
Abstract
Reactive Oxygen Species in Plant–Microbe Interactions
Although ROS are central to preventing parasitism of plants, they are also core to the ability of microbes to successfully invade and colonize plant hosts. Plant–microbe symbiotic interactions, such as plant–mycorrhizal interactions and nitrogen-fixing bacterial symbioses, are dependent on ROS. Successful establishment of a symbiotic association is characterized by an initial oxidative burst response, induced by a chemical cross-talk and leading to ROS accumulation in both the invading microbe and its host (104). Fungal pathogens utilize NOXs and the oxidative burst to initiate developmental stages that are essential for plant infection. For example, mutants for NOX in the rice pathogen Magnaporthe oryzae do not accumulate ROS in appressoria (the structure that attaches to the leaf surface and coordinates penetration) and are unable to penetrate host tissue (52). NOX mutants of other fungi, including the ergot pathogen Claviceps purpurea, have been found to be defective in their ability to colonize host tissue even if they successfully penetrate the host (59, 119). In one of the most ingenious examples of fungal adaptability, it is now known that some necrotrophic fungi (fungi that kill their host cells before colonization) have evolved to cause host cell death, not by production of toxins or enzymes, but by inducing the plant's own programmed cell death responses against itself (65, 82, 89, 91).
This review focuses on another mechanism by which pathogens of plants evolved to utilize ROS in order to successfully parasitize plant hosts: the production of photoactivated perylenequinone toxins that utilize light to produce ROS which damage the host cells, allowing for successful pathogenesis. Plants require light for survival, and these pathogens have exploited this requirement to utilize photosensitization as a tool in host colonization. Furthermore, these toxins primarily produce singlet oxygen (1O2), a highly reactive, non-radical form of ROS that is not a common component of cellular reactions and to which cells have few resistance mechanisms. This review will address the production, biosynthesis, and mode of action of these toxins and their role in disease development. Recent research on cellular resistance mechanisms against perylenequinone toxins and 1O2 will be discussed.
Perylenequinones
Perylenequinones (Fig. 1) are a group of ROS-generating photosensitizers that include well-known compounds which are being exploited for medical and agricultural uses as well as having important functions in nature. For example, the perylenequinone Calphostin C is a potent inhibitor of protein kinase C (130) and has also been investigated for its use in cancer therapy (9, 50, 108). Hypocrellins and their derivatives have been used in China for medicinal purposes and have been extensively studied for their use in photodynamic tumor therapy as well as for their antiviral properties (3, 46, 72, 90). The related polycyclic quinone hypericin is a component of the medicinal herb St. John's Wort (Hypericum perforatum) that is used as a herbal remedy for depression (103). Although hypericin has deficiencies as an anticancer agent, derivatives of hypericin are being synthesized and also tested in photodynamic tumor therapy (143). In agricultural applications, the effectiveness of the fungal biocontrol agent Stagonospora convolvuli against two species of bindweed weeds was correlated with their production of the perylenequinone elsinochrome A (1). Perylenequinones are also used by protozoa, where they function as photoreceptors involved in motility and light-avoidance responses (60, 124, 132).

Despite some widespread distribution in occurrence in nature, the majority of these compounds characterized so far are produced by fungi that are associated with plants, in either parasitic or symbiotic relationships (Table 1). All of the perylenequinone producers are in the phylum Ascomycota, and most cluster within one class, the Dothidiomycetes; thus, production appears to be primarily restricted to a narrow taxonomic group. One exception is the observation that some lichens produce perylenequinone compounds (95); these fungi form symbiotic relationships with algae and are classified in other classes within the Ascomycota. Within a genus or species that produces these compounds, production varies both between species and between strains of a single species (8, 64).
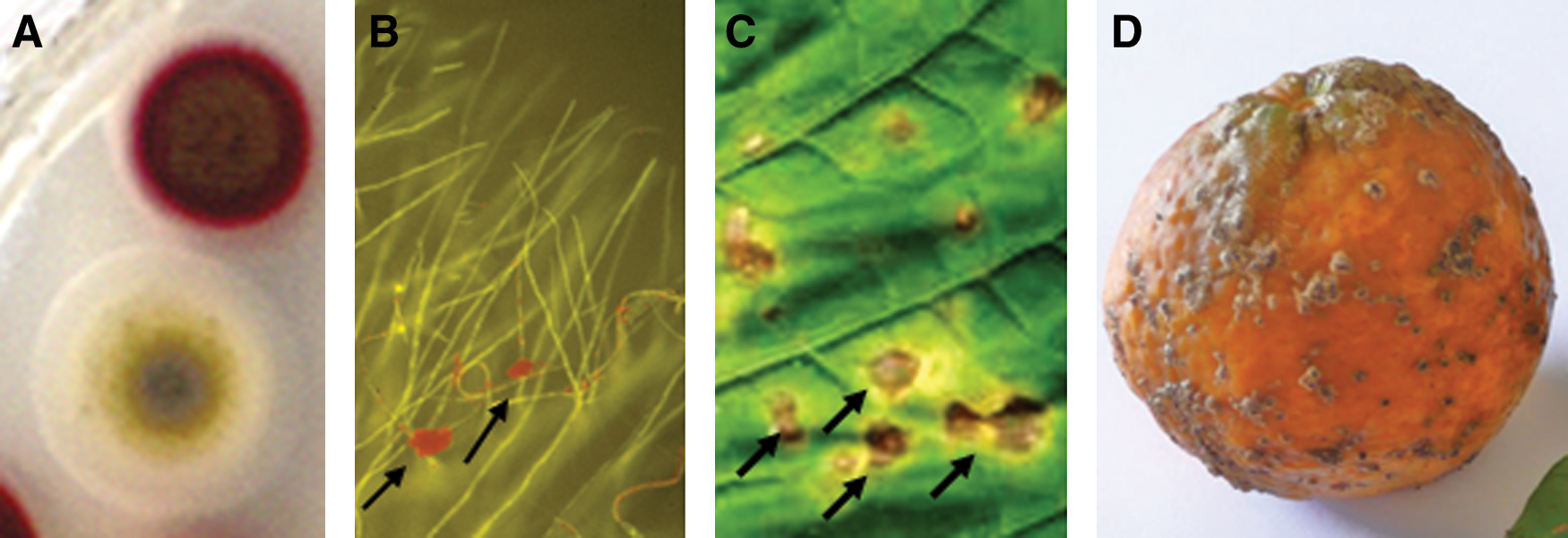
Most of the well-characterized fungal perylenequinones share a common 4,9-dihydroxy-3,10-perylenequinone core and differ in the composition of their side chains (Fig. 1). Their chemical properties were reviewed by Weiss et al. (145). These compounds are bright red pigments that give off a red fluorescence and turn green when dissolved in alkalines (Fig. 2). The compounds vary in their solubility in organic solvents. When reduced, they become yellow–green in color and give off an intense green fluorescence (84, 145). Compounds produced by Alternaria and Stemphylium species, such as altertoxins, alteichin, and stemphyltoxins (Table 1), are partially reduced forms of the other perylenequinones, and are yellow–orange in color (145). Although somewhat structurally different, these compounds have been shown to have similar photoactive, ROS-generating activity as the other perylenequinones (see section below) as well as similar toxicity to plant and animal cells, mutagenic activity, and activity in assays for tumor-suppressing activity (68, 109, 113, 126, 145).
Perylenequinones as ROS-Generating Photosensitizers
Perylenequinones are photosensitizers, a term historically referring to colored compounds that “sensitize” cells to visible wavelengths of light. The term was coined in 1900 when it was observed that common laboratory dyes, such as acridines and xanthenes, killed paramecia that are exposed to visible wavelengths of light (112). Photosensitizers are grouped together based on their mode of action, and compounds of diverse structure have photosensitizing activity. Commonly used dyes such as acridine orange, methylene blue, and rose bengal are photosensitizers. Many natural products found in plants, animals, protozoans, and microorganisms are photosensitizers. These include chlorophyll, porphyrins, and flavins such as riboflavin, coumarins, and polyacetylenes (125).
