
Abstract
Introduction
The legume–Rhizobium symbiosis
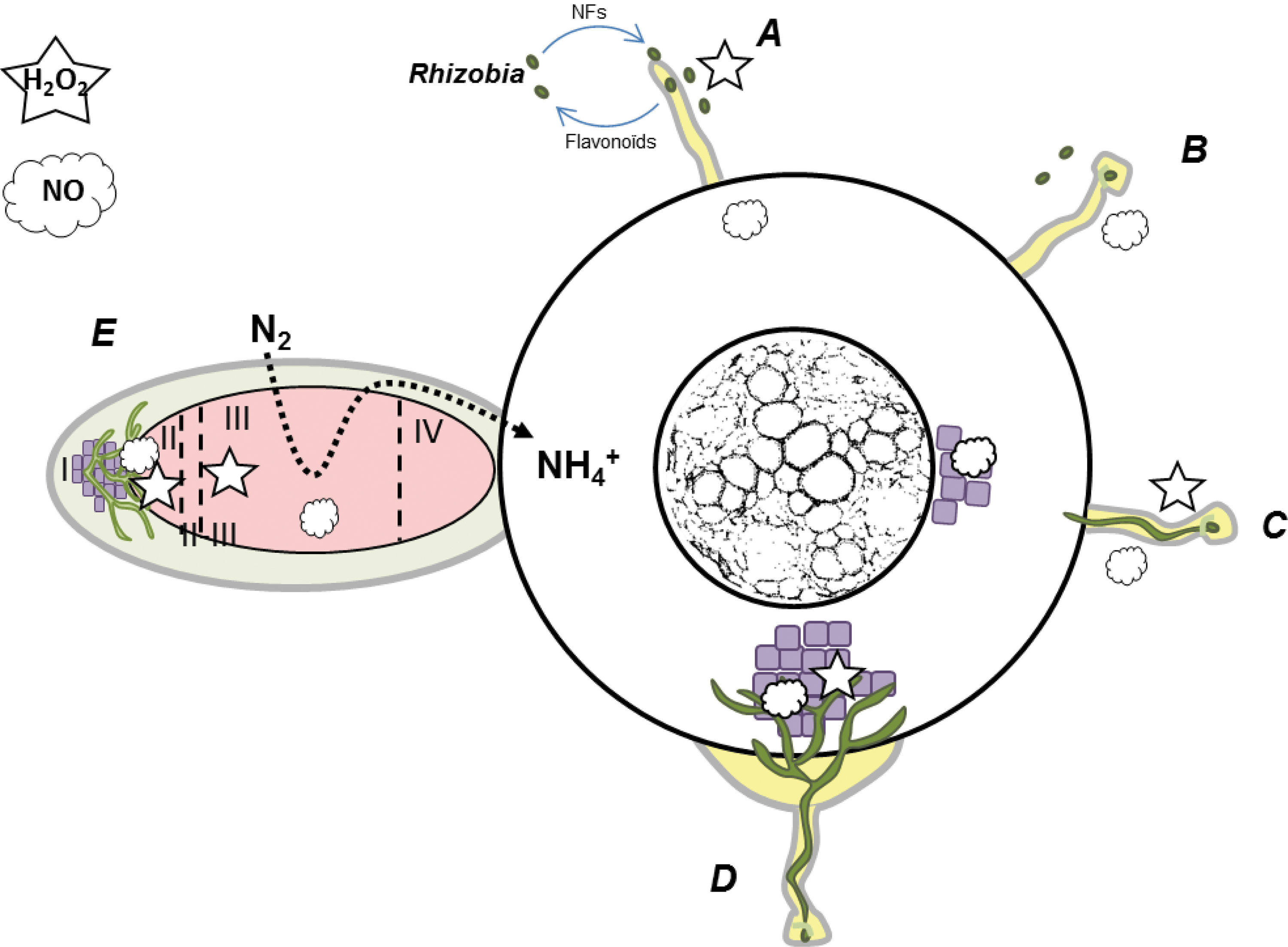
Nodules can be of determinate or indeterminate type (59). Determinate nodules are formed in legumes of tropical origin (bean, soybean, or cow pea). They are spherical and lack a persistent meristem; cell expansion, rather than cell division, is responsible for nodule growth (42). Indeterminate nodules are formed in legumes of subtropical and temperate origins (pea, clover, and Medicago truncatula) and have a persistent meristem that often yields a cylindrical or branched structure. Five zones can be distinguished in these nodules (Fig. 1): zone I made of meristematic cells; zone II where cells enter the nodule differentiation program, characterized by division arrest, endoreduplication phenomena accompanied by increased cell size; these cells are infected by bacteria (interzone II-III) that differentiate into bacteroids; zone III where bacteroids convert N2 into ammonia, which is exported to the plant in exchange for carbohydrates; zone IV where the symbiotic partnership is lost and senescence occurs; and zone V where bacteria are essentially free living (143).
Reactive Species Accumulation During the Symbiotic Interaction
Transient increasing levels of reactive oxygen species (ROS) have been detected at the tip of actively growing root hair cells within seconds after NF addition in Phaseolus vulgaris (21). Interestingly, the situation may be different after several minutes where the production of hydrogen peroxide (H2O2) appears to be inhibited by NF (81, 131). In the same way, a Sinorhizobium meliloti nodC− mutant, defective in NF biosynthesis, showed an increase in H2O2 accumulation (17). Moreover, the compatible interaction between Medicago sativa and S. meliloti is linked, at least in part, with an increase of the antioxidant defence (particularly catalase and lipoxygenase) during the preinfection period (17). More recently, the role of ROS in root hair deformation in the M. truncatula–S. meliloti symbiosis has been highlighted (81). Exogenous application of ROS prevented root hair swelling and branching induced by NF. In addition, transient treatment of roots with diphenyleneiodonium (DPI; a flavoprotein inhibitor) mimicked NF treatment and resulted in root hair branching in the absence of NF. Despite possible side effects of DPI (flavoproteins, such as dehydrogenases, and oxygenases maybe DPI targets), these results suggest a role for the transient reduction of ROS accumulation in governing NF-induced root hair deformation in legumes (81). The generation of ROS in the cortical cells of M. truncatula roots after inoculation with S. meliloti was observed in vivo (114). A gradual increase of ROS production was observed in inoculated plants, which peaked at 24h after inoculation and remained high even after 48h (112, 113). During the infection process, superoxide anion (O2 −) and H2O2 can be detected in ITs and in infected plant cells (Fig. 2), indicating their production during this process (120, 124, 129). Moreover, this production was not observed when M. truncatula plants were inoculated with a S. meliloti nodD1ABC mutant, unable to produce NF, indicating that the oxidative burst is activated downstream of NF perception (120). This production was different in response to pathogenic and incompatible bacteria, both in terms of timing and in terms of subcellular localization (113). In the semiaquatic legume Sesbania rostrata, the formation of cortical infection pockets depends on an NF and was shown to be associated with localized cell death and the production of large amounts of H2O2 (28).

