
Abstract
Introduction
Reactive oxygen species (ROS) may be generated as byproducts of a respiration process or by certain enzymes such as NADPH oxidase at various sites in cells during aerobic metabolism. Oxidative damage to macromolecules, including proteins, has been implicated in the incidence of a variety of diseases and aging. Among the naturally occurring amino acids, the sulfur-containing amino acids, methionine and cysteine, are most sensitive to oxidation. Free and protein-based methionine residues can be oxidized by ROS to a mixture of two diastereomers, methionine-S-sulfoxide and methionine-R-sulfoxide, because of the prochiral nature of its sulfur atom. Oxidation of methionine may cause significant alterations of protein structure and function; however, organisms have evolved a reductase system to reverse this oxidation (50). Msrs, which are responsible for the reduction of methionine sulfoxide, are present in most organisms, including anaerobic organisms.
Sec Biosynthesis and Incorporation into Proteins in Eukaryotes
Unlike biosynthesis of the other 20 amino acids, Sec biosynthesis occurs on its own tRNA, designated Sec tRNA[Ser]Sec (34, 71). Mammalian genomes encode a single copy of the Sec tRNA[Ser]Sec gene (Trsp), which generates two isoforms: mcm5U (5-methylcarboxymethyluridine; unmethylated form at position U34) and mcm5Um (5-methylcarboxymethyluridine-2′-O-methylribose; methylated form) (21). The biosynthesis pathway of Sec in bacteria was established in the early 1990s (6, 27), but the Sec biosynthesis pathway in eukaryotes has only recently been defined (118). The active selenium donor for the biosynthesis of Sec is monoselenophosphate, which is synthesized from selenide and ATP by the enzyme selenophosphate synthetase 2 (SPS2). Remarkably, SPS2 itself is a selenoprotein (31). There is also a Cys-containing of homolog of SPS2, designated SPS1. This protein cannot synthesize selenophosphate, but may be involved in Sec recycling (109). Sec biosynthesis in eukaryotes (118) is initiated by the attachment of a seryl group to the Sec tRNA[Ser]Sec by seryl-tRNA synthetase, which generates Seryl-tRNA[Ser]Sec. The Seryl-tRNA[Ser]Sec is then phosphorylated by phosphoseryl-tRNA kinase, after which selenophosphate synthase catalyzes the replacement of phosphate with selenium using the active selenium donor, selenophosphate. The resulting selenocysteyl-tRNA[Ser]Sec delivers Sec into the growing polypeptide at the UGA codon. Eukaryotic Sec biosynthesis is summarized in Table 1.
A cis-acting stem–loop structure within selenoprotein mRNA, designated Sec insertion sequence (SECIS) element, is required for Sec insertion into proteins (5). The SECIS element is located in 3′-untranslated regions of eukaryotic selenoprotein genes, whereas it is positioned in the coding region of bacterial mRNAs, immediately downstream of the Sec-encoding UGA codon (Fig. 1). The eukaryotic SECIS elements are characterized by the highly conserved secondary stem–loop structure. The conserved nucleotides (or motifs) required for the function of SECIS include an AA (or CC) sequence (AAR motif) in the apical loop, a quartet of SECIS core (non-Watson–Crick motif) in the stem, and a conserved A (or G) nucleotide preceding the SECIS core in the internal loop (92). There are two forms of eukaryotic SECIS elements (Form I and Form II) distinguished by their structure, in which the difference is an additional short helix present in Form II. The SECIS structure of human MsrB1, corresponding to Form II, is shown in Figure 1A.
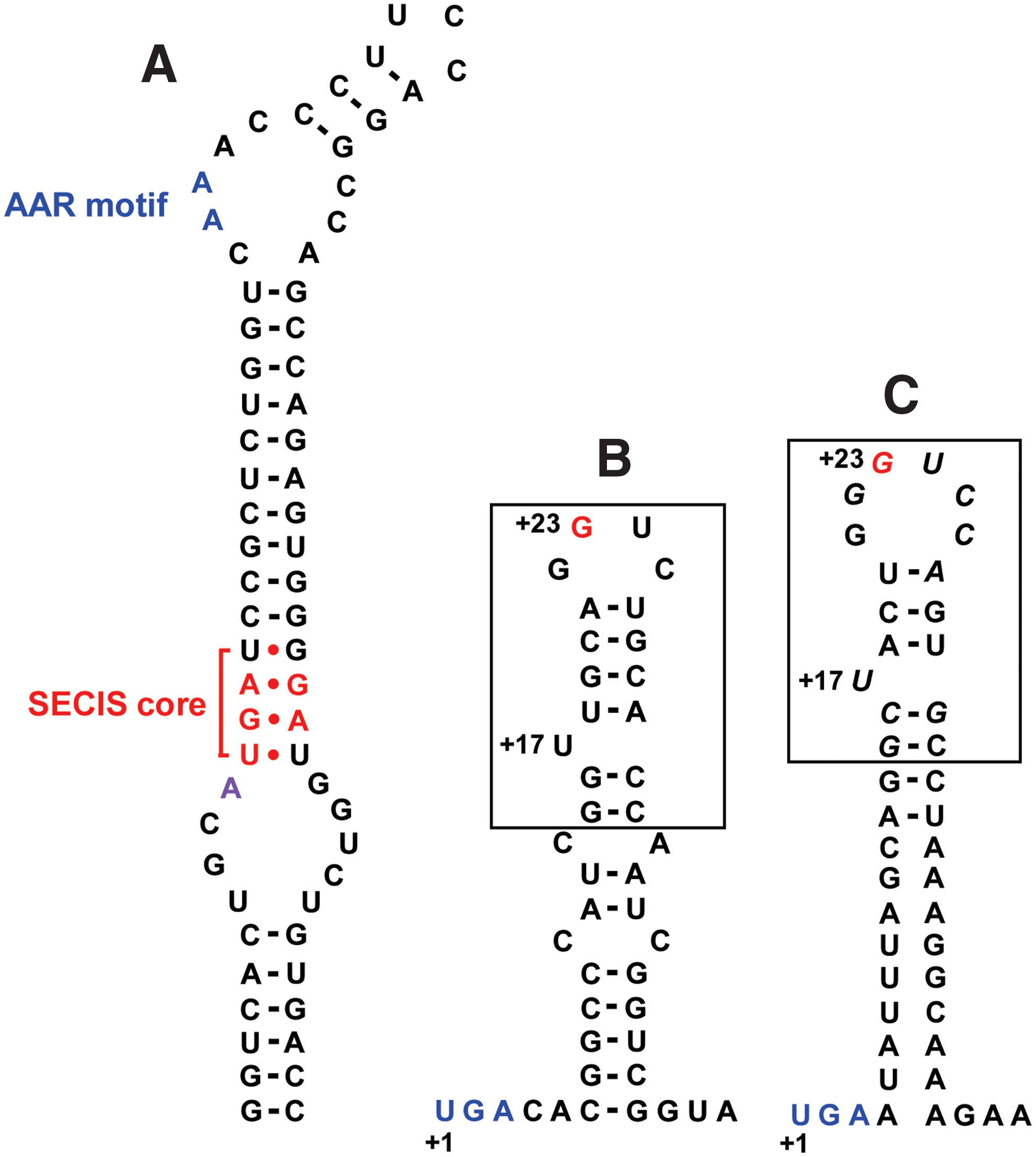
In addition to the Sec tRNA[Ser]Sec and the SECIS element, Sec insertion machinery requires several protein factors, including SECIS-binding protein 2 (SBP2) and Sec-specific elongation factor (eEFSec). Human SBP2 consists of 854 amino acids (17). Interestingly, the first ∼400 amino acids of the N-terminal region are not essential to Sec incorporation (18). The C-terminal region of the protein contains an Sec-incorporation domain (SID) and an RNA-binding domain (RBD). The RBD binds SECIS element and interacts with 28S rRNA. The SID includes several functional motifs required for Sec incorporation. eEFSec exclusively carries the Sec tRNA[Ser]Sec and can interact with Sec tRNA[Ser]Sec, but not Seryl-tRNA[Ser]Sec or canonical aminoacyl-tRNAs (25). Other trans-acting factors, a ribosomal protein L30 and a 43-kDa RNA-binding protein (Secp43), appear to function in the Sec insertion complex (14, 22).