Photosensitizers are toxic to cells, because they generate ROS (Fig. 3). In the presence of light, photosensitizers are converted to an electronically active triplet state. Triplet-state photosensitizers may react directly with biomolecules (61, 125), but they more commonly react with oxygen through one of two reactions. In the presence of reducing substrates, triplet photosensitizers react by electron transfer reactions. These so-called “type I” reactions result in the production of free radical forms of ROS, such as superoxide (O2 •−), H2O2, or OH•. Photosensitizers may also react directly with oxygen through an energy transfer reaction to form the non-radical, but highly reactive 1O2. This latter reaction is known as a “type II” reaction (125). The fungal perylenequinones are very potent 1O2 producers, with high quantum yields (1O2 yield per quantum of light absorbed). 1O2 quantum yields of cercosporin, elsinochrome A, and hypocrellins A and B have been measured at 0.81–0.97, 0.98, 0.82, and 0.76, respectively (41, 48, 49, 160). A study of eight perylenequinones (cercosporin, hypocrellin A and B, elsinochrome A and D, calphostin C, hypomycin B, erythroaphin, and hypericin) documented that all of them react by both type I and type II reactions and generate 1O2 and O2 •−, which is consistent with previous work documenting the importance of the 4,9-dihydroxy-3,10-perylenequinone core for activity (66).
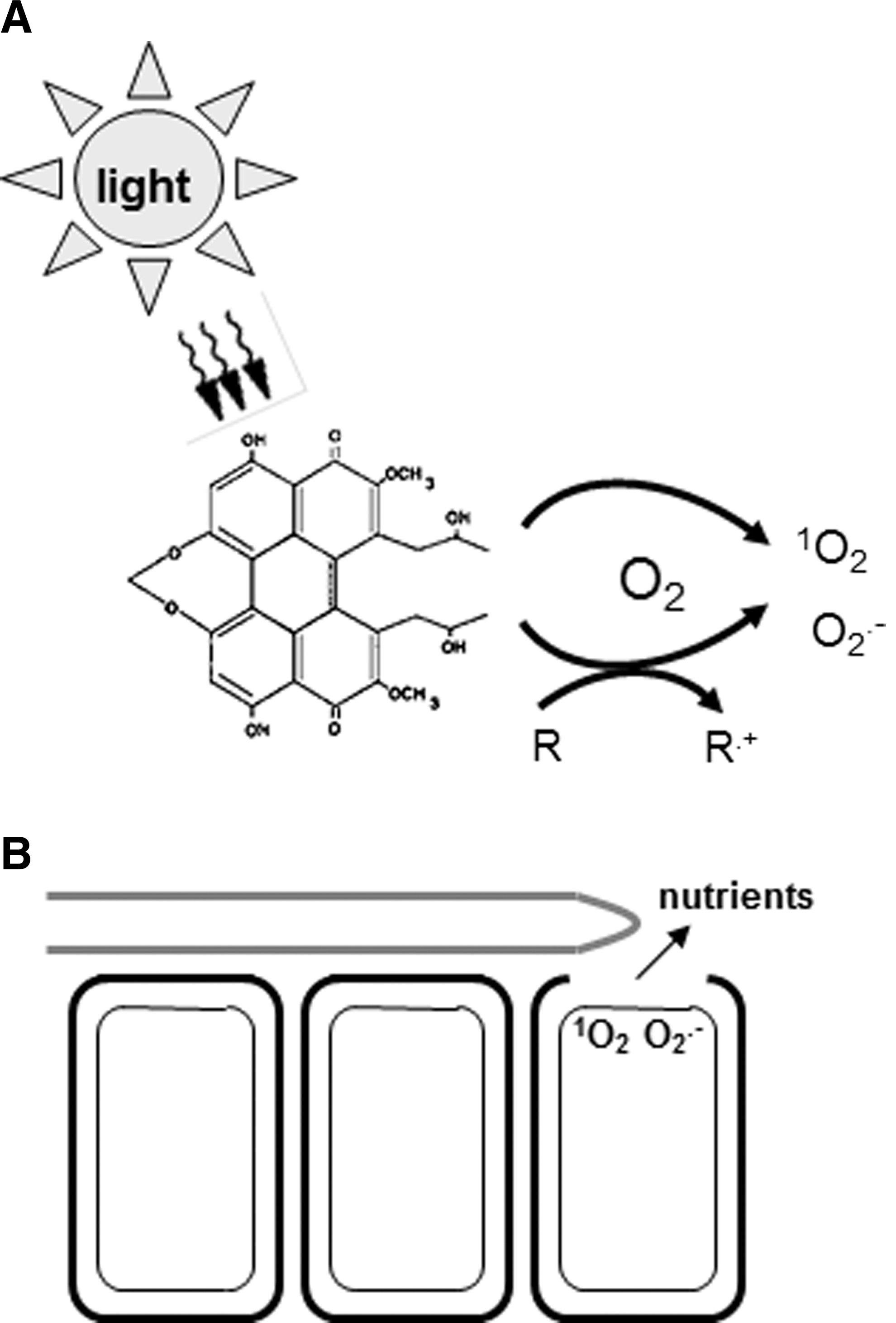
Photosensitizers cause damage to diverse macromolecules and cellular components (62). Lipids, proteins, and nucleic acids are common targets of photosensitizer activity. 1O2 commonly reacts with macromolecules via addition to unsaturated double bonds; thus, unsaturated lipids are highly susceptible. In proteins, aromatic amino acids or those that contain sulfur are particularly susceptible, and nucleic acid damage occurs via a reaction with guanine residues. 1O2 has a short life span [measured at ∼3 μs in a rat neuron model (122)], and does not diffuse long distances in cells. Thus, photosensitizer damage is often determined by the cellular compartment where the photosensitizer localizes (75, 100). For example, membrane-localized photosensitizers commonly result in lipid peroxidation, and nuclear-localized photosensitizers are mutagenic. Localization of the photosensitizer can impact the efficiency of cellular damage, as hydrophobic photosensitizers have been shown to cause more damage than hydrophilic ones that have similar 1O2 quantum yields in vitro (100). Efficiency as well as mode of toxicity (type I vs. type II) is also affected by binding of the photosensitizer to the substrate (74).
Studies on the cellular localization of perylenequinone photosensitizers document that they are lipid soluble, penetrate into cells, and localize in multiple cellular compartments, including cellular membranes and in lysosomes, endoplasmic reticulum, and golgi, and are excluded from nuclei (35, 47, 96). Interest in use of perylenequinones for medical purposes has led to studies of structural features that affect toxicity. Although the 4,9-dihydroxy-3,10-perylenequinone core is central to ROS generation and yields, a study of nine structurally similar perylenequinones showed a large range in activity against membrane-enveloped viruses that did not correlate with 1O2 yields, indicating effects of side groups on activity (72). Studies of side-chain derivatives of hypocrellin B and elsinochrome A, done to increase solubility for clinical applications, showed lower yields of 1O2 and O2 •− as well as decreased toxicity to cancer cells (150, 160), demonstrating that the side chains also affect ROS production.
Perylenequinones in Plant Disease: Cercosporin and the Elsinochromes
Photoactivated perylenequinones are overwhelmingly made by fungi that parasitize plants, suggesting that these compounds play an important role in the evolution and establishment of plant parasitism. Several genera of plant pathogenic fungi produce perylenequinones, but most studies have been done with two, Cercospora species and Elsinoë species. The genus Cercospora includes more than 600 named species of asexual fungi that cause economically important diseases on a wide range of host plants, including important crops such as corn, sugar beet, coffee, and soybean as well as vegetable crops, ornamental plants, native species, and even succulents such as the tequila Agave. These pathogens infect plants through the production of conidia (asexual spores) that germinate on the host leaf surface, penetrate through the stomata, and colonize the intercellular spaces between the cells within the leaf. Cercospora infection results in necrotic lesions (Fig. 2) that often expand to blight the leaf tissue, leading to loss of photosynthetic capacity and severe reductions in yield. Elsinoë species are best known as pathogens of citrus; they not only cause a disease called citrus scab on diverse citrus genotypes, but can also infect other crops such as mango, strawberry, grape, pecan, and cowpea. Unlike Cercospora species, Elsinoë species have a sexual stage, although infection is also via asexual conidia. Elsinoë species cause lesions on leaves, stems, and fruit. Leaf infections can lead to yield losses through stunting of leaves or defoliation. Lesions have a scab-like appearance, and most commonly cause losses by scarring of the fruit, rendering them unsuitable for sale (Fig. 2).
Cercospora species produce cercosporin (Fig. 1), which was first isolated in the 1950s from the soybean pathogen C. kikuchii (83). Since that time, it has been isolated from a large number of Cercospora species as well as from lesions on infected plants [for review see ref. (33)]. Production varies within the genus, with some species, such as C. arachidicola, C. zeina, and C. sojina, not known to synthesize cercosporin (58, 64). Analysis is complicated, however, by the fact that not all isolates which synthesize cercosporin in plants produce it in vitro. Although previously thought to be limited to the genus Cercospora, cercosporin has recently been isolated from an endophytic fungus classified within the closely related sexual genus Mycosphaerella (102). Cercosporin's structure and stereochemistry were determined in the early 1970s (92, 151).
