with HS− in a cell-free system. The electrophilic nitro-moiety underwent nucleophilic substitution, the yield being the novel product 8-SH-cGMP (Fig. 3).

Such electrophile sulfhydration by HS− occurs with a broad array of biological electrophiles, including 15-deoxy-Δ12,14-prostaglandin J2, HNE, acrolein, and fatty acid nitroalkene derivatives. The metabolic fate of these sulfhydrated derivatives allows the sorting of the electrophiles into three groups (Fig. 4). In the first group (including 8-nitro-cGMP and 15-deoxy-Δ12,14-prostaglandin J2), the SH derivative is so stable that no additional reaction occurs, except for oxidative degradation of SH by ROS and other reactive species. The second group (including nitro-oleic acid, 1,2-naphthoquinone, 1,4-naphthoquinone, tert-butylbenzoquinone, N-ethylmaleimide, diethylmaleate, and monobromobimane) involves a relatively stable bis-product formation. The third group may include other highly reactive electrophiles, such as HNE and acrolein, for which additional metabolism and secondary chemical reactions follow SH addition. 8-SH-cGMP seems to be an extremely stable end product among sulfhydrated derivatives of the above-mentioned electrophiles.

Endogenous HS−-dependent sulfhydration occurs in various cultured cells, as evidenced by 8-SH-cGMP formation after addition of exogenous 8-nitro-cGMP (37). Knockdown of CBS markedly inhibited formation of 8-SH-cGMP in A549 cells, which indicated that CBS was a major source of endogenous H2S/HS−. 8-SH-cGMP contains sulfhydryls and is therefore presumably oxygen labile or ROS susceptible, so it may be oxidized further (Fig. 4). This idea is supported by the fact that 8-SH-cGMP treated with H2O2 or ONOO− decomposed readily, at least in vitro in a cell-free reaction mixture. Unexpectedly, however, the sole product observed in this reaction was cGMP, rather than oxidized derivatives of 8-SH-cGMP such as 8-sulfenyl-, 8-sulfinyl-, and 8-sulfonate-cGMP, or 8-OH-cGMP. This result provides the first demonstration of oxidant-induced desulfhydration of SH-containing compounds. All these findings may thus suggest that H2S produced in cells is a critical factor in 8-nitro-cGMP metabolism. It may be possible that, however, other sulfhydryl-related antioxidant systems such as thioredoxin and peroxiredoxin could mediate the metabolism of 8-nitro-cGMP, which may require further investigation.
Phosphodiesterase resistance
Another biochemical characteristic of 8-nitro-cGMP that may determine its pharmacological activities is its potent molecular interaction with cyclic nucleotide (cGMP) receptor proteins such as PKG and other cGMP-binding proteins. The function of S-guanylated proteins may therefore be affected by the structural features of the parental cGMP. Also, of great importance is that all these C8-position-modified cGMP derivatives, including 8-nitro-cGMP, 8-SH-cGMP, 8-Cys-cGMP, and other peptide/protein Cys-cGMP (S-guanylated) adducts, acquire resistance to phosphodiesterases (PDEs), which may sustain the strong, steady stimulation conferred during signal transduction by PKG, cGMP-gated ion channels, and even signaling via protein S-guanylation, discussed below (50). With regard to this PDE-dependent metabolism, especially in terms of reversible metabolism of 8-nitro-cGMP, it is quite conceivable that after 8-nitro-cGMP is sulfhydrated and converted to 8-SH-cGMP and then to cGMP via ROS- or RNOS-mediated oxidative desulfhydration, cGMP is readily metabolized and decomposed to GMP by the many PDEs present in cells and tissues. 8-Nitro-cGMP, being affected by H2S, may serve as an effective electrophilic regulator of intracellular signaling by virtue of its unique, elegant chemistry involving electrophilic reactions with protein sulfhydryls and resistance to metabolic degradation by PDEs, which are highly expressed by many types of cells and tissues.
