
Abstract
Introduction
ROS (1O2, O2
The Plasma Membrane as Stress Target
The cell membrane (plasma membrane, PM) is one of the first targets confronted with biotic or abiotic factors that trigger oxidative stress. Besides signal transduction and stress response, the PM is responsible for nutrient uptake, transport and cell wall related processes. Involvement of PM redox systems in these processes becomes more and more established (78, 91, 104, 152, 177).
Both in vivo and in vitro studies demonstrated the existence of different redox systems in plant PM (25, 106). Application of artificial electron acceptors like ferricyanide (hexacyanoferrate (III), HCF III) to intact plant roots caused (i) a depolarization of the PM, (ii) a reduction of HCF III outside the cell (Fig. 1), and (iii) a simultaneously acidification of the medium. These observations revealed different hypothetical models that postulated a constitutive transmembrane electron transport from a cytosolic electron donor (NAD(P)H or ascorbate, ASC) to an extracellular or intracellular electron acceptor (e.g., iron or oxygen) and a stimulation of the PM H+-ATPase.

The transmembrane HCF III reductase activity appears to be due to a constitutive and ubiquitous existing system. In vivo experiments with maize (Zea mays L.) roots suggest that this so-called standard system may be oxygen sensitive, stimulated by naphthoquinones (vitamin K-type) and inhibited by coumarins (vitamin K antagonists) and auxins (104). A comparison of in vitro studies demonstrated that NADPH appears to play a major role in the transmembrane electron transfer to HCF III in maize roots (118), whereas ASC-loaded or NADH-loaded PM vesicles of other plant tissues and species showed significant lower electron transfer rates (9, 14, 36). Additionally, absorption spectra of PM vesicles revealed reduction of HCF III via a b-type cytochrome (Cyt) in all tissues and species (9, 12, 15, 109).
Iron-deficiency caused an increase in the transmembrane ferric-chelate reductase activity named the turbo reductase or the turbo system (152). The turbo system was not found in grasses, which supports the hypothesis that the standard and turbo reductase are two distinct redox systems (29, 106). It was further shown that grasses evolved an iron uptake strategy related to prokaryotes (Fig. 2), whereas all other plant families revealed an iron uptake strategy comparable to yeast (78, 94, 170). A recent study of our team showed significant higher rates of transmembrane ferric-chelate reduction for sealed and NADH-loaded right-side-out PM vesicles compared to NADPH-loaded vesicles of iron-deficient pea (Pisum sativum L.) roots (Meisrimler and Lüthje, unpublished observation). Thus, NAD(P)H may be suggested as a natural electron donor for the standard system and NADH as an electron donor for the turbo reductase.

Although electron transport processes at the PM were discovered more than 30 years ago and several redox compounds have been identified (Table 1), electron transfer mechanisms and physiological functions of these systems are not very well understood. A lack in full-length amino acid sequence identification of biochemically characterized PM proteins hampers their functional analysis (21, 42, 74, 109, 157, 183, 185). In contrary, most redox systems identified in plant PM preparations by proteomic approaches have not been further characterized (34, 75, 97, 115, 117, 125).
Accession numbers were given according to UNIPROT (
NCBI record removed from database. This whole-genome-shotgun sequence record was removed because it has been superceded by a new assembly of the genome
Purification and identification of transmembrane proteins by mass spectrometry are still a major challenge. Up to now those proteins are under-represented in biochemical and proteomic studies. Nevertheless, proteomic approaches demonstrated that the PM presents one of the most complex subproteomes in plant cells, which may be organized in microdomains (149, 163). The identification of redox systems in detergent insoluble membranes supports the idea that redox systems are localized in those microdomains and may have important functions in signal transduction, stress responses, cell wall metabolism, and transport processes (104, 108). Besides other proteins, b-type cytochromes, members of the flavodoxin and flavodoxin-like protein families, copper-binding proteins, monodehydroascorbate (MDHA) reductase, and malate dehydrogenase (MDH) have been identified in PM preparations (34, 75, 97, 115, 117, 125, 132, 172). Now critical evaluation of the data will be necessary to discriminate between cytosolic or endomembrane contaminants and real PM constituents (20, 107). Most of the redox proteins identified are peripheral and only some of these appear to have transmembrane domains or modifications that allow a direct interaction with the PM. Thus, artefacts and protein–protein interactions have to be carefully elucidated (83, 107, 183). Contamination of PM preparations by ER and cell wall compounds appear the most critical because of the PM interaction with these compartments (95).
Molecular biological approaches revealed several homologous genes of NOX2 (NADPH oxidase; gp91phox, for glycosylated protein MWr 91 kDa phagocyte oxidase), the α-subunit of the neutrophil NADPH oxidase and founding member of the flavocytochrome b family, in animal, yeast, algae, and plants (88, 146, 162, 177, 191). In analogy to the neutrophil system, the PM localization has been postulated for these gene products in the plants discussed below.
PM Redox Systems
The Cyt composition of the PM has been characterized for several plant species by the measurement of absorbance spectra and redox titrations (10). The observation of split α-bands, the identification of low midpoint potentials (E0′=−20 to –40 mV), and the relatively high antimycin A-insensitive NADH-Cytc reductase activity found with some PM preparations were taken as the indirect evidence for the presence of Cytb 5 (12, 15). Cytb 5 identified by proteomic approaches, however, was discussed as contamination by the endoplasmic reticulum (132).
The major Cyt of plant PM appears to be a high-potential Cytb561 (E0′≈+150 mV), which is present in all plant materials investigated with the apparent exception of maize coleoptiles (10). A low potential haem comparable to the neutrophil Cytb558 (E0′≈−245 mV) could not be detected in plant PM so far (10, 15, 128).
Cytb561 protein family
In vitro studies suggested the possible involvement of an ASC-reducible Cytb561 in the transmembrane electron transfer from cytosolic ASC to apoplastic HCF III (9, 76). Later on an ASC-reducible Cytb561 with transmembrane spanning topology was detected in PM preparations of bean (Phaseolus vulgaris L.) and Arabidopsis thaliana (L.)
Cytb561
The bean Cytb561 has a relative molecular mass of 27 kDa in SDS–PAGE and was found as a dimer (60 kDa) by size-exclusion chromatography (141, 181). Peptides identified for PhvCytb561 were conserved in AtCytb561-4 (CYBASC1; Cytb561D; ascorbate ferrireductase 4), OsCytb561, and ZmCytb561-1. Arabidopsis and maize proteins were cloned and biochemically characterized after heterologous expression (26, 129, 130, 184). Properties of ZmCytb561-1 were comparable to that of bean and arabidopsis homologs (26, 141). The proteins have six conserved transmembrane α-helices and two haem ligation sites (18). Midpoint potentials of haem groups were slightly lower for recombinant ZmCytb561-1 (+123 and +15 mV) compared to recombinant AtCytb561-4 (+165 mV and +57 mV) (26, 130). Both amino acid sequences contain conserved binding-motifs for ASC and MDHA, which were used as substrates of the recombinant proteins. A concerted transport of electrons and protons was demonstrated for ZmCytb561-1 (130). While localization of ZmCytb561-1 needs further studies, PM detection of PhvCytb561 and AtCytb561-4 appeared to be due to a tonoplast contamination (63, 141). A function in iron reduction has been demonstrated for the recombinant AtCytb561-4 (26).