Evidence for cercosporin as a photosensitizer came from diverse studies, including demonstration of the requirement for light and oxygen in toxicity to mice and bacteria (152), a matching of the cercosporin absorption spectrum with the action spectrum for killing of plant cells (31), and protection against toxicity by antioxidants (94). Cercosporin has been shown to react by both type I and type II mechanisms both in vitro and in plants. Production of 1O2 has been measured directly (via luminescence at 1270 nm) from both purified cercosporin and fungal cultures (13, 49, 84). The importance of 1O2 in in vivo toxicity has been supported by studies showing that 1O2 quenchers such as carotenoids protect cells against cercosporin toxicity (31, 42). Production of O2 •− has also been confirmed in the presence of reducing substrates (38, 67), and sugar beet plants transformed for high expression of superoxide dismutase (SOD) were more resistant to both cercosporin treatment and infection by C. beticola (133). Extensive studies have documented that cercosporin is a membrane-bound sensitizer and it damages cells by causing peroxidation of membrane lipids. Ultrastructural studies of cercosporin-treated leaf tissue or of Cercospora-infected leaves document membrane damage (127, 128), and rapid ion leakage is the earliest measure of cercosporin toxicity to cells (32, 94). Cercosporin-treated cells accumulate lipid peroxidation products and show a decrease in membrane fluidity, indicating lipid peroxidation (35).
Elsinoë species produce several perylenequinones called elsinochromes (Fig. 1) that share the common perylenequinone structure and differ in side chains (25). These compounds were isolated and characterized in the mid 1950s–1960s (144, 146). Elsinochromes have been isolated from several species, including E. randii and E. fawcettii, as well as from lesions on affected leaves. Elsinochromes have also been reported from species in other fungal genera, including Hypocrella and Stagonospora (25).
As with cercosporin, early studies on the photosensitizing activity of elsinochromes demonstrated toxicity to bacteria that was dependent on light and oxygen (144). A recent study characterized elsinochrome photoactivity on plants (87). Elsinochromes isolated from E. fawcettii were shown to be toxic to cells of both hosts (citrus) and non-hosts (tobacco) when exposed to light. 1O2 quenchers such as the carotenoid bixin and 1,4-diazabicyclooctane and antioxidants such as ascorbate protected against death of citrus protoplasts and cultured tobacco cells and also against necrosis caused by elsinochromes on citrus leaves. Production of 1O2 and O2 •− was demonstrated through the production of cholesterol 5α-hydroperoxide and SOD-inhibited nitroblue tetrazolium reduction, respectively. Elsinochromes were also shown to cause electrolyte leakage from citrus leaves incubated in the light.
As previously stated, the production of so many perylenequinones by fungal plant pathogens suggest a role for these compounds in disease development. The importance of cercosporin and elsinochromes in successful pathogenesis by the producing fungi has been well established. The production of a light-activated virulence product by Cercospora species was, in fact, initially pursued due to extensive observations on the importance of light in development of disease symptoms caused by these fungi. This effect is most evident in crops with a canopy, such as coffee, where it has been long observed that shaded leaves have a fewer lesions than those exposed to bright sunlight (51). Studies of symptom development by C. beticola on sugar beet as well as by Alternaria alternata (another perylenequinone producer) on cotton were also shown to be dependent on high light intensity (18, 117). It is known that Cercospora species produce cercosporin in lesions, and ultrastructural changes in infected leaves (membrane damage) are consistent with cercosporin's mode of action (127). This correlative evidence has been supported by several studies with Cercospora or Elsinoë mutants that are unable to synthesize cercosporin or elsinochromes. Gene knock-out mutants of C. kikuchii for a membrane-bound transporter (cercosporin facilitator protein [CFP]) involved in cercosporin production produce less cercosporin and were significantly reduced in symptom development on their soybean host (17). Knock-out mutants of the corn pathogen C. zeae-maydis for a mitogen-activated protein kinase (MAPK), also impaired in cercosporin production, were unable to produce typical blighting necrotic lesions (121). The recent characterization of the biosynthetic pathways for both cercosporin and elsinochromes has allowed for generation of knock-out mutants for biosynthetic pathway genes. Knock-out mutants of C. nicotianae for the cercosporin polyketide synthase (PKS) produced a limited number of small lesions when inoculated on tobacco in contrast to the large blighting lesions of wild type (23, 39). Elsinoë knock-out mutants for either the elsinochrome PKS or a pathway transcriptional regulator produced a fewer lesions when inoculated on citrus; normal pathogenicity was restored when the strains were complemented with the intact genes (25, 86). These results demonstrate that both cercosporin and elsinochromes are virulence factors which markedly facilitate fungal colonization and the production of necrotic lesions on their respective hosts.
Apoptosis Versus Necrosis
Although it is clear that cercosporin and elsinochromes play a central role in disease development, there is a debate about whether this effect is mediated by direct toxicity or by induction of a programmed cell death response by the host. As noted earlier, some necrotrophic fungal pathogens of plants, which kill the cells of their hosts in order to colonize tissue, do so by inducing the plant host's programmed cell death response against itself (65, 82, 89, 91). Cercospora and Elsinoë species are hemibiotrophs; they are able to initially infect and colonize living tissue, but then switch to a necrotrophic mode of growth where they kill cells in advance of colonization. Thus, they differ from the true necrotrophs that are shown to induce host programmed cell death. Cercosporin's mode of action has long been considered to be due to direct toxicity, as cercosporin directly damages macromolecules, and toxicity to cells is very rapid. Electrolyte leakage, for example, occurs within 1–2 min after exposure of plant cells to 5 μM cercosporin, and 100% of protoplasts are killed within 45 min after treatment (32). Elsinochromes at a concentration as low as 1 μM result in 100% death of cultured tobacco cells within 4 h (87).
There is evidence, however, to support the hypothesis that these compounds induce programmed cell death. Dickman et al. genetically engineered tobacco plants to express antiapoptotic genes, including the human bcl-2, nematode ced-9, and baculovirus op-iap (45). Expression of bcl-2 almost totally inhibited lesion development after the inoculation of transgenic plants with C. nicotianae, and ced-9 and op-iap transformed plants had reduced symptoms. These results suggested that Cercospora fungi require induction of host programmed cell death for symptom development. Studies of perylenequinones being investigated for photodynamic tumor therapy have confirmed apoptotic activity. Hypericin, calphostin C, and hypocrellin have been shown to act through both direct toxicity (necrosis) and induction of apoptosis (50, 108, 134). Mirossay et al. (97) investigated the mode of cell death by hypericin and hypocrellins using a human leukemia cell line. Both compounds showed apoptotic activity on illumination. However, hypocrellin was found to be directly toxic at concentrations that were not sufficient to induce apoptosis. Thus, studies suggest that perylenequinone toxicity may be mediated through both direct toxicity and induction of apoptosis. The relative role of these modes of action in plant disease development is not known.
Biosynthesis of Cercosporin and the Elsinochromes
In contrast to a large body of information on the production of mycotoxins and host-selective toxins of fungal origin, relatively less is known about the biosynthetic pathway and regulation that leads to the formation of perylenequinones. Accumulation of cercosporin and elsinochromes is regulated by a number of environmental and physiological factors (78, 142, 156). Many have similar effects on the production of both compounds, while others have opposite effects. Light is absolutely required for the production of cercosporin and elsinochromes; their production is suppressed under complete darkness. Using ammonium as the sole nitrogen source also suppresses their production. Although elsinochromes are preferentially produced by E. fawcettii when grown under alkaline conditions, pH has little effect on the production of cercosporin toxin by C. nicotianae. Moreover, calcium has been shown to promote cercosporin production by C. nicotianae and to repress the accumulation of elsinochromes by E. fawcettii, further indicating the complexity of perylenequinone biosynthesis.