In the later stages of the nodulation process, H2O2 and O2 − can be observed in infected cells of young nodules, revealing a prolonged production of these radicals during nodule development. H2O2 production was detected in ultrathin sections of mature 6-week-old nodules as an electron-dense precipitate with cerium chloride (124, 129). A large cerium oxide labeling was observed in the cell walls of infected cells and in some ITs around bacteria. In both cases, the H2O2 stain was associated with the plant cell wall (124, 129). Moreover, a strong cerium oxide precipitate can also be observed around peribacteroid and bacteroid membranes of senescent infected cells, strongly suggesting that H2O2 is involved in the senescence process (1, 124).
During the last decade, increasing evidence of the occurrence of nitric oxide (NO) during legume–Rhizobium symbiosis, from the early steps to the nitrogen-fixing nodules, has been reported. Through the use of NO-fluorescent probes (4-amino-5-methylamino-2′,7′-difluorofluorescein [DAF-FM], diaminorhodamine-4M acetoxymethyl ester [DAR-4M AM]), NO production was transiently observed in L. japonicus and M. sativa roots 4 h after inoculation with their cognate symbionts, suggesting that NO production could result from the specific recognition of the plant and bacterial partners (100, 132). In M. truncatula roots, using both a specific NO biosensor S. meliloti strain and the cell-permeable fluorescent probe 4,5-diaminofluorescein-2 diacetate (DAF-2DA), del Giudice et al. (31) demonstrated that NO is produced at different sites during the infection process: in the shepherd's crooks of root hair, in the ITs leading the bacteria to the cortical cells, and in the nodule primordial (Fig. 2). In mature nodules, an NO synthase (NOS)-like activity has been measured in lupine nodules (27), but the first evidence of NO occurrence came from the detection of NO complexed to leghemoglobin (Lb), the major hemoprotein of legume nodules, using electron paramagnetic resonance. The presence of such nitrosylleghemoglobin (LbNO) complexes was reported in several legumes (69, 84, 86, 88, 127), representing determinate and indeterminate nodule types. Based on the use of DAF-2DA fluorescent probe, NO formation has also been detected in bacteroid-containing cells of the fixation zone in M. truncatula–S. meliloti nodules (6, 110). Recently, using pharmacological and genetic approaches to modulate NO level inside nodules, Cam et al. (20) provided evidence that NO regulates senescence in M. truncatula nodule, suggesting that it is also produced in the late phase of the symbiosis process.
The origin of the accumulating ROS during rhizobial infection and nodule organogenesis is still not completely solved. As mentioned earlier, the formation of superoxide radicals in the ITs (129), the abolished ROS production using DPI (114), and the correlation between the decrease in transcript accumulation for two putative NADPH oxidase (Nox) genes and the decrease of the H2O2 efflux during the early steps of the M. truncatula–S. meliloti symbiosis (81) favor the hypothesis of the involvement of Nox in ROS synthesis. In this context, attention was recently paid to the identification and the characterization of Nox in legumes (81, 83, 98). Indeed, several RBOHs genes (respiratory burst oxidase homologues) have been identified in legume genomes (83). Their involvement in the synthesis of symbiotic H2O2 needs to be fully evidenced, but down-regulation of legume Rbohs led to symbiotic phenotypes mimicking H2O2 deficiency (Table 1) (67).
Arrows indicate the increase (↑) or decrease (↓) of the corresponding RUS/RNS.
ITs, infection threads; nd, not determinated; KO, knock out; OX, overexpression; Hmp, hemoprotein; H2O2, hydrogen peroxide; Kat, catalase; NO, nitric oxide; NR, nitrate reductase; Rboh, respiratory burst oxidase homologue; ROS, reactive oxygen species; nd, not determined.
Other possible sources for H2O2 are cell wall peroxidases, germin-like oxalate oxidases, and diamine oxidases (152). Indeed, a germin-like oxidase from Pisum sativum exhibiting a superoxide dismutase (SOD) activity was associated with root nodules (50). In the tropical legume S. rostrata, a gene encoding a functional class III peroxidase (Srprx1) was shown to be transiently up-regulated during the nodulation process (33). Its expression was not induced after wounding or pathogen attack, suggesting that this peroxidase is a symbiosis-specific isoform (33).
In plants, NO synthesis is still a matter of debate (14, 24, 99). Several NO-generating systems have been proposed, such as NOS-like proteins (24), nitrate reductase (NR) (30, 122), mitochondrial electron transfer chain (ETC) (52, 64), polyamine oxidase (154), and nitrite-NO reductase (136). The origin of NO in the nitrogen-fixing symbiosis (NFS) is unclear, and various NO-sources have been analyzed depending on the developmental stage of the symbiotic interaction. Several studies investigated the potential involvement of an NOS-like enzyme in young and mature nodules. Thus, NO and L-[14C] citrulline were found to be produced in an L-arginine-dependent manner in lupine (Lupinus albus) nodule extracts (27), and the authors showed that L-citrulline production was inhibited by NG-monomethyl-L-arginine (L-NMMA), an NOS inhibitor, in a competitive way. Using the NO fluorescent probe DAF-2DA, Baudouin et al. (6) showed that the addition of L-NMMA in M. truncatula-S. meliloti nodule slices impaired NO detection. More recently, another NOS inhibitor, Nω-nitro-L-arginine (L-NNA) was found to negatively affect the growth and viability of soybean-Bradyrhizobium japonicum nodules (76). These data suggest that an NOS-like enzyme is operating and required for the development of functional nodules, but the molecular identity of such an enzyme remains unknown. Using both pharmacological and genetic approaches, Horchani et al. (61) addressed the role of plant NR and mitochondrial ETC in NO production in M. truncatula-S. meliloti nodules. NO production was found to be inhibited by tungstate (Tg), a NR inhibitor. Moreover, nodules obtained with plant NR RNA-interference double knockdown (MtNR1/2) exhibited reduced NR activities and NO production levels (Table 1). The reduction of NO production was reversed by nitrite addition, both in the Tg-treated nodules and in MtNR1/2 RNAi nodules, indicating that NO synthesis depends on NR activity, but that NR does not produce NO directly. The inhibition of NO production by ETC inhibitors such as rotenone, antimycin A, or myxothiazol indicated that mitochondrial ETC was the site of NO2 − reduction into NO (61). Thus, in M. truncatula nodules, NO3 − may be reduced into NO in a two-step mechanism involving successively NR and ETC.