Methionine Sulfoxide Reductases
Msrs catalyze the reduction of free and protein-based methionine sulfoxides to methionine. To accomplish the complete reduction of a mixture of methionine-(R,S)-sulfoxide residues in proteins, organisms have evolved two stereospecific enzyme families. MsrA is specific for reduction of the S-epimer of methionine sulfoxide, whereas MsrB can only act on the R-form. MsrA reduces both free and peptidyl methionine sulfoxide (87). Additionally, MsrA can reduce other methyl sulfoxide compounds, including dimethyl sulfoxide and sulindac (24, 67, 87). In contrast to MsrA, MsrB only acts efficiently on peptidyl methionine sulfoxide and exerts very weak activity toward the free form of methionine sulfoxide (62). For example, MsrB from Escherichia coli shows activity 1000-fold lower than MsrA from the same organism at reducing free methionine sulfoxide (30). It has been reported that MsrB is unable to reduce methyl sulfoxide compounds that are reducible by MsrA (67).
MsrA and MsrB typically exist as separate enzymes in most organisms. However, in several bacteria, including Streptococcus pneumonia and Helicobacter pylori, these two enzymes are directly fused to form a single protein (20, 55, 59). In addition, Neisseria MsrA–MsrB fusion proteins include a Trx-fold domain that may be functionally linked to Msrs. Indeed, it has been reported that the Trx-fold domain of MsrA–MsrB fusion protein from N. meningitidis is a disulfide reductase that can reduce Msrs (117). Fusion proteins containing other domains such as glutaredoxin (Grx) have also been identified (59). Msr genes are present in most organisms, but are absent in some hyperthermophiles and intracellular parasites (20, 59). It is currently not known why certain thermophilic microorganisms (e.g., Thermotoga maritima) do not possess Msr genes. It is possible that a catalyst may be not required for reduction of methionine sulfoxide at high temperatures, or low-molecular-weight thiol compounds may be used for the methionine sulfoxide reduction. Additionally, parasites lacking Msrs may use metabolic pathways of the host to overcome this deficiency. The number of MsrA and MsrB genes varies among organisms. Single MsrA and MsrB genes are present in organisms such as E. coli, yeast, and many animals (e.g., Caenorhabditis elegans and Drosophila melanogaster), while multiple MsrA and MsrB genes are identified in plant genomes such as Arabidopsis thaliana and Chlamydomonas reinhardtii (20). Multiple MsrA and/or MsrB genes may also be present in bacteria. Human and mouse genomes contain a single MsrA and three MsrB genes (46).
MsrA was first discovered three decades ago as an enzyme that can restore biological activity of a ribosomal protein L12 (9). This protein was later shown to stereospecifically reduce the S-form of methionine sulfoxide (104). However, it was not until 2001 that the other Msr family, MsrB, was identified (30). A YeaA protein from E. coli was first found to catalyze the specific reduction of methionine-R-sulfoxide and was then renamed MsrB enzyme (30). Soon after, mammalian selenoprotein R (SelR) was identified as MsrB that contains zinc metal (59). Interestingly, in 2007, a new type of Msr was identified in E. coli in a study involving an MsrA/MsrB double-deletion strain and was designated as fRMsr (77). This enzyme only reduces the free form of methionine-R-sulfoxide and exerts no activity toward the protein-based form. fRMsr contains a GAF domain typically present in cyclic GMP phosphodiesterases, but does not bind cGMP (36). fRMsrs are found in unicellular organisms, including yeast, but not in multicellular organisms (65).