Cercosporin and elsinochromes contain two highly symmetrical polyhydroxynaphthalenes. Their biosynthetic pathways also share considerable similarities. Biosynthesis of these two compounds was independently investigated by feeding fungal strains with 14C-labeled acetate and analyzing the distribution of radioisotope within the compound purified from culture filtrates with nuclear magnetic resonance and mass spectrometry (20, 107). The carbon skeleton of cercosporin and the elsinochromes is derived from a naphthalenoidheptaketide intermediate. Cercosporin (107) and elsinochromes (20) were separately proposed to be biosynthesized from condensation of acetate and malonate via a fungal polyketide pathway, a process resembling fatty acid biosynthesis. The fungal polyketide pathway condenses one acetyl-CoA and six malonyl-CoAs by iterative decarboxylation (Fig. 4). In each cycle, the malonylketo group is reduced, and two carbons are incorporated to form a linear polyketide, which undergoes a series of oxidation, hydration, and methylation reactions to yield a cyclized polyhydroxynaphthalene. The final product is formed by oxidative coupling of two identical phenols.
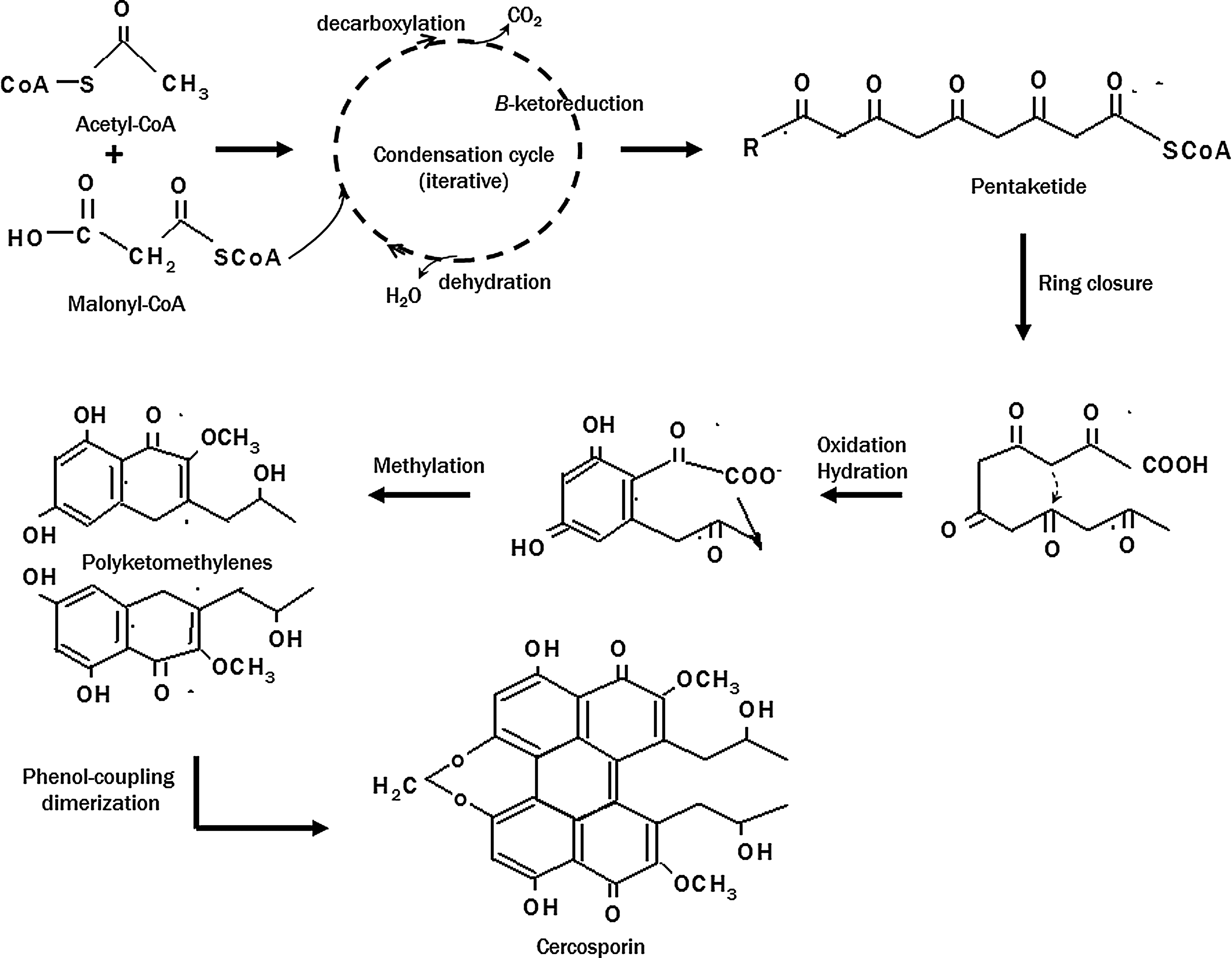
The genes required for the production and accumulation of secondary metabolites are often present as a cluster in the fungal genome (80). Recent studies identified a mini-gene cluster containing eight major genes required for cercosporin biosynthesis from C. nicotianae (36) (Fig. 5). Through a plasmid-tagged mutagenesis strategy (27), several C. nicotianae mutants defective in cercosporin production were recovered, which led to the identification of the first cercosporin biosynthetic gene (CTB1) encoding a fungal PKS. Targeted gene disruption of the CTB1 gene in C. nicotianae abolished cercosporin production, confirming CTB1's role in biosynthesis (23). Subsequently, a chromosome-walking approach was employed to identify the core cercosporin biosynthetic gene cluster that includes genes encoding the PKS, a monooxygenase, a major facilitator superfamily (MFS) transporter, a Zn(II)Cys6 transcription regulator, two O-methyltransferases, and three putative oxidoreductases (Fig. 5). When the CTB8 gene, encoding the transcriptional regulator, was disrupted, expression of all of the core cercosporin cluster genes was abolished or markedly decreased, indicating that CTB8 plays an important role in regulation of the pathway (21). Targeted gene disruption of each of the other genes in the cluster resulted in complete elimination or significant reduction in cercosporin production (21 –23, 44). Re-introduction of functional gene cassettes into each of the respective mutants restored the wild-type level of cercosporin production, thus confirming their function in the biosynthesis.
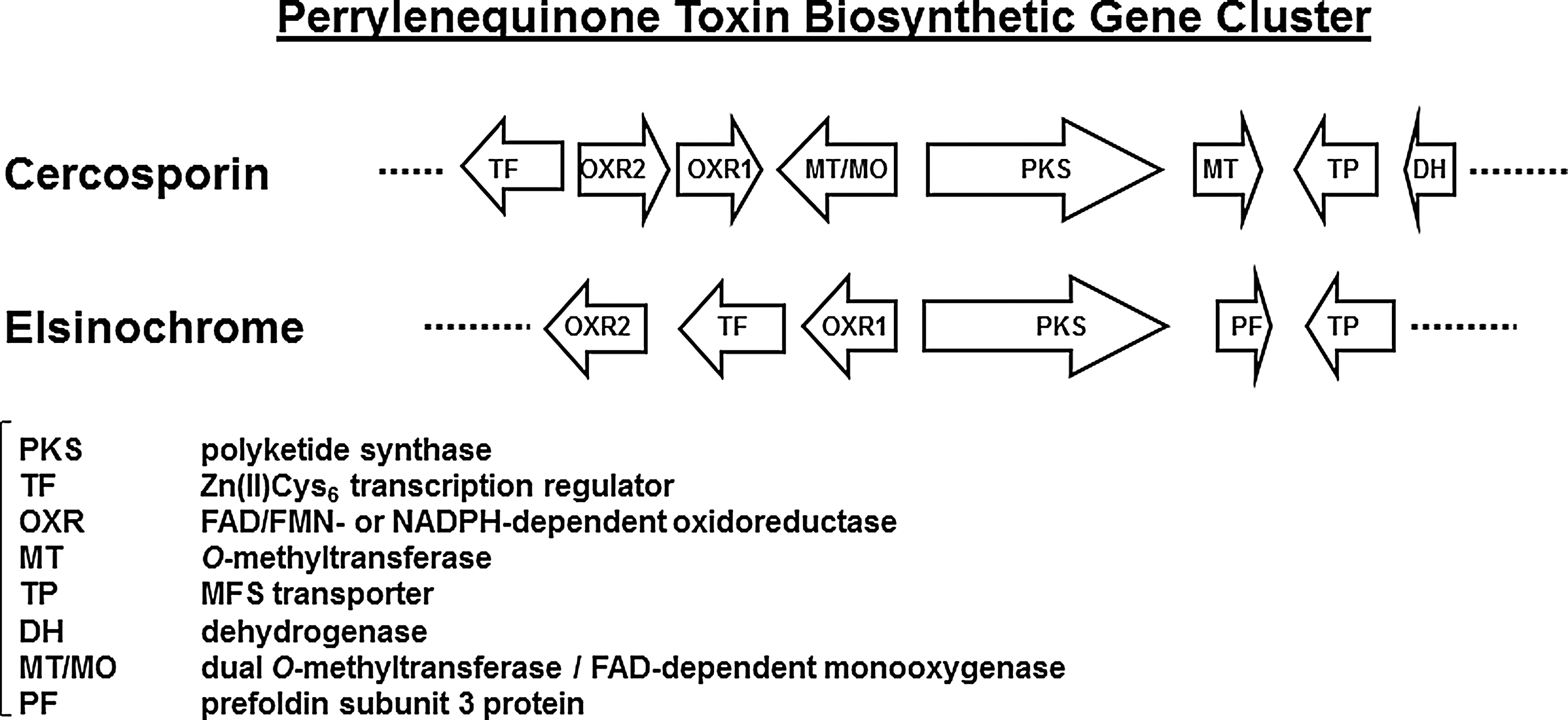
Similar to PKS proteins involved in fatty acid synthesis, the CTB1 PKS contains a keto synthase (KS), an acyltransferase (AT), a thioesterase (TE), and two acyl carrier protein (ACP) domains that function synergistically to build the polyketomethylene backbone of cercosporin. Successive ring closure, oxidation, hydration, and methylation reactions are catalyzed by the proteins encoded by the genes residing in the mini cluster to form the mature cercosporin (Fig. 5). The MFS transporter is likely responsible for cercosporin export (24). Interestingly, other membrane transporters whose coding genes are not found in the cluster have also been shown to play a role in exportation, including the ATP binding cassette (ABC) transporter ABC transporter 1 (ATR1) and the MFS transporter CFP (4, 17). The respective roles of these three transporters in cercosporin export are not known.