In rhizobia, denitrification is dependent on the napEDABC, nirKV, norCBQD, and nosRZDYFLX genes that encode NR, nitrite reductase (NiR), NO reductase, and N2O reductase, respectively (7). The expression of the denitrification genes nirK, norC, and nosZ has been reported in soybean-B. japonicum functional nodules (93). Using napA- and nirK-deficient mutants, Delgado's team (88, 127) showed that bacteroid NR and NiR contribute to the NO formed in soybean-B. japonicum nodules, particularly under hypoxic conditions (Table 1). Similarly, using the mutant approach, Horchani et al. (61) showed that around one-third of the NO generated by the M. truncatula-S. meliloti nodules is produced by the bacteroid denitrification pathway. In free-living S. meliloti cells, an L-arginine-dependent NO production has been reported (115). Thus, although no bacterial NOS-like activity has been evidenced in symbiotic nodules, the involvement of such an enzyme cannot be excluded.
Role of H2O2 and NO in Symbiosis
Role in the establishment of the symbiotic interaction
The putative role of H2O2 during the legume-Rhizobium symbiosis was first evidenced using bacterial catalase mutants impaired in their antioxidant abilities (67, 68). Since these mutants are affected at various steps of nodule development, it is assumed that the regulation of H2O2 level may be involved in different developmental processes (67, 68). H2O2 appears to somehow control a key step of the interaction: the bacterial differentiation into the symbiotic form. A S. meliloti catalase mutant (katB/katC) displayed a drastic phenotype during symbiosis, with nodule abortion and absence of bacteroid differentiation (68). Interestingly, a S. meliloti glutaredoxin mutant displayed a similar phenotype (12), indicating that a fine tuning of the ROS balance is essential to the differentiation process. It is known that in indeterminate nodules, the bacterial differentiation process is under the control of nodule-specific cysteine-rich (NCR) antimicrobial peptides. These redox-sensitive peptides (53) are targeted to the plant cell secretory pathway (92), and some of them are delivered into the bacteroid and trigger bacterial differentiation and/or membrane damage and permeabilization (146). Thus, ROS may regulate, via the NCR redox status, the differentiation process.
On the other hand, another S. meliloti mutant, overexpressing a catalase gene, displayed a delayed nodulation phenotype associated with aberrant IT (67). The catalase over-expression likely diminished the internal H2O2 concentration of the bacteria progressing inside the IT, as observed in free-living conditions. Thus, a positive role for H2O2 during IT elongation has been proposed; it may be related to IT rigidity (121) or to a cytoplasmic signal used by the bacteria to regulate symbiotic function (111). This mutant appeared to be deregulated in a set of pSYMb clustered genes localized closed to the exopolysaccharides (EPS) genes (Mandon et al. unpublished observations). Moreover, redox-dependent regulators might be affected in the reduced environment of the mutant cytosol. CbrA, which is essential for bacterial infection (46) and that is a possible link between bacterial colonization and redox signalisation (47), may be a good candidate. Alternatively, specific post-translational H2O2 protein modification might occur in IT as observed for nitrogen fixing bacteroids (102).
On the plant side, initial reports using pharmacological approaches have evidenced that ROS are essential to the symbiotic process. Indeed, the alteration of ROS production with inhibitors (such as DPI) and ROS scavengers showed that in lateral root base nodulation of S. rostrata, ROS were required for nodule primordia initiation (28). The DPI treatment also suppressed the root hair curling and IT formation, further pointing to ROS involvement in the early steps of symbiotic interactions (114). Indeed, the involvement of Nox has been strongly suggested in different reports (81, 85, 114). Moreover, it has been shown very recently that a gene encoding a Nox in common bean is crucial for successful rhizobial colonization and probably maintains proper ITs growth and shape (98).
Ras-related C3 botulinum toxin (RAC)/Rho of plant (ROP) proteins (rho-related GTPases of plants) are small G proteins that function as molecular switches within elementary signal transduction pathways, including the regulation of ROS generation during early microbial infection via the activation of plants RBOH. RNA interference silencing of MtROP9 has been studied in M. truncatula - microbe interactions (71). MtROP9i transgenic lines showed a clear growth-reduced phenotype and revealed neither ROS generation nor MtROP9 and MtRBOH3/E gene expression after microbial infection (symbiotic arbuscular mycorrhiza fungus, S. meliloti, and the pathogenic oomycete Aphanomyces euteiches). Interestingly, MtROP9 knockdown clearly promoted mycorrhizal early hyphal root colonization, while rhizobial infection was clearly impaired (71). These results suggest a key role of MtROP9 in ROS-mediated early infection signaling.
The production of NO along the infection process points to a role of this reactive nitrogen species (RNS) in the specific recognition between the plant and the bacterial partners. In the M. truncatula - S. meliloti interaction, both NO-scavenging by 2-[4-carboxyphenyl]-4,4,5,5-tetramethyl-imidazoline-1-oxyl-3-oxide (cPTIO) and hmp (a bacterial hemoprotein involved in NO detoxification) (117) over-expression in the plant partner led to a delayed nodulation phenotype (31), indicating that NO is required for an optimal establishment of the symbiosis process. In contrast, the over-expression of class-1 non symbiotic hemoglobins (ns-Hb1) genes LjnHb1 or AfnHb1 in L. japonicus induced a decrease in NO production and resulted in an increased nodule number (133), suggesting that NO negatively affects the nodulation process. The apparent contradiction in nodulation phenotypes raises the question of the effect of Hmp/Hb on NO level (31, 115, 133), and suggests that a tightly regulated level of NO is necessary for successful establishment of the symbiotic relationship. It was suggested that, in the legume-Rhizobium symbiosis, early production of NO by the plant could serve to induce the expression of bacterial genes which are necessary to adapt bacteria to NO encountered at later symbiotic stages. The lower competitiveness of the hmp ++ strain as compared with the WT is an argument in favor of such a role of NO during the infection process. It was also shown that during the symbiotic association between fungi and lichens, the mycobiont-driven NO had an important role in the regulation of the oxidative stress of lichens (i.e., during rehydration) (23). A similar role could be conceived in the symbiotic interaction where an oxidative burst occurs in alfalfa early after infection with symbiotic bacteria (129).