Mammalian Msrs
A single MsrA gene exists in the human and mouse genomes. The most abundant mammalian MsrA form contains a typical mitochondrial signal peptide at the N-terminus. However, subcellular localization experiments have revealed that rat and mouse MsrAs are present in the cytosol as well as the mitochondria (48, 116). The cytosolic MsrA form migrates faster than the mitochondrial form on sodium dodecyl sulfate–polyacrylamide gel electrophoresis, which was found to be a result of modification of the cytosolic form with myristoylation at the N-terminus (42). Structural and functional elements of mouse MsrA have been reported to play a role in the cellular distribution of the protein (48). However, a recent study argued that cytosolic MsrA is generated by alternative translational initiation at Met-21 (42), suggesting a role for dual sites of protein initiation in controlling the cellular location of MsrA. An additional MsrA form generated by alternative first-exon splicing has also been identified (49) and shown to be present in low abundance, but catalytically active, in the cytosol and the nucleus.
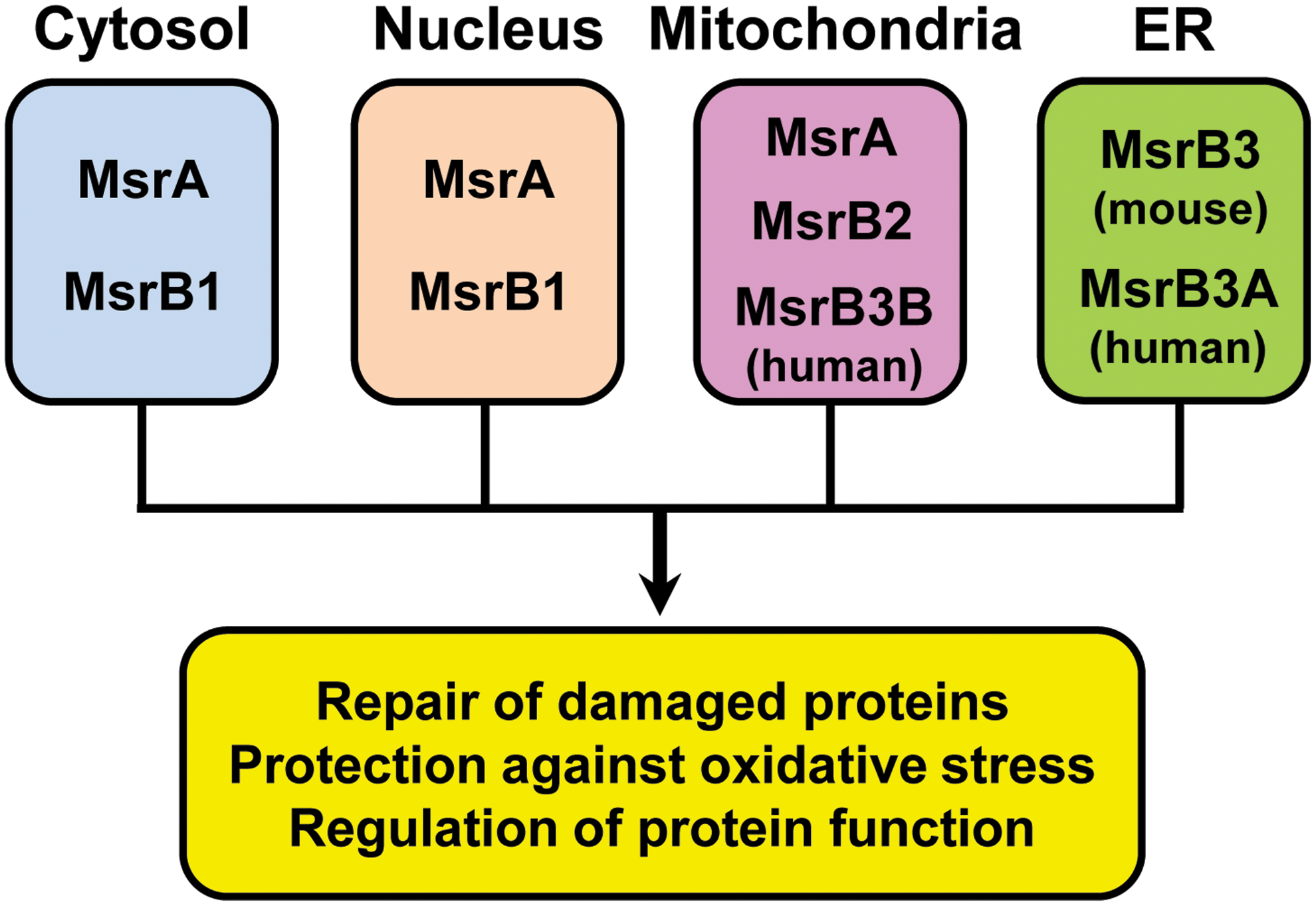
In contrast to a single MsrA gene, there are three MsrB genes (B1–B3) in mammals that code for proteins targeted to different cellular compartments (46) (Fig. 2). MsrB1 was the first selenoprotein identified by computational analysis in 1999 (58, 72). To identify selenoprotein genes from mammalian cells, a bioinformatics tool (SECISearch) that recognizes SECIS elements was developed (58). This program identified SelR as a new selenoprotein with no homology to known proteins. MsrB1 was also independently identified as SelX by another group (72). Subsequent comparative genomic analyses linked the function of SelR to the pathway of methionine sulfoxide reduction, and further biochemical studies verified SelR as a methionine-R-sulfoxide reductase containing a zinc atom (59). SelR was later renamed MsrB1 (46). MsrB1 has Sec as the catalytic residue and localizes to the cytosol and the nucleus. Two other MsrBs, MsrB2 and MsrB3, are Cys-containing isozymes in which catalytic Cys are present in place of Sec. MsrB2 was initially reported as a protein with high similarity to bacterial PilB (37). This enzyme is targeted to mitochondria owing to the N-terminal mitochondrial signal peptide (46). In humans, MsrB3 gives rise to two forms, MsrB3A and MsrB3B, via alternative first-exon splicing (46). MsrB3A contains an endoplasmic reticulum (ER) signal peptide at the N-terminus and an ER retention signal KAEL tetrapeptide at the C-terminus and is targeted to the ER. However, the other alternatively spliced form, MsrB3B, contains an N-terminal mitochondrial signal peptide that targets the protein to the mitochondria. In contrast, there is no evidence for alternative splicing of MsrB3 in mice (45). Instead, mouse MsrB3 contains consecutive ER and mitochondrial targeting signals at the N-terminus with a C-terminal ER retention signal (RAEL sequence). This protein is targeted to the ER, because the function of the mitochondrial signal appears to be masked by the ER signal peptide. The distinct cellular compartmentalization of Msr proteins in mammalian cells is believed to maintain the methionine sulfoxide reduction system in different cellular compartments for repair of oxidatively damaged proteins, protection against oxidative stress, and regulation of protein function (Fig. 2).