In addition to the CTB8 transcriptional activator, expression of the cercosporin biosynthetic genes and production of cercosporin are coordinately regulated by another Zn(II)Cys6 transcriptional activator, cercosporin resistance gene 1 (CRG1) (21). The CRG1 gene, not present in the mini-gene cluster, was originally identified as a gene involved in cercosporin resistance, but disruption mutants also had a reduction in production (26, 28). Accumulation of cercosporin biosynthesis gene transcripts was considerably reduced in a fungal mutant for CRG1. Thus, CTB8 is the pathway-specific transcriptional regulator that regulates the entire cluster of genes involved in cercosporin biosynthesis and accumulation. By contrast, CRG1 has a dual regulatory function, including both cercosporin biosynthesis and resistance.
In some fungi, biosynthetic gene clusters also contain genes encoding proteins for self-protection (2, 111). However, none of the genes in the cercosporin biosynthetic cluster plays a role in self-defense against cercosporin toxicity; mutants of each of the cluster genes retain normal wild-type levels of resistance to exogenous cercosporin and to other 1O2-generating photosensitizers (21, 22).
As stated earlier, the synthesis of elsinochromes in Elsinoë species also occurs via a fungal polyketide pathway. A gene encoding a type-I fungal PKS was cloned and inactivated in the citrus pathogen E. fawcettii and yielded fungal strains that are completely unable to accumulate any detectable elsinochromes (86). The E. fawcettii PKS contains a KS domain, an AT domain, a TE domain, and two ACP domains that are involved in the formation of polyketomethylenes. Sequencing followed by a chromosome walk strategy further identified four additional genes whose products may have a role in the production of elsinochromes in E. fawcettii. They include a Zn(II)Cys6 transcriptional regulator, an MFS membrane transporter, two oxidoreductases, and a putative prefoldin subunit 3 protein (Fig. 5). Although the role of prefoldin in the biosynthesis of elsinochromes remains uncertain, the protein has been shown to function as a molecular chaperone by binding and stabilizing newly synthesized polypeptides for proper folding (138). Since prefoldin is a hexamer containing multiple subunits, additional prefoldin subunits whose coding genes are not located in the gene cluster are likely required to fulfill the chaperone function. Moreover, there is no methyltransferase-coding gene in the biosynthetic gene cluster, even though elsinochromes have four methyl groups at positions C3, C7, C8, and C12. It appears that the identified gene cluster does not contain the entire entourage of genes responsible for the biosynthesis of elsinochromes.
Northern-blot analysis revealed that expression of the elsinochrome PKS is up-regulated on exposure to light, in the presence of higher amounts of glucose, during nitrogen starvation or at alkaline pH, all conditions which are highly conducive to the accumulation of elsinochromes (86). However, accumulation of transcripts of the other genes in the cluster did not always correspond to the conditions conducive for elsinochrome accumulation (29). All putative genes were up-regulated under nitrogen starvation and in response to light. All genes except for the OXR1 gene encoding an oxidoreductase were preferentially expressed under alkaline conditions. Glucose suppressed the gene encoding the Zn(II)Cys6 transcriptional regulator, but elevated expression of the other genes in the cluster. Mutants for the transcriptional regulator were unable to accumulate elsinochromes and exhibited a marked reduction in transcripts of other genes in the cluster. Thus, the Zn(II)Cys6-containing protein acts as a core transcriptional activator and plays an important role in the biosynthesis of elsinochromes.
In addition to the Zn(II)Cys6 transcriptional regulator, genetic analyses revealed that a homolog for the yeast STE12 transcription factor also has a regulatory role in the biosynthesis of elsinochromes (153). The EfSTE12 gene is not present in the biosynthetic gene cluster. In Saccharomyces cerevisiae, STE12 is required for a wide array of developmental and physiological functions and is activated by a pheromone response MAPK pathway (118). Interestingly, an E. fawcettii mutant lacking the EfSTE12 gene shows decreased expression of the elsinochrome cluster PKS gene, but not the gene encoding the cluster Zn(II)Cys6 transcriptional regulator. EfSTE12 apparently activates the biosynthetic genes without regulating the pathway-specific transcription regulator. Moreover, E. fawcettii mutants lacking the PKS or the Zn(II)Cys6 transcriptional regulator are incapable of producing asexual spores (29, 86), implying an important role of elsinochromes in spore formation. However, EfSTE12 is primarily required for the regulation of toxin biosynthesis, but not for spore formation (153).
Cellular Resistance Mechanisms to Perylenequinones and 1O2
The cellular basis of resistance to perylenequinone photosensitizers has been of considerable interest, as engineering crop plants with resistance genes may have utility in reducing the current significant reliance on pesticides to control damaging diseases. Perylenequinones have almost universal toxicity, killing or causing oxidative damage to plants, mice, bacteria, oomycetes, and viruses as well as most fungi (36). A major challenge to identifying resistance is the production of 1O2 as the primary form of ROS by these perylenequinones. Unlike free-radical forms of ROS where resistance mechanisms are well understood, the cellular basis of 1O2 resistance is not well defined and is still being investigated. Free-radical forms of ROS are common products of cellular metabolism, and cells have multiple defenses, such as SOD, catalases, peroxidases, and many others. 1O2, by contrast, is not commonly generated in cells with the exception of photosynthetic organisms, where it is a byproduct of photosynthesis. Much of the information on the cellular basis of 1O2 resistance comes from studies of plants and photosynthetic bacteria. Studies of auto-resistance mechanisms in perylenequinone-producing fungi show many commonalities with the photosynthetic systems and add significantly to the understanding of 1O2 resistance mechanisms.
1O2 quenchers, antioxidants, and antioxidant enzymes
There have been many compounds identified that quench 1O2 (11, 147). Although many of these have been only identified in chemical studies, some very effective quenchers are found in living cells and have been shown to be an effective mechanism of protection. Of the cellular quenchers, carotenoid pigments are highly effective, due to their ability to quench not only 1O2, but also the activated triplet state of photosensitizers (57, 135). Carotenoids are the primary mechanism identified in plants for protection against 1O2 produced as a by-product of photosynthesis and that damages chloroplasts (157). Other 1O2 quenchers found in cells include the amino acids histidine, methionine, and tryptophan, vitamin B6 (pyridoxal phosphate and its vitamers), and some phenols, amines, and peptides (14, 147). Although antioxidant enzymes such as SOD, catalases, and peroxidases are not directly active against 1O2, they can be effective against photosensitizers acting via type I reactions and also against free-radical chain reactions that occur as the result of photosensitizer damage.
