
Abstract
Introduction
It is important to note that in addition to its role as a light energy collector and converter, the photosynthetic apparatus also acts as a sensor that reconfigures its components in response to changes in its environment and metabolic needs. Photosynthetic organisms are continuously subjected to the changes in environmental conditions such as light quality and intensity, temperature, water availability, and nutrients. These changes impact the redox poise of the electron transfer chain, especially at the level of the PQ and FD/TRX pools. Both the redox state of these compounds and the pH gradient across the thylakoid membrane are sensed by the photosynthetic machinery, which is then able to trigger an appropriate response for restoring the proper redox poise.
Changes in irradiance profoundly affect the photosynthetic machinery. These changes can occur within seconds or minutes due to clouding or movement of leaves or within hours, days, or months because of the daily and seasonal changes. Moreover, changes both in the intensity and in the spectral quality of light with an enrichment in the far-red region occur under the canopy in forests and crop fields.
The light-harvesting system of PSII plays a key role in this adaptation. This system acts both as a light energy collector and dissipator under certain conditions. When the absorbed light exceeds the capacity of the photosynthetic apparatus, the excess energy is dissipated as heat through a mechanism called nonphotochemical quenching (NPQ) (36, 105). NPQ is triggered through acidification of the lumen under strong light and results in conformational changes in the antenna systems that lead to quenching of fluorescence and heat dissipation. Over-reduction of the electron transport chain and photodamage are thereby prevented. In contrast, when the absorbed light energy is limiting, and under conditions where the two photosystems are unequally excited, the system readjusts by redirecting a part of the excitation energy toward the photosystem that is limiting. This reallocation involves both a short-term response, called state transitions, and a long-term response (LTR) in which the stoichiometry of the photosystems is adjusted so as to optimally match the new environmental conditions. State transitions are mediated through protein kinases and protein phosphatases, some of which are regulated by the redox state of the PQ pool and of the FD/TRX system (72, 135). Changes in the redox state of the photosynthetic electron transport chain can also induce changes in both nuclear and chloroplast gene expression through signaling chains that are still poorly understood (48, 74). How are these kinases and phosphatases connected with each other within chloroplast signaling chains? How do they participate in light acclimation, in the maintenance of the PQ-redox poise, in the control of thylakoid membrane folding and protein turnover, and in the adjustment to metabolic needs (ATP/NADPH)? How do they mediate the changes in the redox state of the photosynthetic electron transport chain into changes in chloroplast gene expression? The aim of this review was to provide some answers to these questions. Earlier work on this topic is covered in several recent reviews (7, 39, 48, 74, 90, 100, 115).
Electron Transport Pathways
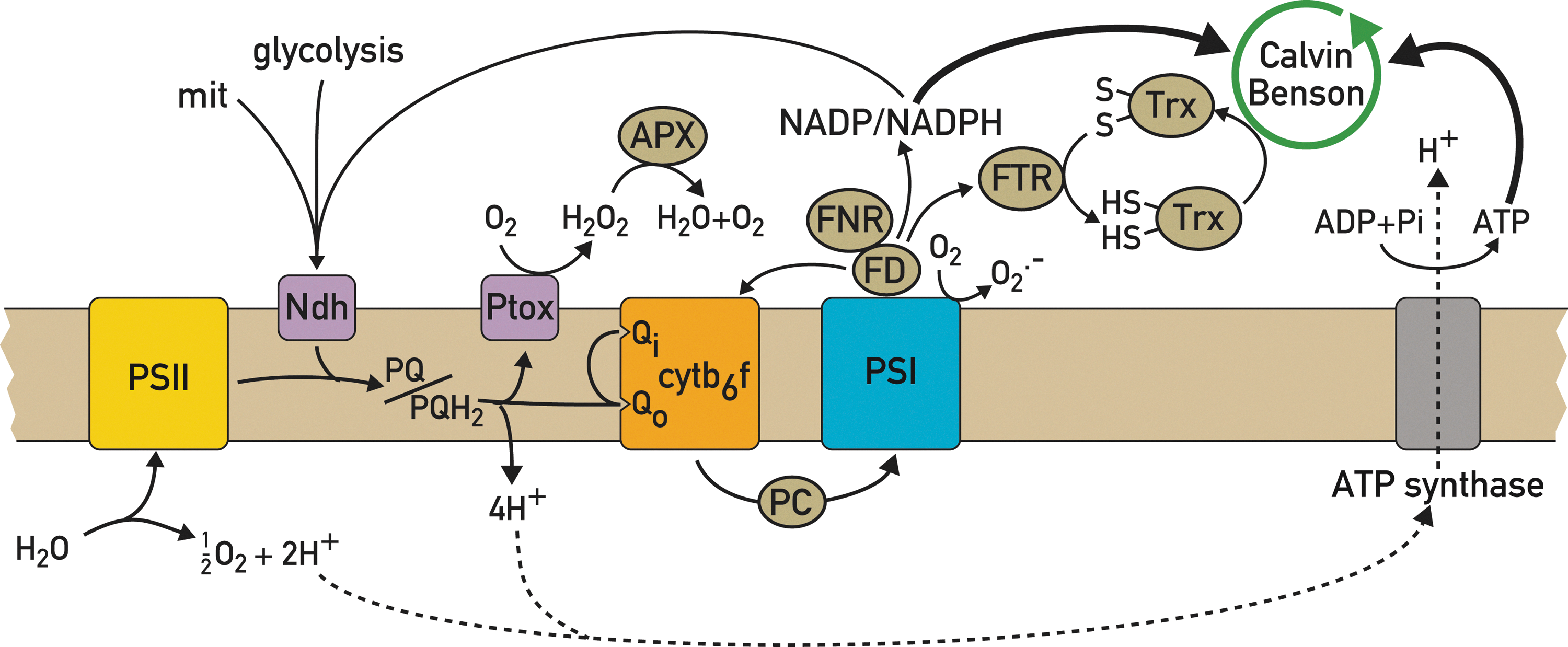
The redox state of the PQ pool and the FD/TRX system depends to a large degree on the relative activities of the major photosynthetic electron transport pathways involved in LEF and CEF. The choice between these pathways occurs downstream of PSI where FD, the terminal acceptor of PSI, transfers its electrons to NADP+ and subsequently either to the Calvin–Benson cycle or back to the PQ pool and Cytb 6 f. Besides its well-established role in cyanobacteria, unicellular algae, and C4 plants, CEF is also important in C3 plants, especially under specific stress conditions of low CO2, high light, elevated temperature, and during dark-to-light transitions. Under these conditions, CEF leads to increased acidification of the thylakoid lumen and thereby to more ATP synthesis and higher NPQ, which downregulates PSII and thus prevents over-reduction of the electron transfer chain. CEF has been proposed to follow several different routes. The first pathway occurs through electron transfer from FD to NADP+ catalyzed by FNR and subsequently to the NAD(P)H dehydrogenase complex and the PQ pool. Because of the low abundance of the NAD(P)H complex, this pathway can only contribute in a modest way to ATP production (21). Since FNR is localized both in the stroma and on the thylakoids, it has been proposed to be involved in the partitioning between CEF and LEF. The second route for CEF involves a direct electron transfer from FD to PQ catalyzed by a FD-PQ oxidoreductase (25), which has however not yet been identified. Because this pathway is sensitive to antimycin, and because antimycin does not bind to Cytb 6 f, it was thought that the enzyme is sensitive to this drug (13, 59). Finally, a third possibility is that FD transfers its electrons to PQ through cytc’, a newly discovered heme in the Cytb 6 f complex (60, 118, 125).
Because a decrease in CEF is expected to diminish proton pumping into the thylakoid lumen and hence NPQ, this property was used in a mutant screen for Arabidopsis mutants deficient in NPQ. In this way, pgr5 was found, a mutant that lacks the thylakoid protein PGR5, and in which both FD-dependent PQ reduction and NPQ are decreased (78). The precise role of PGR5 is not well understood as conflicting results have been reported on its importance for CEF. In one study, pgr5 was crossed to crr2, a mutant deficient in NDH dehydrogenase. The pgr5-crr2 double mutant was strongly affected in its growth, suggesting that PGR 5 plays an important role in CEF (78). However, other studies indicated that PGR5 has a regulatory role in controlling the switch between LEF and CEF based on the observation that in the pgr5 mutant, the ability of CEF to compete with LEF is diminished (81). Another protein, PGRL1 was shown to play a role in the antimycin-sensitive CEF, because its loss as in the case of PGR5 leads to perturbations in CEF (31). In Chlamydomonas, PGRL1 is also involved in iron-induced remodeling of the photosynthetic apparatus (88). PGR5 and PGRL1 interact with each other and form a complex that is most likely involved in CEF (31).