During the early steps of symbiosis establishment, NO was also detected in dividing cortical cells of the root, not yet invaded by the bacteria (31). This NO production is similar to that detected in lateral root primordia for which NO was reported to have a role in the cell cycle regulation and in the control of primary and adventitious root organogenesis (26, 75). As a consequence of NO decrease, delayed nodulation and reduced expression of genes involved in nodule development (MtCRE1, MtCCS52A) reinforce the hypothesis that NO could play a similar function in nodule organogenesis (31).
Role in the functioning nodule
In S. meliloti, various ROS scavenging enzymes are expressed in the nitrogen fixing zone. The SodB was detected in nitrogen fixing bacteria (36), and a sodA mutant displayed different symbiotic phenotypes depending on the strain used (128). Similarly, the disruption of the Mesorhizobium loti sodA gene led to variable symbiotic defect depending on the lotus cultivar used (125). The S. meliloti katA/katC double mutant displayed a reduce nitrogenase activity, and bacteroids underwent quick senescence (68). M. loti contains two catalases, the monofunctional catalase KatE and the bifuctional catalase peroxidase KatG. According to their expression profiles, the katG mutant exhibited no symbiotic phenotype, while katE mutant formed nodules with a decrease in nitrogenase activity (55). Rhizobium etli and B. japonicum genome possess only one catalase gene, and corresponding mutants were not affected in nodule functioning (37, 108). However, other ROS-scavenging enzyme might reduce H2O2 in addition to the metal-containing enzyme catalase. Good candidates are peroxiredoxins: These peroxidases that use redox-active cysteines for catalyses display a wide variety of substrate from H2O2 to organic and fatty acid peroxides. For instance, bacteroids from the double 2-Cys peroxiredoxin/catalase mutant of R. etli displayed a 40% reduction of nitrogen fixation capability (37). The S. meliloti prxC gene is fully expressed in the bacteroïds (Mandon et al., unpublished observations), and the corresponding protein was previously detected by proteomic analysis (36). Most peroxiredoxins reduce hydroperoxides using thioredoxin (Trx) or other thiol-containing reducing agents; Trx, in turn, is reduced by NADPH through Trx reductase (153). Interestingly, S. meliloti (CE52G) Trx-like (trxL) mutant displayed a slight decrease in nitrogen fixation efficiency (22), and the presence of TrxB in nitrogen-fixing bacteroids was also reported (36). In the same way, a mutant affected in glutathione biosynthesis exhibited higher catalase activity and induced earlier-senescing nodules (56). Rhizobacteria have evolved redundant strategies to maintain a reducing environment that is compatible to nitrogenase activity. This should be crucial for the nodule functioning, as the redox state may influence numerous enzyme activities. Among the 20 sulfenylated enzymes detected in the nitrogen-fixing bacteroids, six belong to the tricarboxylic acid cycle, which is essential for energy metabolism (102). In addition, in B. japonicum, the key regulator of the nitrogen fixation enzymes FixK2 is reversibly susceptible to oxidation (96).
The first genetic evidence of the importance of an RBOH for optimal functioning of the M. truncatula–S. meliloti symbiotic interaction was recently reported (83). The expression of an RBOH-encoding gene (MtRbohA) was found to be restricted to the N2-fixing zone. A strong link between nodule N2 fixation and up-regulation of MtRbohA was reported. Furthermore, down-regulation of MtRbohA expression led to a decrease in nodule N2-fixation activity and down-regulation of the microsymbiont genes encoding proteins of the nitrogenase complex (83). Moreover, MtRbohA expression was strongly increased by hypoxic conditions, suggesting a role for RBOHs in sensing the microaerophilic environment of the nodule. Altogether, these data indicate that MtRBOHA plays a role in communication between the plant and the microsymbiont.
Until recently, the presence of NO in mature nodules was puzzling. Indeed, Baudouin et al. (6) showed that NO is significantly produced in the fixation zone of M. truncatula nodule, but at the same time, nitrogenase activity is known to be strongly inhibited by NO (70, 144). These observations raised two questions: (i) How can nitrogenase be protected from the deleterious effects of NO produced in the fixation zone? and (ii) What is the role of NO in the mature nodule? An element of response to the first question comes from the presence of Lb and ns-Hb1 in symbiotic nodules. Lbs, which accumulate to a millimolar concentration in the cytoplasm of infected nodule cells, are thought to buffer free oxygen in the nanomolar range, avoiding inactivation of nitrogenase, while maintaining high oxygen flux for respiration (3, 106). In addition, deoxy-Lb was shown to bind NO with a high affinity to form stable complexes (86), and it was suggested that Lb could function not only as an O2 − supplier for respiration, but also as an NO scavenger to protect nitrogenase and preclude NO-damaging effect on N2-fixing metabolism (57, 58). Furthermore, ns-Hb1 proteins have been found to present a higher affinity for oxygen than Lb, implying that they could not supply O2 for terminal oxidase in mitochondria (4), and they have been supposed to play the role of NO scavenger in NO detoxifying pathways (51, 63). A similar function was devoted to the flavohemoprotein Hmp in the bacterial partner (90). Several studies on the role of plant and bacterial hemoglobins (Hbs) in N2-fixing nodules support this hypothesis. Thus, in L. japonicus and M. truncatula nodules, the over-expression of either plant ns-Hb1 in the plant partner (100, 133) or bacterial Hbs in the rhizobial partner (90, 119) led to an enhanced symbiotic N2 fixation. Conversely, a rhizobial hmp mutant impaired N2 fixation in M. truncatula-S. meliloti nodules (90). Taken together, all these observations argue in favor of a protective role of plant and bacterial Hbs to preserve nitrogenase from the NO produced either by the bacterial denitrification pathway, or by the plant NR and ETC.