Studies of responses of plants to cercosporin have documented the importance of carotenoids and antioxidant enzymes in protection against cercosporin toxicity. The only plant identified thus far with resistance to cercosporin is a wild rice species, “Louisiana red rice,” a weed that has been studied because it exhibits resistance to the Cercospora species (C. oryzae) pathogenic on cultivated rice. Much of the resistance appears to be due to the plant's resistance to cercosporin itself, and this resistance was correlated with higher levels of carotenoids as well as the exclusion of cercosporin from cells (10). In maize, treatment with cercosporin results in the induction of antioxidant enzymes, including catalases and SOD (148). Efforts to engineer plants for Cercospora and cercosporin resistance using antioxidants and antioxidant enzyme defenses have had mixed results. Tobacco cell mutants selected for resistance to paraquat and having elevated levels of catalase and SOD activity were not more resistant to cercosporin (73). By contrast, sugar beet genetically engineered for higher SOD activity was more resistant to cercosporin and also showed increased resistance to C. beticola infection (133). Tobacco plants genetically engineered for elevated levels of vitamin B6 did not show enhanced cercosporin resistance (71).
Studies of quenchers, antioxidants, and antioxidant enzymes in Cercospora resistance to cercosporin support a role for some as defense mechanisms. Exogenously supplied reducing agents ascorbate, cysteine, and reduced glutathione (GSH) protect cercosporin-sensitive C. nicotianae mutants against cercosporin toxicity, although the mutants did not differ in endogenous levels of these compounds or in total soluble or protein thiol content as compared with wild type (77). A study of the Cpd1 protein in yeast, a FAD pyridine nucleotide reductase similar to GSH reductase and thioredoxin reductase, showed increased resistance to cercosporin when the gene was overexpressed in yeast and in tobacco (110, 141). Cercospora fungi produce high levels of β-carotene, and carotenoid-deficient mutants of the fungi Neurospora crassa and Phycomyces blakesleeanus are significantly more sensitive to cercosporin than carotenoid-producing wild-type isolates (42). β-Carotene-deficient mutants of C. nicotianae, created through targeted gene disruption of the gene encoding phytoene dehydrogenase, were not, however, altered in resistance to cercosporin (54). These results suggest that carotenoids function in fungi to protect against cercosporin, but may be just one of many resistance mechanisms in Cercospora. Mutants of C. nicotianae deficient in biosynthesis of vitamin B6, which are also good 1O2 quenchers, are highly sensitive to cercosporin (53). Analysis of cellular levels of vitamin B6 vitamers also showed that C. nicotianae has 2–3-fold higher levels of B6 vitamers than cercosporin-sensitive fungi N. crassa and Aspergillus flavus (71). The fact that B6 is an essential vitamin for fungal growth complicates further characterization of its precise role. In contrast to 1O2 quenchers, there is little evidence for a role of antioxidant enzymes in cercosporin resistance in Cercospora. C. nicotianae does not have higher levels of SOD, catalase, or peroxidase activity as compared with cercosporin-sensitive fungi, and there is also no difference in overall antioxidant activity (34).
Transporters
Another highly effective defense against perylenequinone photosensitizers are membrane transporters, which presumably function by transport of the perylenequinones out of the cell. Both MFS and ABC transporters have been identified in Cercospora species with documented activity against cercosporin. The first transporter identified was CFP, an MFS transporter originally identified in a study of light-induced transcripts in the soybean pathogen C. kikuchii (55). C. kikuchii mutants disrupted for CFP produce less cercosporin, are less virulent on soybean, and are also more sensitive to cercosporin toxicity than is wild type (17). Subsequent studies expressing CFP in a cercosporin-sensitive fungus (Cochliobolus heterostrophus) and in tobacco documented increased cercosporin resistance in both species, as well as increased Cercospora disease resistance in tobacco (136, 137). An ABC transporter, ATR1 was identified in C. nicotianae from a subtractive library between the wild-type fungus and a cercosporin-sensitive mutant deficient for the CRG1 transcription factor required for resistance (see below). As with CFP, atr1-disruption mutants produce less cercosporin and are significantly more sensitive to cercosporin than wild type (4). Interestingly, CTB4, the MFS transporter in the cercosporin biosynthetic cluster (Fig. 5) does not appear to play a role in cercosporin resistance, as mutants disrupted for the CTB8 pathway regulator required for expression of all the other pathway genes are not more sensitive to cercosporin than wild type (21).
Other studies have documented a role for transporters from other organisms in perylenequinone resistance. The yeast ABC transporter Snq2p provided resistance to cercosporin when over-expressed in yeast (141). An MFS transporter from the fungus Botrytis cinerea (Bcmfs1) was shown to provide resistance to cercosporin as well as to camptothecin, a quinone alkaloid produced by the plant Camptotheca acuminata (69). MgMfs1, an MFS transporter from the wheat pathogen Mycosphaerella graminicola, provided increased resistance to cercosporin and other compounds when expressed in yeast (116); M. graminicola disrupted for MgMfs1 became highly sensitive to cercosporin as well as to strobilurin fungicides. Finally, a study of cellular responses that limit the effectiveness of hypericin against cancer cells identified two ABC transporters, MRP1 and BCRP, whose expression was increased in response to hypericin treatment (76). The authors documented the need for inhibitor treatment to allow for sufficient accumulation of hypericin for effective therapy.
Photosensitizer degradation
Enzymatic degradation of photosensitizers is also a documented mechanism of resistance. Insects that feed on photosensitizer-containing plants have evolved mechanisms for degradation of the compounds; for example, a cytochrome P450-mediated degradation of photosensitizing furanocoumarins by swallowtail caterpillars that feed on furanocoumarin-containing plants (85). Several studies have characterized enzymatic degradation of cercosporin. Studies of biological control agents active against C. beticola infection of sugar beet identified a number of Basidiomycete fungi with biocontrol potential. A further study identified the production of laccase, a phenol oxidoreductase, as the basis for control activity and documented the effectiveness of laccase in cercosporin degradation (16). Laccase from Pleurotus ostreatus or Tramete versicolor was shown to degrade cercosporin in vitro and protect Escherichia coli and sugar beet leaves from cercosporin toxicity.
Another study screened 244 bacterial strains from 12 genera and 23 species for cercosporin-degrading activity (99). Isolates of Xanthomonas, Pseudomonas, and Ralstonia (all plant-associated species) were the most active in cercosporin degradation. All 32 isolates of Xanthomonas campestris pv zinniae (a zinnia pathogen) were the most active, degrading more than 90% of cercosporin in assays. The degradation product was identified as xanosporic acid (Fig. 6) (98), which was shown to be non-toxic to tobacco. Complementation of non-degrading mutants of X. campestris pv zinniae identified an oxidoreductase required for cercosporin degradation, and Southern blot analysis showed a correlation between presence of the gene and cercosporin-degrading activity in diverse strains (131). Efforts to engineer cercosporin degradation in non-degrading bacteria and plants through transformation with the oxidoreductase gene were, however, not successful. It is not yet clear whether other enzymes are also required or whether other factors (localization, other substrates, or co-factors) are required.

Reductive detoxification
The most significant mechanism documented for cercosporin resistance in Cercospora species is the ability of these fungi to maintain cercosporin within the cell in a reduced form. In vitro, cercosporin can be reduced by strong reducing agents such as zinc dust or dithionite (84). Reduced cercosporin (Fig. 6) is labile, and readily reoxidizes on aeration or removal from the reducing agents. Reduced cercosporin (assayed as stable methylated and acetylated derivatives) was shown to be a poor generator of 1O2, with quantum yields of less than 20% of that of cercosporin. 1O2 producing activity by reduced cercosporin is particularly poor in aqueous solution, whereas cercosporin's photosensitizing activity is unaffected by the solvent (41). The reduced derivatives are highly fluorescent, with fluorescence intensities of 80–160 times greater than cercosporin, supporting the hypothesis that weak photosensitizing activity is due to dissipation of light energy via fluorescence rather than transfer to O2 for ROS production. Toxicity assays against cercosporin-sensitive fungi and lipid peroxidation assays confirmed that the reduced derivatives were not toxic and were weak agents of lipid peroxidation.