A large supercomplex was characterized in Chlamydomonas that contains PSI and its light-harvesting system, the major trimeric light-harvesting complex (LHC) II and the minor monomeric CP29 and CP26, Cytb 6 f, and PetO, a subunit of Cytb 6 f uniquely found in Chlamydomonas, PGRL1, and FNR, but not PGR5 (56). This complex plays a major role in CEF as revealed by spectroscopic measurements that show that upon illumination, reducing equivalents produced by PSI are transferred to Cytb 6 f, and reducing equivalents from Cytb 6 f can rereduce oxidized PSI. The identification of this complex strengthens the view that FD can transfer its electrons directly to Cytb 6 f and suggests that the FD-PQ reductase may act through the concerted action of PGRL1, PGR5, and FNR. The location of this supercomplex remains to be determined, although it is clearly separated from PSII (56). The existence of this supercomplex is also in agreement with a model in which CEF can operate independently from LEF, which would avoid competition between CEF and LEF, as well as perturbation of the redox poise of CEF components by reduced LEF components. A supercomplex of this sort has not yet been isolated from land plants, although complexes comprising PSI, LHCI, and LHCII have been detected by blue native gel electrophoresis (87) and by electron microscopy (67).
Under conditions that could induce over-reduction of the electron transport chain and that could potentially compromise electron flow, pseudo-CEF occurs whereby electrons are transferred from water to PSI with molecular oxygen as an alternative electron acceptor, a process called the Mehler reaction. It leads to the formation of superoxide, which is converted by superoxide dismutase and catalase to water and oxygen. This water–water cycle could in principle restore the redox poise under conditions of over-reduction of the electron transport chain (84) by restoring CEF and ATP production for the Calvin–Benson cycle that would in turn oxidize NADPH and re-establish LEF. Excess PSI-generated reducing equivalents can also be exported to the mitochondria through the malate or triose phosphate transporter and consumed by the mitochondrial respiratory chain (82), resulting in increased ATP production. Interestingly, enhanced respiration was detected in a pgrl1 knockdown mutant of Chlamydomonas reinhardtii with impaired CEF, in which over-reduction of the acceptor side of PSI was prevented (88).
The PQ pool is shared by CEF and LEF together with the chlororespiratory chain in which stromal reducing equivalents are transferred to dioxygen through the PQ pool (16). Several components of this chlororespiratory chain have been identified that catalyze the nonphotochemical reduction of the PQ pool. The first is the NAD(P)H dehydrogenase complex that participates in both CEF and chlororespiration. It consists of a large number of chloroplast- and nucleus-encoded subunits. Surprisingly, none of the identified subunits contains a pyridine nucleotide-binding motif involved in catalysis (106). Besides structural subunits, mutant screens have revealed several nucleus-encoded factors that are required for the synthesis of the structural subunits and act at post-transcriptional steps, in particular in the editing of chloroplast NDH transcripts (66) and in the processing of polycistronic transcripts and in the regulation of their expression (52). While this NAD(P)H dehydrogenase complex is present in plants, it is absent in Chlamydomonas in which PQ reduction appears to be mediated instead by the NDH-2 enzyme (57).
Plastid quinol terminal oxidase (PTOX) is the ultimate component of the chlororespiratory chain (23, 137). Together with the NDH complex, it could play a regulatory role in maintaining the redox poise of the PQ pool in the thylakoid domains where CEF is prevalent, especially under environmental stress conditions. It could also act as an electron safety valve by preventing the over-reduction of the electron transport chain under high light (5). Indeed, PTOX diminishes the PQ reduction state during a dark-to-light transition when the Calvin–Benson cycle is not yet activated (58, 104). However, over expression of PTOX did not confer enhanced protection against photoinhibition in tobacco and Arabidopsis. The variegated immutans mutant of Arabidopsis is deficient in PTOX expression (23). The leaves of this mutant have green and white sectors with plastids lacking lamellar structures and unable to produce chlorophyll and carotenoids (23). The analysis of this mutant revealed that PTOX is important during the early stages of chloroplast biogenesis for carotenoid synthesis (5) and for lowering the excitation pressure of PSII (103), a process that provides a protection against photo-oxidation. In C. reinhardtii, PTOX acts as an electron sink in mutants lacking Cytb 6 f or PSI. Moreover, it was recently shown that PTOX plays an important role in lowering the PSII excitation pressure in the gun4 mutant of C. reinhardtii, in which the electron flow is perturbed (47). Treatment of this mutant with propylgallate, an inhibitor of PTOX, led to a rise in PSII excitation pressure. PTOX also appears to play an important role in marine cyanobacteria under conditions of iron limitation. These organisms adapt by lowering their Cytb 6 f and PSI levels, two complexes with several iron–sulfur proteins that constitute an important sink for iron in the chloroplast (9). To avoid over-reduction of their electron transport chain, they use an alternative electron pathway in which electrons from water are extracted from the electron transport chain at the level of the PQ pool through PTOX to reduce dioxygen (9). In this way, PTOX lowers the PSII excitation pressure and prevents the formation of reactive oxygen species, which could be produced even at low irradiance in these organisms.
Thylakoid Protein Phosphorylation and Dephosphorylation and Redox Control
Stt7/STN7 kinases and state transitions
PSII and PSI are connected to their specific light-harvesting systems, LHCII and LHCI, respectively, each of which has a distinct pigment composition with specific light absorption properties. While PSII absorbs mostly in the blue and red region of the solar spectrum, PSI absorbs in the blue and far-red regions. Hence, changes in the light quality will lead to unequal excitation of PSII and PSI. Because these two photosystems are serially connected through the electron transport chain, this may lead to over-reduction or overoxidation of the PQ pool and thus cause serious perturbation of photosynthetic electron flow. This problem is avoided because plants and algae have evolved a process called state transitions that corrects for any imbalance in the redox state of the PQ pool (101, 135). If PSII is overexcited, the PQ is reduced, and plastoquinol (PQH2) docks to the Qo site of Cytb 6 f (Fig. 2). This event leads to the activation of a protein kinase that phosphorylates LHCII. Upon phosphorylation, the mobile part of LHCII is displaced from PSII to PSI, thus re-equilibrating the cross-sections of the antennae of PSII and PSI and their respective light excitation. This state is called state 2. The process is reversible, as over-excitation of PSI causes the oxidation of the PQ pool, deactivation of the kinase, and dephosphorylation of LHCII by a phosphatase and the return of LHCII to PSI. This state is called state 1.
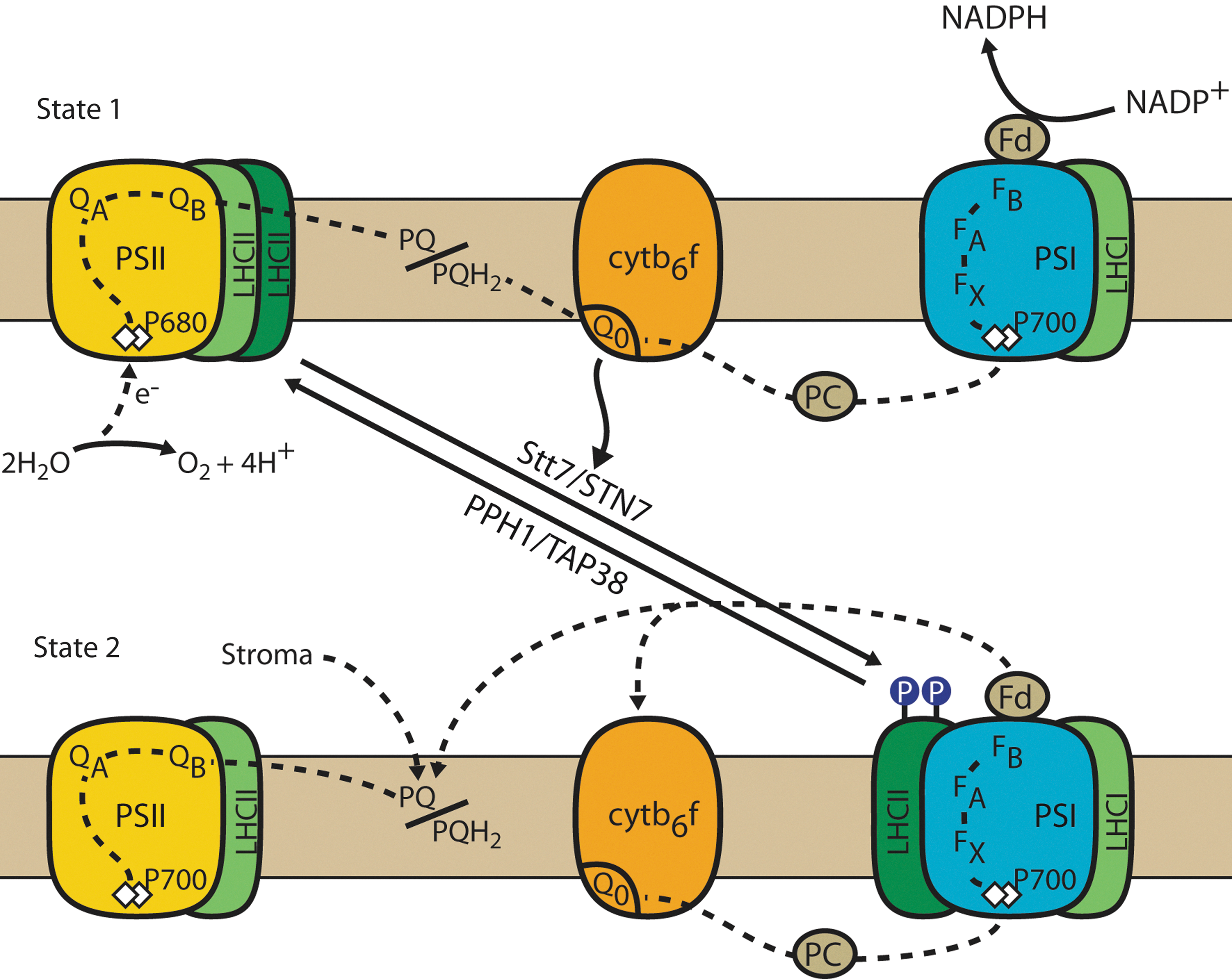
State transitions were discovered independently by Bonaventura and Myers and by Murata in 1969 (18, 79). A few years later, Bennett found that light induces the phosphorylation of LHCII, and that the kinase activity is associated with the thylakoid membranes (14, 15). It was later recognized that the activation of the LHCII kinase was controlled by the redox state of the PQ pool (4, 54). In spite of many efforts, the biochemical hunt for the LHCII kinase was unsuccessful until new molecular genetic approaches were utilized (28, 29, 51, 122). Kohorn and coworkers identified the TAK kinase family in Arabidopsis, a group of three kinases of which at least one appears to play a role in the phosphorylation of the LHCII proteins and in state transitions (120, 121). However, no further studies have been performed on these kinases, and their exact role is still not clear. Another approach involved the search for mutants deficient in state transitions in Chlamydomonas reinhardtii. This green unicellular alga is particularly well suited for such an approach, because the mobile part of LHCII constitutes 80% of the total LHCII, whereas in land plants, only 15%–20% of LHCII is mobile (37). The massive transfer of LHCII from PSII to PSI in C. reinhardtii during state transitions leads to significant changes in room-temperature fluorescence from PSII, which can be used for the screening of mutants affected in state transitions. In this way, several mutants were isolated and characterized (46, 69). One of the most valuable mutants was stt7 deficient in the thylakoid Ser/Thr protein kinase Stt7 (38). This kinase was shown to be required for the phosphorylation of LHCII. It displays a high sequence similarity with another kinase called Stl1. Both Stt7 and Stl1 are conserved in other algae and land plants, and they have been named STN7 and STN8, respectively (12, 38).