Signal Transduction via Protein S-Guanylation
Moderate electrophilicity of 8-nitro-cGMP and selective cell signaling
That 8-nitro-cGMP as an electrophile has unique chemical reactivity with protein Cys residues is now clear. As one example of such reactivity, 8-nitro-cGMP undergoes nucleophilic substitution with a protein sulfhydryl, releases the nitro group, and forms a protein Cys-cGMP adduct in a process called protein S-guanylation, which is also said to be a protein PTM (Fig. 2). Indeed, endogenous protein S-guanylation occurring in cells is currently a major focus of our studies, because its identification may stimulate new oxidative stress and redox signaling research.
During physiological signal transduction via protein S-guanylation, the relatively low electrophilicity or redox reactivity of 8-nitro-cGMP may be a critical factor that can determine its specificity in protein S-guanylation (Fig. 5). For electrophiles that are highly reactive, nucleophilic amino acids other than Cys, especially histidine and lysine, also become targets in electrophilic reactions, that is, Michael additions. Because these electrophiles have a very high electron-withdrawing potential, they undergo unstable, reversible S-alkylation, with the alkyl group at some point being transferred to other sulfhydryls of Cys of different proteins, in a process called transalkylation. This finding indicates that many endogenous electrophiles, but not 8-nitro-cGMP, may not necessarily generate site-specific PTMs, which can transduce signaling (Figs. 2 and 5). Such electrophiles may require specific reaction conditions or compartmentalization, so that they can act as signaling molecules and have significant effects on sulfhydryls of acceptor proteins. For example, particular unique structural characteristics, yet unidentified, may be requirements for much of the stable covalent binding of electrophilic protein S-alkylation occurring near PTM sites.
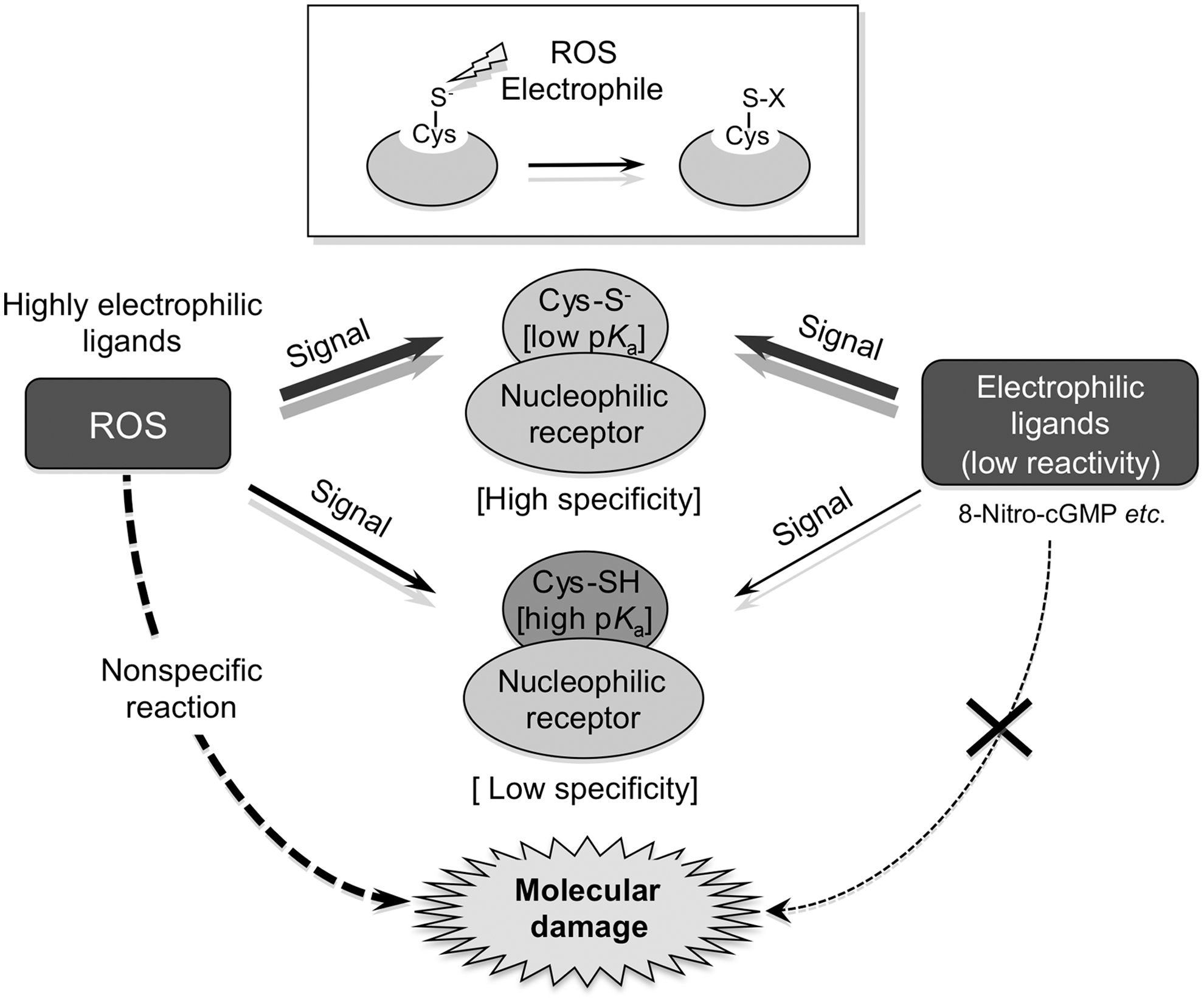
The major distinction between S-guanylation and other S-alkylation reactions is that S-guanylation appears to be a quite stable, irreversible sulfhydryl modification, because the nucleophilic nitro-moiety of its purine structure is lost during formation of adducts with protein Cys residues. Another important characteristic of 8-nitro-cGMP is that because of its inert chemical reactivity as an electrophile compared with other electrophiles, S-guanylation occurs almost solely with sulfhydryls having high nucleophilicity, as determined by the pK a of the Cys-moiety sulfhydryls (Fig. 5). This evidence is strongly supported by the finding that a particular Keap1 Cys residue is highly susceptible to a specific 8-nitro-cGMP-induced S-guanylation site, as described below (2, 18, 46, 48, 50, 63). Also, our preliminary studies suggested that S-guanylated proteins may be readily degraded via autophagy, so that this irreversible PTM can be compensated by a rapid protein turnover (54; our unpublished observation).