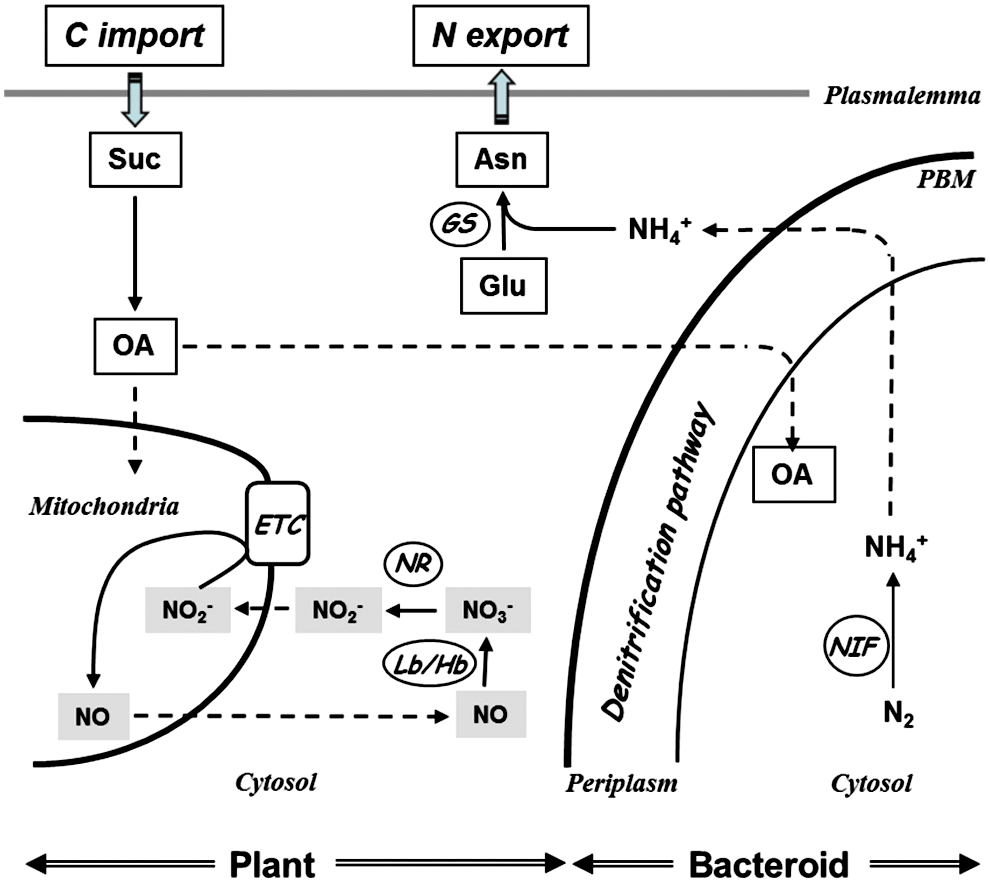
In response to the second question, a metabolic role for NO in mature nodules has been recently highlighted (61, 89). NO production has been shown to be induced in the roots of plants submitted to hypoxia, and this production was supposed to be linked —via a cyclic respiration process—with improved capacity of the plants to cope with hypoxic stress and to maintain cell energy status (64). This cyclic respiration, called “nitrate-NO respiration,” involves four successive steps (Fig. 3): (i) the reduction of NO3 − in NO2 − by the cytosolic NR; (ii) the translocation of NO2 − from the cytosol into the mitochondria, presumably through a nitrite transporter; (iii) the reduction of NO2 − in NO, via the mitochondrial ETC, thus allowing respiration functioning and ATP regeneration; and (iv) the passive diffusion of NO from the matrix to the cytosol, where it is oxidized by Hb into NO3 − . Functional nodules of G. max (88) and M. truncatula (61) have been shown to increase their production of NO when submitted to hypoxic conditions, and it was suggested that, under the microaerobic conditions prevailing in nodules, the nitrate-NO respiratory pathway may contribute to energy supply for nitrogen fixation (61). Several lines of evidence argue in favor of this hypothesis. First, plant NR and ETC, and bacterial denitrification pathway contribute to NO production, via NO3 − and NO2 − reduction, particularly under hypoxic conditions (61, 127). Second, Lb and class-1 ns-Hb have the capacity to efficiently react with NO to produce NO3 − with an elevated rate constant (58), and the NO generated at ETC level may, therefore, be oxidized into NO3 − by Lbs and/or ns-Hbs. Third, the energy status of the nodules depends either significantly, or almost entirely, on NR functioning under normoxic or hypoxic conditions, respectively (61). Thus, in symbiotic nodules, a role related to NO scavenging and metabolism may be fulfilled by Hbs in the plant partner and Hmp in the bacterial ones. The high affinity of these Hbs for NO and their capacity to oxidize NO into NO3 − would be favorable to keep NO concentration below toxic level for nitrogenase, and to supply the nitrate-NO respiratory cycle in order to maintain the energy status required for nitrogen fixation under limiting oxygen conditions.
Using a S. meliloti hmp mutant strain, Cam et al. (20) recently showed that an increase in the NO level inside the nodule causes its premature senescence, whereas over-expression of Hmp in nodules leads to a significant delay in nodule senescence, and partly relieves dark-induced senescence of the nodules. In addition, coinoculation experiments with either wt/hmp or hmp/hmp++ S. meliloti strains showed that, on the same plant root, an increase in NO level in one nodule (hmp) triggers senescence in wt nodules, but has no effect on hmp++ nodules. Taken together, these observations support the hypothesis that NO is produced during aging of legume nodules and is able to act as a local senescence signal, either directly or via the plant root.
H2O2 and NO Molecular Targets in the Interaction
Gene targets
Although progress has been recently made on the identification of genes regulated by ROS in plants (44, 49, 66, 101), only little data are available with regard to the rhizobial symbiosis. MtRip1 (Rhizobium-induced peroxidase 1) has been shown to be induced by H2O2 (120). In this context, by using a combination of pharmacological (DPI, a widely used Nox inhibitor) and transcriptomic approaches, we identified genes putatively targeted by H2O2 during the establishment of the M. truncatula–S. meliloti symbiosis (2), including genes involved in hormone synthesis or signaling, in cell wall synthesis (potentially related to IT progression), and in transcription regulation. Moreover, 22 M. truncatula genes displayed a sequence similarity to 19 Arabidopsis genes previously described as regulated by ROS (2, 44). This is in line with a performed meta-analysis of H2O2-induced gene expression in eukaryotes (including Arabidopsis), showing potentially strong conservation of the regulation of gene expression by ROS across kingdoms (148). A candidate gene, MtSpk1 (Symbiotic Protein Kinase 1) was induced by H2O2 and during nodulation (29). The functional analysis indicates that MtSpk1 plays an important role in establishment of the rhizobial symbiosis (2).