Evidence that Cercospora fungi maintain cercosporin in a reduced form comes from several studies. Fluorescence microscopy of cercosporin-producing cultures using bandpass filters specific for fluorescence emission from cercosporin and reduced cercosporin confirmed that fluorescence emission from within hyphae of Cercospora species is consistent with reduced cercosporin (40) (Fig. 2). Hyphae of cercosporin-resistant Alt. alternata also reduced cercosporin; however, hyphae of cercosporin-sensitive fungi (Asp. flavus, N. crassa) or of Cercospora cultures killed by chloroform vapor, steam, or UV irradiation did not reduce cercosporin. Confocal microscopy showed reduced cercosporin localized in the cytoplasm, an aqueous environment that, as noted earlier, is most inhibitory to 1O2 production by reduced cercosporin (41). Ability of fungi to resist cercosporin and to reduce cercosporin correlates with cell surface reducing power, which was estimated by assaying the ability of fungi to reduce a series of 20 tetrazolium dyes differing in redox potential (123); Cercospora species and Alt. alternata reduced significantly more dyes and dyes with more negative redox potentials than did the sensitive fungi Asp. flavus, N. crassa, or two Penicillium species. The formal redox potential (Eo′) for cercosporin was determined to be −0.16 V versus the standard hydrogen electrode (pH 7.1) (30, 40). This compares to −0.72 V for rose bengal, the only 1O2-generating photosensitizer that shows some toxicity to Cercospora species and which cannot be reduced by Cercospora hyphae (40).
A summary of cercosporin resistance mechanisms identified in Cercospora is shown in Figure 7.

Proteomics of 1O2 resistance in Rhodobacter sphaeroides
The photosynthetic, Gram-negative bacterium Rhodobacter sphaeroides has been extensively studied as a model for photooxidative stress and 1O2 resistance. It is a versatile microbe that is able to grow under either aerobic or anaerobic conditions and has been the subject of many studies focused on prokaryotic photosynthesis (93, 158). Its use for understanding 1O2 and photooxidative stress is due to the fact that 1O2 is the main contributor to light-induced cellular damage in photosynthetic organisms. Thus, these organisms have evolved to both sense 1O2 and remove toxic byproducts and defend themselves against photosensitization [reviewed in refs. (62, 81)].
Many studies have focused on regulatory networks governing 1O2-induced stress and cellular responses in R. sphaeroides and include the role of sigma transcription factors RpoE and RpoHII. RpoE activity induced by exposure to light wavelengths conducive to bacteriochlorophyll sensitization or to the 1O2-generator methylene blue correlated with the transcriptional up-regulation of ∼180 genes predicted to be involved in stress responses and 1O2 protection (5). A mutant lacking RpoE was sensitive to bacteriocidal properties of 1O2, indicating that one or more genes regulated by RpoE are essential for the viability and resistance to 1O2. Proteomic analysis conducted after 1O2 exposure of R. sphaeroides wild type and RpoE mutant strains identified proteins involved in protein synthesis and turnover, and amino acid metabolism and transport (63). The RpoE-mediated signal transduction cascade includes the downstream sigma transcription factor RpoHII (62). A genome-wide search for RpoHII target sequences identified 115 genes preceded by putative RpoHII promoters that were predicted to play a role in energy metabolism, transport, regulatory processes, protein turnover, amino acid metabolism, and stress response (105). In addition, protein synthesis under photooxidative stress of a rpoHII deletion mutant differed from that in wild type and revealed that RpoHII-dependent proteins are involved in (i) GSH-dependent detoxification of reaction products formed from cellular macromolecules which interact with 1O2; (ii) proteins involved in the degradation of peroxides; and (iii) proteins involved in methylglyoxal removal generated by disturbances in carbohydrate metabolism. It has been proposed that a GSH-dependent and -independent 1O2 detoxification pathways controlled by RpoHII exist in R. sphaeroides (62, 106).
In addition to transcriptional regulation by sigma factors, post-transcriptional RNA-mediated regulation plays a role in 1O2 response. The bacterial RNA chaperone (Hfq) binds small RNAs and stabilizes base pairing between them and their targets (63, 101, 159). Its involvement in photooxidative stress resistance was first reported in R. sphaeroides (12). When compared with wild type, bacteria lacking Hfq were more sensitive to 1O2, had lower RpoE activity, and diminished induction of several RpoHII-dependent proteins, indicating complex interactions among these regulatory entities. Proteomic analysis between a wild-type strain and a hfq deletion mutant has been conducted either under control conditions (no 1O2) or after a 60-min exposure to 1O2 (12). A total of 79 soluble proteins were differentially expressed and classified as (i) up-regulated in wild type or the hfq mutant under control conditions or (ii) up-regulated in wild type or in the hfq mutant on 1O2 exposure. In the absence of 1O2-induced stress, most proteins up-regulated in wild type or the hfq mutant were involved in amino acid metabolism or transport, respectively. Under 1O2 stress conditions, many of the differentially expressed proteins between wild type and the hfq mutant were those previously identified as depending on RpoHI, RpoHII, or both (105, 106). Taken together, the studies on regulatory networks demonstrate that the cellular responses triggered by 1O2 stress are diverse and include DNA repair, quinol oxidation, GSH peroxide reduction, 1O2 scavenging, GSH-dependent and -independent detoxification, protein turnover, membrane transports, scavenging of divalent metal ions, methionine sulfoxide reduction, and aerobic respiration (5, 15, 63, 105, 106).
Cercosporin-resistance genes in Cercospora and similarities with R. sphaeroides
The resistance of Cercospora species to cercosporin allows use of this organism as a model to understand the basis of 1O2 and oxidative stress resistance in a non-photosynthetic organism. As with R. sphaeroides, characterization of regulatory components has led to the recovery and characterization of genes important in autoresistance. A forward-genetics approach using a collection of UV-mutagenized cercosporin-sensitive strains of C. nicotianae identified CRG1 belonging to the binuclear zinc cluster family of transcription factors found only in fungi (26, 28, 77, 79). As previously stated, mutants of C. nicotianae lacking CRG1 are strongly inhibited by cercosporin and also show medium-regulated impairments in cercosporin production, suggesting pleiotropic effects of this transcription factor on both production and resistance. Expression of CRG1 is not constitutive and depends on medium composition (156). Sensitivity of crg1-null strains to cercosporin is both medium and pH-dependent (70). The CRG1-specific upstream and downstream regulatory networks and subsequent cellular responses triggered by these environmental cues are yet to be elucidated. However, it has been proposed that signal transduction and regulatory controls affecting cercosporin biosynthesis, including Ca/calmodulin signaling, MAPK signaling, and/or G-protein/cAMP/Protein Kinase A signaling pathways, may, in turn, up- or down-regulate CRG1 (36, 156). A sequence comparison of CRG1 to those of sigma factors RpoE and RpoHII show no common domains; however, analogous to CRG1, different sigma factors involved in stress responses are induced by environmental cues such as nitrogen limitation and starvation.
In an attempt to elucidate the downstream cellular processes regulated by CRG1 and shed light on its role on 1O2 resistance, suppressive subtractive hybridization was used to identify genes differentially expressed between wild-type and crg1-null strains. This subtractive library identified 185 unique expressed sequence tags (ESTs) (80 and 105 up-regulated, respectively, in wild type and the crg1 null mutant) that were classified into different functional categories (70). These studies identified many commonalities with the R. sphaeroides studies described earlier (Table 2). The most striking difference between C. nicotianae wild type and the crg1 mutant was found for energy-related processes. Several ATPase synthases and a cytochrome b were up-regulated in the cercosporin-sensitive crg1 null mutant (Fig. 8). This result is similar to the reported impact of 1O2 exposure in R. sphaeroides on energy metabolism and respiration (105, 106). In addition, proteins are a primary target of 1O2, and proteins involved in amino acid metabolism and transport and protein turnover were identified in R. sphaeroides (12, 63, 105). Similarly, we identified a total of 16 genes differentially expressed between wild type and the crg1-null strain involved in amino acid metabolism (70). One of these genes, a O-acetylhomoserine(thiol)-lyase is involved in protein turnover and shows significant homology to its counterpart in R. sphaeroides, where it is down-regulated in the 1O2-sensitive hfq mutant. In addition, we identified a cysteine synthase B and a disulfide isomerase precursor. Although these proteins lack significant homology to their R. sphaeroides counterparts (a cysteine synthase A and a thiol-disulphide interchange protein, respectively), they have similar roles in cells and exhibit similar expression patterns in both organisms; specifically, the cysteine synthase is up-regulated in both crg1 and hfq; and the putative thiol-disulphide interchange protein is up-regulated in crg1 and induced upon 1O2 exposure in wild-type R. sphaeroides. Cysteine is a potent antioxidant known to provide protection against 1O2, and the up-regulation of production in 1O2-sensitive mutants is expected.