Keap1 S-Guanylation and a Cytoprotective Adaptive Response
One of the most important target proteins for S-guanylation is Keap1, which is well known as a potent redox-sensing protein. Keap1 is a negative regulator of nuclear factor erythroid 2-related factor 2 (Nrf2), a transcription factor that regulates phase II detoxifying and antioxidant enzymes for electrophiles and ROS (11, 35). Binding of Keap1 to Nrf2 maintains the cytosolic localization of Nrf2 and mediates rapid degradation of Nrf2 by proteasomes. Because Keap1 has highly reactive Cys residues, chemical modification of the sulfhydryl group of Cys residues by electrophiles and ROS has been proposed to trigger dissociation of Nrf2, which would lead to its stabilization and nuclear translocation. Activated Nrf2 would then bind to the antioxidant-responsive element to induce expression of various cytoprotective enzymes, which would contribute to the adaptive response to oxidative stress (10, 24).
In our studies, we identified Keap1 as a target protein for S-guanylation in cells and thereby provided the first proof of protein S-guanylation in cells (50). Considerable evidence from our recent work shows that Keap1 S-guanylation and subsequent Nrf2 activation were mediated by 8-nitro-cGMP formed via NO derived from NO donors or generated by iNOS (2, 18, 46, 48, 50, 63). For example, in C6 cells treated with LPS plus cytokines, we discovered evidence of Keap1 S-guanylation, by means of Western blotting using anti-S-guanylation antibody after Keap1 isolation via immunoprecipitation, along with 8-nitro-cGMP formation in cells, as described above (18). We observed similar Keap1 S-guanylation in macrophages after Salmonella infection, which depended on iNOS-derived NO production and subsequent formation of 8-nitro-cGMP (63). General proteomic profiling of S-guanylated proteins, which we recently clarified, revealed that Keap1 is one of the major target proteins that is S-guanylated after NO exposure and that its S-guanylated structure derives solely from RS-cGMP adducts (18). We did not observe other chemical modifications of Cys residues, such as sulfhydryl oxidation and S-nitrosylation, which are often caused by ROS and NO (8, 25, 43) in Keap1 in cells treated with an NO donor (18). Therefore, that Keap1 may play a predominant role in the NO-signaling pathway that depends on 8-nitro-cGMP produced in cells is highly plausible.