Msrs as Selenoproteins
Selenoprotein forms of MsrA and MsrB have been found, but their distribution differs among species (50, 66). The selenoprotein forms of MsrA have been identified in bacteria, algae, and invertebrate animals, but none have been found in vertebrates. The selenoprotein forms of MsrB have been found in animals, including mammals and some invertebrates, as well as in the alga Aureococcus anophagefferens (29). Interestingly, there is no evidence of the occurrence of selenoprotein forms of fRMsr.
Mouse MsrB1 was first engineered to elucidate the role of Sec in catalysis (46). For expression in E. coli, the minimal E. coli SECIS element was introduced immediately downstream of the UGA Sec codon by site-directed mutagenesis (Fig. 1C) (46). The recombinant selenoprotein MsrB1 exhibited an 800-fold higher activity than the corresponding Cys version, indicating that Sec is essential to catalysis of MsrB1. The native selenoprotein MsrB1 form was also characterized (47). The purified MsrB1 selenoenzyme from transfected mammalian cells had a 100-fold higher activity than the Cys mutant form. Additionally, an MsrB from Clostridium sp. (also known as Alkaliphilus oremlandii) OhILAs was engineered to a selenoprotein form (70). This engineered selenoprotein MsrB showed 100-fold higher activity than the Cys-containing wild type. The role of Sec in catalysis has also been studied in MsrAs, and naturally occurring selenoprotein MsrAs from Chlamydomonas and Clostridium exhibited at least 10-fold higher activities than their Cys mutant forms (44, 52). Taken together, these studies demonstrated that the Sec residue confers a higher activity in Msrs as a catalytic advantage, likely in other thiol-dependent oxidoreductases.
There are different sets of active-site features between Sec- and Cys-containing MsrBs (47). The Cys-containing MsrBs contain three highly conserved residues (His-77, Val/Ile-81, and Asn-97; numbering based on the mouse MsrB1 sequence). However, these three residues are modified to Gly-77, Glu-81, and Phe-97, respectively, in Sec-containing forms. The three conserved residues in Cys-containing MsrBs are important for the activity in MsrB2 and MsrB3. The three conserved residues in Sec-containing MsrBs were also found to be critical for the activity of selenoprotein MsrB1. The activity of selenoprotein MsrB1 was highly impaired by substitution of the residues conserved in Cys-containing MsrBs (47). These data imply that Sec- and Cys-containing MsrBs evolved distinct sets of active-site features to maximize their catalytic efficiencies.
Remarkably, a multiple Sec-containing MsrB from Metridium senile has recently been identified by computational analysis (68). This selenoprotein MsrB contains four Sec residues, one of which corresponds to the conserved catalytic Sec. However, M. senile MsrB has no enzyme activity, because its sequence near the active site is not fully conserved. Further biochemical analyses revealed that M. senile MsrB binds iron, with the possibility of binding a sulfur–iron cluster. This study demonstrated a noncatalytic function of Sec in Msr.
Regulation of Selenoprotein MsrB1
Mammalian MsrA and MsrBs are ubiquitous in tissues, but with distinct expression patterns (61, 86, 88). MsrA and selenoprotein MsrB1 are highly expressed in the kidney and liver, and selenoprotein MsrB1 is the main Msr in these organs in mice (26, 88). However, higher MsrB2 and MsrB3 expression levels are detected in heart and skeletal muscle (32, 39).
MsrB1 is one among the selenoproteins that are highly regulated by dietary selenium, such as Gpx1 (88). MsrB1 expression and activity in the liver and other organs of mice are largely reduced by selenium deficiency (88). Additionally, its expression level decreases with age in mice with decreased MsrB activity (88). It has also been suggested that expression of MsrB1 may be regulated at the transcriptional level by Sp1 transcription factor and epigenetic modifications such as methylation (19). A recent study by computational analysis reported that MsrB1 has a different gene structure between rodents and most other mammals, including humans (73). The mouse MsrB1 gene consists of five exons, whereas the corresponding human gene contains four exons. An extra intron (intron 4) in mouse MsrB1 exists in the 3′-untranslational region immediately following the stop codon. Interestingly, this sequence can be either intronized or nonintronized, thereby generating two spliced forms in mice. However, they give rise to identical open reading frames of protein (73).
Knockout mice deficient in selenoprotein MsrB1 have recently been reported (26). MsrB1-deleted mice showed normal development and growth, indicating that MsrB1 is a nonessential selenoprotein. However, it was revealed that selenoprotein MsrB1 significantly contributes to redox regulation in the liver and kidneys (26). Protein oxidation, lipid peroxidation, and oxidized glutathione were elevated in knockout MsrB1 mice, while levels of free and protein thiols were reduced. In addition, the study of MsrB1 knockout mice revealed a 5-kDa selenoprotein that had not been previously described (26). This 5-kDa selenoprotein specifically disappeared in MsrB1 knockout mice and was identified as the C-terminal portion of MsrB1 that contains Sec residue. It was suggested that the 5-kDa form is generated by proteolytic cleavage between Gly-75 and Asn-76. However, the molecular basis for generation of the 5-kDa MsrB1 form and its function are unknown.
Catalytic Mechanisms of Msrs
Although MsrA and MsrB completely differ in structure and sequence, they employ similar catalytic mechanisms characterized by a common sulfenic acid chemistry (7, 8). The general catalytic reaction consists of three steps. A catalytic Cys attacks the sulfur of methionine sulfoxide to form Cys sulfenic acid, with concomitant release of the product, methionine. The catalytic Cys sulfenic acid then interacts with resolving Cys, leading to an intramolecular disulfide bond. Finally, the disulfide bond is reduced by reducing agents, and the enzyme becomes active for the next cycle of the reaction. Trx is generally thought to be a natural reductant for Msrs, whereas dithiothreitol can be used as the in vitro reductant.