As often occurs in screens of this sort, some of the identified mutants are indirectly affected in state transitions. As an example, mutants deficient in the phylloquinone biosynthetic pathway appeared in this screen, because the loss of phylloquinone perturbs PSI function and indirectly affects state transitions (70). Other interesting mutations that had an indirect effect on state transitions identified proteins outside of the chloroplast involved in mitochondrial light acclimation (109) and in the light-regulated expression of the LHCII proteins (80).
During a state 1-to-state 2 transition, the chlorophyll a/b-binding protein CP29 dissociates from the PSII-LHCII complex and associates with the PSI core in the vicinity of the PsaH/L/O/P domain as seen by electron microscopy in C. reinhardtii (61). It appears that redox-dependent multiple reversible phosphorylations of CP29 induce conformational changes that determine the affinity of LHCII to either PSII or PSI. Further analysis of large-molecular-weight complexes from solubilized thylakoid membranes from C. reinhardtii cells in state 2 revealed the presence of a PSI-LHCI supercomplex containing the minor monomeric LHCII proteins CP29, CP26, and Lhcbm5 (126).
Transition from state 1 to state 2 can be induced by red-light illumination, which preferentially excites PSII relative to PSI. However, the same transition can also be induced in C. reinhardtii in the dark through ATP depletion induced by treating the cells with an uncoupler such as carbonyl cyanide 4-(trifluoromethoxy)phenylhydrazone (FCCP), by anaerobia or by using mitochondrial mutants deficient in respiration (20). In these mutants, ATP depletion is thought to stimulate glycolysis and the accumulation of NADH, which will reduce the PQ pool and trigger a transition to state 2 (96). Under those conditions, a switch from LEF to CEF occurs in Chlamydomonas (44, 45). Thus, the major role of state transitions in this alga is to maintain the redox poise of the PQ pool and to respond to metabolic needs rather than to adjust the relative antenna sizes of PSII and PSI. This may explain why the mobile LHCII antenna is markedly larger in Chlamydomonas than in land plants. In plants, LHCII phosphorylation also responds to the redox state of the PQ pool, which in turn is modulated by the metabolic state and the environmental conditions (42, 53). Hence, in this respect, the difference between state transitions in algae and plants may only be quantitative.
In mutants of C. reinhardtii deficient in mitochondrial respiration, ATP deficiency is partly compensated by an increase in CEF. Although this shows that photosynthetic electron flow and, in particular, the redox poise of the PQ pool are influenced by mitochondrial respiration, it is surprising that the growth phenotype of the stt7 mutant is nearly undistinguishable from that of wild type (46), suggesting some sort of energetic compensation between mitochondrial respiration and state transitions when either of the two systems is perturbed. This is indeed the case as shown by the fact that the phototrophic growth of double mutants affected in both mitochondrial respiration and state transitions is severely impaired compared to each single mutant (22). Thus, state transitions play an essential role in C. reinhardtii for maintaining the cellular ATP level by enhancing CEF when mitochondrial respiration is compromised.
The Stt7/STN7 kinase is associated with the Cytb 6 f complex (72). Although it has not yet been demonstrated that this kinase directly phosphorylates LHCII, this is likely to be the case because this kinase can be coimmunoprecipitated with LHCII, and phosphorylation of several LHCII sites under state 2 conditions requires Stt7 (72). Interestingly, the kinase is phosphorylated under state 2 conditions. This probably represents an autophosphorylation, because in vitro recombinant Stt7 kinase autophosphorylates readily (G. Fucile and J.D. Rochaix, unpublished results). Removal of the phosphorylation sites of Stt7/STN7 both in Chlamydomonas and Arabidopsis did not affect state transitions, indicating that phosphorylation of the kinase is not required for this process (72, 134). However, the marked decrease in the level of STN7, which is observed after prolonged state 1 conditions in wild-type plants, did no longer occur when the four phosphorylation sites were changed to Asp, but not when they were changed to Ala. The specific interfering effect of phospho-mimetic amino acids strongly suggests that the phosphorylation status of STN7 affects its turnover in Arabidopsis (134).
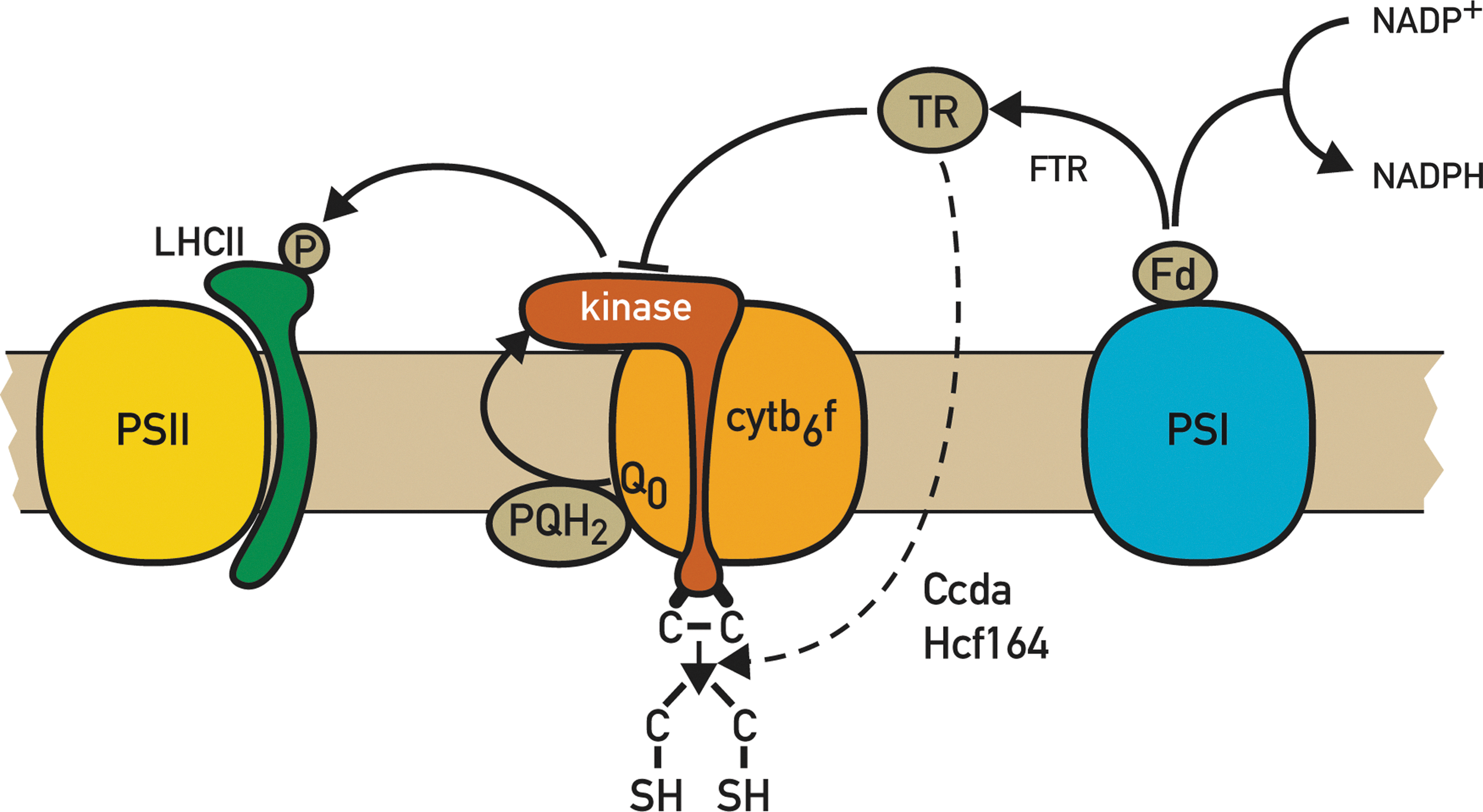
An intriguing feature is the presence of two conserved Cys residues within the N-terminal domain on the lumen side of Stt7/STN7 (Fig. 3). These Cys residues appear to be essential because changes of Cys to either Ala or Ser through site-directed mutagenesis abolished the kinase activity and state transitions both in Chlamydomonas (72) and Arabidopsis (A. Shapiguzov and J.D. Rochaix, unpublished results). It is tempting to propose a role for these Cys residues in the inactivation of the LHCII kinase, which occurs under high light, and which is mediated by FD and TRX, at least in land plants (99). The fact that the latter proteins are on the opposite side of the thylakoid membrane with respect to these Cys residues is not necessarily incompatible with this proposal, because thiol-reducing equivalents can be transferred across the thylakoid membrane through CCDA and HCF164 (Fig. 3) (73, 86). These two proteins are required for heme attachment on the lumen side and for Cytb 6 f biosynthesis and may also regulate the redox state of the two luminal Cys of Stt7/STN7. However, recent experiments indicate that these Cys residues form a disulfide bridge in Chlamydomonas, which appears to be essential for kinase activity, and the redox state of this disulfide bridge does not change during state transitions. Moreover, contrary to the expectation, it is not reduced by high-light treatment in Chlamydomonas (A. Shapiguzov and J.D. Rochaix, unpublished results). Whether this process is different in land plants remains to be explored. In this respect, a TRX-like CxxxC motif in the stromally exposed kinase domain of STN7 has been proposed as a potential target for TRX (92). However, this motif is unlikely to play an essential role in the regulation of the kinase, because site-directed mutagenesis of its Cys residues did not affect LHCII phosphorylation and state transitions in Arabidopsis (A. Shapiguzov and J.D. Rochaix, unpublished results). Moreover, while this motif is conserved in land plants and several algae, it is absent in C. reinhardtii (92). An important task will be to determine whether the redox control of Stt7/STN7 differs between Chlamydomonas and land plants and, in particular, whether it involves different regions of the kinase. It is unlikely that proteolysis of Stt7, which occurs under extended state 1 conditions, is involved in the control of the activity of the kinase, because the decay of the kinase is slower than that of its activity under high light, and because in C. reinhardtii, phosphorylation of LHCII and state transitions occur readily even after prolonged state 1 conditions when the amount of kinase is strongly decreased (72, 112, 134). However, there may be differences with land plants, and it cannot be excluded that in this case, regulation of the STN7 kinase activity also occurs partly through changes in its accumulation.
The Stt7/STN7 kinase appears to have a broader role than state transitions and is also required for adaptation to light fluctuations. Reversible LHCII protein phosphorylation can be induced by subjecting Arabidopsis plants to alternative periods of low light and high light (127). Under these conditions, LHCII is phosphorylated during the low-light and dephosphorylated during the high-light periods. However, these changes in phosphorylation do not significantly change the distribution of light excitation energy between the two photosystems. In the absence of STN7, the excitation pressure of PSII rises during the low light phases, suggesting that this kinase ensures a balanced distribution of excitation energy to both photosystems and thereby optimizes electron transfer under fluctuating light. The loss of STN7 in plants subjected to this fluctuating light regime leads to a severe decrease in growth (127).
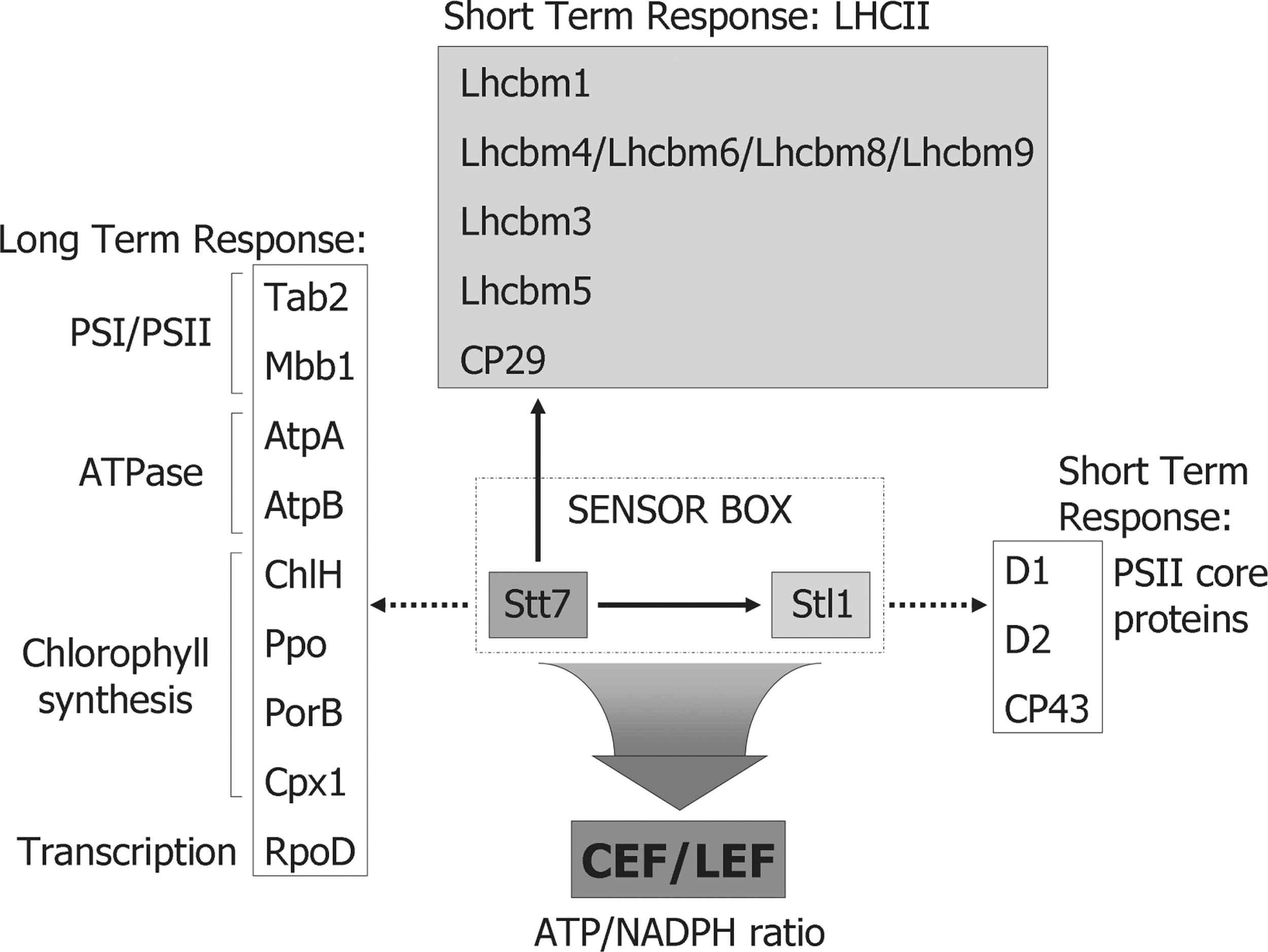
The STN7 kinase is involved both in a short-term response such as state transitions and in an LTR that comprises changes in the nuclear and chloroplast gene expression. These changes induce compensatory alterations in the thylakoid protein composition that ultimately lead to balancing of the excitation energy between PSII and PSI and to the maintenance of the redox poise of the PQ pool (19, 87). In the absence of STN7, the chloroplast psaAB operon is upregulated at the transcriptional level, and the amount of Lhca3, a nucleus-encoded PSI antenna protein, is upregulated post-transcriptionally. It was shown that the short-term response and LTR diverge at or immediately downstream of STN7, and that the latter does not involve LHCII phosphorylation or any other step of state transitions (87). This implies that Stt7/STN7 must have other targets that act in the LTR signaling cascade, but that are still largely unknown. In this respect, a comparative analysis of the Stt7 known target sites in the different LHCII proteins of C. reinhardtii led to the identification of a consensus motif for these sites in which the phosphorylated Ser/Thr is flanked on each side by two basic residues (Fig. 4) (71). Interestingly, a similar motif was also proposed as specific signature for the STN7 phosphorylation sites in Arabidopsis (133). Additional potential Stt7/STN7 phosphorylation sites were identified by searching the predicted chloroplast proteins from Chlamydomonas according to their sequence similarity with the predicted plastid proteins from Arabidopsis for the presence of the (K/R/H)(K/R/H)TX(K/R/H)(K/R/H) motif. Of particular interest is the fact that among these new potential substrates of Stt7, several are low-abundance chloroplast- and nucleus-encoded proteins involved in the post-transcriptional steps of chloroplast gene expression (Fig. 5). They include Tab2 required for psaB mRNA translation and the chloroplast σ-factor RpoD. These findings raise the possibility that Stt7 may have additional roles in the regulation of the synthesis of chloroplast proteins involved in photosynthesis. Other potential Stt7 target sites were also found among proteins involved in chlorophyll biosynthesis (ChlH, Cpx1, PorB, and Ppo) and the two ATP synthase subunits AtpA and AtpB. It will be important to test whether these newly predicted sites are indeed phosphorylated in vivo both in Chlamydomonas and Arabidopsis.

Under standard laboratory conditions, the stn7 mutant does not display any growth phenotype as compared to the wild type. However, when this mutant was subjected to fluctuating light, its growth was impaired (12), and it had a severely stunted phenotype (127). Moreover, the fitness of this mutant tested under natural field conditions and measured by seed production was significantly decreased relative to the wild type (49). The fitness of the stn7-npq4 double mutant deficient both in state transitions and NPQ was even more affected. It is however not clear whether it is the loss of state transitions or the loss of the kinase with the LTR, which is the cause of these phenotypes.
Further insights into the relationship between state transitions and LTR came from the analysis of several Arabidopsis mutants. This analysis revealed that state transitions are not required for LTR as shown by the existence of mutants such psaI-1 lacking the PSI subunit PsaL and as a consequence deficient in the docking site of PSI for LHCII (75). This mutant is strongly affected in state transitions, but still has an LTR (87). Moreover, the psaD1 and psaE1 mutants lacking one of the isoforms of the PsaD and PsaE PSI proteins, respectively, which still accumulate PSI, are impaired in the photosynthetic electron flow and unable to perform LTR and state transitions, but their LHCII is phosphorylated under all light conditions because of the over-reduction of the PQ pool (87). These results and the observation that the growth defect of the stn7 psaD1 and stn7 psaD2 double mutants was exacerbated suggest that the STN7 kinase does not work properly when it is permanently activated and that its activity needs to be modulated for an efficient LTR to occur.
Stl1/STN8 kinase and PSII repair
The STN8 kinase is mainly required for the phosphorylation of the PSII core proteins D1, D2, and to some extent CP43 (17, 133). It is not yet entirely clear how the activity of STN8 is regulated. Analysis of the phosphorylation of the PSII core proteins of Arabidopsis upon transfer from the dark to low light, corresponding to a shift from an oxidized to a more reduced state of the PQ pool, revealed an increase in phosphorylation of the D1 and D2 proteins, but not of CP43(12, 17, 133, 138). High light irradiance caused a further increase in D1 and D2 phosphorylation, which correlated with an increase of STN8 (138). However, in another study, no significant increase of D1 and D2 phosphorylation was observed in high light as compared to low light (17). This discrepancy may be due to slight differences in the light conditions used in these two studies.
Natural variation of the activity of STN8 was observed by comparing three Arabidopsis thaliana accessions. Phosphorylation of the D1 and D2 proteins decreased by 50% in the accession Wassilewskija-4 (Ws-4) as compared to Columbia-0 and Landsberg erecta (138). This difference could be correlated with a 50% decrease in the level of STN8 in Ws-4 relative to the two other accessions. These results suggest that the amount of the STN8 kinase is limiting, and that its activity may be regulated by changes in its accumulation through changes in its rate of synthesis and/or proteolysis.
Analysis of the protein phosphorylation profiles of the stn7 and stn8 mutants of Arabidopsis deficient in STN7 and STN8, respectively, indicates residual phosphorylation of the LHCII and PSII core proteins. However, these phosphorylations are undetectable in the stn7stn8 double mutant, indicating some substrate overlap between the STN7 and STN8 kinases (17, 50, 133). Only a small amount of phosphorylation of D2,PsbH and LHCII could be detected in the double mutant that must be due to a still-unidentified chloroplast kinase. Both kinases appear to act synergistically as shown by the fact that in field tests, the fitness of stn8 alone is not significantly reduced as compared to the wild type. However, the stn7 stn8 double mutant is considerably more affected than stn7 (49).
Analysis of the stn8 mutant revealed that the folding of the thylakoid membranes is altered in the absence of STN8 (50). Interestingly the granum size, which is usually conserved even among different plant species, is significantly increased in stn8,and there are less granum stacks on average. This change in membrane folding can be correlated with changes in the PSII repair cycle. Because PSII generates highly oxidizing species during the water-splitting reaction, photodamage is unavoidable, particularly at the level of the D1 reaction center protein. During the PSII repair cycle, damaged PSII moves from the grana to the stroma lamellae, and D1 is degraded, mainly by the FtsH and Deg proteases and replaced by newly synthesized D1. In the absence of STN8, the movement of PSII from the grana to the stromal region is slowed down, indicating that PSII core protein phosphorylation is involved in this process. Moreover, the access of FtsH to the granum region is hindered in stn8 (50).
Besides the PSII core proteins, STN8 has additional targets, including CAS, the chloroplast calcium-sensing protein (132), and PGRL1 (97), a protein involved in CEF, which is a part of the PSI-LHCI-LHCII supercomplex (31). The loss of STN8 could be correlated with a reduced CEF stability, raising the possibility that this phosphorylation may be important for fine-tuning of the photosynthetic machinery to meet the NADPH/ATP demands of the plastid metabolism.
PPH1/TAP38 and PBPC phosphatases
Among all the state transition mutants of C. reinhardtii found, none was affected in a phosphatase in contrast to the kinase mutants where three independent alleles of Stt7 were identified. This may be due to some redundancy between different phosphatases or to duplications of the genes of these enzymes in Chlamydomonas. Therefore, a reverse genetics approach was chosen using Arabidopsis (91, 117). A total of 159 phosphatase genes were identified in its genome. Of those predicted to be targeted to the chloroplast, 84 homozygous TDNA insertion lines with disruptions of the genes encoding 60 putative phosphatases were screened for the inability to dephosphorylate LHCII during a state 2-to-state 1 transition (91, 117). This screen revealed PPH1/TAP38, a protein phosphatase 2C (PP2C) that is required for the dephosphorylation of LHCII proteins. In the absence of this phosphatase, the dephosphorylation of LHCII is strongly impaired. PPH1/TAP38 appears to act in a specific way, as dephosphorylation of the PSII core proteins is not affected in its absence. Consistent with its function, this phosphatase localizes to the stromal thylakoid lamellae, where at least a part of phosphorylated LHCII is associated with PSI (117). A similar screen identified PBPC (PSII core protein phosphatase), another PP2C that is involved in the dephosphorylation of the PSII core proteins CP43, D1, D2, and PsbH, the known substrates of the STN8 kinase (107). Interestingly, thylakoid folding is affected in the absence of this phosphatase, and its overexpression alters the kinetics of state transitions with a faster transition from state 2 to state 1 and a slower transition from state 1 to state 2. Phosphorylation/dephosphorylation of the PSII core proteins may influence the diffusion of macromolecular complexes in the thylakoid membrane. Earlier findings showed that the PSII repair cycle requires a cycle of phosphorylation and dephosphorylation reactions in the thylakoid membrane (98) and were further confirmed by the analysis of the stn8 mutant, in which D1 repair is slowed down (50). This membrane is one of the most crowded biological membranes with a very high protein content that poses a challenge for the diffusion of macromolecular complexes (64). PPH1/TAP38 and PBPC, which counteract the activity of STN7 and STN8, respectively, could have an important role in this process.
In vitro phosphatase assays revealed that both phosphatases require Mn2+rather than Mg2+ for optimal activity. They are conserved in land plants and also have orthologs in algae, in particular in C. reinhardtii. It is not yet known whether these phosphatases are constitutively active or whether they are under redox control as their partner kinases. It is intriguing that these phosphatases contain conserved Cys, which might be involved in such a control. Although these two phosphatases display different substrate specificities and physiological functions, their activities might overlap to some extent (107). It is however clear that these two phosphatases are not sufficient to account for all the observed changes in phosphorylation within the thylakoid membrane and that additional phosphatases are involved.
The picture that emerges from these studies is that of an intricate signaling network in the chloroplast in which the redox state of the PQ pool plays a central role (Fig. 6). It is modulated by the spectral quality and quantity of light, the ATP/ADP balance, and the CO2 level. The Stt7/STN7 kinase appears to play a central role in this network, as it is able to sense the redox state of the PQ pool through its close association with Cytb 6 f and most likely also that of the FD/TRX system, and it acts as a trigger for both the short-term response and LTR. It controls the phosphorylation of the LHCII proteins together with its corresponding PPH1/TAP38 phosphatase, and similarly, the Stl1/STN8 kinase regulates the phosphorylation of the PSII core proteins with the PSII core protein phosphatase (PBCP). How these two kinases are connected with the chloroplast sensor kinase (CSK) and plastid transcription kinase (PTK) remains to be established, although there are some hints that Stt7/STN7 may regulate the activities of CSK and PTK directly or indirectly (see below).

Chloroplast Gene Expression and Redox Control
It is well established that the chloroplasts contain their own genetic system comprising a relatively small genome coding for ca. 100 genes and a protein synthesizing system. Chloroplasts are thought to be descendants of cyanobacteria-like organisms that invaded an ancestral cell through endosymbiosis two billion years ago. During evolution, a large part of the genome of the invading cyanobacteria was transferred to the nucleus. The question why this process did not reach completion with a complete loss of the chloroplast genome is not clear, although various hypotheses have been proposed. One is that the chloroplast genome encodes some very hydrophobic proteins with multiple transmembrane domains, 11 in the case of the PSI reaction-center proteins, which may be difficult to translocate across the chloroplast envelope. However, there are examples for the transport of proteins with several transmembrane domains across the biological membranes. Another possible explanation is the redox control proposed by John Allen (2, 89). Chloroplasts need to respond swiftly to changes in the environmental conditions. Therefore, having the chloroplast genes in the organelle would allow for a rapid response to redox changes of the photosynthetic electron transport chain induced by changes in environmental cues such as light quality and quantity, temperature, and nutrient availability.
Light has profound effects on plastid gene expression. Transfer of plants or algae from the dark to the light induces a massive increase in chloroplast gene expression both at the transcriptional and post-transcriptional level. Genetic screens in maize and Arabidopsis, and also in Chlamydomonas, have revealed a large number of nucleus-encoded factors that are specifically required for chloroplast post-transcriptional steps such as RNA processing, splicing, RNA stabilization, and translation (10, 41). In several cases, these factors act in a gene-specific manner, especially in Chlamydomonas (Fig. 7).
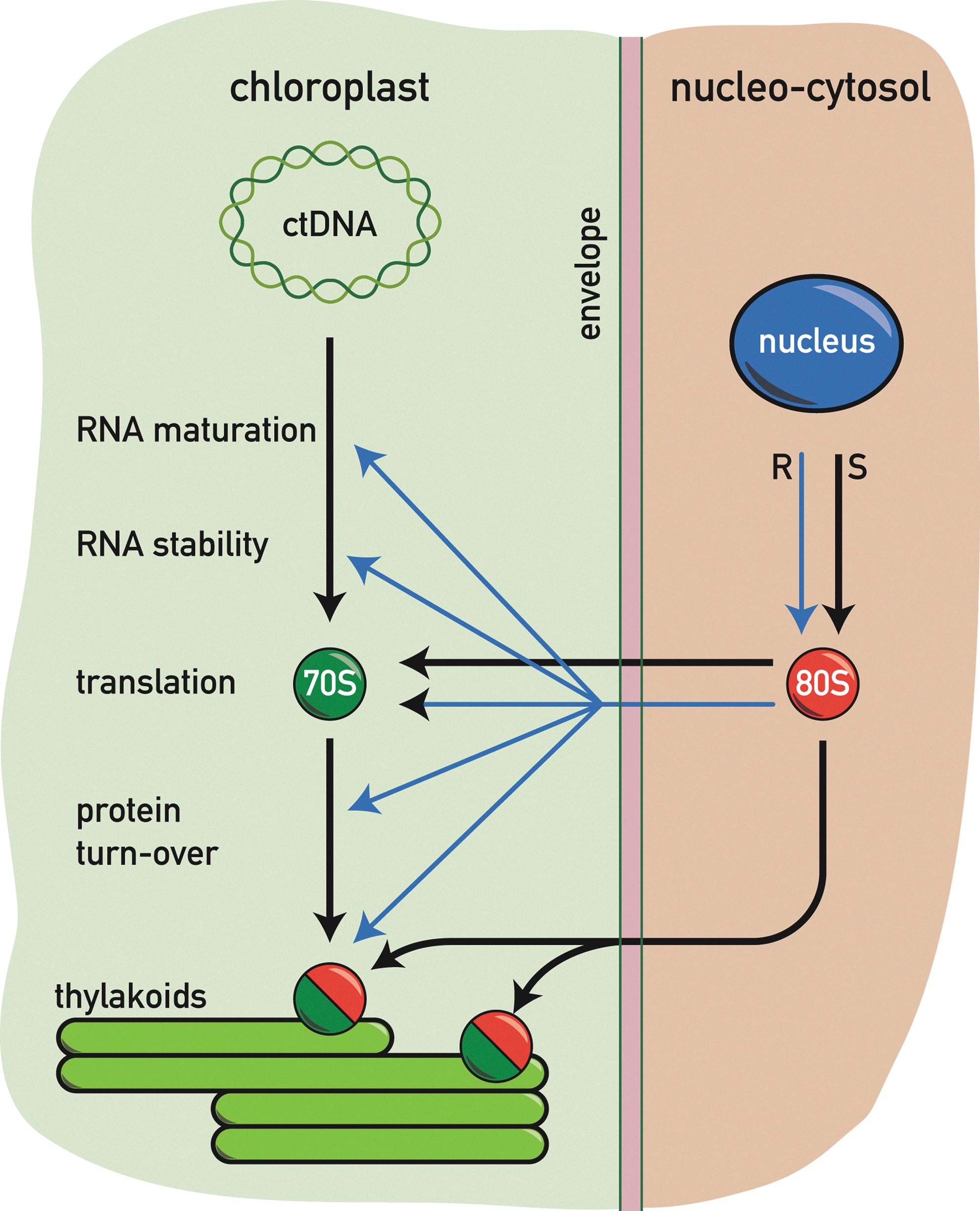
Redox regulation of transcription
The redox state of the PQ pool can be oxidized or reduced through unbalanced excitation of PSI and PSII, thereby triggering a signal that leads to state transitions and/or to the LTR, which results in changes in gene expression both in the chloroplast and nuclear compartments. These changes tend to re-equilibrate the excitation for an optimal photosynthetic performance through readjustment of the stoichiometry of the photosynthetic complexes. An important target of photosynthetic redox signals is the plastid-encoded RNA polymerase (PEP), which resembles the bacterial enzyme and consists of the α−, β,β′-, and β′′-subunits encoded by the plastid genes rpoA, rpoB, rpoC1, and rpoC2, respectively (55) and is associated with several σ factors. In contrast, the activity of the nucleus-encoded single-subunit plastid polymerase does not appear to be significantly affected by redox signals (124).
Plastid Ser/Thr protein kinase
A convenient system to study LTR during light acclimation is to shift plants from light preferentially absorbed by PSII (PSII light) to light preferentially absorbed by PSI (PSI light) or vice-versa (PSII–PSI and PSI–PSII light). This system was used to study the chloroplast protein fraction associated with PEP together with the DNA-binding proteins in terms of promoter recognition and specificity and the protein phosphorylation state (43, 124). This fraction was shown to contain a kinase activity called PTK. PTK is a Ser/Thr protein kinase of the casein kinase II family (8) and is mainly associated with the PEP polymerase. In vitro, it is less active in a phosphorylated state and activated by reduced glutathione (8). PTK acts as a global regulator of chloroplast transcription, which is modulated by its phosphorylation state and that of the σ factors (74, 83). It was initially proposed that phosphorylation of σ factors enhances promoter binding and results in transcription arrest in the etioplasts, but this phosphorylation does not occur in the chloroplasts (128). Thus, phosphorylation and dephosphorylation of the σ factors confer etioplast and chloroplast type-specific promoter usage, respectively. However, both positive and negative effects on transcription have been observed depending which σ factor is phosphorylated and which chloroplast promoter is considered (116). Moreover, some σ-factors contain multiple phosphorylation sites, some of which appear to be the substrates of PTK, which itself responds to phosphorylation and redox regulation by glutathione (8, 83).
The redox regulation through glutathione does not appear to play a significant role in vivo during PSII–PSI and PSI–PSII light shifts because the redox state of glutathione remains stable during these light changes (43). Instead, the chloroplast kinase activity during PSI–PSII light shifts was lower than in PSII–PSI light shifts. It was therefore suggested that STN7, which is activated under PSII light, directly or indirectly mediates the phosphorylation of PTK and thereby its inactivation under PSII light (124). The targets of PTK include σ factors. Among these, SIG1 has been studied in some detail. Redox-mediated regulation of transcription occurs through the phosphorylation status of SIG1 (119). Reduced PQ leads to dephosphorylation of SIG1, which in turn enhances transcription of the psaAB genes encoding the reaction center proteins of PSI. In contrast, phosphorylation of SIG1 is triggered by oxidized PQ and leads to a decrease of psaAB transcription. The expression of psbA encoding the reaction center protein of PSII is not affected significantly by the redox state of the PQ pool. Other targets of PTK include small DNA-binding proteins whose phosphorylation state changes during PSI–PSII light acclimation. These phosphorylation events appear to be essential for mediating the redox signals from the photosynthetic electron transport chain toward transcription of the psaA and psaB genes and require in addition the synergistic action of the thiol redox signals (124). The significance of these changes in the thiol redox state is not clear. However, one interesting possibility is that they could induce conformational changes, thereby granting access of the masked sites to phosphorylation (115).
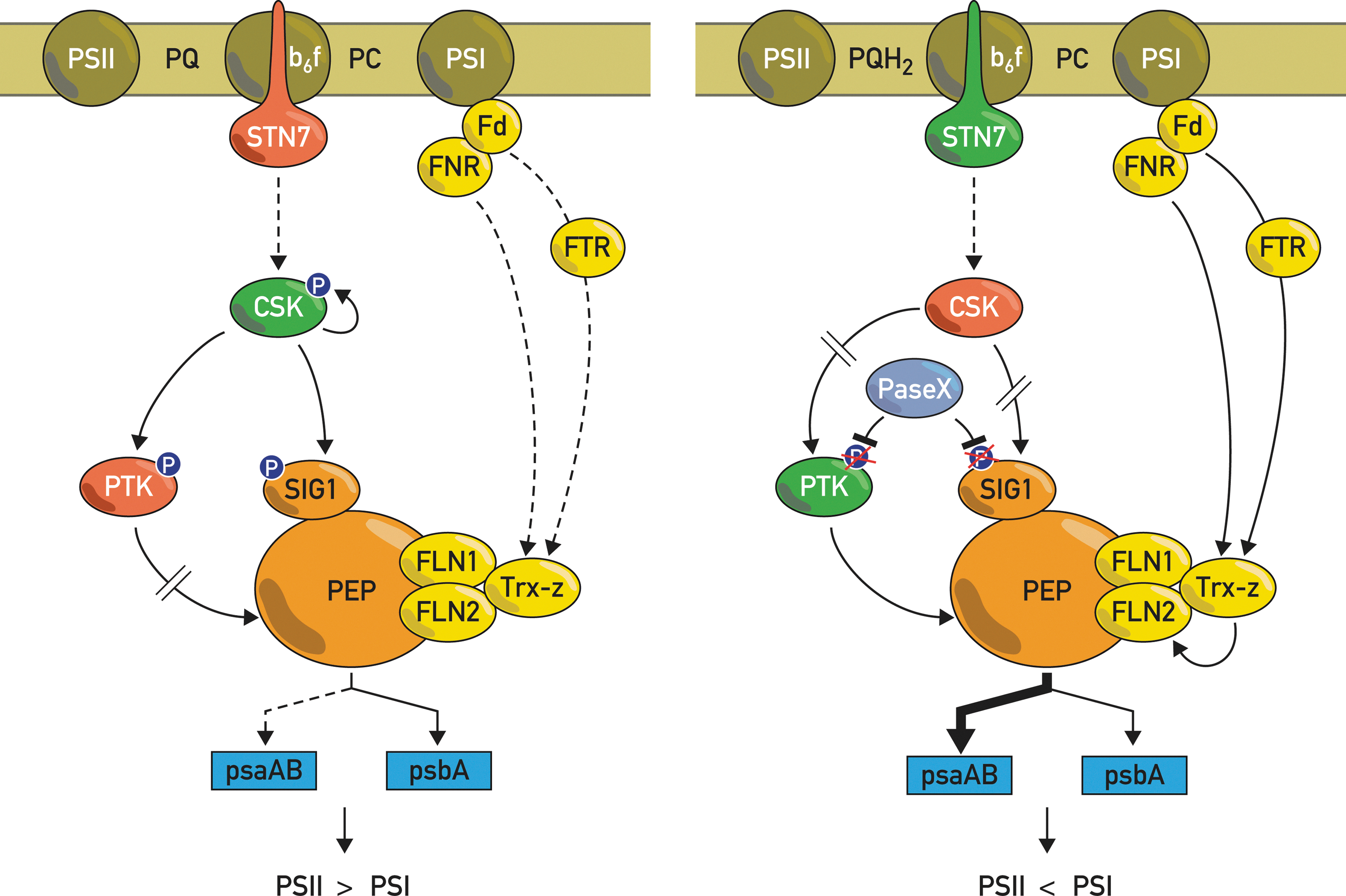
CSK kinase
The chloroplast CSK kinase was proposed to act as a sensor kinase based on its sequence similarity to cyanobacterial sensor histidine kinases (95). It is conserved in all major plant and algal lineages, except for C. reinhardtii. However, while CSK is present as a typical sensor histidine kinase in cyanobacteria, red algae, diatoms, and phaeophytes, it has been modified in green algae and plants in which the conserved His phosphorylation site has been lost (95). The chloroplast transcript levels are altered in CSK knockout lines of Arabidopsis, indicating that this kinase is involved in chloroplast gene expression either directly or indirectly (95). Interestingly, CSK autophosphorylation correlates with SIG1 phosphorylation when the PQ pool is oxidized, and these phosphorylations also correlate with a decrease in psaAB gene expression (93). Moreover, CSK interacts with PTK and SIG1 based on yeast two-hybrid assays. By taking into account the results discussed above, the model proposed by Puthiyaveetil et al. (94) can be extended as follows: It is assumed that the activity of CSK is regulated by the redox state of the PQ pool through the STN7 kinase. When the PQ pool is oxidized, CSK phosphorylates and inactivates PTK. CSK would then phosphorylate SIG1, which leads to the repression of the psaAB genes, whereas psbA expression is not markedly affected (Fig. 6). When the PQ pool is reduced, SIG1 is dephosphorylated, and repression of the psaAB genes is relieved. This might be catalyzed by an unidentified phosphatase (PaseX), which would also dephosphorylate PTK in addition to SIG1. Interestingly, a phylogenetic analysis revealed that photosynthetic organisms thar possess CSK fall into two groups. The first lacks PTK, but instead contains Ycf29, a response regulator of cyanobacterial origin that has been proposed to be the cognate partner of CSK (108). The second group contains PTK, but lacks Ycf29. This suggests that during evolution, the prokaryotic Ycf29 response regulator was replaced by PTK, an analogous eukaryotic-type protein kinase as the functional partner in a two-component regulatory system (94).
With the available tools, it should be possible to test the model in Figure 6. The question remains how CSK activation is linked to the redox state of the PQ pool, and how it relates to the STN7 kinase. The fact that the stn7 mutants are unable to perform a proper LTR suggests that these kinases operate in the same pathway. It should be noted however that the activation of these two kinases is regulated in opposite ways by the redox state of the PQ pool. Reduction of this pool activates STN7 and deactivates CSK, whereas the opposite occurs when the pool is oxidized. Current evidence favors a complex network in which both phosphorylation- and thiol-dependent signals are involved and may act synergistically (110). Recent studies have revealed a new player in this network, Trx-z, a Trx-like protein (6, 110). The Arabidopsis knockout mutant of Trx-z displays an albino phenotype with a strong impairment of chloroplast development. This feature is rather unusual, as Trx mutants can usually be rescued because of the functional redundancy of the Trx system. Expression of genes transcribed by PEP was specifically decreased in the absence of Trx-z, indicating that this protein plays an important role in chloroplast gene expression (6). Based on yeast two-hybrid assays, Trx-z interacts with two fructokinase-like proteins, FLN1 and FLN2, which are members of the pfbB-sugar kinase family. However, these two proteins have amino acid substitutions within the predicted active site and have no detectable sugar-phosphorylating activity, indicating that these proteins have acquired a new, most probably, regulatory function. This view was further strengthened by the observation that FLN2 undergoes redox changes during light–dark transitions, which are mediated by Trx-z (6). Analysis of purified PEP by mass spectrometry showed that Trx-z and FLN2 are intrinsic subunits of this complex (110). Although it is not clear how Trx-z relates to the other components of the regulatory network, it is likely that Trx-z and the two FLNs belong to a signaling chain, which conveys redox regulation of PEP function and could couple transcriptional control to light signals through the photosynthetic electron transport chain (Fig. 8). The picture that emerges from these studies is that of a complex network of interacting proteins that transmits signals from the thylakoid membrane to the plastid gene expression system through multiple phosphorylations and protein redox changes.
Redox control of translation
During the assembly of the photosynthetic complexes in the thylakoid membrane, coordination of the synthesis and stoichiometric accumulation of their subunits is mediated by two different processes. First, subunits made in excess are usually rapidly degraded by chloroplast proteases. Second, assembly and translation are coupled through the control by the epistasy of synthesis (CES) process. CES was first uncovered during the analysis of mutants of C. reinhardtii deficient in Cytb 6 f (24). In the absence of the Cytb 6 subunit, cytochrome f cannot assemble, and in its free unassembled form, it represses its own translation probably indirectly through another factor. A similar negative feedback inhibition has been observed for unassembled subunits of other photosynthetic complexes. In the case of PSII, unassembled D1 represses its own translation in the absence of D2, and in the absence of D1, unassembled CP47 represses its translation (77). A similar scenario occurs with PSI where in the absence of PsaB, PsaA inhibits its own translation, and in the absence of PsaA and PsaB, unassembled PsaC exerts a negative feedback effect on its own translation (136). Finally, in the case of the ATP synthase, in the absence of the β-subunit, the unassembled α-subunit represses its own translation (40).
Translation represents not only an important regulatory step for the coordinate assembly of the photosynthetic complexes, but it is also greatly influenced by changes in environmental cues such as light. Considerable differences in the rate of synthesis of the major thylakoid proteins occur between light- and dark-grown plants and algae (76). This is particularly true for the D1 reaction center protein of PSII. Because of the highly oxidizing water-splitting reaction in PSII, photodamage of D1 is unavoidable, and the protein needs to be resynthesized continuously. It is therefore not surprising that the translation of this protein is strongly stimulated by light. A protein complex consisting of RB60, RB55, RB47, and RB38 was identified in C. reinhardtii, which binds specifically to the 5′-untranslated region (5′UTR) of psbA mRNA (33). Binding of the complex to the psbA RNA is strongly increased by light and mediated through RB47, which belongs to the polyA-binding protein family (140). Mutants lacking this protein are unable to synthesize D1 (139, 140). The RNA-binding activity of the psbA complex correlates with psbA mRNA translation and its association with polyribosomes, which are considerably higher in light-versus-dark-grown cells. Because the proteins of the complex accumulate to the same extent in the dark and the light, this points to a post-translational regulation. It was proposed that this control occurs through FD/TRX based on the observation that the RNA-binding activity is regulated by the redox state of the complex, which in turn is regulated by TRX, at least in vitro (35). RB60 is a chloroplast protein disulfide isomerase (63). It was identified as the redox sensor of the psbA RNA-binding complex. This protein carries a redox-active regulatory site consisting of a vicinal dithiol. Together with TBA1, an oxidoreductase (123), RB60, regulates the RNA-binding activity of RB47 in response to the chloroplast redox state by reacting with a regulatory disulfide of RB47, indicating that these two redox partners are probably coupled (1). In its reduced, but not in its oxidized, state, RB47 binds to the psbA 5′UTR, which leads to an increase of translation initiation.
An important issue in redox control is to achieve a proper redox poise, which is only possible for a given redox factor if counteracting oxidizing and reducing activities are available. This is indeed the case for RB60. Upon transfer to the light, a priming event occurs in which the vicinal dithiol is oxidized and reduction of the disulfide bridge is likely to be mediated by the FD-TRX system (129, 130). This system is known to regulate the activity of a large set of chloroplast redox-responsive proteins by using reducing equivalents generated by the photosynthetic electron transport chain. Indeed, at higher light intensities, the amount of reduced RB60 parallels the rate of psbA mRNA translation (130). Although the priming event is unknown, it is probably mediated through the redox state of the PQ pool. Whether the Stt7/STN7 kinase is involved in this pathway remains to be explored, especially in view of the fact that, as mentioned above, this kinase could act on factors involved in chloroplast post-transcriptional steps. The finding that both RB47 and RB60 are tightly associated with the thylakoid membrane is compatible with the idea that photosynthetic electron transport is responsible for the oxidizing equivalents for RB60 (131). Thus, two signaling pathways have been proposed for the light-regulated translation of RB60. The first is initiated by the reduction of the PQ pool and the second by PSI, the primary reducer of the FD-TRX system. While the light-induced FD-mediated signal can be replaced by dithiothreitol in the dark, the priming signal cannot be generated by protein thiol reduction in the dark (129). Moreover, RB60 also appears to be involved in the attenuation of psbA mRNA translation in the dark through phosphorylation of RB60 by ADP (34).
The molecular identity of RB55, another component of the psbA 5′UTR complex, is still unknown, whereas RB38 is insensitive to the redox state and binds specifically to U-rich sequences of the psbA 5′UTR (11). It should be noted that RB38 is identical to RB40, which is involved in psbD rather than in psbA translation in C. reinhardtii (114). As for psbA, psbD translation is also enhanced by light. RB40 binds to a polyU-rich region of the psbD 5′UTR, thereby destabilizing a stem–loop structure that allows the small ribosomal subunit to bind to the psbD initiation codon (65, 85). D2 synthesis in the light depends on the formation of a dynamic high-MW complex of 700 kDa in which NAC2 and RB40 form an interprotein disulfide bridge (114). In the dark, this bridge is opened probably through the NADPH-dependent thioredoxin reductase, and the dissociation of RB40 from the complex correlates with the arrest of translation initiation (113). This would support a direct link between chloroplast gene expression and carbon metabolism during cell adaptation in the dark.
Redox regulation is widely used by plants to dynamically adjust chloroplast metabolism. While most reducing equivalents generated by photosynthesis are used for producing NADPH, a small portion is transferred from FD to TRX via FD-dependent TRX reductase and then used to reduce disulfide bonds of chloroplast target proteins (111). In contrast to this well-studied reductive pathway, the oxidizing pathways are still largely unknown as discussed for RB60. However, new insights have come from studies of the oxidation of a thylakoid-associated atypical TRX-type protein, ACHT1 of Arabidopsis (32). When plants are illuminated with moderate light, this protein is first reduced, but subsequently oxidized by 2-Cys peroxiredoxin, which is in turn oxidized by peroxides produced by the photosynthetic reactions, especially under changing light conditions. Whether a similar system also operates for the oxidation of RB60 remains an open question.
Expression of both subunits of Rubisco is known to be tightly regulated, as a decrease in RBCS through RNA silencing (102) or through deletion of the RBCS genes (62) leads to the specific loss of RbcL. Also, when expression of RbcL is blocked through a nonsense mutation, RBCS is rapidly degraded (62). In the case of tobacco, a CES process underlies the coordinate expression of the two subunits during the assembly of Rubisco (136). Further insights have come from the studies on the effect of oxidative stress in C. reinhardtii (26, 27). Light can cause oxidative stress whenever the reducing power generated by the primary reactions of photosynthesis in the thylakoid membrane exceeds the capacity of the downstream reactions resulting in an increase of oxygen radicals and in a shift of the glutathione pool toward its oxidized form. Under these conditions, translation of the large subunit (RbcL) of Rubisco is arrested and resumes only after the cells recover from this oxidative stress. The proposed mechanism of this translational arrest of RbcL is that oxidation of the glutathione pool leads to the oxidation of the thiol groups of RbcL (26). This event unmasks a domain in the N-terminal part of RbcL, which is normally buried, and which resembles the RRM domain of RNA-binding proteins (141). This exposed motif binds any RNA in its vicinity, in particular its own mRNA, and thereby inhibits the translation and assembly of Rubisco (26). Interestingly, the RNA-binding activity of RbcL was only detected under oxidizing conditions in the presence of oxidized glutathione. An RNA-binding activity with a dissociation constant in the nanomolar range as well as the translational arrest under oxidizing conditions were also found for Rubisco from a wide evolutionary range of photosynthetic organisms, including photosynthetic bacteria and land plants, suggesting that this proposed autoregulatory mechanism of translational control of RbcL in C. reinhardtii may also be valid for these organisms. Because photosynthetic bacteria contain a Rubisco dimer without small subunit, it is likely that the masking of the RNA-binding motif occurs when the RbcL dimer is formed or when a chaperone interacts with RbcL before its dimerization during the assembly of Rubisco (27).
Conclusions and Perspectives
In recent years, there have been major advances in our understanding of the composition, structure, and assembly of the photosynthetic complexes in the thylakoid membranes. An important feature of this membrane system is its extraordinary dynamics, which becomes apparent under changing environmental conditions, which include changes in light intensity, spectral light quality, temperature, and nutrient and CO2 availability. Photosynthetic organisms have evolved complex processes that allow them to adapt to these changes. A particularly interesting aspect of these acclimation responses is that the photosynthetic apparatus can switch from a light energy collector to a light energy dissipator within a few seconds depending on whether the incident light is limiting or exceeds the absorption capacity of the system. The first case is illustrated by state transitions, which involve a dynamic remodeling of the thylakoid membrane and diffusion of macromolecular complexes from the grana to the lamellar thylakoid regions, leading to a rebalancing of the light energy absorbed by the two photosystems. In the second case, high light leads to the formation of a large proton gradient across the thylakoid membrane, which induces conformational changes in LHCII, resulting ultimately in a harmless heat dissipation of the excess absorbed excitation energy. Another remarkable feature is that in the case of PSII, this complex is able to repair the photodamage that occurs as an unavoidable consequence of its particular photochemistry. A complex repair cycle occurs, involving sequential phosphorylation and dephosphorylation of the PSII core proteins and diffusion of the damaged complexes from the grana to the stromal regions, where the damaged D1 is replaced by newly synthesized D1 protein. While these processes take place within the second-to-minute range, other responses occur over longer periods and involve the chloroplast and nuclear gene expression systems, both of which are influenced by the redox state of the PQ pool and the FD/TRX system. This redox state is in turn modulated through environmental conditions such as irradiance, CO2, and nutrient availability. How the redox state of the PQ and the FD/TRX pool triggers signals within the plastid to the plastid genetic system and from the chloroplast to the nucleus to elicit an appropriate response is still largely unknown, although several factors that participate in these signaling chains have been identified through genetic and biochemical means. A challenging task will be to determine how the identified chloroplast kinases and phosphatases are linked with each other within a large network, and how changes in the redox state of the photosynthetic electron transport chain are transduced into changes in the chloroplast and nuclear gene expression ultimately leading to the acclimation response.
Footnotes
Acknowledgments
I thank Nicolas Roggli for preparing the figures and Michel Goldschmidt-Clermont for critical reading of the manuscript. The work in the author's laboratory was supported by the grant 31003A_133089/1 from the Swiss National Foundation.