An intriguing result of proteomic analysis for endogenously modified Keap1 with matrix-assisted laser desorption/ionization time-of-flight-MS/MS was that the Cys434 residue of Keap1 was S-guanylated by 8-nitro-cGMP in a site-specific manner (18). In recombinant Keap1 reacted with 8-nitro-cGMP in an in vitro cell-free system, however, 18 of 25 total Cys residues were S-guanylated, and Cys434 S-guanylation was not dominant. Therefore, in cells, the Cys434 of Keap1 may have structural or chemical properties that are related to its specific S-guanylation and that are distinct from those of other Cys residues of Keap1. A number of studies have investigated Keap1–electrophile adducts generated in vitro in the presence of high concentrations of electrophilic reagents (8, 10, 28, 29, 31, 38). In addition, Yamamoto's group reportedly showed the in vivo importance of Cys151, Cys273, and Cys288 in Keap1 functions by means of a transgenic complementation rescue experiment in mice (61). Although the in vivo significance of Cys434 modification remains to be clarified, one study found that Cys434 in Keap1 is one of the Cys residues that are most sensitive to S-glutathionylation, which causes marked structural changes in the Nrf2-binding surface of the Keap1 molecule (22). X-ray crystallographic analysis data showed that Cys434 lies close to the Nrf2-binding region of the DC domain of Keap1, which may have an impact on Keap1–Nrf2 binding, so S-guanylation of Cys434 may facilitate dissociation of Nrf2 from Keap1. The specific S-guanylation of Cys434 by 8-nitro-cGMP may thus have a critical role in molecular sensing performed by Keap1 for RNOS and/or ROS (18).
That 8-nitro-cGMP was in fact involved in the cytoprotective response through S-guanylation of Keap1 was verified by the finding that treatment of C6 cells with 8-nitro-cGMP reduced cell death induced by oxidative stress related to H2O2 exposure (18). Our study also showed that 8-nitro-cGMP increased Nrf2 nuclear accumulation and heme oxygenase-1 (HO-1) expression in C6 cells (18). NO-induced expression of HO-1 reportedly mediated cell survival in solid tumor models and during bacterial infection (12, 40, 55, 63). All these data therefore indicate that the cytoprotection against oxidative stress that 8-nitro-cGMP confers is associated, at least in part, with increased HO-1 expression. Therefore, the idea that 8-nitro-cGMP is a major component of the NO- and ROS-signaling pathway that confers cytoprotection from or adaptive responses to ROS and oxidative stress is highly conceivable.
H-Ras S-Guanylation and Cellular Senescence
The small GTP-binding protein Ras is a multifunctional signaling protein that regulates various cellular process such as growth, proliferation, and differentiation (20). For many years, ROS have been implicated in the regulation of Ras signaling. For example, oxidative inflammatory reactions involving NO-derived species induce an H-Ras oncogenic cellular response (30, 41), which in turn can activate p53-dependent cellular senescence (52, 60). However, the precise molecular mechanism of the effect of RNOS and ROS on Ras signaling is not yet fully elucidated.