MsrA has been well characterized biochemically and structurally (41). The conserved active-site sequence, including catalytic Cys in MsrAs, is GCFW(G/H) located in the N-terminal region. Resolving Cys residues in MsrAs are located in the flexible C-terminal region. MsrAs can be divided into three groups based on involvement of the resolving Cys. Group I MsrAs such as the enzymes from E. coli and Bos taurus have two resolving Cys (3-Cys MsrA) and are the best characterized (78, 114). A thiol–disulfide exchange reaction involving the two resolving Cys occurs, and the resulting disulfide bond is reduced by reducing agents. Group II MsrAs, such as the enzyme from Mycobacterium tuberculosis, include a single resolving Cys (2-Cys MsrA) (113). Group III MsrAs have no resolving Cys (1-Cys MsrA). For example, selenoprotein MsrAs from Clostridium and Chlamydomonas (44, 52) and an MsrA from Synechocystis sp. (111) lack a resolving Cys. Since there is no resolving Cys in 1-Cys MsrA, the catalytic mechanism of this class of MsrA enzyme would be different from that of 2-Cys or 3-Cys MsrAs. Further studies are needed to verify the catalytic mechanism for 1-Cys MsrA regeneration. Three-dimensional structures of MsrAs from organisms, including E. coli, B. taurus, M. tuberculosis, and Populus trichocarpa, have been described (Fig. 3) (79, 98, 113, 114). The central feature of the MsrA structure is an α-/β-plait motif, which includes the conserved active site, GCFWG.
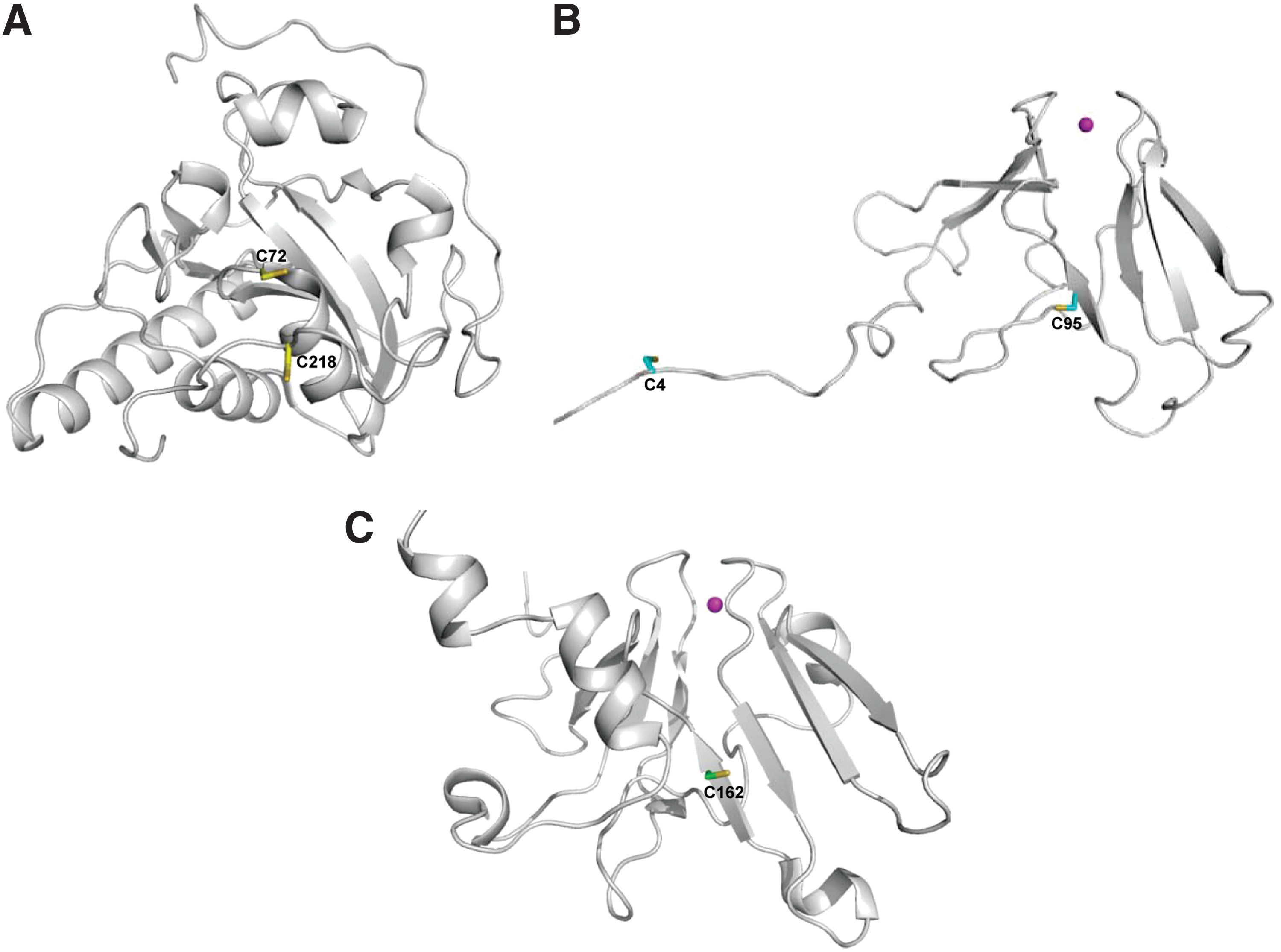
Structures of MsrBs from organisms, including N. gonorrhoeae, Xanthomonas campestris, and Mus musculus, have been reported (Fig. 3) (1, 2, 80, 96). As mentioned above, although the MsrB fold is completely different from that of MsrA, structural comparison analysis of MsrB and MsrA revealed a mirror-like relationship of their active sites (80). In MsrBs, a conserved catalytic Cys is located in the C-terminal region and a conserved resolving Cys is present in the middle region. However, about 40% of MsrB proteins, including all three mammalian MsrBs, do not have this conserved resolving Cys. It was reported that mammalian MsrB2 and MsrB3 do not need resolving Cys, and the Cys sulfenic acid in these enzymes can be directly reduced by Trx (47, 51). Interestingly, in the case of selenoprotein MsrB1, an alternative Cys (Cys-4) in the N-terminal region was identified as the resolving residue (2, 47). Cys-4 residue is conserved in selenoprotein MsrB forms and essential for selenoenzyme MsrB1 regeneration by Trx. It was also reported that an MsrB from X. campestris uses an N-terminal-resolving Cys to reduce the sulfenic acid intermediate (96). Notably, many MsrBs, including all three mammalian enzymes, are metalloenzymes that contain a zinc atom. The zinc is coordinated by two CXXC (X, any amino acid) motifs. Mutation of any zinc-coordinating Cys led to complete loss of metal and catalytic activity in Drosophila MsrB (60). It has been suggested that zinc plays a structural function in metalloprotein MsrBs rather than being directly involved in the catalytic function (60, 91).