Large-scale transcriptome analyses have been performed to identify NO-responsive genes in plants, but only a few have been conducted on symbiotic interaction. Most of these transcriptome analyses were carried out in Arabidopsis with the use of NOS inhibitors, NO donors, and the NO scavenger cPTIO, with the main objective of determining the NO molecular targets during pathogenic interactions (62, 109, 157). Although the exploitation of data from different biological systems and experimental conditions is a challenging issue, the comparative analysis of these transcriptomic data revealed a significant number of genes related to plant adaptive responses to biotic and abiotic stresses in these NO-responsive targets (13). The first transcriptomic study conducted on a symbiotic interaction from M. truncatula roots treated with two NO donors, SNP and GSNO, enabled the identification of 999 putative NO-responsive genes (40). Of these genes, 290 also were regulated during the nodule development. Interestingly, genes involved in the control of cellular redox responses (peroxidase and germin-like oxalate oxidase) were also identified. Complementary analysis on the two genes involved the glutathione synthesis, γ-glutamylcysteine synthetase, and glutathione synthetase (GSHS), provided the first evidence that GSH synthesis is stimulated by NO in plants (65). In M. truncatula, NO scavenging resulting either from overexpression of a bacterial flavohemoglobin hmp gene, or from cPTIO treatment, was found to down-regulate several genes involved in nodule development, such as MtCRE1 and MtCCS52A (31). MtCRE1 encodes a cytokinin receptor that plays a major role in the development of the symbiotic interaction between M. truncatula and S. meliloti (48). This suggests that NO may control the nodulation process through the regulation of cytokinin perception.
Recently, a transcriptomic analysis using RNA-Seq technology was performed with M. truncatula inoculated roots treated with cPTIO to identify genes potentially regulated by NO during the nodule primordium development (16). One of the main results of this study lies in the reversal, on cPTIO treatement, of the down-regulation of defense genes provoked by the inoculation with rhizobia. A number of genes involved in terpene, Flavonoid, and phenylpropanoid pathways, and genes encoding PR-proteins, GSH S-transferases, and cytochrome P450 were notably affected by cPTIO. In contrast, cPTIO triggered the repression of genes potentially involved in nodule development such as regulation of transcription, and protein transport, degradation, and synthesis. These findings suggest for the first time that NO could be involved in repressing the defense reaction during the symbiotic interaction, thereby favoring the establishment of the beneficial plant-microbe interaction. This action differs markedly from the signaling functions of NO in pathogenic interactions, in which NO cooperates with ROS to induce hypersensitive cell death, and functions independently of them in the induction of defense-related genes (32).
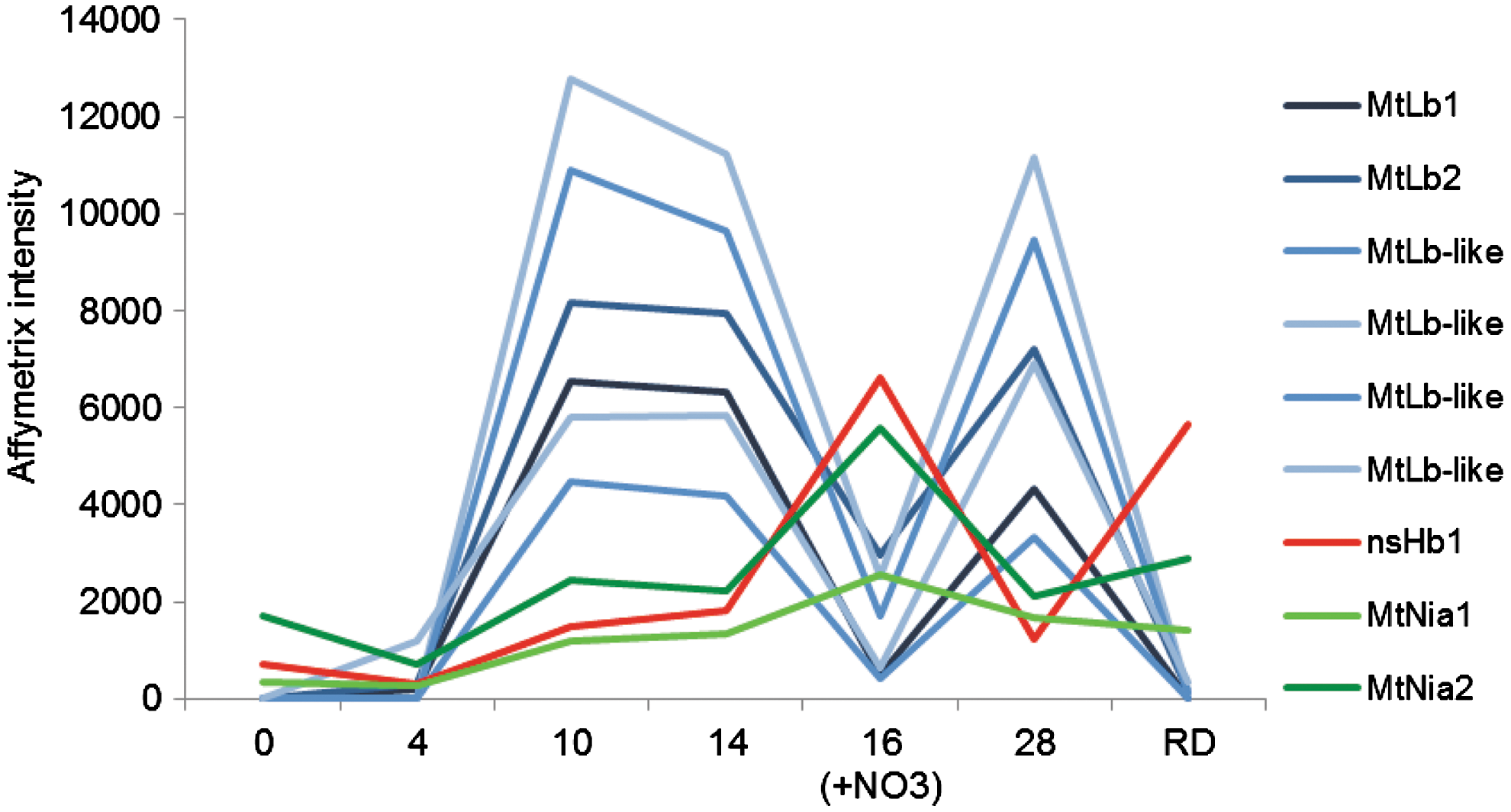
Among the NO-responsive genes in early legume–rhizobium interactions, ns-Hb1 holds particular attention. As mentioned earlier, an NO-scavenging activity has been associated with ns-Hb1 in NO-detoxifying pathways (51, 63). In M. truncatula, we observed a decrease in the transcript level of ns-Hb1 at four days postinoculation, suggesting that a decrease in ns-Hb1 level could be related to an increase in the NO level in the root during the establishment of the symbiotic interaction (16). In mature nodules, the plant NO-responsive genes are still to be deciphered. However, an in silico analysis of the expression profile of the different Hbs expressed in M. truncatula nodules (Medicago truncatula gene expression Atlas, Noble foundation) showed that after 2 days of nitrate treatment, all Lbs are strongly repressed; whereas at the same time, ns-Hb1 and the 2 NRs are strongly induced (Fig. 4). In a number of species, ns-Hb1 was reported to be nitrate- and NO regulated (18, 103, 118, 130). Considered together, these data argue in favor of the role of ns-Hb1 in the recycling of NO through the nitrate-NO respiration process in the nodules (Fig. 2) (61).