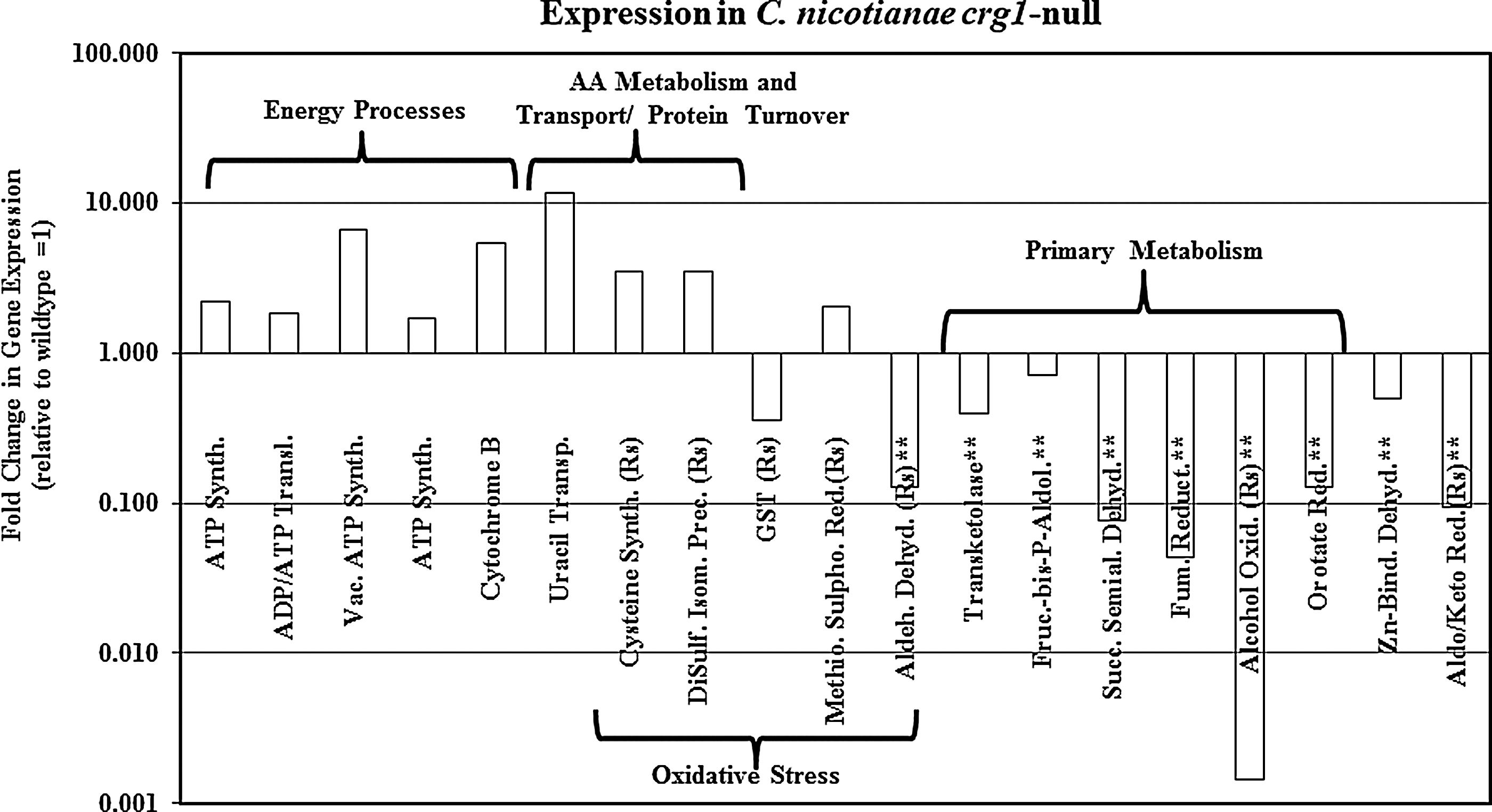
Proteins not found to be significantly homologous between C. nicotianae and R. sphaeroids, but sharing similar nomenclatures and a cellular role.
1O2, singlet oxygen; GSH, glutathione; N/D, not determined; WT, wild type.
As noted earlier, both ABC and MFS membrane transporters have been shown to be involved in cercosporin resistance, similar to the findings of Berghoff et al. (12) that many of the genes up-regulated in the hfq mutant are involved in transport of amino acids, peptides, and polyamines. Based on functional assignment, a total of nine transport-related ESTs were identified in the C. nicotianae subtractive library, including two MFS and three ABC transporters, two of which play a role in cercosporin transport (CFP, ATR1) (4). We also found ESTs homologous to a protein transport protein, a protein translocation complex component, a uracil transporter, and a zinc transport protein that were identified upon transcript enrichment in the crg1-null strain and possibly up-regulated in this mutant. The zinc transport protein may be involved in scavenging of dimetal ions, a mechanism known to protect cells against 1O2 (62).
As noted earlier, proteins involved in GSH-dependent and -independent detoxification of oxidation products have been found to be differentially expressed in R. sphaeroides on exposure to 1O2. Two genes identified in the C. nicotianae subtractive library are likely to be involved in these processes, a glutathione S-transferase involved in GSH-dependent degradation and a putative aldehyde dehydrogenase that shows significant homology to an NADH-dependent aldehyde dehydrogenase from R. sphaeroides involved in GHS-independent degradation. These genes are up-regulated in R. sphaeroides after exposure to 1O2 and down-regulated in the cercosporin-sensitive crg1 mutant. In addition, a methionine sulfoxide reductase involved in protein repair caused by oxidative damage was up-regulated in the hfq-null strain on 1O2 exposure and in C. nicotianae (Table 2). One of the homologues of this enzyme found in the model plant Arabidopsis thaliana has been shown to be up-regulated by oxidative stress, especially by high light (115). Finally, we found an aldo/keto reductase in C. nicotianae that is homologous to an oxidoreductase in R. sphaeroides. This enzyme is down-regulated in the crg1 null mutant and is induced by 1O2 in R. sphaeroides. The specific role of this enzyme in the cell remains unknown (Table 2). Thus, in spite of the fundamental differences between R. sphaeroides (prokaryote, phototroph, and non-pathogenic) and C. nicotianae (eukaryote, heterotroph, and pathogenic) and their life styles, there seems to be significant commonalities on how they respond to 1O2 and cope with its toxicity, including cellular adaptations that involve energy-related processes and respiration, protein transport and turnover, amino acid metabolism, and an induction of general protective mechanisms against oxidative stress.
Finally, our subtractive library contains genes that may play a role in providing reducing capacity required for reductive detoxification of cercosporin as described earlier. Other studies with microorganisms (E. coli, yeast) have demonstrated that the ability to reduce oxidized molecules may constitute the overarching defense mechanism against oxidative stress in microbial organisms (19). Some of the enzymes identified in our subtractive library are directly or indirectly involved in the pentose cycle (transketolase, fructose-bisphosphate-aldolase) and the TCA cycle (succinate semialdehyde dehydrogenase, succinyl-CoA ligase, pyruvate dehydrogenase, fumarate reductase, and alcohol oxidase). These two pathways provide a significant amount of the reducing power present in the cell, and any disruptions would significantly compromise redox homeostasis and ability to provide a reductive environment. The succinyl-CoA ligase and the alcohol oxidase found in our studies are homologous to a succinyl-CoA ligase (α-subunit) and phosphoenolpyruvate carboxykinase from R. sphaeroides, respectively. In R. sphaeroides, these genes were up-regulated in wild type and hfq on exposure to 1O2, respectively; they were down-regulated in the C. nicotianae crg1 mutant (Table 2). Both of these enzymes generate NAD(P)H. Similarly, additional enzymes known to generate NAD(P)H were also down-regulated in the cercosporin-sensitive crg1 strain, including a Zn-binding dehydrogenase, an aldehyde dehydrogenase, a dihydroorotate reductase, and an aldoketo reductase. Efforts to characterize the role of the subtractive library genes in cercosporin resistance in Cercospora through targeted gene disruption are in progress.
Summary
Photoactivated perylenequinones have significant utility in medical and scientific studies, and in nature play a major role in mediating associations between the producing fungi and their host plants. Significant progress has been made in elucidating the biosynthesis and regulation of production as well as the role of these compounds in fungal pathogenesis in plants. Studies have also significantly furthered our understanding of fungal resistance to cercosporin, and Cercospora species provide a model system for understanding 1O2 and oxidative stress resistance in non-photosynthetic organisms. All evidence to date indicates that resistance is multifaceted and complex, and there is not a single mechanism governing resistance to these compounds and to the 1O2 which they generate. Understanding the genes and mechanisms involved in perylenequinone resistance holds promise both for more effective medical use of these compounds and for engineering crop plants for resistance to these agents and the diseases in which they are involved.