A recent study reported the first structure of the selenoprotein MsrB1 determined by high-resolution NMR spectroscopy (2). The overall MsrB1 structure consists of only a β-fold with no α-helix, and is characterized by a highly flexible N-terminal region (Fig. 3). This NMR study verified that resolving Cys-4 can interact with catalytic Sec-95 to form an intramolecular selenenylsulfide bond, as previously suggested.
Reducing Systems for Regeneration of Msrs
The Trx and Grx systems are two major redox systems that maintain cellular redox homeostasis (74, 75). The Trx system is composed of NADPH, Trx, and Trx reductase, while the Grx system is comprised of NADPH, glutathione (GSH), GSH reductase, and Grx. As mentioned above, Trx has long been considered an in vivo reductant for the regeneration of Msrs. However, it has been reported that Trx is unable to reduce certain 1-Cys MsrAs. For example, it cannot reduce selenoprotein MsrAs from Clostridium and Chlamydomonas and their Sec-to-Cys mutants (44, 52). These data suggest that there must be an alternative reductant for these non-Trx-reducible MsrAs. Trx and Grx are thiol–disulfide oxidoreductases that are characterized by a common structural fold known as the Trx fold and can compensate for each other's functions to a large extent. Thus, Grx is predicted to be a possible alternative for the reduction of MsrA forms that are not reducible by Trx. Indeed, it was found that Grx can serve as a reductant for 1-Cys Clostridium MsrA (54). Moreover, Grx can reduce 3-Cys mouse MsrA containing resolving Cys with reduction ability comparable to that of Trx (43). Grx was also found to reduce 1-Cys MsrA more efficiently than 3-Cys MsrA (43).
Grx can also be used for the reduction of MsrB. It was previously reported that Grx was able to reduce a plastidial MsrB with substrate specificity (112, 115), only acting on the 1-Cys form, but not the 2-Cys form. However, a recent study reported that Grx can reduce 2-Cys MsrB as well as the 1-Cys form (43). Furthermore, that study suggested that Grx may serve as a more efficient reductant for 1-Cys MsrB than Trx, whereas Trx was a better reductant for the 2-Cys form than Grx. Future studies should be conducted to determine whether Grx can act as a reductant in vivo for Msrs. The reducing systems for regeneration of Msrs are summarized in Figure 4.

Additionally, zinc-containing metallothionein (a low-molecular-mass cysteine-rich protein) has been shown to serve as a reductant for both MsrA and MsrB (101), while selenium compounds such as selenocystamine have been found to act as reducing agents for MsrBs (100).
Use of Sec in Msr Reactions
It is well known that Sec provides selenoprotein oxidoreductases, including Trx reductase and Msrs, with higher catalytic efficiency than their Sec-to-Cys mutant forms (50, 69). However, there are also Cys-containing orthologs of selenoprotein forms with catalytic efficiencies comparable to those of Sec-containing forms. It has been shown that the selenoprotein MsrB1 has a similar activity to the non-selenoproteins MsrB2 and MsrB3 (47), although the Sec residue is essential for its catalytic activity. In general, selenoproteins have evolved via the replacement of Cys with Sec. However, the evolution of selenoproteins is a very complex process that should balance enhanced catalytic efficiency conferred by the Sec residue with limitations such as the dependence on selenium and prior occurrence of the Sec insertion system. Previous studies suggest that the utilization of Sec in MsrB enzymes may depend on a balance between the catalytic advantage of higher activity and the disadvantage of less regeneration efficiency provided by this residue (70).
A recent study has provided an interesting reason for Sec utilization in the Msr system (54). Specifically, selenoprotein Grx was found to be able to efficiently reduce a Clostridium selenoprotein MsrA, but not the Cys version of selenoprotein MsrA: tandem use of Sec in the selenoprotein MsrA regeneration by selenoprotein Grx. This high specificity for MsrA reduction could explain why Sec is utilized in Grx, providing a novel example of the use of Sec in biological systems.
Physiological Roles of Msrs
Methionine oxidation/reduction has been implicated in a variety of biological and pathological processes, including cellular signaling, oxidative stress, and aging (50, 66). The primary role of Msrs is to repair oxidatively damaged proteins. The protein repair role of Msrs has been supported by a recent study that showed that they more efficiently reduce unfolded oxidized proteins than folded proteins (110). Msrs also function as antioxidants to protect against oxidative stress. Some surface-exposed methionine residues may be oxidized without any impact on protein function. Thus, the cyclic methionine oxidation/reduction is proposed to be an important antioxidant defense mechanism to scavenge cellular ROS (81). Finally, Msrs are able to regulate the function of proteins by modulating specific methionine sulfoxide residues involved in activation or inactivation of proteins. Several target proteins regulated by Msrs are described below.