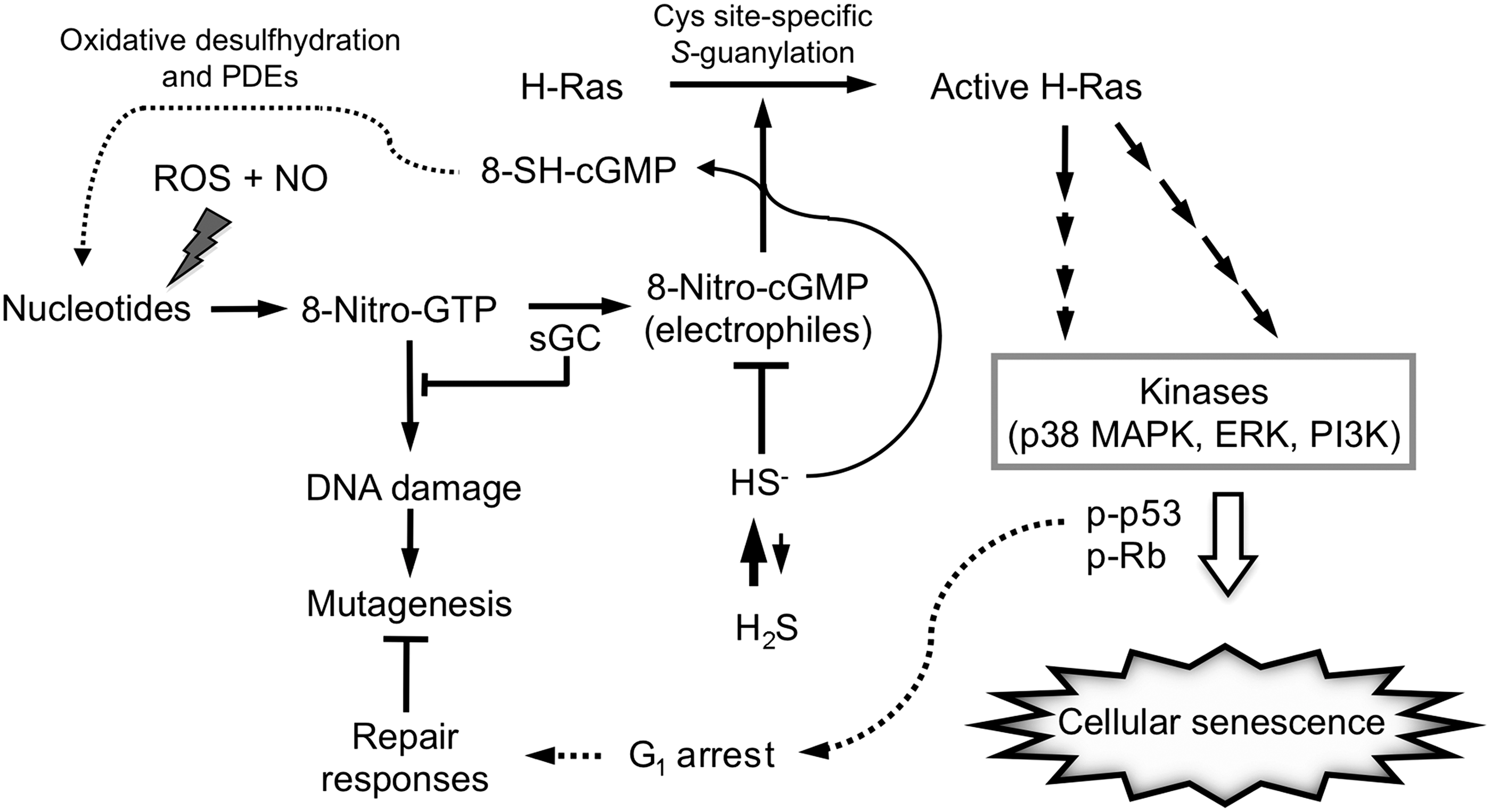
Our recent study clarified that S-guanylation of H-Ras induced by 8-nitro-cGMP activated H-Ras and was involved in cellular senescence (Fig. 6) (37). Exogenous 8-nitro-cGMP, but not NO or cGMP, produced growth arrest and significantly increased the senescence of rat cardiac fibroblasts and myocytes in culture. iNOS-inducing stimuli (LPS and ATP) induced cellular senescence, with marked 8-nitro-cGMP formation and concomitant S-guanylation of H-Ras, in cardiac cells. Because H2S is an important regulator of 8-nitro-cGMP metabolism, as described above, the effect of H2S treatment was investigated in terms of H-Ras activation induced by 8-nitro-cGMP. H2S treatment of cardiac cells markedly reduced 8-nitro-cGMP formation and S-guanylation of H-Ras induced by the stimuli, which resulted in suppressed cellular senescence. Notably, knockdown of CBS in various cultured cells induced both S-guanylation and activation of H-Ras, and H2S treatment suppressed these changes. These data thus suggest that H-Ras activation via S-guanylation by 8-nitro-cGMP is critically involved in cellular senescence, and that endogenously produced HS− is a potent regulator of this signaling process.