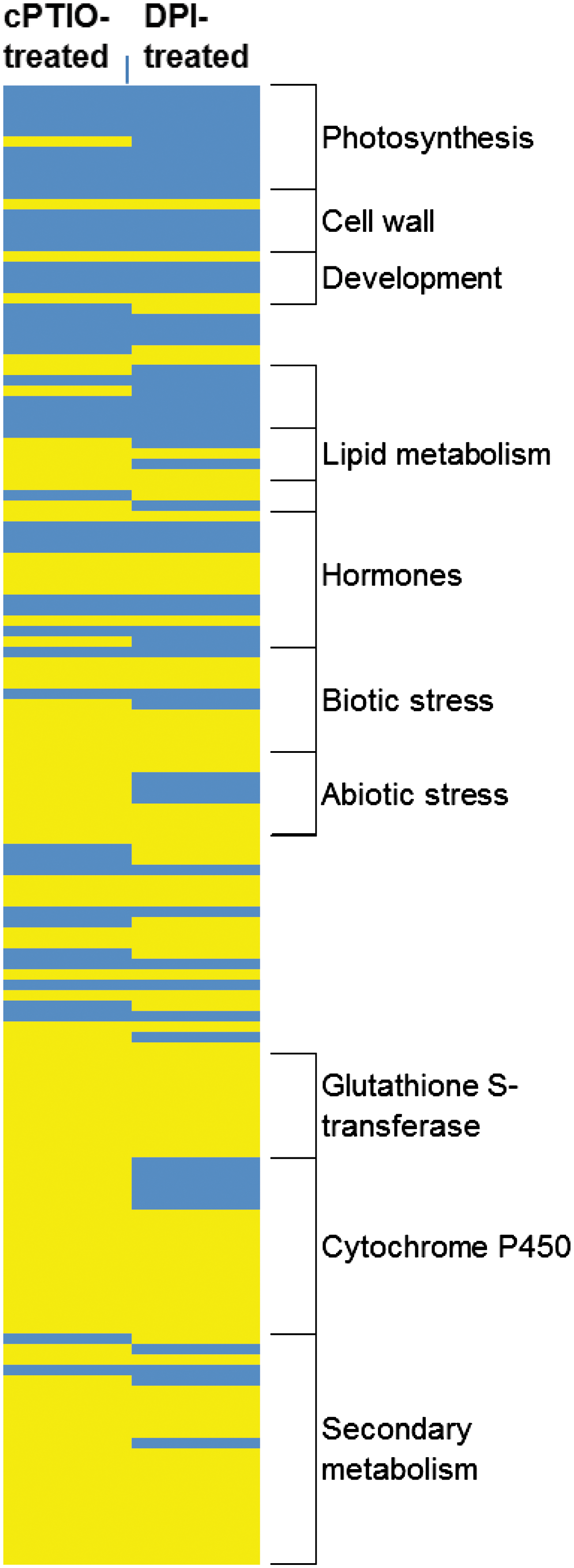
Interestingly, the comparison of M. truncatula inoculated roots treated with either DPI (2 dpi) or cPTIO (4 dpi) reveals that 316 genes (75% of the genes deregulated by cPTIO and DPI) are similarly regulated (Fig. 5). It cannot be excluded that DPI affects NO generation, as observed in mammalian cells where DPI was shown to inhibit NOS (137). However, in our analyses, only 316 out of the 2030 cPTIO-regulated genes (15%) were affected by DPI, suggesting a limited effect of DPI on NO regulated genes. Indeed, gene families involved in photosystem synthesis, cell wall formation, and development processes are mainly down-regulated, while gene families involved in plant defense and secondary metabolism are up-regulated. These results point to a significant cross-talk between ROS and RNS during the establishment of the symbiotic process as previously highlighted during cell death induction (156).
The NO-responsive genes of the bacterial partners, B. japonicum, S meliloti, and R. etli, have also been studied essentially in culture conditions. A transcriptomic approach was undertaken to identify the set of bacterial genes induced by NO in culture of S. meliloti. About 100 genes were clearly identified as up-regulated by NO (90), with a 70% overlap between the NO and microaerobic stimulons (15, 90).