A number of studies have demonstrated that MsrA and MsrB function as antioxidant enzymes. In experiments of gene overexpression or deletion in microorganisms, including S. cerevisiae, MsrAs were found to protect against oxidative stress (85, 107). MsrAs were also found to protect various mammalian cells, including fibroblast, lens, neuronal, and skin cells, against oxidative stress-induced cell death (40, 89). Overexpression of bovine MsrA in fruit flies led to significantly increased survival under oxidative stress conditions when compared to control flies (99). Moreover, MsrA knockout mice were more susceptible to paraquat-induced oxidative stress than wild-type mice (84). Recent studies reported that MsrA protects against ischemia/reperfusion (I/R) injury in the heart and kidneys of mice (53, 122). In addition, the antioxidant role of MsrBs has been characterized. Specifically, the knockdown of each MsrB gene in human lens cells increased oxidative stress-induced cell death (82), while overexpression of MsrB2 protected cells against oxidative stress (10). Furthermore, heterologous expression of MsrB2 or MsrB3 in yeast protected cells against oxidative stress (63). Recent studies revealed that overexpression of either MsrB1 or MsrB3 increases resistance to paraquat-induced oxidative stress in Drosophila (76, 105). It should be also noted that fRMsr can function as an antioxidant in yeast cells (65).
Substrate Proteins of Msrs
Oxidation of methionine residues may impair biological activities of proteins if they are involved in the protein function, whereas the reduction of methionine sulfoxides by Msrs can restore these activities. As mentioned above, MsrA was first identified as an enzyme that could restore the activity of ribosomal protein L12 (9). Other proteins previously identified as substrates for MsrA include α-1-proteinase inhibitor, calmodulin, apolipoprotein A-I, inhibitor of Kappa B-alpha, and the shaker potassium channel, whose functions are damaged by methionine oxidation and restored by MsrA (90). Interestingly, it was reported that methionine oxidation can also activate the biological activity of protein (23). Calcium-/calmodulin-dependent protein kinase II (CaMKII) is activated by calcium, but can also be activated in an ROS-dependent manner when oxidized at Met-281 and Met-282 in the regulatory domain of the protein. MsrA was found to reverse the activation of CaMKII caused by oxidation (23). Additionally, MsrB1 was recently found to be capable of recovering TRPM6 magnesium channel activity under oxidative conditions (12).
Involvement of Msrs in Methionine Metabolism and the
Trans
-Sulfuration Pathway
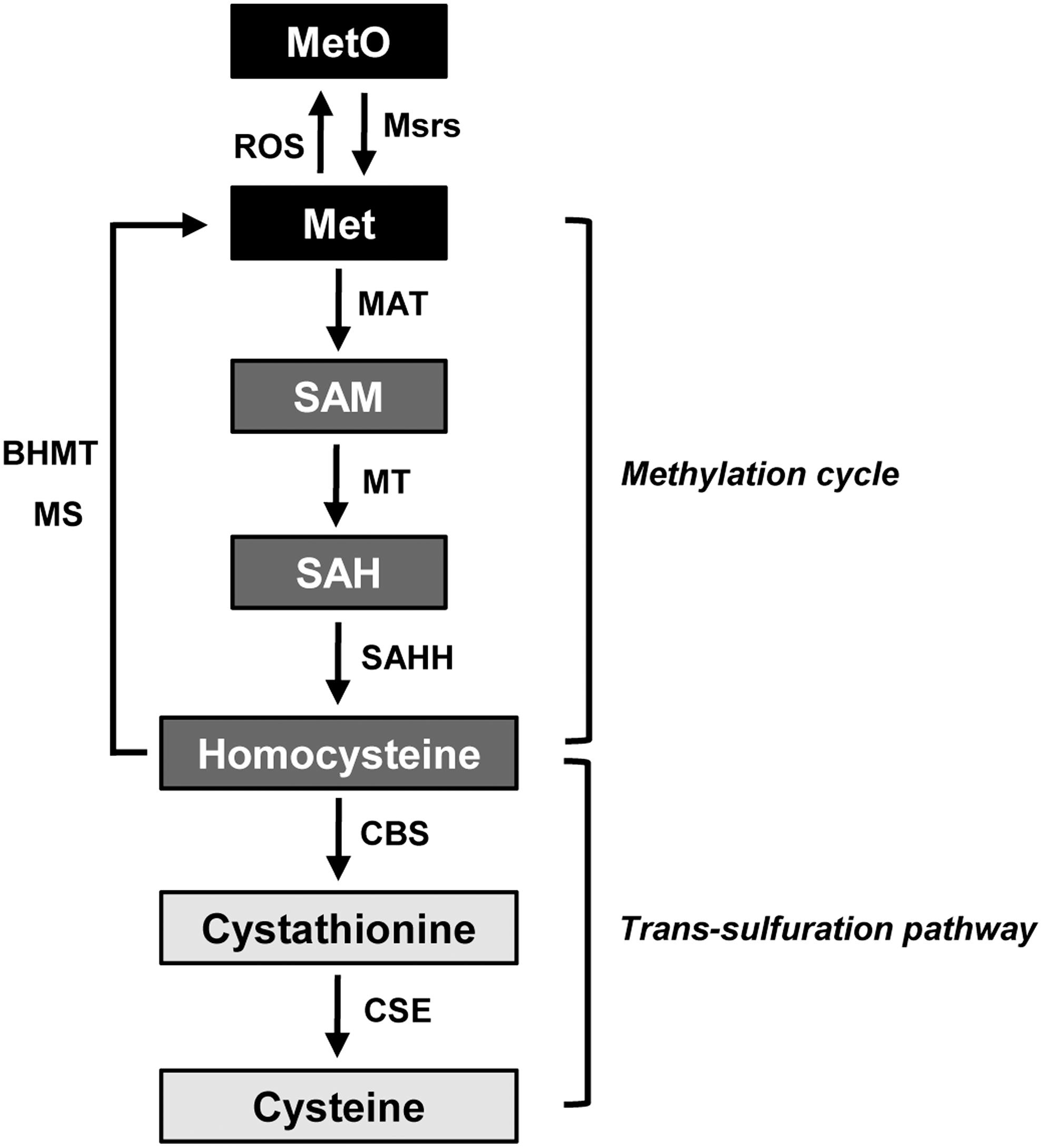
Methionine is an essential amino acid in mammals that is a key molecule for supporting the methylation cycle involving S-adenosylmethionine and for the biosynthesis of cysteine (Fig. 5). Homocysteine is generated as an intermediate amino acid during methionine metabolism. Specifically, it is metabolized to cysteine by the consecutive actions of the two pyridoxal-5′-phosphate-dependent enzymes, cystathionine β-synthase (CBS) and cystathionine γ-lyase (CSE), in the trans-sulfuration pathway. It has been reported that restriction of methionine reduces CBS levels, leading to downregulation of the trans-sulfuration pathway (95). Cysteine is a substrate of both CBS and CSE enzymes to generate hydrogen sulfide (H2S) (103), which has been recognized as a gaseous physiological molecule with potent antioxidant and anti-inflammatory effects (3, 103, 120). Recent studies have demonstrated the cytoprotective role of H2S in a variety of organs, including the heart, brain, intestine, and kidney, against I/R injury (11, 97, 119). Therefore, it is plausible that Msrs may be involved in methionine metabolism, regulation of the trans-sulfuration pathway, and H2S production.