Site-specific H-Ras S-guanylation and its inhibition by H2S were verified by Western blotting and proteomic analysis of recombinant H-Ras protein and its various Cys mutants treated with 8-nitro-cGMP. Moreover, LC-MS/MS analysis revealed that only Cys184 of the H-Ras protein was S-guanylated. H-Ras Cys184 was thus identified as a nucleophilic sensor that was highly susceptible to 8-nitro-cGMP-induced protein S-guanylation. Further extensive analysis using Cys-mutant H-Ras and cultured cells revealed the impact of site-specific H-Ras Cys184 S-guanylation in signaling, leading to cellular senescence. This result indicates that Cys184 of H-Ras is functionally critical for sensing electrophilic ligands such as 8-nitro-cGMP (Fig. 6).
With regard to the H-Ras protein structure, Cys184 of H-Ras is one of two palmitoylation sites located at the C-terminal domain. Cys184 is not essential for the targeting of H-Ras to the plasma membrane, but it is necessary for the control of GTP-regulated lateral segmentation of H-Ras between lipid rafts and nonrafts, which is required for efficient Raf activation (42, 44). Also, inhibiting Cys184 palmitoylation efficiently delivers H-Ras to the plasma membrane with little Golgi pooling (44), which suggests that S-guanylation of H-Ras at Cys184 promotes localization of H-Ras at the plasma membrane and association with Raf by inducing dissociation of H-Ras from lipid rafts.
Indeed, 8-nitro-cGMP treatment caused sustained activation of downstream signaling cascades, such as extracellular signal-regulated protein kinase (ERK), p38 mitogen-activated protein kinase (p38 MAPK), p53, and Rb. Inhibition of ERK, p38 MAPK, and phosphatidylinositol-3-kinase (PI3K) suppressed 8-nitro-cGMP-induced senescence in cardiac cells. We therefore suggest that the Cys184 of H-Ras is a functionally critical sensor for endogenous electrophiles such as 8-nitro-cGMP in their H-Ras-mediated downstream signal transduction (Fig. 6). The Cys184 of H-Ras may be chemically modified not only by 8-nitro-cGMP but also by HNE and 15-deoxy-Δ12,14-prostaglandin J2 and causes H-Ras activation. However, downstream signals of H-Ras, such as ERK and p38 MAP kinase, are only activated by 8-nitro-cGMP, but not necessarily activated by other reactive electrophiles such as HNE and 15-deoxy-Δ12,14-prostaglandin J2 (our unpublished data), probably because these reactive electrophiles may nonspecifically affect signaling molecules other than H-Ras and may perturb specific signaling pathways as was observed with 8-nitro-cGMP. In view of the much higher electrophilicity of HNE and 15-deoxy-Δ12,14-prostaglandin J2 compared with that of 8-nitro-cGMP, these findings strongly suggest that 8-nitro-cGMP acts as a specific physiological ligand for H-Ras activation.
The physiological relevance of this signaling mediated by H-Ras S-guanylation and its regulation by H2S was substantiated by data obtained with the mouse model of MI-induced chronic heart failure. In vivo H2S treatment greatly inhibited 8-nitro-cGMP formation and markedly reduced H-Ras activation and H-Ras S-guanylation in hypertrophic lesions of the post-MI heart. Phosphorylation of p38 MAPK, ERK, p53, and Rb proteins increased in post-MI hearts, with H2S treatment suppressing their activation as well as cardiac dysfunction (37).