At least in barley (Hordeum vulgare L.) and wild watermelon (Citrullus lanatus L.), a PM localization of a Cytb561 has been demonstrated by fluorescence protein-fusion constructs (28, 131). The barley HvCytb561 colocalised with synthaxin at the side of pathogen invasion suggesting a function in pathogen response (28). Both CLb561A and CLb561B of the wild watermelon showed significant increases in expression levels and protein abundance during drought stress and high light conditions (131). The induction was correlated to the occurrence of excess reducing equivalents under these stresses and a simultaneous increase of apoplastic ASC oxidase. The authors assumed that excess of reducing equivalents may be transported from the chloroplast via the cytosol to the apoplast by malate-oxaloacetate (OAA) shuttle, cytosolic Asc–glutathione cycle, and accelerated Asc regeneration in the apoplast triggered by CLb561A induction. Finally, ASC may be used by apoplastic ASC oxidase for reduction of oxygen to water. Thus, PM-bound Cytb561 appears to regulate the apoplastic redox state (11) and may have a function for excess light energy dissipation (131).
DoHCytb561
Shot-gun proteomics revealed DoHCytb561 homologs in barrel clover (Medicago truncatula
Dopamine-β-hydroxylase
Proteomic approaches detected auxin inducible in root cultures (AIR12) in the PM and detergent insoluble membranes of arabidopsis, Medicago truncatula, and maize, which belongs to the DOMON domain superfamily (34, 75, 97). AIR12 was identified as the major ASC-reducible compound in the PM of bean and soybean (Glycine max L.) hypocotyls (142, 181). PhvAIR12 has a native molecular mass of 55 to 63 kDa, is glycosylated and most likely attached to the apoplasmic surface of the PM by a glycosylphosphatidylinositol-(GPI)-anchor (33, 181). The recombinant protein (65 to 90 kDa, 27 kDa after deglycosylation) showed a symmetrical α-band at 561 nm, caused by a single, highly axial low-spin haem that could be fully reduced by ASC and fully oxidized by MDHA (142). This observation was in agreement with conserved binding motifs for these substrates in the amino acid sequence of AIR12 (184). Additionally, phylogenetic analyses reveal that the auxin-responsive genes AIR12 represent a new protein family specific to flowering plants (142).
Accumulation of AIR12 mRNA starts 4 to 8 h after application of auxins and a function of the gene product in lateral root formation was postulated (135). The induction of AIR12 by auxin also suggests that the protein could be another mediator of catecholamine function in plants (93, 187). The higher abundance of AIR12 in maize roots after long-term treatment with high concentrations of iron suggests an additional function in oxidative stress (75).
Flavocytochrome b family
Plant homologs of the neutrophil NADPH oxidase (NOX2) can be classified as ferric-chelate reductase (FRO, ferric reduction oxidase) and respiratory burst oxidase homologs (Rboh) (87, 89, 146, 162, 191). Flavocytochromes b especially Rboh are most intensively investigated among PM-bound redox systems. Several reviews are published on the structure, regulation, physiological functions, and evolution of this protein family and are recommended for further details (88, 113, 169, 171). Functions of Rboh in the pathogen response and symbiosis are discussed elsewhere (58, 114, 178).
Turbo system
The turbo reductase was identified by molecular biological approaches as a member of the flavocytochrome b family (88, 99, 146, 191). Based on the sequence similarity to AtFRO2 (ferric reductase oxidase), the major ferric-chelate reductase in arabidopsis, eight genes have been identified in the genome of this plant species (193). Cloning and heterologous expression of these genes in yeast revealed the ferric-chelate reductase activity for AtFRO2, AtFRO3, AtFRO4, AtFRO5, AtFRO7, and AtFRO8. As summarized in Figure 3, AtFRO2 and AtFRO3 were mainly expressed in roots, AtFRO5 and AtFRO6 in shoots and flowers, and AtFRO7 in cotyledons and trichomes, whereas the transcription of AtFRO8 was specific for leaf veins (193). It was further shown that FRO7 plays an essential role in iron delivery to chloroplasts, whereas FRO3 and FRO8 are localized in the mitochondria membrane and are most likely involved in iron homeostasis of these organelles (80).
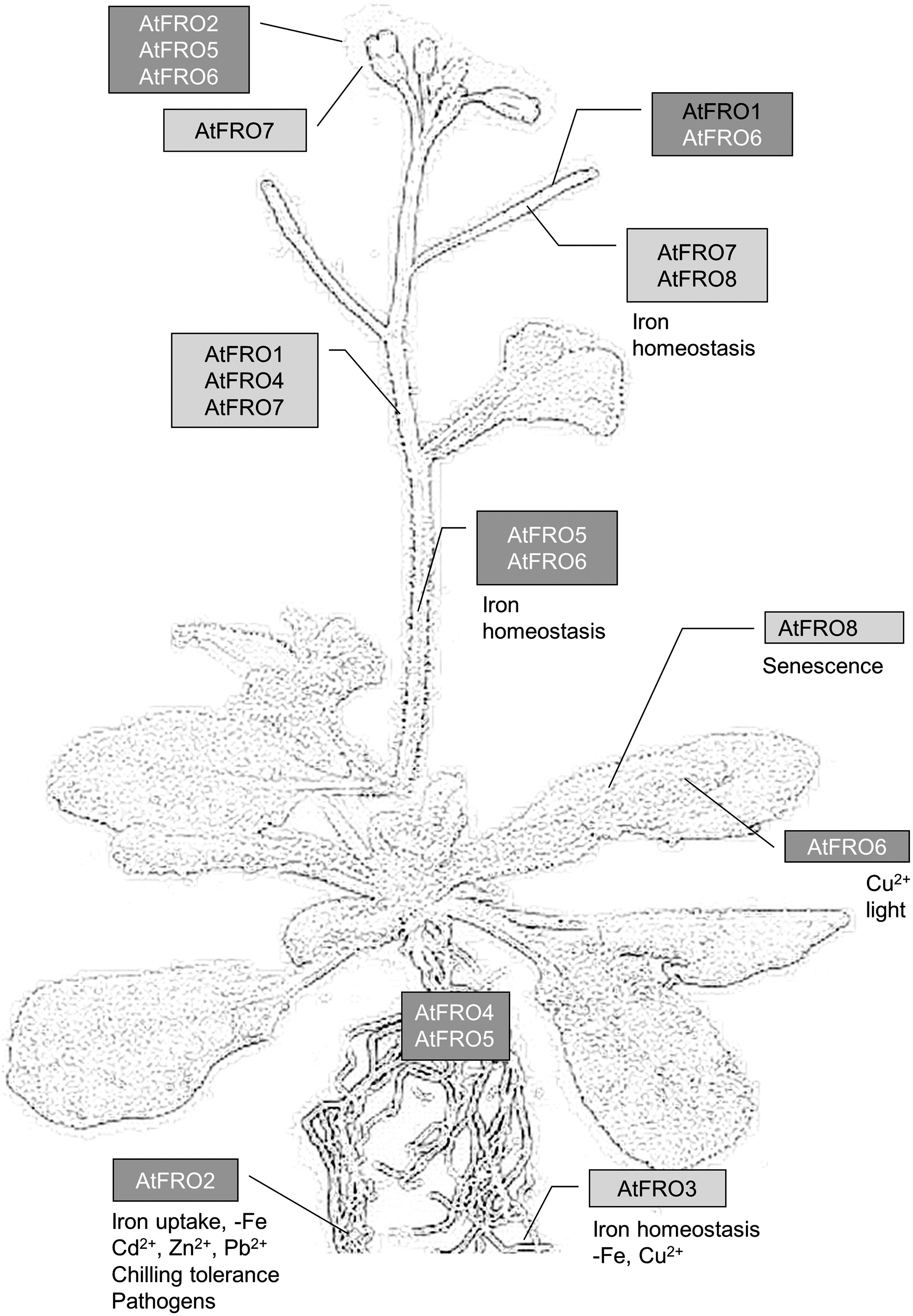
In silico analysis of FRO amino acid sequences predicted eight to 12 transmembrane domains, haem, flavin, and NAD(P)H-binding sites (99, 146, 191). Experimental data for AtFRO2, however, revealed a topology of eight transmembrane helices (151). Helices III, V, VII, and VIII belong to the conserved and membrane-embedded core domain of the flavocytochrome b family. Haem binding-sites are presented by conserved His residues, which are located in helices V and VII. Additionally, a large water soluble domain with NADPH, FAD, and oxidoreductase sequence motifs is located on the inside of the membrane.
In vitro studies revealed higher rates of the transmembrane ferric-chelate reductase activity with NADH compared to NADPH and a preference for Fe(III)-citrate over Fe(III)-EDTA as a substrate in iron-deficient pea roots (Meisrimler and Lüthje, unpublished observations). The native PsFRO1 showed comparable preferences for coenzymes and substrates after partial purification from the PM. This observation fits to the data on the partially purified ferric-chelate reductase activity of tomato roots (73, 74). PsFRO1 was inhibited by low amounts of the flavoprotein inhibitor diphenylene iodonium chloride (DPI), supporting the presence of a flavin cofactor (Meisrimler and Lüthje, unpublished observations).
The physiological function of FRO in ferric-chelate reduction has been demonstrated on the transcriptional level (99, 146, 191). A coordinated regulation of FRO2 together with the major iron transporter (IRT1) of the high iron uptake system has been found for arabidopsis (41). Localization of both IRT1 (iron transporter) and FRO2 mRNA in epidermal root cells confirmed the proposed role of these proteins in the iron uptake from the soil. IRT1 belongs to the zinc and iron transporter protein (ZIP) family, which is divided in zinc-regulated (ZRT) and iron-regulated (IRT-like protein) transporters (66). The ZIP family is selective for bivalent cations like Fe2+, Zn2+, Mn2+, and Cd2+ (41, 188). In accordance to this property, mRNA levels of IRT1, but also of FRO were upregulated by treatment with zinc and cadmium (41, 60). In contrary to this observation, the ferric-chelate reductase activity of sugar beet was inhibited after pretreatment with cadmium or lead (37). Long-term treatment with cadmium caused symptoms similar to iron deficiency. For example, an increase in the ferric-chelate reductase activity revealed high concentrations of iron in the roots, but iron deficiency in young leaves (60). A comparison of iron deficiency and cadmium treatment revealed that at the physiological and molecular levels, the time-course of the patterns for alterations in Fe-deficiency responses induced by cadmium treatment were different in intensity and timing from those induced by iron deficiency. Due to these observations, it was postulated that cadmium itself may act as a local signal and plays a role in regulating iron deficiency responses (60).
In contrast to the upregulation of AtFRO1 by metal stress and pathogens (summarized in Fig. 3), downregulation of the PsFRO2 transcript and the corresponding enzyme activity was observed in iron-deficient pea roots after elicitor treatment (117). The control of iron homeostasis appears to play a central role in pathogen defense (52). Iron is used by the host as a modulator of the oxidative burst (32). Alterations in plant iron trafficking have been described during the establishment of plant innate immunity (48, 52, 102, 155). Additionally, microbial infection caused competition for iron between the host and the pathogen. Thus, iron pools of the host plant could be significantly depleted by siderophore production of the pathogen. Vice versa innate immune responses of the host may deprive the pathogen of nutritional iron by downregulation of FRO (117, 155).
Respiratory burst oxidase homologs
Expression profiles of several Rboh genes indicated that their gene products are involved in a massive production of ROS (so-called oxidative burst) that occurs during the first phase of pathogen defense (89, 162, 177). Additionally, ROS production and Rboh expression correlated with a function in growth regulation and stress responses (56). Rboh genes belong to a small multigenic family with ten members in arabidopsis and nine in rice, composed of five groups of orthologous sequences (113). At least four Rboh genes with different splicing have been identified in maize (101). Tissue-specific expression patterns were found for Rboh during plant development in a bitter apple (Citrullus colocynthis (L
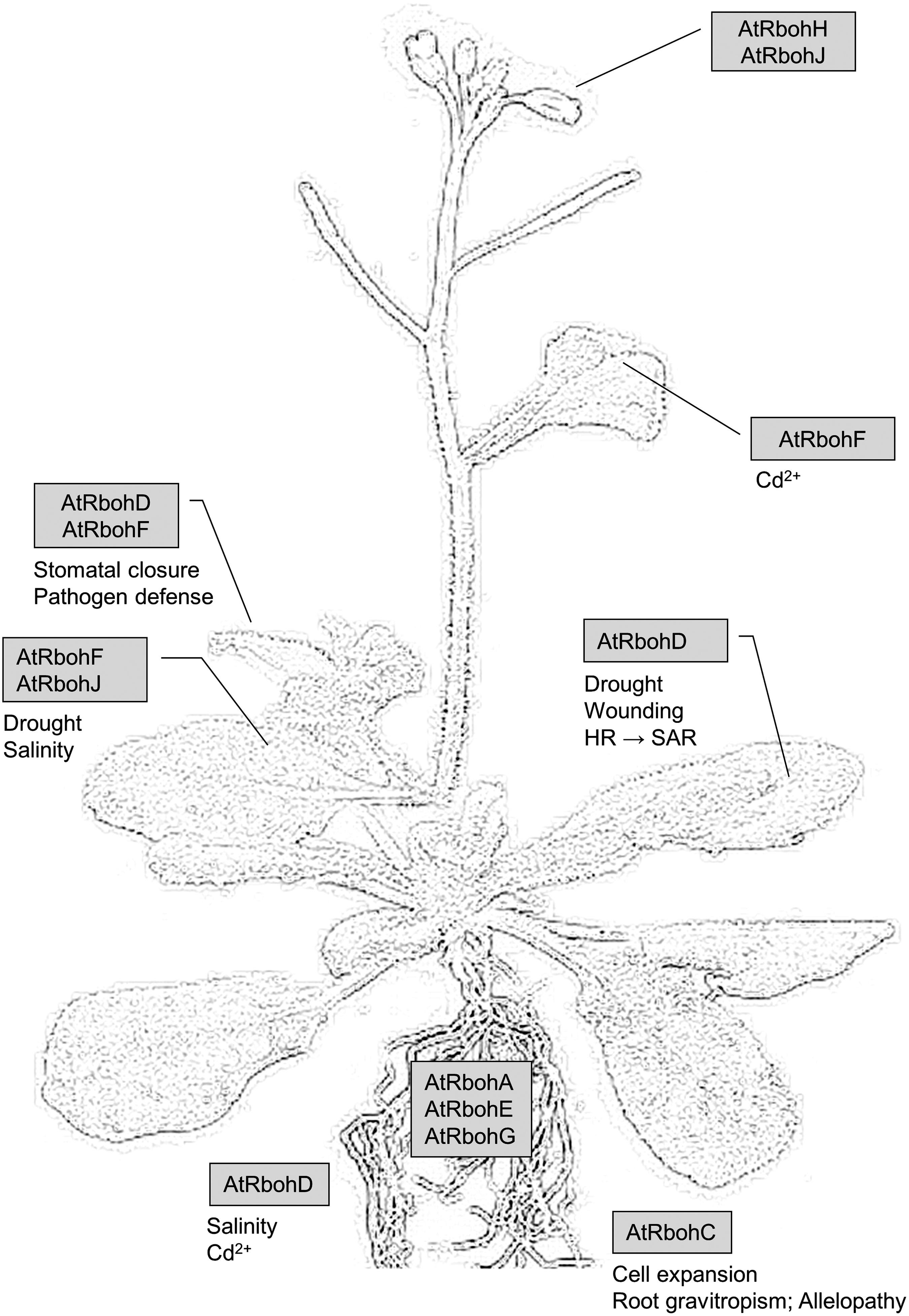
In recent years, the structure and regulation of the whole protein family were intensively reviewed and functions of Rboh in the pathogen response and ROS signaling are discussed in detail elsewhere (19, 113, 169, 171). Expression and functions of Rboh are summarized in Figure 4.
The PM localization of AtRbohA, NtRohD, StRbohA, and StRbohB was demonstrated (87, 89, 162). In silico analysis of Rboh amino acid sequences predicted up to ten transmembrane domains, two haem, FAD and NADPH binding-sites, and conserved EF-hand motifs at the N-terminus (4, 64, 87, 162, 179). The NADPH oxidase-like activity was purified from the PM of bean hooks (185). Enzymatic characteristics of the putative bean Rboh pointed to similarities with NOX2 with respect to inhibitor sensitivities, but some properties were contradictory to in silico predictions for plant Rboh. For example, the partially purified enzyme contained FMN instead of FAD and a b-type Cyt could not be detected. Histidin residues for haem binding were highly conserved in plant Rboh, suggesting an active center comparable to NOX2 (-245 mV). However, redox titration analysis of highly enriched PM preparations revealed that a low potential haem group could not be detected in plants (10, 128). Assuming that the amount of Rboh in plant PM is comparable to that of NOX2 in phagocyte membranes, 15% of the total cytochrome b in plant PM should be formed by this flavocytochrome b. This amount [80 pmol cytochrome b mg1 protein, (43)] should be well above the limits of detection. Experiments with sealed and NADPH-loaded right-side-out PM vesicles of maize roots, however, revealed a transmembrane NADPH oxidase activity (118). Thus, the electron transfer mechanism from cytosolic NADPH to molecular oxygen by Rboh needs further elucidation.
Most regulatory processes of Rboh, found in the last years, influence the transcriptional level. ROS production by NtRbohD appears to be regulated by a protein–protein interaction at the C-terminus (50). Besides Nt14-3-3h/omega, protein phosphatase 2C was identified as an interaction partner. Transcripts of Nt14-3-3h/omega were upregulated after infiltration of tobacco leaves with the fungal elicitor cryptogein. Regulation of ROS accumulation by this protein was verified by tobacco cells transformed with an antisense construct of Nt14-3-3h/omega that showed strong inhibition of ROS accumulation. This regulation mechanism has to be further analyzed. Besides the protein–protein interaction, Rboh is regulated by phosphorylation and calcium binding in its N-terminal cytosolic region (136, 173, 174). The current consensus on activation and inhibition of Rboh involves calcium-dependent protein kinases (CDPK) and Ca2+-signaling: for activation, two conserved serins are phosphorylated by CDPK, administrating the ability to bind to Rac-GTP. ROS production in the apoplast causes an elevated calcium influx, promoting Rac-GTP dissociation from Rboh in favor of Ca2+ binding to two EF-hands (90, 192). Additionally, regulation of Rboh takes place by the surrounding lipids. At least two types of lipid modulators take part: phosphatidic acid (PA) and phosphatidylinositol (PtdIns). When abscisic acid (ABA) signals reach stomatal guard cells during drought stress, phospholipase Dα1 produces PA. The lipid is then bound by four arginin residues in the N-terminal region of AtRbohD. ROS production increases and the stomata are closed to prevent a loss of water (198). When knocking out phosphatase 7 (At5PTase7) that produces PtdIns, arabidopsis probably fails to react properly to salt stress. This effect is similar to a direct inhibition of RbohH (85). Normally, stress activation of AtRbohD would lead to downstream induction of gene expression of AtRbohF and AtRbohJ, the isoforms responsible for drought and salt stress, respectively (Fig. 4). In agreement with these observations, ABA treatment of maize leaves induced a biphasic response (phase I and phase II) in the expression of ZmRbohA to ZmRbohD and in the NADPH oxidase activity (101). Phase I was not affected by pretreatments with mitogen-activated protein kinase (MPKKK) inhibitors or H2O2 scavengers, whereas phase II was blocked. Treatment with H2O2 alone induced phase II, but not phase I, MAPKK inhibitors caused a permanent induction. These data suggest a positive feedback loop involving NADPH oxidase, H2O2, and ZmMPK5 in ABA signaling (101).
Rboh transcripts are upregulated by high salinity (61, 175, 194). Meanwhile, it has been proposed that ROS originating from Rboh supports mRNA stability of the salt overly sensitive 1 (SOS1) gene, a Na+/H+ antiporter in the PM of arabidopsis (197). It is activated in reaction to salinity stress, pumping Na+ out of the cell and an increasing pH in the apoplast. It was also proposed that a higher pH could induce a positive feedback loop with Rboh, since its activity has been reported to be preceded by apoplastic alkalinization (39).
Effects of mechanical damage on ROS homeostasis and calmodulin (CaM) signaling were investigated in arabidopsis (173). Two signal transduction cascades were discovered that were activated by wounding. First, MAPKK3 is phosphorylated and, in turn, phosphorylates MAPK8 twice. Second, CaM binds to Ca2+, and then to another TDY-motif in MAPK8. Thus modulated, the MAPK8 represses the expression of AtRbohD, maintaining ROS homeostasis.
Finally, upregulation of AtRbohF and NtRbohD was demonstrated after exposure to cadmium (46, 61, 77). In Bright Yellow 2 (BY-2) cells, millimolar concentrations of cadmium caused cell death within 2 to 3 h. This process was preceded by two waves of ROS: (i) a transient ROS production by NtRbohD and (ii) by ROS accumulation in the mitochondria. Finally, cell death occurred by a wave of hydroxyl peroxide in the membranes (61). A similar result has been found in arabidopsis, AtRbohF transcripts were doubled within 15 min by micromolar concentrations of cadmium (77).
Peroxidases
At least three haem-containing peroxidases (Prx) have been characterized after partial purification from PM of maize roots (119, 121). These Prx, ZmPrx01, ZmPrx66, and ZmPrx70, share characteristics common for class III Prx (secretory pathway). All proteins have an N-terminal signal peptide directing to the Golgi or PM, possess haem and calcium-binding sites, and are glycosylated. The amino acid sequence of class III Prx is highly conserved. Due to this fact, three-dimensional (3D) structures could be predicted based on Prx with known structures and hypothetical 3D structures have been published for PM-bound Prx (121).
In silico analysis of putative membrane-bound maize Prx revealed six to 21 α-helices and between two to 11 ß-sheets in a conserved orientation. Haem, substrate, and calcium-binding sites could be identified in the amino acid sequences. The calculated MW is 38.3 kDa for ZmPrx01 and 33.4 kDa for ZmPrx66 and ZmPrx70, the theoretical pI is 6.81 for ZmPrx01, 8.39 for ZmPrx70, and 8.94 for ZmPrx66. In other plant species, putative PM-bound Prx has been identified by proteomic studies (Table 1). Some of these isoenzymes have been found in detergent insoluble microdomains of the PM (97, 108, 125).
Due to their ability to perform two independent catalytic cycles, plant Prx are involved in several cellular and extracellular processes. Via a hydroxylic cycle, they can regulate H2O2 levels and release ROS (138) and via a peroxidative cycle, various substrates can be used for oxidation (Fig. 5). The functions of the Prx range from cell wall-related reactions (crosslinking or separation of cell wall components) to basic metabolic pathways and stress-related processes (3, 138, 164).
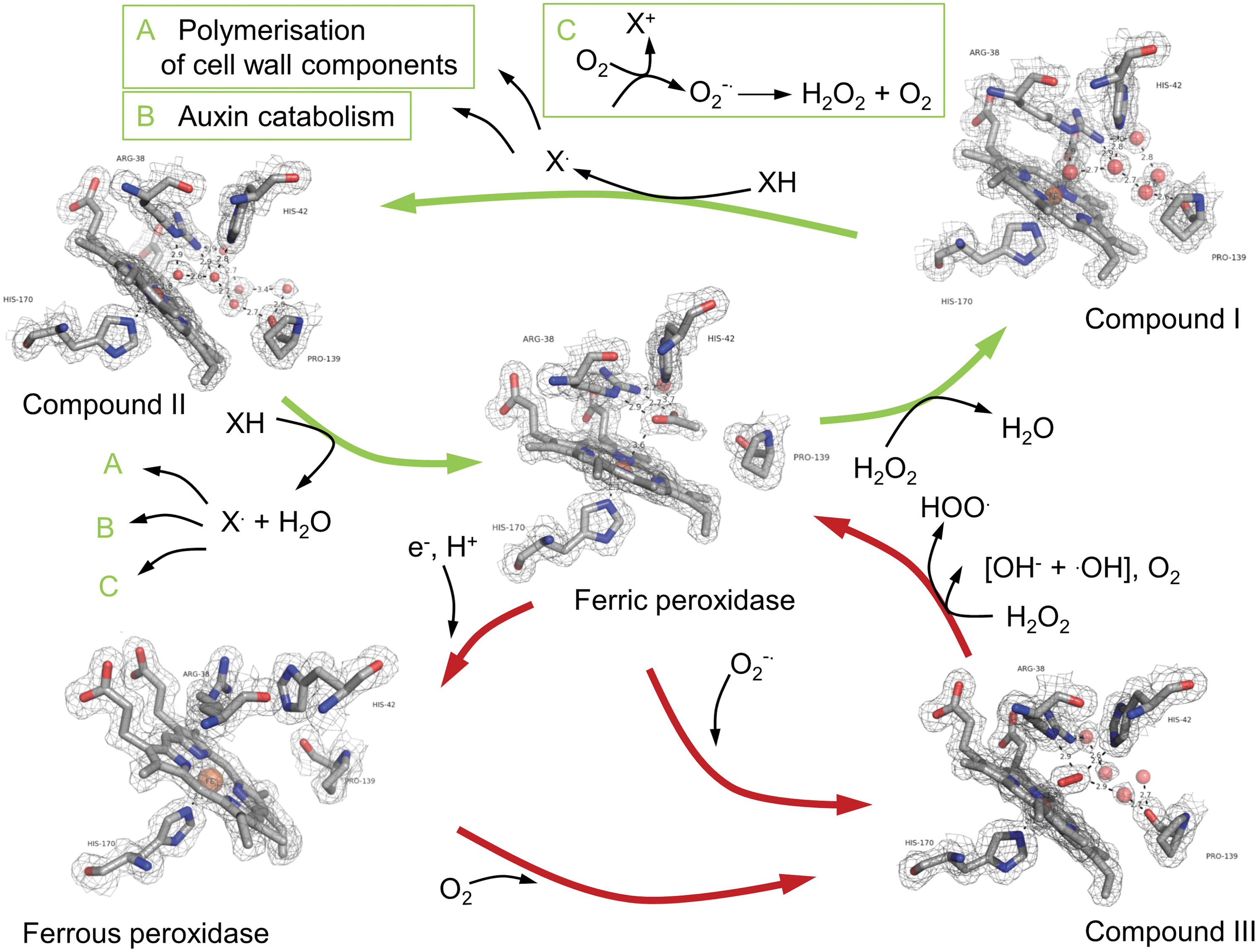
Lipid peroxidation and changes in membrane permeability and stability have been observed by oxidative stress (144, 145). To remain the PM integrity and to ensure the cell function under different stress conditions, PM-localized Prx have been suggested to play an important role in detoxification of the cell and repair of the PM (108). This hypothesis is supported by analysis of the gene structure of ZmPrx70, which revealed several putative cis-regulatory elements that suggested a regulation of its gene by wounding, methyl-jasmonate, salicylic acid, and elicitors (121). The similarity to the family of antioxidant-responsive elements (ARE) in the 5′-untranslated region of the gene is additionally supporting a regulation by oxidative stress (72).
Experimental evidence for this prediction was given by proteomic approaches. The response has been studied of soluble and membrane-bound class III Prx to biotic and abiotic stressors. Besides a comparison of soluble and microsomal fractions, PM-bound Prx isoforms were characterized. The protein levels of four haem Prx in response to stress and signal compounds, were investigated using isoelectric focusing and nonreducing, modified SDS-PAGE (120).
Due to the fact that a binding of apoplastic Prx may be possible under certain stress conditions (32), an extra washing step was integrated in the PM preparation to exclude peripheral Prx (120). Different stresses, from biotic stress factors like chitosan and Fusarium extracts to abiotic stress factors like wounding or H2O2 treatment, were used. In the majority of stresses, only slight changes were visible in the total activities of microsomal fractions and soluble Prx. Strong effects up to 30% were visible after H2O2, chitosan, and cantharidin treatment. This observation may be due to the compensating effects of a large number of Prx isoenzymes.
In contrast, PM preparations showed multiple strong changes in the Prx activity by the effectors tested. Additionally, the comparison of PM vesicles with the total Prx activities of the soluble enzymes and washed PM vesicles (wPM) indicated the presence of the putative peripheral membrane-bound Prx.
This approach revealed a different regulation of isoforms under the same stress conditions as discussed above. The strongest effects were found after treatment with methyl-jasmonate, salicylic acid, and H2O2. These data showed that all PM-bound isoforms may be involved in pathogen defense with different responses and that they may have a broad function in oxidative stress.
Involvement of soluble Prx in metal stress has been demonstrated (35, 111, 124). Preliminary data of our team showed that the PM-bound Prx in maize roots seem to be differentially affected by micromolar concentrations of cadmium. Due to the haem prosthetic group of class III Prx, changes in the protein abundance and activity may be postulated under iron deficiency. The weakening of PM-bound redox systems was tested under iron conditions and addition of the elicitor chitosan in pea roots (117). Three membrane-bound Prx were detected with isoelectric points of 9.8, 9.6, and 8.1, which differentially depend on iron. In comparison to the control plants treated with iron, the iron-deficient plants showed a significant decrease of the activity of Prx at pI 9.8, but no differences for Prx at pI 9.6 and Prx at pI 8.1. Treatment with chitosan and iron caused a highly significant increase of all the three detected Prx. The effect, however, was lower in the iron-deficient pea.
A putative Prx1 precursor (MWtheor. 37.6 kDa; pItheor 5.63) was found in PM preparations of rice that is influenced by salt stress (38). This Prx was presented by eight protein spots in 2D-PAGE. Changes of mass and isoelectric points in addition to a different alteration of these Prx spots suggest possible posttranslational modifications of one isoform, for example, glycosylation (MW shift) or phosphorylation (pI shift).
Copper-binding proteins
Proteomic approaches revealed several proteins that belong to the blue copper protein family (Table 1). Among these protein homologs of skewed5 (SKU5), namely, SKU5 similar 1 (SKS1) and C-terminal peptide-binding protein 1 (CBP1) have been identified in the PM. The SKU5 gene belongs to a small gene family (SKS) with 19 members in arabidopsis. Gene products of this protein family are structurally related to the multicopper oxidases, the ASC oxidase and laccase. SKU5 was identified as the putative C-terminal receptor and interaction partner of the auxin-binding protein (ABP1) in arabidopsis (154). Later on, CBP1 (73 kDa) was identified as the SKU5 homolog in maize (159). SKS proteins are attached to the PM by a GPI-anchor and are localized in detergent-resistant membranes (86, 97, 174). SKU5 was also found in cell wall fractions (154).
Copper-binding motifs appeared not very well conserved in SKS protein family members. This suggestion was supported by a lack in the enzymatic activity for the native AtSKU5 protein.
Nevertheless, copper and zinc have been detected in CBP1 (159). Thus, the redox activity of this protein family needs further proof.
Expression screens for SKU5 demonstrated high levels of transcripts in expanding tissues. A function in the two-directional growth processes and cell wall expansion was postulated for SKU5 homologs (154, 159). ABP1 appears to contribute to the directional cell elongation process, whereas the direction of cell elongation may be provided by its putative receptors the SKS proteins (159).
An evidence for a function of the SKS protein family in oxidative stress has been presented. SKU5 was enriched in detergent insoluble membrane fractions after treatment with the bacterial pathogen-associated molecular pattern flagellin (86). In arabidopsis, a blue copper protein was found to be induced by aluminium stress that caused increased resistance to oxidative stress (53).
NAD(P)H:quinone oxidoreductases
Quinones are oxidized derivatives of aromatic compounds that are produced from reactive aromatic compounds such as phenols and catechols. Long chain quinones act as antioxidants able to regenerate tocopherol inside biomembranes or as mobile electron and proton carriers in cell membranes (81, 104, 161, 189). It appears that tocopherol or ubiquinone (Coenzyme Q, CoQ) are not present in plant PM (13, 109, 150). The evidence for a membrane-embedded quinone in the PM has been given for carrot cell cultures and maize roots (17, 104, 109). Although the origin of this compound may need further proof, its relatively high concentration together with in vivo observations suggests that this compound may have a function analogous to CoQ in animal PM (104).
Autoxido-reduction of the quinolic pool, however, may produce toxic products like semiquinol radicals and finally, ROS (31, 105). Animal DT-diaphorase (NAD(P)H-quinone oxidoreductase) decreases the rate of semiquinol production by a two-electron transfer mechanism, and thereby decreases the production of ROS (49). A function of DT-diaphorase in regeneration of membrane-embedded quinones was also demonstrated (49).
NAD(P)H-dependent flavoproteins with a quinone reductase activity have been described for PM preparations of several plant species and tissues (21, 27, 42, 103, 109, 153, 157, 158, 183). In addition to these biochemical studies, proteomic approaches identified members of the flavodoxin and flavodoxin-like protein families in PM preparations (34, 75, 115 –117, 125). Studies on detergent insoluble membranes (so-called microdomains or lipid rafts) isolated from arabidopsis and tobacco leaves indicated at least three quinone reductases (34, 125). None of these enzymes, however, has a transmembrane domain or modification that could explain its interaction with the PM.
AtNQR (NAD(P)H:quinone reductase, flavodoxin protein family) and AtFQR1 (flavin quinone reductase, flavodoxin-like protein family) appear to functionally replace DT-diaphorase in plants. The structure of the founding member of the flavodoxin-like family, NADH:quinone reductase WrbA (tryptophan (W) repressor-binding protein) from Escherichia coli showed significant differences in its structure compared to DT-diaphorase (5, 54). Similar to the E.coli WrbA, both plant proteins contain FMN instead of FAD and have subunits of the similar size (96, 139, 167). AtNQR
A homolog of AtNQR was isolated from PM fractions of soybean (Glycine max L.) (153). Although this protein was demonstrated to reduce quinones by a two-electron transfer mechanism, ROS production was observed at alkaline pH values using high concentrations of menadione. This reaction appears to occur by a nonenzymatic autoxido-reduction of the quinolic pool at an alkaline pH (Fig. 6). The intermediate semiquinols are able to react with molecular oxygen, produce superoxide anion radicals, and finally, hydrogen peroxide. ROS generation by NQR and other PM-bound NAD(P)H:quinone oxidoreductases under in vivo conditions will need future research (104, 153). At least sensitivity of NAD(P)H-quinone oxidoreductase to the flavoprotein inhibitor DPI and lower abundance of flavodoxin-like proteins in PM preparations after elicitor treatment may support this suggestion (117).
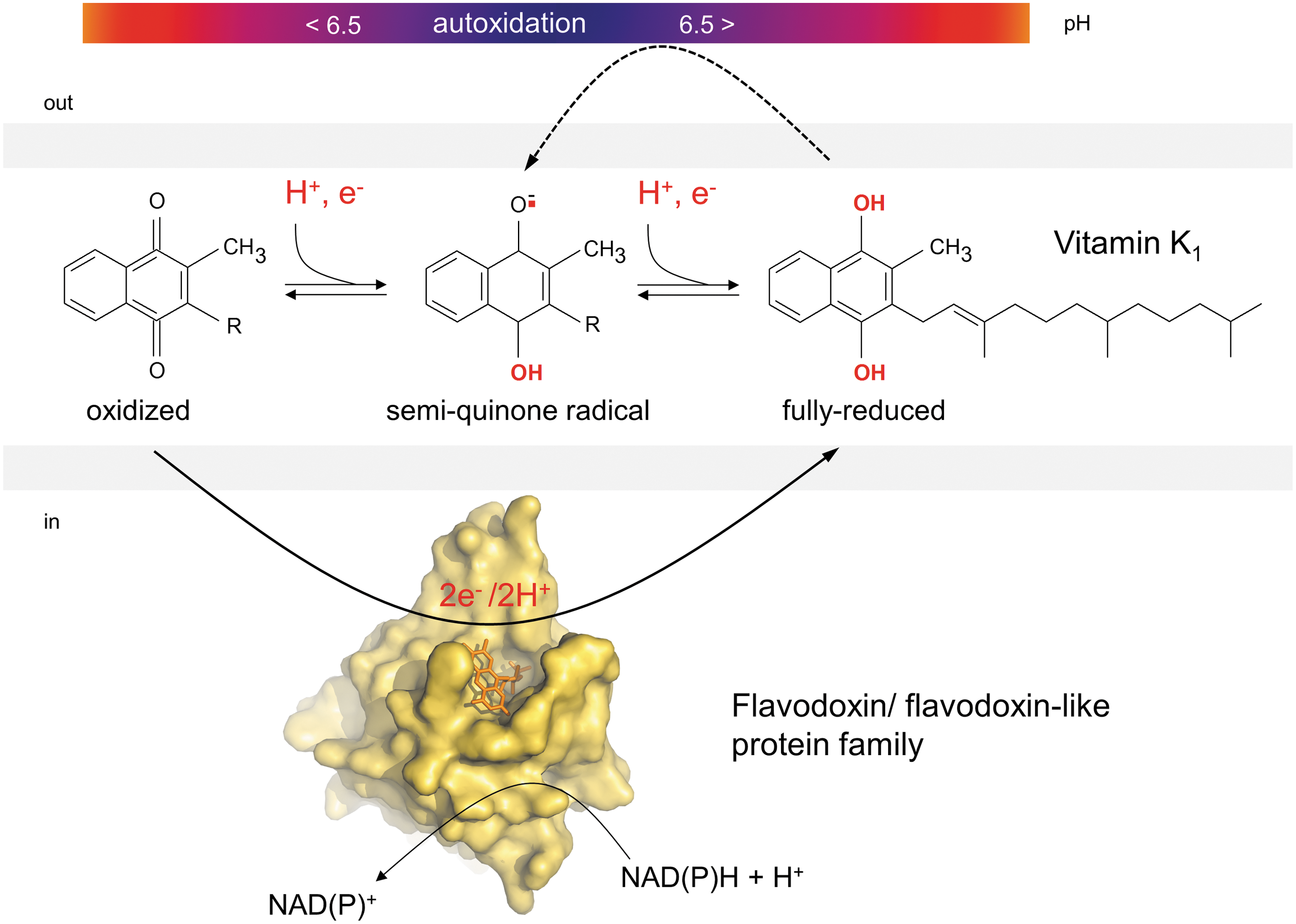
Additionally, NADH-specific quinone reductase activities were detected in root PM. The activity consisted of a single polypeptide of about 31 kDa with an isoelectric point of 8 and a pH optimum of the activity at pH 6 (157). The enzyme reduced HCF III>ferric-chelates>juglone (naphthoquinone group) and was strongly inhibited by p-hydroxymercuribenzoate, dicumarol, and thenoyltrifluoroacetone. In contrast to the protein purified from onion (Allium cepa L.) roots (157), two isoenzymes were detected in the PM of maize roots (109). A protein with properties similar to the NADH-specific quinone reductase from maize PM was characterized after purification from microsomal fractions (165). The enzyme catalyzes a one-electron transfer mechanism and was identified as cytb 5 reductase (16). Localization of this protein at the PM, however, needs further proof.
While AtFRQ1 is an auxin-regulated protein possibly involved in growth response (96), the function of NQR appears to be the protection of intracellular components from oxidative damage. This hypothesis is supported by the observation that abundance and activity of members of the flavodoxin and flavodoxin-like protein family change during a stress response (38, 110, 117). For example, salt treatment of rice (Oryza sativa) caused an increase in abundance of a 1,4-benzoquinone reductase in both salt-sensitive and salt-tolerant lines (38, 110). A similar result has been found for tomato with an increase of protein abundance of a quinone reductase by high salinity (112). The increase in abundance of flavodoxin-like proteins was also found in the PM of roots grown under iron deficiency (75, 117). In pea, a simultaneous decrease of PM-bound Prx has been observed under these conditions (117). So in all these cases, an increase of quinone reductases may protect the cell for oxidative damage by scavenging of semiquinone radicals.
In contrast, abundance of the flavodoxin family and quinone reductase activities decreased in pea roots after elicitor treatment (117). As a hypothesis, lower abundance of quinone reductases may lower the antioxidant capacity of the cell, enhances semiquinone radical release and supports ROS production during a pathogen response (Fig. 6).
MDHA reductase
A MDHA reductase (45 kDa) was isolated from the PM of spinach (Spinacia oleracea L.) leaves and identified by N-terminal amino acid sequence analysis and cross reactivity with anti-MDHA reductase antibodies (14, 21, 23). FAD acts as cofactor of the enzyme that showed characteristic substrate specificity, kinetics, and inhibitor sensitivity of MDHA reductases. The protein appeared to be strongly associated with the cytosolic surface of the PM. MDHA reductase was suggested to regenerate ASC, which may act as an antioxidant at the surface of the PM. In addition, the product may be used by a PM-localized cytb561 for transmembrane electron transfer and regeneration of ASC in the apoplast (8, 21).
Malate dehydrogenase
MDH is involved in several cellular processes, including plant development, nutrient uptake, and oxidative stress. A PM-associated cytosolic MDH (cMDH) was identified in different plant species (Table 1). The recombinant protein of arabidopsis MDH (Table 1) was found as a dimer after heterologous coexpression with recombinant thioredoxin in E. coli (71). In contrast to the monomer of the recombinant AtcMDH, the dimer was inactive. This observation suggests a redox regulation of the enzyme activity by thioredoxin. Green fluorescence protein-fusion constructs of apple cMDH support an additional localization of the cytosolic enzyme at the PM (196). The cMDH appears tightly bound to the membrane and enzyme kinetics and properties were significantly different from soluble, chloroplast, and mitochondrial isoenzymes, but also between orthologs (42, 44, 67, 68, 82, 109; Menckhoff et al., unpublished observation). Thus, reduction of a putative cMDH dimer in the cytosol may present a regulatory mechanism that allows activation of the soluble cMDH and attachment to the PM (Fig. 7). Besides this hypothesis, the mode of interaction of cMDH with the membrane has to be elucidated in the future. A cMDH isolated from kidney PM had two transmembrane spanning domains (69, 70). The sequence similarity of plant cMDH to kidney cMDH is low (60%) and transmembrane helices were not predicted within the amino acid sequence of these proteins. Solubilization of cMDH from the PM by Triton X-100 suggests a possible interaction with phospholipids, for example, phosphatidyl choline, the dominating phospholipid in plant PM (30).
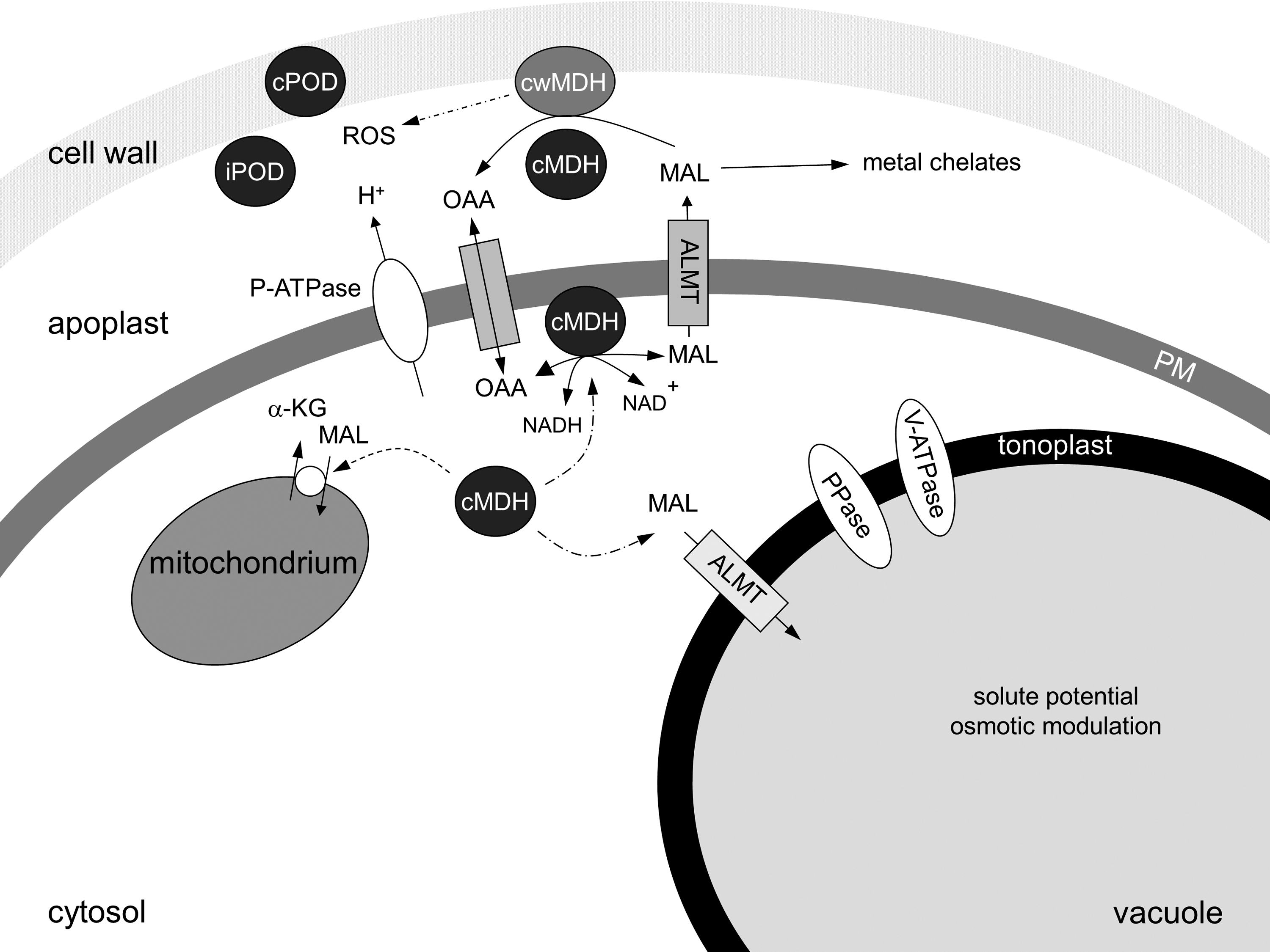
Physiological functions of PM-associated cMDH are not yet clear. Proteomic and molecular biological approaches on PM-associated cMDH suggest a function related to plant growth and stress response. Iron deficiency, cold and high salinity caused a decrease in protein abundance and a downregulation of the transcript corresponding to the PM-bound cMDH in different plant species (75, 84, 112, 195).
Functional analysis of apple cMDH by overexpression and suppressor mutants demonstrated a relation to growth and suggests a protective mechanism in the stress response (195). In comparison to the control, the overexpressor showed an increased growth, whereas growth of suppressors was decreased. Ectopic expression of the apple cMDH in tomato further supported a function in growth.
Lower abundance of cMDH at the PM was found after cold stress and high salinity (112). In apple, these stressors caused downregulation of cMDH (195). The data at hand showed that overexpression of cMDH enhanced H+-ATPase activities and accumulation of V-ATPase and PPase transcripts and revealed an increase in stress tolerance. Thus, alterations in the electrochemical gradient at the tonoplast may increase the transport of sugars, organic acids, and ions into the vacuole, which may cause alteration in solute potential (Fig. 7). Additionally, V-ATPase is the driving force for Na+-transport into the vacuole. Thus, an increase in V-ATPase may be also responsible for osmotic modulation under high salinity (195). These data suggest a regulatory function of soluble cMDH for the vacuole and its function in growth and stress response.
Additionally, a malate shuttle was postulated at the PM (42). As shown in Figure 7, cMDH convert OAA to malate, which is transported across the PM by a specific transporter (100). In the apoplast, malate fulfils miscellaneous functions such as metal complexation or production of ROS via a cell wall MDH (42, 67). A cMDH with a high-sequence similarity to PM-associated cMDH was found in the apoplast (199).
The control of pH and apoplastic redox state may depend on the direction of the reaction catalyzed by the PM-associated cMDH. For example, the H+-ATPase activity may be stimulated by protons released by malate consumption and modulate apoplast acidification (Fig. 7). Simultaneously, redox equivalents may be produced for NADH-dependent redox processes at the PM or in the cytosol. This hypothesis was supported by the Km values for malate in roots (1.8–2.5 mM) and leaves (0.3–0.5 mM) that were found for the native PM-associated cMDH (67; Menckhoff et al., unpublished observation).
In contrast to this hypothesis, cMDH from the kidney was found as a regulatory subunit of a nucleic acid channel located in the PM (70). In stomata guard cells, modulation of a voltage-dependent ion channel (SLAC1, slow anion channel) has been found by malate (156). Anion channels located in the PM are the major pathway for malate efflux (100). This implicates a key role of these channels in linking solute flux with metabolism. A recent study on stomatal guard cells demonstrated the capacity for OAA to coordinate organic acid metabolism with the anion channel current through the direct effect of the organic acid pool size (190). Thus, it appears possible that PM-associated cMDH may have a regulatory function for ion channels as well. This hypothesis was further supported by OAA-dependent activation of PM-bound anion channels that triggers a cell response to ozone stress (180). Copurification of cMDH from root PMs with a smaller protein was suggested to be due to the protein–protein interaction (42, 109). Possible interacting partners of cMDH, however, could not be identified so far.
Conclusions
Several redox systems have been identified in the past. So far, ZmDoHCytb561 appears to be a constitutive system with an unknown function. Other redox systems are differentially regulated in dependence on oxidative stress and physiological conditions. AtDoHCytb561, AIR12, FQR1, the SKS family, cMDH, Rboh, and class III Prxs appear to play important roles in plant development and growth regulation. NtRbohD was demonstrated as a major source in ROS production at the PM involved in several functions, including elicitation, wounding, and other stress responses. Elicitation caused regulation of SKU5, flavodoxin and flavodoxin-like proteins, Prxs, cytb561, and FRO. It was demonstrated that the control of iron homeostasis, growth regulation, and cell wall stiffening play central roles in pathogen defense. Metal stress regulates not only FRO, but also Rboh, Prxs, quinone reductases, AIR12, and SKU5. Differential regulation of flavodoxin and flavodoxin-like proteins, MDH, Prxs, and Cytb561 under high salinity, drought, and light stress support functions of these redox systems in regulation of the apoplastic redox status. Additionally, functions of flavodoxin and flavodoxin-like proteins, Prx and MDHA reductase in membrane protection have been discussed.
The evidence was found for a localization of PM redox systems in microdomains and participation in protein–protein interactions. Alterations in protein abundance and regulation of transcripts have been demonstrated for different redox compounds under stress conditions. In dependence on the tissue of expression, isoenzymes may fulfil distinct functions and in dependence on that may be differentially regulated during stress response. Roles of root PM are more complex than those of leaf PM. Additionally, the biotic and abiotic environment that the root surface counteracts with may be more complex. Thus, isolation of PM-bound proteins from root tissue may cause an overlay of several isoenzymes, and thereby, identification of a higher number of redox proteins. At least 2D-PAGE analysis suggest that the redox proteome of root PM is more complex compared to that of leaf.
Characterization and identification of real membrane proteins, however, is still a major challenge. Studies on structural predictions and biochemical properties of PM-bound redox systems have been presented. Experimental data of the flavocytochrome b family, but also of other protein families, showed significant differences to in silico predictions. Although a localization of the flavocytochrome b family in the PM was postulated and proved for some homologs. FRO3, FRO7, and FRO8 are localized in chloroplast and mitochondria. Members of the Cytb561 family have been detected in the PM and tonoplast. Thus, localization of Rboh in other compartments appears likely.
It is clear by now that FRO represent the turbo reductase involved in the high-affinity iron uptake system of strategy I plants. Cytb561 appears to act as iron reductase at the tonoplast (Fig. 2). In contrast, in vivo observations made for the standard system cannot be explained by any of the transmembrane compounds identified so far. In other words, a HCF III-reducible and NAD(P)H-dependent compound with transmembrane topology, stimulated by quinones and inhibited by dicumarol and auxins has not yet been identified. It may be possible that such a protein could not be identified due to the difficulties regarding identification of transmembrane proteins by mass spectrometry. However, evidence has been presented for a trans-PM electron transport chain. The data at hand suggest that electrons are transferred from cytosolic NAD(P)H to the apoplast via cytosolic quinone reductases (flavodoxin and flavoxodin-like family), a membrane embedded or soluble vitamin K, and a transmembrane Cytb561 (Fig. 8). In tandem with these electrons, protons might be transported to the apoplastic space. This hypothesis, however, will need further verification. Both Cytb561 and DoHCytb561 may be candidates for this transmembrane electron transfer chain. Cytb561, however, utilizes ASC and the natural electron donor of DoHCytb561 has not been identified so far.

Several redox compounds have been found to be induced or upregulated by auxin; thus. it appears obvious that PM-bound redox systems are involved in growth processes. In this context, members of the SKS protein family may be involved in signal transduction and Rboh in ROS production that may support cell wall loosening, whereas the role of AIR12 and other redox compounds is not yet clear. Functions of PM-bound redox systems in oxidative stress, however, have been demonstrated by several studies.
Future Perspective
Physiological functions of FRO and Rboh have been intensively studied in the past. Discrepancies of in silico predictions and biochemical studies showed that the electron transfer mechanism of the flavocytochrome b family in plants has been poorly understood. Although a central role of Rboh in ROS production has been demonstrated, a haem comparable to the neutrophils Cytb558 could not be detected in plant PM so far. The electron transfer mechanism of Cytb561 has been investigated in detail, some DoH fusion products, however, appear to have less conserved ASC-binding motifs and in some cases, transmembrane helices do not contain any of the conserved His residues for haem binding. Thus, substrates and transmembrane electron transfer by these proteins will need future research. Additionally, mutants of the Cytb561 and other protein families that occurred in the plant PM should help to elucidate their physiological functions.
Finally, several redox compounds have been identified by proteomic approaches. Detection of those proteins in detergent-resistant membrane fractions suggests a localization in microdomains. If these compounds are colocalized in the same microdomain or in different ones has to be elucidated.
Footnotes
Acknowledgments
The authors wish to thank Michael Böttger and Christian Rensch (the University of Hamburg) for providing the picture of the root staining. The work was supported by the Deutsche Forschungsgemeinschaft (DFG Lu-668/4-4).