A recent study of MsrA gene-deleted mice provided the first in vivo evidence of the involvement of MsrA in the regulation of methionine metabolism and the trans-sulfuration pathway (53). Concentration of homocysteine in the plasma of MsrA knockout mice was found to be significantly lower than that in wild-type mice. Consistent with the reduced homocysteine levels, plasma H2S levels were also significantly lower in MsrA deletion mice than wild-type mice. The kidney I/R injury significantly decreased the levels of plasma homocysteine and H2S in both MsrA wild-type and knockout mice; however, these reductions appeared to be more profound in MsrA knockout mice than wild-type mice. The I/R was also found to induce significant reductions in H2S and homocysteine in the kidneys of MsrA knockout mice when compared to wild-type mice. Overall, this study showed that MsrA can modulate the expression and activity of the CBS and CSE enzymes in the kidneys, thereby regulating the production of H2S. It will be interesting to determine if MsrB is involved in methionine metabolism and the regulation of the trans-sulfuration pathway similar to MsrA.
The involvement of Msrs in methionine metabolism has also recently been supported by another study, which tested the hypothesis that Msrs provide methionine for growth in mice (121). Wild-type, single MsrA or MsrB1 deletion, and double MsrA/MsrB1 deletion mice were fed either methionine-depleted (0.1% methionine) or methionine-sufficient (0.45% methionine) diet. Since methionine is an essential amino acid in mammals, mice of all genotypes showed retarded growth when raised on the methionine-depleted diet. The growth retardation was significantly greater in the double knockout mice than in wild-type mice; however, there were no significant differences in retardation between the single-MsrA or MsrB1 knockout and the wild-type mice. Overall, this study revealed that Msrs can supply methionine for growth in mice when methionine intake is limited through regulation of methionine metabolism.
Methionine Sulfoxide Reduction and Aging
One of the prominent aging theories is the free radical (oxidative stress) theory (4, 33), in which the accumulation of oxidatively damaged proteins with age is considered a major cause of aging. Msrs have been implicated in regulation of the aging process of organisms due to their functions as protein repair enzymes and antioxidants. Reduced expression levels of MsrA and MsrB2 were observed in senescent WI-38 fibroblasts (94), and decreased total Msr activity was observed in aged rat livers and kidneys (93). Overexpression of bovine MsrA and Drosophila homolog can increase fruit fly lifespan (16, 99). Conversely, knockout of MsrA in yeast and nematodes shortens the lifespan of these organisms (56, 83). However, the effect of MsrA deletion in mice is controversial. A former study reported that deletion of MsrA decreased lifespan in mice by 40% (84), whereas a later study showed that MsrA knockout in mice did not alter lifespan (102).
In the case of MsrB, its deletion or overexpression in yeast has no significant effect on the lifespan under normal conditions (56). However, overexpression of MsrB leads to increased lifespan of yeast under caloric restriction conditions. Recent studies reported that overexpression of mouse cytosolic selenoprotein MsrB1, mitochondrial MsrB2, and Drosophila cytosolic MsrB had no influence on the lifespan of fruit flies (105, 106). In contrast, the expression of human MsrB3A, which is an ER-type of MsrB, was recently reported to significantly extend the lifespan of fruit flies (76). Moreover, MsrB3A overexpression significantly delayed onset of the age-related decline in locomotor activity and fecundity, similar to MsrA overexpression. This study suggested that methionine-R-sulfoxide reduction in the ER, but not in the cytosol or mitochondria, can be a determinant for regulating the fruit fly lifespan (76). Furthermore, MsrB in the ER played an important role in protecting against ER stress (64).
There have been a few mechanistic studies of the roles of Msrs in the aging process. The transcription factor FOXO3a or its homolog directly activates the MsrA gene in human cells, nematodes, and fruit flies (16, 83). Additionally, MsrA overexpression in Drosophila cells leads to activation of FOXO with increased nuclear translocation (16). These data imply that there is a positive feedback loop mechanism between MsrA and FOXO that prolongs the lifespan of Drosophila. A recent study revealed that MsrA can regulate normal cell proliferation (15). Specifically, depletion of MsrA inhibited cell proliferation, while its overexpression did not have a positive effect on cell growth. MsrA deficiency led to elevated acetylation levels of p53, thereby activating p21, and causing cell cycle arrest. This regulation of the p53-p21 pathway by MsrA may be another potential mechanism for involvement of MsrA in the aging process.
Concluding Remarks
Msrs constitute an important defense mechanism against oxidative stress and have been implicated in a variety of physiopathological processes, including cellular signaling and the aging process. Selenium is important in the Msr system, and the Sec residue is essential for catalysis of selenoprotein forms of Msrs. Additionally, recent studies have provided important insights into functions of Msrs involving redox signaling, cell proliferation, and methionine metabolism. The effects of mammalian MsrBs on the regulation of aging have been addressed with regard to their locations using a Drosophila model, and the ER-type of MsrB has been shown to be critical to fruit fly aging. Furthermore, recent progress in characterizing a novel reducing system for Msrs and selenoprotein Msr forms has provided critical insights into the biochemistry of Msrs and the reasons for Sec utilization in the Msr system. Future studies will seek to understand the mechanistic functions of Msrs in redox signaling, the aging process, and metabolism. There will also be continuous efforts to explore new cellular targets of Msrs and the identity of the selenoproteome of Msrs will be further investigated, together with characterization of the identified selenoprotein forms.