Biological Relevance of 8-Nitro-cGMP Formation
One of the most important physiologically relevant functions of 8-nitro-cGMP formation is potential cellular protection against ROS/RNOS-induced genotoxicity or nucleotide damage occurring in cells: 8-nitro-cGMP serves as an endogenous ligand for H-Ras activation, which leads to its downstream signaling pathways for p53-mediated growth arrest (Fig. 7). The G1 cell cycle arrest brought about by a p53-dependent signaling mechanism is known to protect oxidatively injured cells from genetic mutation and genotoxicity caused by oxidative stress, provided that many DNA repair systems are stimulated to repair damaged cells and tissues. Abundant GTP stored in the cellular nucleotide pool, as noted above, could be nitrated, and thereby functions as a sensor for nucleotide modification caused by RNOS (1, 18). 8-Nitro-GTP thus generated may serve as a substrate for formation of 8-nitro-cGMP, which is believed to be a crucial step in the response of cells to oxidative stress. Indeed, we demonstrated earlier that 8-nitro-cGMP forms readily in cells via the sGC-catalyzed cyclization of 8-nitro-GTP, which is eventually generated through RNOS-dependent GTP nitration (18). Also, we and other groups showed that 8-nitro-GTP becomes an endogenous mutagen when incorporated into DNA and RNA (27, 62). If the phosphate of GTP undergoes cyclization by GC to become a cyclic phosphate, the resultant cyclic nucleotide cannot be used for DNA or RNA synthesis. Therefore, sGC not only contributes to electrophilic 8-nitro-cGMP signaling but also is likely to eliminate 8-nitro-GTP to prevent induced mutagenesis. In this context of nucleotide base modifications, we propose a unique function of HS−. Because 8-nitro-GTP is much less electrophilic than 8-nitro-cGMP, HS− reacts very poorly with 8-nitro-GTP, and selective sulfhydration of 8-nitro-cGMP, among different 8-nitrated guanine nucleotides, can be expected. It is therefore logical to suggest that H2S functions as a kind of backup repair system, as it modulates H-Ras activation leading to p53 responses and more critically generates the natural DNA substrate GTP from GMP, after 8-SH-cGMP is oxidatively desulfhydrated (to form cGMP) and subsequently undergoes decyclization to GMP by PDEs (Fig. 7).
Conclusion
The seminal studies that we and others have published in recent years have now verified the NO-dependent formation of 8-nitro-cGMP and the simultaneous S-guanylation of sensor proteins, such as Keap1 and H-Ras, induced by 8-nitro-cGMP in animal cardiac tissue and cells in culture. S-Guanylation of Keap1 leads to Nrf2 activation and cytoprotective gene expression, which seems to be involved in the general adaptive response to oxidative stress. H-Ras S-guanylation by 8-nitro-cGMP contributes to cellular senescence and, under some circumstances, functions as a DNA repair system. These findings therefore suggest that protein S-guanylation induced by 8-nitro-cGMP is a unique redox-signaling pathway for modulating various cellular functions mediated by NO and ROS. Another striking finding is that endogenous HS− can regulate 8-nitro-cGMP-signaling pathways in a critical way in cells and whole organisms. We therefore suggest that all these data related to protein S-guanylation induced by 8-nitro-cGMP and its regulation by endogenous HS− warrant additional extensive investigations that may reveal new aspects of redox signaling, NO- and ROS-related physiology and pathology, and pharmaceutical chemistry, with greater understanding of many oxidative stress-related diseases and development of new therapeutics.
Footnotes
Acknowledgments
We thank J.B. Gandy for her editing of the manuscript. We gratefully thank M. Nishida, T. Sawa, H. Ihara, H. Motohashi, M. Yamamoto, M. Suematsu, A. van der Vliet, B.A. Freeman, K. Uchida, and Y. Kumagai. This work was supported in part by grant-in-aid for scientific research and grant-in-aid for scientific research on Innovative Areas (Research in a Proposed Area) from the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan.