
Abstract
Singlet Oxygen in Photosynthetic Organisms
All organisms living in an oxygen-rich atmosphere are frequently confronted with oxidative stress. Plants are especially exposed to oxidative stress evoked by 1O2 because of their pigments that act as photosensitizers. Some photosensitizers can transfer the absorbed light energy to 3O2 and directly produce 1O2 (52). In photosynthetic organisms, 1O2 is most commonly produced by the photosensitizer chlorophyll (Chl), which acts as the main light-absorbing pigment in organisms able to perform oxygenic photosynthesis. Chl a has a very high extinction coefficient at about 670 nm in vivo, and its excited state is long-lived (up to a few nanoseconds). so that during photosynthesis, efficient conversion of the excitation energy into an electrochemical potential via charge separation in the photosystems (PS) is possible. If no charge separation takes place, the spins of the electrons in the excited state of the Chl a molecule can reorientate and give rise to the lowest energy-excited state, the Chl triplet state (3Chl) (123). This 3Chl can react with 3O2 to produce 1O2 if no efficient quenchers are close by (68) (Fig. 1).
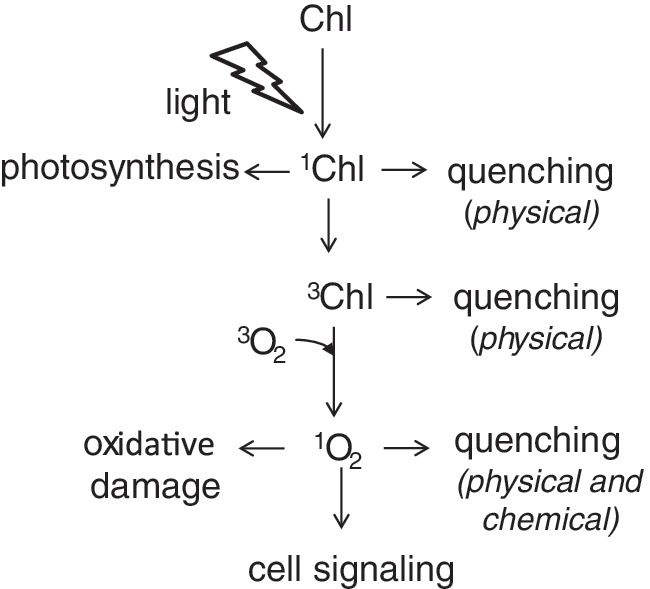
Photosynthetic organisms have evolved a number of molecules that efficiently quench or scavenge 1O2 in the photosynthetic membranes and prevent damage under normal conditions. However, under some environmental conditions such as during cold or dry periods or exposure to high light intensities, these protection mechanisms can be overwhelmed, and 1O2 levels increase in the chloroplasts. Under these conditions, 1O2 was also shown to function as a signaling molecule that can trigger cellular responses such as acclimation, repair mechanisms, and activation of a programmed cell death (PCD) response.
Formation of Singlet Oxygen
Singlet-oxygen generation in the antenna
In plants and algae, Chl is located in the reaction centers of the PS, in the antenna systems, and in the cytochrome (Cyt) b6f complex. The majority of Chl is found in the antenna, and 3Chl might be produced by an intersystem crossing from excited Chl (1Chl). In isolated antenna complexes, the rate of intersystem crossing is significant (75), and the formation of 3Chl (131) and 1O2 (125) has been shown in vitro. However, in these studies, antenna complexes were isolated using detergents. In such preparations, a higher probability of 1O2 generation seems to be more likely than in the native complexes because of detergent-induced instabilities of the protein/pigment complexes. In the native antenna complexes, most of the formed 3Chl is efficiently quenched by nearby carotenoids such as xanthophylls (64). The nature and number of the carotenoids bound in the antenna complexes and in the reaction centers are well known from structural data (4, 26). The PSII complex contains 2 β-carotenes in each reaction center, 1 lutein and 1 violaxanthin in CP24 and 1 lutein, 1 neoxanthin, and 1 violaxanthin in CP29 and CP26, respectively. Further, depending on the light intensity and the length of the photoperiod, the variable number of light-harvesting complex (LHC)II trimers are associated with an individual PSII complex (87), each Lhcb polypeptide containing four carotenoids (1 neoxanthin, 2 luteins, and 1 violaxanthin). At PSI, about 20 carotenoids are located in the reaction center, and two xanthophylls are bound in each of the four Lhca polypeptides of the single LHCI per PSI. In addition, there is a pool of carotenoids that cannot be assigned to individual polypeptides on either the LHCI or PSI core. The physical quenching of 3Chl requires an edge-to-edge distance within the van der Waals distance (3.6 Å) between the Chl and carotenoid, so that the electron orbitals have some overlap. In this spin-exchange reaction, the triplet state of the carotenoid is formed, which can dissipate the excess energy as heat (30, 125).
The capacity of individual xanthophylls in the different locations of the PSII antenna complexes to quench 3Chl has been studied in detail by Mozzo et al. (100). These authors concluded that the xanthophylls bound in LHCII quench 95% of the 3Chl states. When measured, the carotenoid-to-Chl ratio of thylakoids from Arabidopsis thaliana grown in short-day photoperiods is 1.5-fold higher than expected from the above listing, even though this ratio can vary depending on the lipid composition of the membrane and between the A. thaliana ecotypes (163). Still, this indicates the presence of carotenoids also in the lipid matrix. Thus, the remaining 5% of 3Chl from LHCII may generate 1O2 that can either be quenched by carotenoids located at the interface between lipids and proteins or escape into the thylakoid membrane where it may be quenched by the free carotenoids in the lipid matrix (66).
Singlet-oxygen generation in the PS II
Most 1O2 is, however, produced in the reaction center of PS II (Fig. 2) [for extensive reviews, see (76, 77)]. Excitation of Chl leads to charge separations in the reaction center of PSII, and the electrons are transferred in the so-called linear electron flow through the whole electron transport chain to the final electron acceptors, mainly NADP+, at the acceptor side of PSI. The probability of 1O2 generation is very low when sufficient electron acceptors are available at a given light intensity and physiological state of a plant. However, when the light absorption exceeds the capacity of photosynthetic electron transport, the probability of 1O2 generation increases. Under these conditions, the plastoquinone pool is largely reduced, and charge recombination reactions within PSII take place, leading to the generation of 3P680 (P680, the Chl being the primary electron donor). The two β-carotene molecules present in the PSII reaction center are placed too far from P680 to prevent this process [for location of the β-carotenes and P680 in the reaction center, see Fig. 2 and (153)]. 3P680 reacts subsequently with 3O2 to form 1O2 and P680 (Fig. 3).


The pathway of charge recombination depends on the energetics of the electron acceptors of PSII (for details, see Fig. 3). Charge recombination between the primary quinone acceptor, QA, and the positive charge at the donor side of PSII (P680 +) can proceed either via a repopulation of the primary radical pair (P680 +Ph−) (indirect pathway, radiative or nonradiative) or directly into the ground state of P680 (nonradiative pathway). The indirect pathway can lead to the formation of the dangerous triplet states of the primary charge pair and finally to 3P680, while the direct, nonradiative pathway is safe and does not lead to the generation of 1O2. It has been shown that the midpoint potential of the redox couple (QA/QA −) controls the pathway of charge recombination and the yield of 1O2 formation (44, 45, 78). The modification of the midpoint potential of QA in PSII lacking or containing an inactive water-splitting complex without Ca2+ (79) or in the presence of a high proton gradient (81) is an important regulation mechanism to protect PSII from 1O2 generation and subsequent photo-oxidative damage. The influence of the midpoint potential of QA on the yield of 1O2 generation has also been demonstrated by the use of herbicides that bind to the QB-binding pocket (Fig. 2). 3-(3,4-Dichlorophenyl)-1,1-dimethylurea shifts the midpoint potential of QA to a more positive value and lowers thereby the yield of 1O2 generation. Phenolic herbicides lower the redox potential of QA and increase the yield of 1O2 formation (45, 78). In addition, the midpoint potential of QA was changed when the alanine at position 249 of the D2 protein was changed to a serine by site-directed mutagenesis, and this also affected 1O2 generation (44). The midpoint potential of the pheophytin-redox couple (Ph/Ph−) influences also the probability of the nonradiative pathway of charge recombination between the primary charge pair (126). Interestingly, cyanobacteria have two genes for distinct D1 proteins, a main subunit of the PSII reaction center, with different amino acids at positions D1–130. D1-E130 proteins are expressed only during high light conditions (73) and probably differently affect the redox potential of Ph to enhance the charge recombination via the nonradiative pathways and thereby lowering the yield of 1O2 generation (155). Similar to the high-light D1 protein in cyanobacteria, in all higher plants with known sequences, a glutamate occupies the position D1–130.
Photosynthetic organisms have evolved a number of protection mechanisms to prevent the formation of 1O2, including the dissipation of absorbed light energy as heat by the process of nonphotochemical quenching [for review, see (64, 102)]. Still, PSII is the main target of light-induced damage, the so-called photoinhibition, caused by unfavorable environmental conditions such as, drought, cold, and high light illumination. During this process the D1 protein is triggered for degradation, most likely by the action of 1O2 and/or other reactive oxygen species (ROS), even though direct experimental evidence for the oxidation of specific amino acid residues of D1 is still missing. During the turnover of the D1 protein, the pigments and other cofactors have to be stored in a safe way, so that no photosensitizing reactions take place by the free pigments. Free Chls may be bound by Early Light-Induced Proteins (ELIPs) and related proteins to prevent the formation of 1O2 (1). Heddad et al. showed that the transcripts of one isoform of ELIPs increased linearly with light intensity and the extent of PSII photodamage (54). In aging cauliflower leaves, a water-soluble Chl-binding protein (WSCP) is expressed. This protein is a tetramer, binds two Chl molecules, and does not contain any carotenoids. Upon excitation, 3Chl, but no 1O2, is produced; thanks to a diffusion barrier that prevents the interaction of 3Chl with 3O2 (124, 132).
Singlet-oxygen generation by other components of the photosynthetic apparatus
It is generally accepted that PSI does not contribute to 1O2 generation in photosynthetic organisms. It has recently been proposed that back-reaction pathways in PSI leading to 3P700 formation are specifically minimized by redox tuning to avoid 1O2 generation (126). Still, in PSI, illuminated under reducing conditions or when vitamin K1 is removed from the reaction center, charge recombination reactions do occur leading to the formation of 3P700 [for a review, see (14)]. However, in PSI, the lifetime of the 3P700 state is about 6 μs and is not shortened by the presence of 3O2, indicating that P700 is screened from O2 (134). On the other hand, it has been shown recently in a mutant with low amounts of carotene that a higher amount of 1O2 is formed in PSI-LHCI supercomplexes compared to the wild-type, and that its PSI is more susceptible to photo-oxidative damage than PSII (17). In this mutant, enhanced photodamage of PSI may be either due to reduced quenching by β-carotene and higher 1O2 formation in the reaction center, less-stable PSI reaction centers lacking a structural component or due to a more vulnerable carotenoid-deficient LHCI. Further experimental evidence is needed to show whether any significant amount of 1O2 is generated via charge recombination reactions and 3P700 formation in PSI.
The Cyt b6f complex is a dimer, and each monomer contains four major and four minor protein subunits (82, 143). Three of the major subunits bind the redox-active cofactors, the Cyt f with a c-type heme, the Cyt b6 with two b-type hemes, and the so-called heme x, and the Rieske protein with a [2Fe-2S] cluster. In addition to these redox-active cofactors, the Cyt b6f complex contains a carotene (β-carotene in most species) and a Chl a molecule that is potentially photoactive. However, structural analyses revealed that specific local protein structures involving the aromatic amino acid phenylalanine in subunit IV are responsible for shorting the excited 1Chl lifetime and thus decreasing the probability of 1O2 formation (24, 161). Since this Chl a is not needed for the electron transfer reactions, the authors concluded that it seems to play rather a structural role in the Cyt b6f complex (161). This is supported by the fact that Cyt b6f complexes from etioplasts contain, instead of Chl a, a molecule of protochlorophyll a required for correct assembly of the complex (121). On the other hand, it has been demonstrated that the Chl a can generate 1O2 when the intact or the Rieske protein-depleted Cyt b6f complex was exposed to high light (130). In this study, the neighboring carotene seemed unable to detoxify the 3Chl state by physical quenching; however, chemical quenching of 1O2 by β-carotene was demonstrated. The same authors showed that the level of 3Chl and 1O2 generation increased with an increased structural disturbance of the Cyt b6f complex, indicating that in intact, enzymatically active complex 1O2 formation is rather low (92). Thus, 1O2 might not be produced in significant amounts by the Cyt b6f complexes in vivo.
Singlet-oxygen generation by Chl precursors and catabolites
Assembly and disassembly of the protein/pigment complexes of the photosynthetic electron transport chain are critical processes that have to be strictly regulated to limit the danger of 1O2 formation by immature, not fully functional complexes and by free metabolites/catabolites that can act as photosensitizers (Fig. 4). Most of these metabolites are Chl precursors such as protoporphyrin IX and protochlorophyllide, even though it has been suggested recently that also carotenoid intermediates such as tetra-cis-lycopene might act as photosensitizers and generate 1O2 (53). In etiolated pea epicotyls, porphyrin-type Chl precursors were shown to be capable of photogenerating 1O2 (32). In a functional system, the accumulation of Chl precursors is prevented by metabolic control and by channeling the metabolites from one enzyme to the next (20, 147). Only the amount of pigments is synthesized, which is used in the specific pigment-binding sites of a given protein. In the A. thaliana fluorescent (flu) mutant a protein is mutated that plays a key role in the negative feedback regulation of Chl biosynthesis, and consequently, protochlorophyllide accumulates in the dark and leads to the generation of a high amount of 1O2 when the flu plants are transferred to the light (109). This mutant was extensively used to study the 1O2 response of A. thaliana (see Cellular Response to Singlet Oxygen). One has to keep in mind that in the flu mutant, protochlorophyllide accumulates in the chloroplast envelope and in the thylakoid membrane (8), and therefore 1O2 is partially produced at a site that is different from its production sites in a wild-type plant. Addition of herbicides that block the protoporphyrinogen oxidase can lead to the accumulation of protoporphyrin IX, a photodynamic pigment that also does produce high amounts of 1O2 in the light (11).
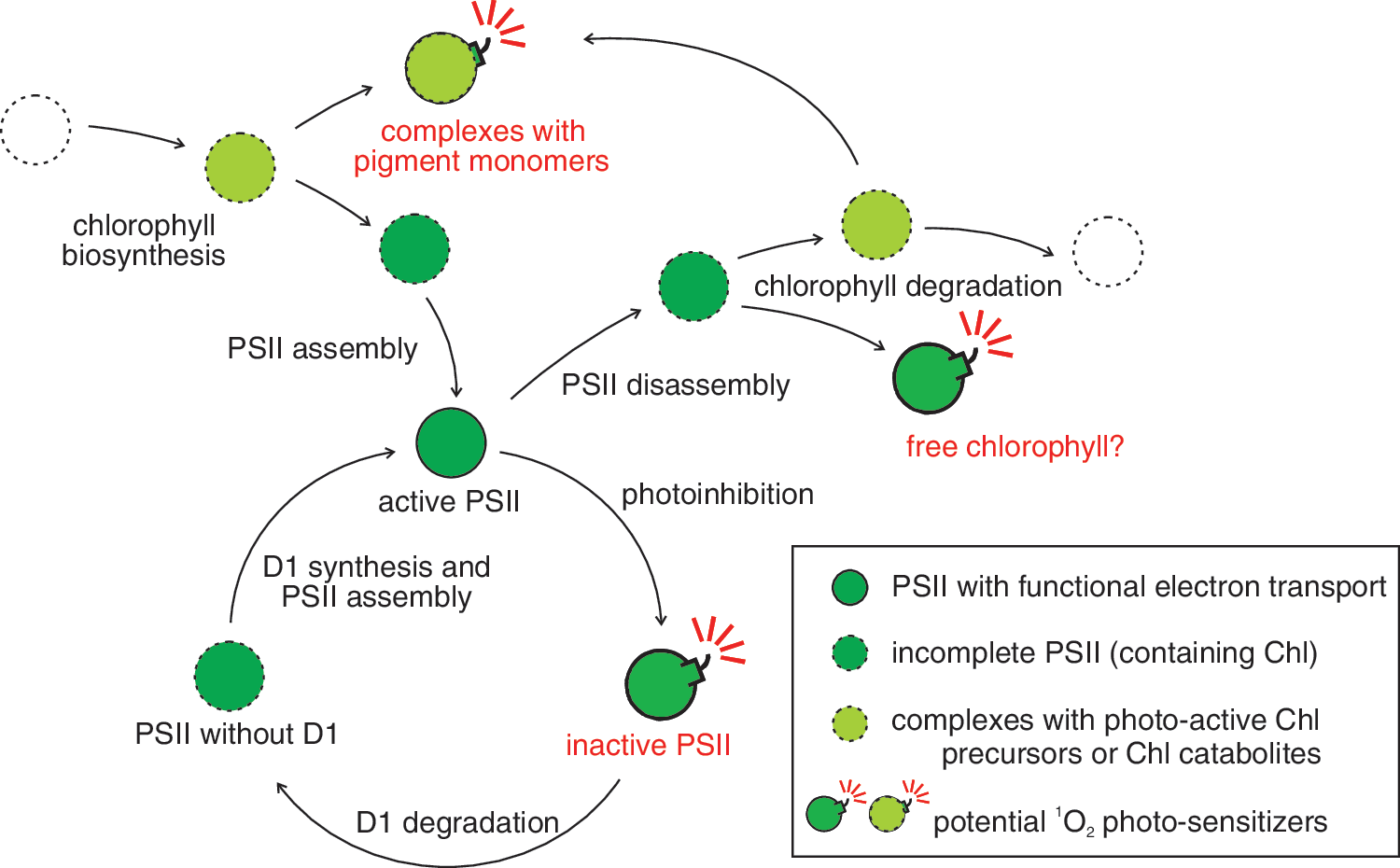
To avoid 1O2 production and photo-oxidative damage during Chl synthesis, the precursors are not set free, but are bound to the enzymes involved in synthesis or to specialized protection proteins. Light-harvesting CP-like proteins (LIL) have been identified to participate in this process, and the isoform LIL3 is synthesized during de-etiolation (122). It carries the late metabolites of Chl biosynthesis (122) and interacts physically with geranylgeranyl reductase (CHLP) (146). It has been suggested that LIL3, CHLP, and other enzymes of late Chl biosynthesis form a protein complex close to the site of Chl insertion into its binding sites at the LHC proteins. The same may hold for the insertion of Chl into the reaction centers. Such a complex could allow the efficient channeling of Chl and its photo-reactive precursors into the PSs and thus prevent the generation of 1O2 during the greening and assembly process of the photosynthetic electron transport chain.
During leaf senescence, Chl is degraded in a controlled way to avoid the accumulation of phototoxic intermediates of Chl breakdown such as pheophorbide and the red Chl catabolite (RCC) [for a review on Chl degradation, see (63)]. It was recently shown that during senescence, the protein STAY-GREEN and five Chl catabolic enzymes, involved in the conversion of Chl a to a primary fluorescent Chl catabolite, localize to LHCII and form a complex (128). Metabolic control by channeling through this complex occurs in Chl breakdown at the LHCII (106), and it is likely that similar complexes exist at LHCI and other CP, so that the risk of accumulating Chl breakdown intermediates is minimized. Overexpression of STAY-GREEN proteins in A. thaliana and rice also stimulated 1O2 production (65, 104). Similarly, mutants deficient in the enzymes Pheide a oxygenase or RCC reductase accumulate phototoxic intermediates and promote light-dependent 1O2 production and cell death (114, 115). RCC reductase expression is induced during pathogen infection, and the protein might play a role during a pathogen-induced cell death response (162). Interestingly, the RCC reductase was recently found to be localized in both the chloroplasts and mitochondria, and increased 1O2 formation was detected in the mitochondria of RCC reductase-deficient mutants (110). The authors speculated that photoactive RCC might penetrate from the chloroplasts to the mitochondria, where a 1O2 burst stimulates pathogen-induced cell death response, and that RCC reductases might control this response. This implements an important signaling role of Chl catabolites and 1O2 in the cellular response to pathogen infection.
Detection of Singlet Oxygen
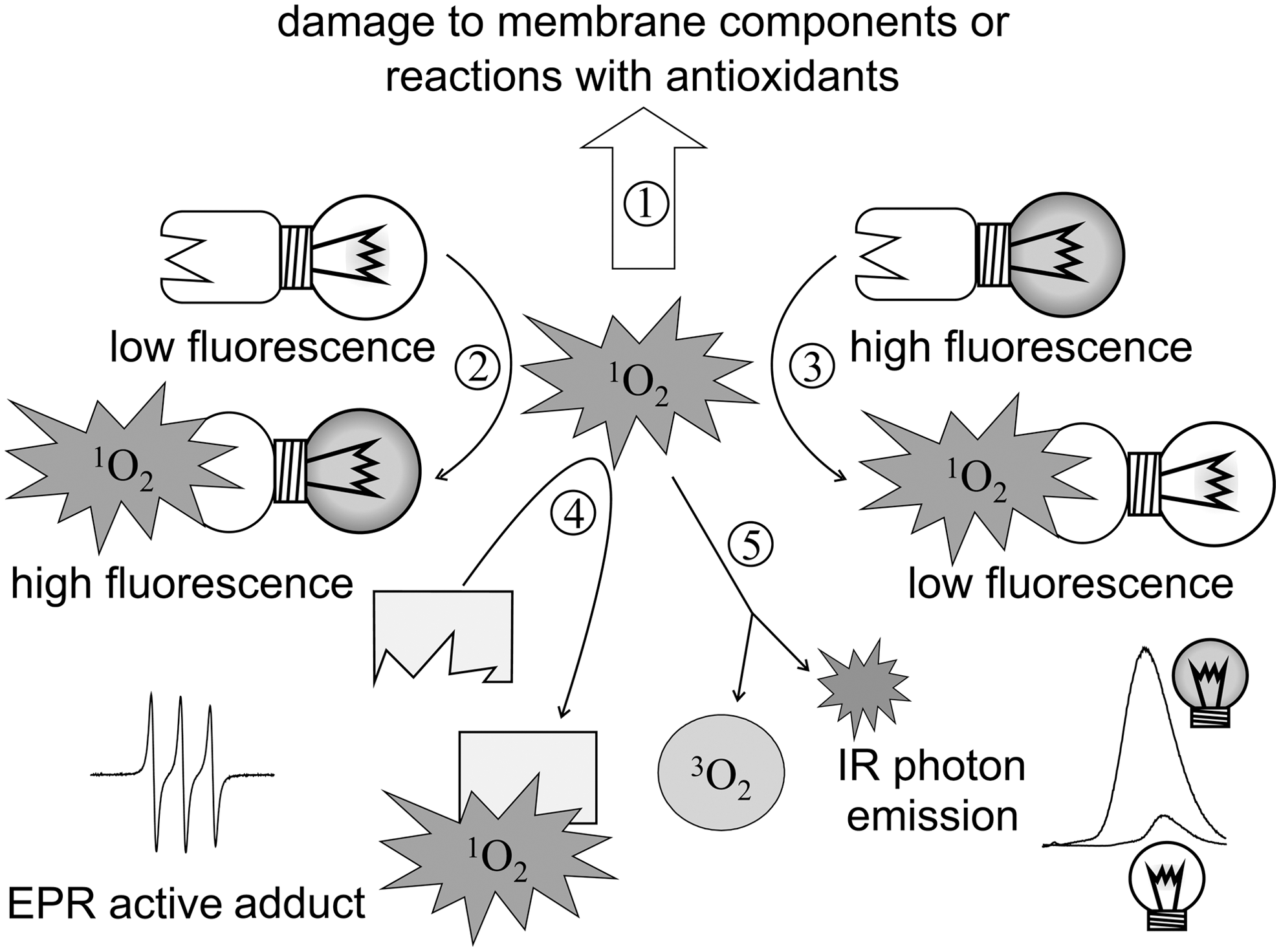
The importance of 1O2 in physiological stress response calls for a reliable means of detection. Direct detection and, what is more, quantification are hampered by the fact that 1O2 is reactive to a variety of biological molecules and is quenched by water. In this way, efficient 1O2 reporter molecules are to compete with target molecules of oxidative damage, 1O2-quenching antioxidants, as well as with solvent molecules. Upon oxidation by 1O2, chemical traps change their optical properties (absorption, fluorescence, or both) or their electron paramagnetic resonance (EPR) properties (Fig. 5).
EPR spectroscopy
Since its introduction as a 1O2 trap more than three decades ago, 2,2,6,6-tetramethylpiperidine (TEMP) and its derivatives have been used extensively. When reacting with 1O2, this sterically hindered amine is converted into a nitroxide radical that (unlike TEMP itself) is paramagnetic and thus detectable by EPR spectroscopy (90). A structurally similar, but more hydrophilic, 1O2 probe is TEMPD (2,2,6,6-tetramethyl-4-piperidone) (99). Because the product is more or less a stable free radical, the method is often referred to as 1O2 spin trapping, although 1O2 is not a radical itself. The first use in the isolated thylakoid membranes (60) showed that the resulting spin-adduct is prone to reduction into an EPR-silent hydroxylamine, and thus the method may underestimate the amount of 1O2 produced in the sample (62). It is also important to note that commercially available TEMP needs to be purified by vacuum distillation before use (45, 57), because even small amounts of contaminants inhibit the PSII electron transport (51, 57). TEMPD-HCl is preferred to TEMPD, because the hydrochloride form is more stable in storage and is only converted to the nonchlorinated TEMPD when dissolved in water-based measuring buffers (57). With care and adequate controls, 1O2 spin trapping is a sensitive tool that has been applied in a number of studies on isolated functional photosynthetic membranes (37, 44, 45, 60, 61, 80). However, the application of TEMP in vivo in leaves is hampered by the sensitivity of both trap and adduct to redox-active metabolites. Attempts to overcome this problem by replacing the piperidine ring with a theoretically less-sensitive pirrolydine were not successful (62), leaving 1O2 trapping by TEMP and its derivatives in the domain of in vitro experiments.
Changes in absorption or fluorescence of specific singlet oxygen probes
1,3-diphenylisobenzofuran (DPBF) reacts with 1O2 rapidly and irreversibly in a cycloaddition reaction, yielding an unstable endoperoxide that decomposes to give 1,2-dibenzoylbenzene. Products have lower blue fluorescence (excitation/emission 410/455 nm) than DPBF itself (159), and DPBF has been a popular 1O2 probe for decades [for examples, see (113, 164)]. DPBF can be useful in pure 1O2-yielding systems such as pure photosensitizers aimed at studying the properties of 1O2 (133), but it must be used with caution for the detection of 1O2 in complex biological samples, because it also reacts with different radical species, such as peroxyl or alkoxyl radicals (16, 158). The potential use of DPBF in plant 1O2 studies is limited by its low solubility in water, and also because its blue fluorescence may be absorbed by Chls.
The 440-nm absorption maximum of a water-soluble compound, p-nitroso-dimethylaniline (RNO), also appears disadvantageous from the perspective of Chl-rich samples. Nevertheless, a photometric technique based on observing RNO bleaching caused by a product of the reaction between 1O2 and histidine (74) has been successfully applied for detecting 1O2 production by PSII reaction centers isolated from Pisum sativum (148). Further, using an exogenous photosensitizer as an artificial 1O2 source, the RNO original method has also been developed into an assay measuring the 1O2-quenching capacity of a plant extract (94). The assessment of this parameter is not a direct indicator of actual 1O2 production rates, but a high quenching capacity certainly indicates a need for enhanced protection against putative high fluxes of 1O2 in the plant itself.
Other probes with more perspectives for in vivo studies include molecules comprised of a trapping moiety and a chromophore. Upon reaction with 1O2, the resulting oxygen adduct changes the intramolecular energetics and thus affects light emission of the chromophore. Such fluorescent or luminescent ROS probes have great perspectives in imaging techniques with potentials to identify 1O2 in situ. In one approach, developed by Kálai et al., 1O2 trapping converts the reporter molecule from a high to a low fluorescent form (67). Fluorescence quenching of 3-(N-diethylaminoethyl)-N-dansyl)aminomethyl-2,5-dihydro-2,2,5,5-tetramethyl-1H-pyrrole (DanePy) and its derivatives have been successfully applied in a variety of experiments to detect 1O2 in A. thaliana (59, 109), Vicia faba (58), spinach (7), or tobacco (55) leaves, and in Chlamydomonas reinhardtii cultures (39). Major disadvantages of this probe are that (i) fluorescence quenching by 1O2 is only partial and thus it is useful in detecting high fluxes of ROS only (58), and that (ii) it requires ultraviolet [345 nm, (67)] fluorescence excitation, which may interfere with plant metabolism. For successful delivery into photosynthetic tissues, DanePy needs to be infiltrated directly into the leaf (56); otherwise, it tends to concentrate in vascular tissues (43). Consequently, the use of this probe in hard leaf tissues such as of grapevine or Tilia was not found to be feasible (Hideg É, unpublished). Although europium (III) complexes have not been tested in plants so far, these probes that are converted into compounds emitting red light upon UV (294–335 nm) excitation by 1O2 (140) may also suffer from the problem of high-energy excitation.
A reverse chemistry governs the functioning of chemical 1O2 traps in which photoemission from the chromophore is quenched by electron transfer from the adjacent trapping moiety before a reaction with 1O2; therefore, these molecules have very low (if any) light emission in the absence of ROS. Upon reaction with 1O2, the resulting oxygen adduct is no longer an efficient intramolecular electron donor, and light emission occurs (140, 141, 145). The advantage of these chemical traps is a wide range of response, from virtually zero to intense light emission.
Nagano and coworkers developed a series of fluorescent probes in which fluorescein is fused with a 9,10-dimethylanthracene moiety as a chemical trap of 1O2. Endoperoxides formed from the probes upon reacting with 1O2 emit green fluorescence (with a maximum intensity at 516 nm) upon blue (492 nm) excitation (145, 154). While these probes have been successfully applied for detecting 1O2 in a variety of medicinal chemistry experiments (105), to our best knowledge, there is only one reported application in plants, in which 1O2 was found in isolated intact chloroplasts under photo-oxidative stress (160).
Singlet Oxygen Sensor Green® [SOSG (141)] features fluorescence excitation and emission in the visible and has been applied to detect 1O2 in photoinhibited Arabidopsis leaves (42). However, the probe has shown sensitivity to both visible (55) and UV light (117), which is undesirable in most plant experiments. In addition, recent studies showed that both SOSG and the immediate product of the reaction between SOSG and 1O2 (the SOSG endoperoxide) are capable of sensitizing the production of 1O2 (47, 117). Although SOSG is marketed as a cell-impermeable compound (141), Gollmer et al. recently reported that it penetrated the living mammalian cells (47). In tobacco leaves infiltrated with SOSG, the probe was mainly found in the epidermal cells, but was also preferentially associated with the nucleus (55). Nevertheless, SOSG may be used for detection of 1O2 in in-vitro assays. SOSG has been applied successfully in isolated antenna (LHCII) and in PSII core complexes and PSII-LHCII, PSI-LHCI supercomplexes (17, 22). Differences in 1O2 generation depending on the pigment composition of these complexes were convincingly detected.
In addition to being prone to artifacts, another limitation of chemical probes is that they may also respond to ROS other than 1O2. Although excluding such cross-reactivity is a key aspect of all reports on new chemical 1O2 probes, testing for each and every reactive agent occurring in biological systems under oxidative stress (including radicals other than oxygen-centered ones) is certainly a formidable task. Another disadvantage of chemical 1O2 probes is that their versatility depends on the successful delivery into the very site of 1O2 production.
Near-infrared luminescence emission of singlet oxygen
The miniature fraction of 1O2 that undergoes radiative deactivation is a unique in situ signature of this molecule. In this way, this low-intensity near-infrared luminescence around 1270 nm provides the only undoubtedly direct method to detect 1O2. However, there are no gains without pains: detecting time-resolved 1O2 luminescence requires sensitive optical detectors with a low signal-to-noise ratio as well as careful signal analysis to separate the true 1O2 signal from scattered light and from phosphorescence and delayed fluorescence emissions from other molecules. In plant samples, where Chl is a major source of 1O2, the excited pigment may also relax through long-wavelength phosphorescence. Separation of infrared Chl phosphorescence (between 900 and 1000 nm) and 1O2 phosphorescence (at 1270 nm) is possible by measuring the emission spectra and also by fitting the time-resolved luminescence signal into components. In addition to its unambiguity, the advantage of detecting 1O2 using luminescence over chemical trapping is that with careful analysis, the luminescence method also reveals the lifetime of 1O2 in the studied system. This parameter is fundamental for studies on both the physiology of oxidative damage and on signaling events, since it has the information on putative distances to which 1O2 may reach from its production site. There are numerous studies on analyzing 1O2 luminescence decay in medical samples or model systems undisturbed by the presence of Chl, but these applications are not reviewed here. Interested readers are referred to a recent comprehensive study and references therein (21).
The first utilization of 1O2 infrared luminescence was the demonstration of 1O2 in isolated PSII reaction center under high-light stress (93). This was followed by a number of reports on 1O2 luminescence decay analyses in PSII reaction center preparations from leaves (88, 149), or PSII particles from Synechococcus (27) or Synechocystis (150), and one report identified the spectrum of 1O2 photoemission in the Cyt b6f complex (92). Analyzing the time-resolved infrared emission spectra of PSII particles, Dědic et al. determined both the submicrosecond lifetime of Chl triplet states and the microsecond lifetime of 1O2 [18 μs and 3–4 μs in D2O and in H2O buffers, respectively (27)]. These authors also showed that the quantum efficiency of 1O2 generation in PSII particles substantially increased by addition of sodium dodecyl sulfate, which inactivated the complex and released Chl molecules from their specific binding sites on PSII proteins and thus abolished the effective interaction with quenchers. Although there are no experimental data available, the presence of a variety of additional quenchers in chloroplasts suggests a much shorter lifetime of 1O2 and thus a more difficult task of detection by infrared emission. Based simply on the concentration of ascorbate in the stroma of chloroplasts [20–300 mM, (139)] and pH-dependent rate constants for quenching of 1O2 by ascorbic acid/ascorbate (12), Li et al. estimate a short, ∼200-ns 1O2 lifetime and thus predict serious technical obstacles in detecting the luminescence of endogenously produced 1O2 in chloroplasts (88).
Reaction of Singlet Oxygen with Cellular Components
Main targets of singlet oxygen damage
The main targets of damage by 1O2 include lipids, proteins (25), and nucleic acids (137). Oxidized products are not only of impaired biological function, but may also initiate further biological damage, either as oxidizing (such as lipid peroxidation products or oxidized amino acid intermediates) or as genotoxic (oxidized DNA) agents. For example, 1O2 reacts with the guanine moiety in DNA to form 8-oxo-7-hydrodeoxyguanosine (8-oxodG; also called 8-hydroxydeoxyguanosine) and 2,6-diamino-4-hydroxy-5-formamidopyrimidine, which stimulate the accumulation of point mutations (137). Further, 1O2 also causes alkali-labile sites and single-strand breaks in DNA (13).
Most reactions of 1O2 with amino acids occur via chemical quenching, although tryptophan has been reported to be capable of physical quenching too (95). During chemical quenching, primary oxidation mainly occurs on the side chains of tryptophan, tyrosine, histidine, methionine, and cysteine. The damage of a protein does depend not only on the number of sensitive side chains but also on the structure of the protein. A conformational change in a protein can either expose or hinder the access of 1O2 to specific amino acid residues. The major consequence of 1O2-mediated protein oxidation is cross-linking and aggregation (25). Oxidation of tryptophan, tyrosine, or histidine includes peroxide intermediates, and the relatively long lifetimes of these hydroperoxides may allow these species to diffuse considerable distances and thereby initiate damage at sites remote from their initial site of generation (49). In the photosynthetic apparatus, the main protein target of 1O2-mediated damage is the D1 protein. Attempts have been made to identify the amino acid residues that are oxidized by 1O2 (136). However, a study with the modern mass spectrometry technology is needed to identify the main targets.
Although oxidative damage of lipids is generally attributed to radical-mediated chain reactions initialized by hydroxyl radicals (type I reaction), studies with phospholipid model membranes showed that 1O2 is also capable of oxidizing PUFA (type II reaction) (Fig. 6). Linoleic acid is a major PUFA in photosynthetic membranes and also the main target of 1O2-induced lipid peroxidation, leading to the formation of different lipid hydroperoxides (LOOH). It was shown that 10′- and 12′-hydroperoxides of linoleic acids (hydroperoxy-octadecatrienoic acid [HPOTE]) are specifically generated by 1O2-stimulated peroxidation, whereas 9′- and 13′-hydroperoxides are formed by both type I and II reactions (142). On the other hand, the accumulation of LOOH formed by type II reactions was shown to stimulate radical-mediated lipid peroxidation in the presence of exited photosensitizers, leading to faster generation of type I LOOHs after a prolonged exposure to 1O2 (142). Moreover, LOOH produced during the oxidation by hydroxyl radicals can generate 1O2 by the Russell mechanism in vitro (97), and similar reactions have been suggested for DNA or protein hydroperoxides (98). Thus, it is important to efficiently detoxify LOOHs by either peroxidases or reductases, leading to the formation of hydroxy fatty acid isomers such as 13-hydroxy octadecatrienoic acid (13-HOTE) or 13-hydroxy octadecadienoic acid (13-HODE). Further, LOOHs can be converted to keto-forms such as 13-keto-octadecatrienoic acid (13-KOTE) and 13-keto-octadecadienoic acid or to other reactive electrophile species (RES), which were shown to play an important role in cellular signaling during 1O2 response (see “Reactive electrophiles as transmitters of singlet-oxygen signals”) (101).

Singlet-oxygen quenchers
The high reactivity of 1O2 with essential biomolecules and the subsequent oxidative stress increase the need for efficient protection mechanisms. Protection of cells from oxidative stress generally includes some ROS-specific enzymes; however, there is no detoxification enzyme targeted to 1O2 only. Still, there are many antioxidants involved in 1O2 detoxification, and several excellent reviews summarize the 1O2-neutralizing pathways (5, 31, 72, 108, 151) of which a detail discussion would go beyond the scope of the present work. The aspect to be emphasized here is the variety of products yielded from reactions of 1O2 with target molecules, including antioxidants.
As illustrated in Figure 6, 1O2 that is produced by triplet-singlet energy-transfer reactions can undergo both physical and chemical quenching (scavenging) (30). The ability of carotenoids to quench 1O2 increases with the number of conjugated double bonds (27). In physical quenching, a singlet-triplet energy transfer dissipates the energy as heat and does not modify the reaction partner (e.g., β-carotene) chemically, while chemical quenching results in oxidized products. Depending on whether the molecule oxidized by 1O2 is regenerated (rereduced) or remains nonfunctional, chemical quenching can be beneficial or damaging. Although theoretically any reaction lessening the amount of 1O2 in the cell is antioxidative, in practice, only pathways including rapid replacement of the oxidized target molecules, are regarded as antioxidants. In plant cells, tocopherol (106), carotenoids (28), and ascorbate (12) are the most abundant 1O2 antioxidants. Tocopherol is oxidized to a free radical product and is regenerated by ascorbate, whereas oxidized ascorbate is reduced in the reactions involving specialized enzymes, glutathione, and NADPH. These regenerative pathways are also included in defense against ROS other than 1O2 (52).
In the reaction center of PSII, β-carotene does not quench the P680 triplet physically (29), but it was shown to be a chemical quencher (148). Recently, specific endoperoxide molecules that resulted from 1O2 oxidation of β-carotene, lutein, and zeaxanthin in Arabidopsis leaves were characterized (118). This carotenoid oxidation results in products such as β-cyclocitral and β-ionone (118, 119), and oxidized carotenoids have to be replaced by de novo synthesis. When plants are exposed to high-light stress, the level of β-carotene endoperoxides increases rapidly and much faster than the level of lipid peroxidation products, indicating that these carotenoid oxidation products are an early index for 1O2 production (152). Plastoquinol has been shown recently to be a more-efficient 1O2 quencher than tocopherol during exposure of plants to high light. Plastoquinone-9 reacts with 1O2 to form plastoquinone-C (107). However, the concentration of plastoquinol is much lower than that of tocopherol, indicating that overall tocopherol may be the more important quencher.
Cellular Response to Singlet Oxygen
The high reactivity of 1O2 with biomolecules such as proteins and lipids is one reason for its cytotoxicity during harsh environmental conditions. Further, analyses of the specific 1O2 reaction products 10-HOTE and 15-HOTE revealed that it is also the major ROS responsible for lipid peroxidation in plants exposed to simultaneous high-light and cold conditions (152). Thus, large amounts of 1O2 produced during strong photo-oxidative stress conditions can cause an irreversible damage to cellular components and cause cytotoxic effects (116).
Induction of PCD responses
Singlet oxygen can also induce cell death in plants at lower, noncytotoxic levels by inducing a genetically PCD response. This cellular response has been extensively studied by Apel and coworkers taking advantage of the Arabidopsis flu mutant [for recent reviews see (70, 103, 151)]. Strong generation of 1O2 in flu after a dark-to-light shift causes growth arrest of mature plants, bleaching and cell death in seedlings accompanied by the accumulation of enzymatically formed lipid peroxidation products, and the induction of 1O2-specific changes in gene expression (109). A similar cell death phenotype for seedlings has also been observed for a mutant in the barley flu-orthologous called tigrina-d.12 and for another Arabidopsis mutant called oep16. This oep16 mutant is deficient in the import of the NADPH:protochlorophyllide oxidoreductase A involved in the protochlorophyllide conversion during Chl synthesis what leads to the accumulation of this photoactive Chl precursor (112, 156). In the flu system, the isolation of two suppressor mutants of the cell death and growth arrest phenotype, executer 1 (ex1) and executer 2 (ex2), confirmed a signaling role of 1O2 in this genetically controlled cellular response (85, 116, 157). Detailed analyses revealed that the transcriptional response of A. thaliana to increased 1O2 formation is distinct from the response to other ROS such as O2 •− and H2O2, and that H2O2 even antagonizes the 1O2 response and induction of PCD (83, 109). On the other hand, several genes associated with the biosynthesis of or signal transduction through the phytohormones ethylene (ET), salicylic acid (SA), 12-oxo phytodienoic acid (OPDA), and jasmonic acid (JA) were upregulated by 1O2 produced in the flu system or by exogenous photosensitizers such as rose bengal (Fig. 7) (23, 91). This includes the genes of the 1-aminocyclopropane-1-carboxylic acid synthase (ACS) and of several ET-responsive element-binding factors (ERFs) involved in ET signaling, the enhanced disease susceptibility gene EDS1 required for the accumulation of SA, and the genes for lipoxigenase (LOX3), allene oxide synthase (AOS) or cyclase (AOC3), and OPDA reductase (OPR3) involved in OPDA and/or JA synthesis, respectively. Thus, increased synthesis of ET, SA, and JA might be involved in the PCD response after increased 1O2 formation.
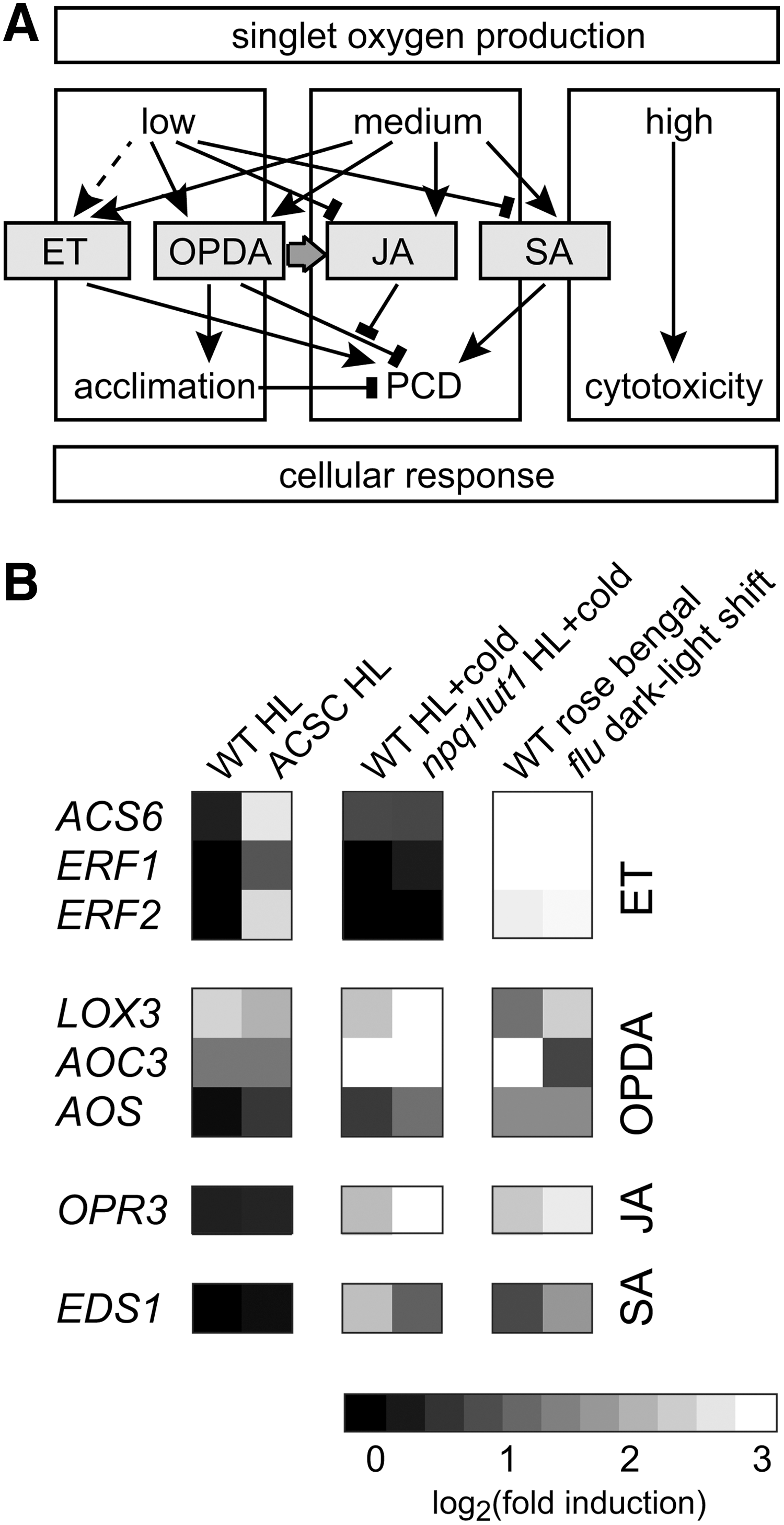
Further analyses confirmed a link between these phytohormones and 1O2-induced PCD response in A. thaliana (Fig. 7). Cell death in the flu mutant after the dark-to-light shift depended on a functional synthesis of ET and SA, both shown to be involved in various physiological responses, including pathogen response (23). SA plays a role in defense against biotrophic pathogens. ET is, together with JA, usually associated with defense against necrotrophic pathogens and herbivorous insects and is mutually antagonistic with the SA response (6). Interestingly, JA was also found to promote 1O2-mediated cell death in flu. Further, the levels of JA as well as of its precursor OPDA and related oxylipins such as 13-HOTE and 13-HODE strongly increase after increased 1O2 formation (23, 116). On the other hand, mutants deficient in JA and OPDA synthesis revealed that these hormones are not essential for the PCD response, showing that they do not function directly as second messengers for 1O2 signaling. An opr3 mutant, disable to convert OPDA to JA, showed a much lower 1O2 response in the flu background compared to the wild-type allele, which could be partially complemented by the addition of exogenous JA (23). This indicates that not JA itself but rather the ratio of JA and OPDA influences the PCD response. Thus, increased OPDA levels might exhibit an antagonistic effect on the 1O2-induced PCD response, which seems to be controlled by the level of JA (Fig. 7). This idea is supported by the abolished induction of OPR3 under conditions producing lower levels of 1O2 compared to flu and rose bengal such as the exposure of wild-type plants or cell cultures to high-light conditions, whereas the genes for OPDA synthesis (LOX3 and AOC3) are still upregulated.
Beside the induction of a PCD response, 1O2 was also shown to function as a signal for other cellular responses. An 1O2-induced transcriptional response in immature Arabidopsis seeds before the onset of dormancy was shown to affect plastid differentiation later during seed germination (69). This long-term effect of 1O2 signaling depended on the recruitment of another plant hormone, abscisic acid. The role of this pregermination 1O2 response in plastid development is unknown. However, mutants disturbing plastid homeostasis were found to have constitutive high expression of 1O2-specific stress-response genes, suggesting a linkage between the chloroplast function and 1O2 response (138). It was argued that the disturbance of the chloroplast homeostasis induces a molecular stress response that stimulates acclimation to the environmental stressors. This acclimation response suppressed the normal 1O2-induced PCD response in the flu background and enhanced the pathogen resistance (138). Similar phenotypes were found for a second class of suppressor mutants of the 1O2-induced bleaching in flu seedlings, called singlet-oxygen-linked death activator (soldat), and for happy-on-norflurazon (hon) mutants. The hon mutants are resistant to the bleaching herbicide norflurazon, which blocks carotenoid synthesis. In hon, norflurazon treatment still increased 1O2 accumulation in the light, but the expression of the nuclear Lhcb gene, normally downregulated by the herbicide, was derepressed (127). It was shown that resistance to norflurazon was due to the induction of an acclimation response caused by a disturbed plastid protein homeostasis in the hon mutants. Similarly, deletion of the SIGMA6 factor of plastid RNA polymerase in the soldat8 mutant and of a gene coding for a putative transcription termination factor in the soldat10 mutant disturbed plastid protein synthesis and stimulated acclimation at the beginning of the seedling development. This acclimation process suppressed the 1O2-induced PCD response in the flu mutant (19, 96). All these examples show that acclimation to a disturbed plastid homeostasis can suppress the normal cellular response to increased 1O2 levels.
Induction of acclimation responses
Acclimation to stressful environmental conditions usually is stimulated by pre-exposure to low levels of the stressor that changes the expression levels of responsive genes helping to protect the organism from the harmful effects of high stress levels. Such an acclimation response to 1O2 has been found in the green alga C. reinhardtii. In this alga pre-exposure to low concentrations of exogenous photosensitizers such as rose bengal or neutral red or to high-light illumination can increase the tolerance to normally toxic levels of the 1O2-producing chemicals (35, 84). This increased tolerance could be linked to the stimulated expression of nuclear defense genes, including the glutathione peroxidase homologous gene GPXH/GPX5 and the glutathione S-transferase gene GSTS1. Overexpression of either of these genes was sufficient to increase the tolerance to 1O2-producing chemicals (84). Both genes were also induced in C. reinhardtii exposed to high-light illumination (41). This indicates that the same response as induced by the exogenous photosensitizers might stimulate acclimation to 1O2 under natural photo-oxidative stress conditions.
Even though no increased tolerance to photo-oxidative stress was shown yet for plants after acclimation to low 1O2 levels, changes in the gene expression profiles associated with an acclimation process have also been detected in A. thaliana. The double-mutant npq1lut2 is deficient in the two 1O2-scavenging xanthophylls lutein and zeaxanthin. This enables the plant to induce an 1O2-specific signal upon a high light shift in the cold, which is weaker than in the flu system (2). As a result, many genes encoding for chloroplast proteins were differently expressed in the mutant compared to wild-type plants. It also increased the expression and synthesis of carotenoids, tocopherol, and other antioxidants and caused a rearrangement of the photosynthetic apparatus (2). Thus, it was claimed that in this mutant 1O2 induced an acclimation response. However, in another study, similar stress treatments of wild-type A. thaliana were able to induce a PCD response after prolonged exposure (96). Thus, in agreement with a rather strong 1O2 response, a high induction of EDS1, AOC3, and OPR3 was detected in npq1lut2 exposed to photo-oxidative stress conditions (Fig. 7B). On the other hand, genes involved in ET response (ERFs and ACS) were not upregulated under these conditions what might explain the missing PCD response. Alternatively, ET response and stimulation of PCD might be delayed under the slightly weaker 1O2 stress conditions than in the flu system.
In Arabidopsis cell suspension cultures (ACSC) exposed to high-light conditions, the 1O2-specific transcriptional response was also similar to the one in the flu system, but no PCD response was detected (48). Thus, under these conditions, lower 1O2 levels might similar to npq1lut2 induce an acclimation response. However, different to npq1lut2, in ACSC exposed to high-light conditions, genes involved in ET response were strongly induced, as were genes of OPDA and/or JA synthesis (LOX3 and AOC3) (Fig. 7B). On the other hand, no evidence for increased SA synthesis such as the induction of EDS1 could be detected in ACSC, supporting an essential role of SA synthesis in PCD response (48). Further, transcripts encoding peroxisome-located transporters for OPDA or enzymes for the synthesis of JA from OPDA such as OPR3 were also not upregulated. This suggests that OPDA, but not JA, synthesis was induced. In agreement with that, a much higher correlation of the high-light response in ACSC with the response of A. thaliana to OPDA than to JA was found (50). OPDA was shown to trigger a transcriptional response different from JA, but more similar to wounding, including the specific induction of many stress response and detoxification genes (144). This indicates that OPDA rather than JA might trigger the acclimation response to low 1O2 levels. Together with the fact that the ratio of JA and OPDA affects the induction of PCD, these findings suggest that the rates of 1O2 production determine the relative levels of OPDA and JA, which might play a key role in the decision between acclimation and PCD responses (50).
Activation of Signal Cascades by Singlet Oxygen
In photosynthetic organisms, 1O2 is produced in the plastid as described above, but it activates the expression of many genes in the nucleus. The induction of nuclear gene expression by plastid signals is called retrograde signaling, and several components of the chloroplast-to-nucleus signal transduction pathways have been identified [for reviews, see (18, 33, 46, 89, 111)]. Singlet oxygen is one of the plastid signals shown to activate nuclear gene expression; however, due to its high reactivity, 1O2 is unlikely to be the signaling molecule that transmits the signal from the chloroplast to the cytoplasm. Still, it has been shown in C. reinhardtii that 1O2 generated in PSII can be detected in the cytoplasm (39). In that study, it was not possible to distinguish whether 1O2 diffuses from its site of production to the cytoplasm or is produced by secondary reactions of LOOH at the outer membrane of the chloroplast envelope (Russell mechanism). However, it is most likely that the 1O2 signal has to be detected by a specific sensor in the thylakoids or the stroma of the chloroplast, and that this sensor than activates a signal transduction pathway directed to the nucleus (39). Several studies were conducted attempting to identify the components of the 1O2 signaling pathways, most of them applying screens to identify mutants affected in the physiological or molecular response to the signal.
Components involved in singlet-oxygen signaling
In A. thaliana, the two plastid-localized proteins EX1 and EX2 have been identified in a mutant screen for plant deficient in the 1O2-induced PCD response (157). The exact function of these two homologous proteins is not known, but they play a crucial role in the 1O2-mediated changes in gene expression and stimulation of phytohormone synthesis in the flu mutant as shown by the loss of 1O2 response in the ex1 single or ex1/ex2 double mutant (Fig. 8) (85, 116). Another approach was used by screening for mutants deficient in the 1O2-specific transcriptional response. The promoter of an AAA-ATPase gene was used for reporter gene construction and screening for constitutive activator of AAA-ATPase (caa) or non-activator of AAA-ATPase (naa) mutants (9). Characterization of these mutants revealed that there is probably not a single linear signaling pathway for the response of this gene that could be easily mutated. In agreement with that, identification of the mutant caa13, containing a mutation in the pleiotropic response locus 1 (PRL1) gene, suggested that 1O2 signaling is strongly interconnected with other signaling pathways by the function of the PRL1 protein (10). Similarly, other 1O2-response mutants (caa33, soldat8, and soldat10) are depleted in processes generally affecting plastid homeostasis and consequently stimulating 1O2 response and acclimation (19, 96, 138). All these examples indicate that the 1O2 signal transduction in plants forms an integral part of a complex signaling network that is modified by other signaling routes (9).

1O2 signaling does not only change gene expression on the transcriptional level, but was also shown to affect translation (120). A reduced expression of many chloroplast proteins as a response to 1O2-producing conditions was detected in the barley flu-orthologous mutant tigrina-d.12 (68). This suppression of protein synthesis was caused by an arrest in translation initiation at the 80S cytoplasmic ribosomes what correlated with a declined phosphorylation level of the ribosomal protein S6. A strong effect of 1O2 production on translation was also observed in the A. thaliana oep16 mutant deficient in the protochlorophyllide conversion. Even though upon the dark-to-light shift oep16 exhibits a similar PCD phenotype to flu, translation was differently affected in the two mutants. This shows that again multiple signaling mechanisms might control translational by 1O2 production (129).
The green alga C. reinhardtii provides another powerful tool to screen for 1O2-response mutants, because its haploid genome and asexual reproduction allow the fast identification of loss-of-function mutations. Further, several 1O2-specific responses have been identified in C. reinhardtii, including acclimation and induction of the defense genes (84, 86). The HSP70A gene was shown to be induced by 1O2 and different HSP70A-based reporter constructs could be created which specifically responded to either 1O2 or H2O2 (135). The GPXH/GPX5 gene is strongly induced by 1O2, but only moderately by other ROS, and it was shown to be specifically induced by 1O2 during exposure to high-light conditions (37, 38). Further, the transcriptional activation of GPXH is dependent on multiple regulatory elements that are distinct from an RES-responsive element (34, 40). A reporter construct containing an 1O2-specific GPXH promoter fragment was used to screen more than 5,000 UV-mutagenized clones for their response to 1O2. The fact that no mutants deficient in signal transduction could be isolated indicates that in C. reinhardtii, similar to plants, the 1O2 signal is not transmitted through a single linear pathway (36). On the other hand, an independent screen for mutants deficient in the response of a GPXH-dependent reporter construct resulted in the isolation of a psbP2 mutant (15). The PSBP2 gene is paralogous to the oxygen-evolving enhancer gene PSBP1, and it was hypothesized that it might function as a sensor for 1O2 to activate GPXH expression. However, induction of the wild-type GPXH gene and acclimation to 1O2 are not depleted completely in psbP2, supporting the fact that multiple signaling pathways might be involved in a specific 1O2 response in C. reinhardtii (Fig. 8).
Reactive electrophiles as putative transmitters of singlet-oxygen signals
An alternative signaling pathway in the response of C. reinhardtii to 1O2 was recently identified in a screen for mutants that are constitutively acclimated to 1O2 and thus more tolerant to high concentrations of 1O2-generating photosensitizers (40). One of these singlet oxygen-resistant (sor) mutants was affected by a transcription factor called SOR1. SOR1 seems to regulate the expression of a number oxidative stress response and detoxification genes, including GPXH and GSTS1. Most of the SOR1-regulated genes are also strongly induced by lipophilic RES and contain a RES-response element (Fig. 8). This indicates a link between the response to RES and the acclimation to 1O2. Such a link was confirmed by the increased tolerance of RES-acclimated C. reinhardtii cells to subsequent exposure to toxic 1O2 levels (40).
RES can be formed after increased 1O2 formation by direct oxidation of PUFA (see “Main targets of singlet oxygen damage”) or enzymatically by the activation of lipid-oxidizing enzymes called LOXs. In A. thaliana, 1O2 was shown to activate enzymatic lipid oxidation by inducing the expression of LOX2 and LOX3 coding for two 13-LOXs catalyzing the synthesis of 13-HPOTE (Fig. 8). This hydroperoxide is a precursor for OPDA synthesis but can also be reduced to 13-HOTE or converted to 13-KOTE. Due to their α/β-unsaturated keto group, 13-KOTEs participate in nucleophilic Michael additions and therefore are RES (101). Similarly, OPDA also belongs to the RES, whereas JA is a saturated ketone and not as electrophilic. Different RES have been shown to activate similar response mechanisms causing comparable gene expression profiles that were also related to the response to other stressors such as pathogens and wounding (3, 101, 144). Exogenous RES can even stimulate the accumulation of OPDA without affecting the JA levels (3). Thus, the much higher overlap of low 1O2-induced genes with the OPDA- than with JA-responsive genes indicates that many genes induced during 1O2 acclimation are responding to reactive electrophilic oxylipins such as 13-KOTE and OPDA (50). Direct requirement of OPDA accumulation was shown for the 1O2 response of the CYP94C and VSP1 genes in the OPDA synthesis mutant aos (116). On the other hand, other OPDA-responsive genes such as OPR1 and HSP17.6 were derepressed in aos, what might be due to released repression by JA, which is also depleted in aos mutants. Finally, a lox1lox5 mutant deficient in 9-HPOTE synthesis was more sensitive to the 1O2 photosensitizer rose bengal. This supports an important role of reactive electrophilic oxylipins such as OPDA, HPOTE, and KOTE in 1O2 signaling and acclimation in photosynthetic organisms (91).
Another group of RES that can be formed by 1O2 are carotenoid oxidation products (118). Carotenoids are the main 1O2 quenchers in the photosynthetic apparatus, and therefore increased levels of the carotenoid oxidation products β-cyclocitral and β-ionone could be measured in A. thaliana exposed to a high light intensity in the cold (119). Exposure to β-cyclocitral induced a very similar response pattern to the flu mutant. It stimulated the expression of many genes specifically induced by 1O2 or 1O2 and RES such as HSP17.6, GST1, and GST6 (Fig. 8). Further, β-cyclocitral also induced the expression of genes for OPDA synthesis (LOX2, AOC1 and AOS), indicating an interaction with oxylipin signaling and the induction of RES responses. The induction of genes by β-cyclocitral increased the tolerance to high light and cold treatment (119). Because the responses to β-cyclocitral were EX1 independent, it was suggested that this carotenoid oxidation product might function as second messenger to stimulate the 1O2-induced acclimation response in A. thaliana. Induction experiments with β-cyclocitral in C. reinhardtii revealed no transcriptional response similar to the specific 1O2 induction, but rather a stimulated expression of detoxification mechanisms similar to other RES treatments (Fischer BB, unpublished). Thus, β-cyclocitral might not be an universal signaling molecule transmitting the 1O2 signal to the nucleus in photosynthetic organisms. However, it might be a part of a larger group of RES molecules that individually or in combination stimulate part of the 1O2-specific response and activate acclimation to photo-oxidative stress conditions (Fig. 8).
Conclusions and Outlook
Despite the increasing body of knowledge in the literature on 1O2 generation and signaling, many important questions remain open for future research. The mechanism of safe transport and storage of Chl during degradation of CP to prevent 1O2 formation is still unidentified. Even though first lines of evidence are provided on how Chl is inserted into the LHC by LIL proteins, it is still unclear how Chl is inserted into the reaction centers of the PSs. Further, the fate of Chl during the turnover of the D1 protein is not known. Proteins of the ELIP family may play a role in this process, and the water-soluble CP may be involved in the protection of old leaves against destruction through 1O2 produced by free Chl; however, the exact protection mechanisms still need to be solved.
Beside its deleterious effects, 1O2 plays an important function in cell signaling by activating a cascade to transmit a chloroplast signal to the nucleus and stimulate a specific gene expression response. However, hardly anything is known about how and where the 1O2 signal is sensed and transmitted to the cytoplasm. Such specific questions also increase the need to establish reliable 1O2 detection methods that allow to detect 1O2 quantitatively and selectively, to localize it inside the cell compartments, and to determine its lifetime in Chl-containing organisms. Studies on 1O2 signaling also revealed that different and multiple mechanisms might be involved in 1O2 response in different photosynthetic organisms, and that strong interactions with other ROS and RES-signaling cascades occur. Unraveling these different mechanisms will help to clarify the specific role of 1O2 in the response to photo-oxidative stress and in the response to other stressors such as pathogen infection. By knowing all the factors, their interaction and sensitivity to abiotic and biotic stressors, we might start to understand under which conditions plants respond to stressors by activating an acclimation or a PCD response.
Footnotes
Acknowledgments
We would like to thank Rik Eggen (Eawag) for critical reading of the manuscript and Roberto Bassi (University Verona) for providing detailed information on the carotenoid composition of the different protein subunits. A. K.L. was supported by the Agence Nationale de Recherche reference ANR-09-BLAN-0005-01. É. H. acknowledges support from the Hungarian Scientific Research Fund (OTKA K 101430).
Author Disclosure Statement
No competing financial interests exist.