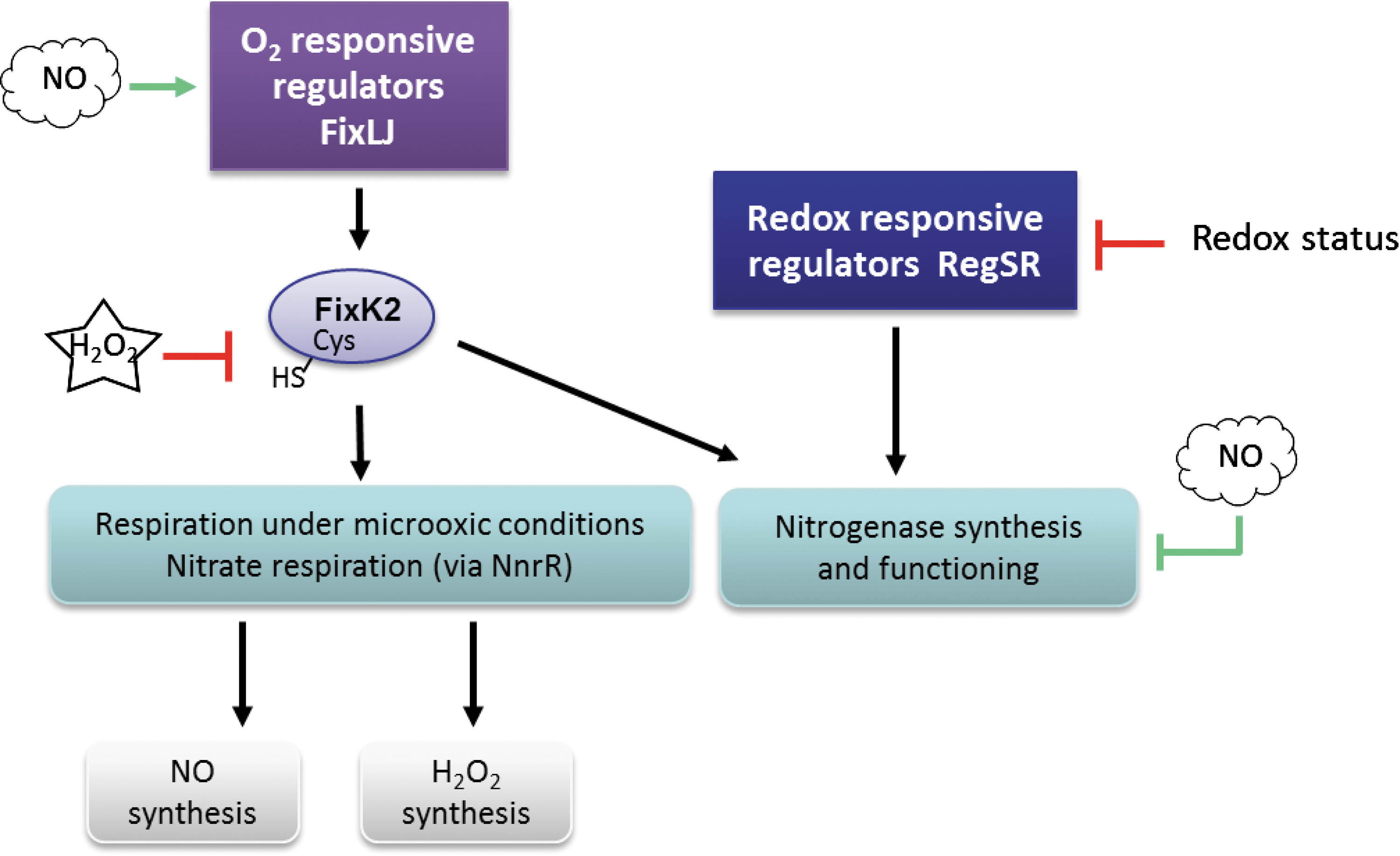
Most of these genes are under the control of the two-component regulator FixLJ, a major regulator of the oxygen limitation response, and the regulator NnrR (Fig. 6) (90). NnrR was shown to be an NO sensor involved in the specific response to NO in a number of bacteria species (145), and in E. coli, a model was proposed in which NsrR senses NO directly via a nitrosylation on its [2Fe-2S] cluster, resulting in the loss of the DNA-binding property of the transcription factor (145). Among the genes induced by NO in S. meliloti are those involved in the denitrification process, namely napEDABC, nirK, norCBQD, and nosRZDYFLX, and also hmp (80, 90). The requirement of the transcriptional regulator NnrR for nir and nor genes activation was also shown in B. japonicum (94). However, NnrR is placed under the control of the FixLJ-K2 regulatory cascade (Fig. 5) and responds either directly or indirectly to NO (94). These studies show similarities between Rhizobium species and others bacteria with the control by the NO-specific regulator NnrR. However, in S. meliloti, NnrR and FixLJ–FixK do not regulate the whole set of NO responsives genes, suggesting that other putative regulators remain to be identified (90).
Post-translational modifications
ROS are routinely synthesized in highly controlled and transient fashions, which constitute key regulators of redox signaling. Mild oxidants, such as H2O2 and NO, react with specific residues of target proteins that result in covalent protein modifications. The oxidation of target Cys residues can result in a number of distinct redox-based post-translational modifications, including S-nitrosylation, S-glutathionylation, S-sulfenylation, and disulfide formation, which result in the modification of protein function, localization, or turnover (72). Cys-based redox switches are now increasingly being found to underpin many different signaling systems and to regulate physiological outputs across kingdoms (135).
Using chemical and genetic probes along with mass spectrometry analyses, about a hundred of sulfenylated proteins were recently identified during the M. truncatula–S. meliloti symbiosis (102). Interestingly, redox state-linked proteins represent the largest functional group in the early interaction (2 days postinoculation), whereas primary metabolism-related proteins are the major ones in nodules (4 weeks postinoculation) (102). These results suggest that sulfenylation may regulate the activity of proteins playing major roles in the development and functioning of the symbiotic interaction (102).
In B. japonicum, the transcription factor Fix K2 was shown to be modified by reversible S-sulfenylation conferring different DNA-binding affinities and transcription activator activities (96). The fixK 2 gene is activated by the two-component regulator FixLJ in response to a moderate decrease of the oxygen pressure (Fig. 5), and the FixK2 protein amplifies this response to approximately 200 target genes (35, 95). Interestingly, a critical, single cysteine residue near the DNA-binding domain confers sensitivity to ROS in vitro as well as in vivo (96). Similarly, the highly conserved cysteine of the sensor RegS might be involved in controlling the redox inactivation of kinase activity as demonstrated for Rhodobacter capsulatus (139).
On the other hand, S-nitrosylation is emerging as a key redox-based post-translational modification in plants and a pivotal mechanism to convey NO bioactivity. Preliminary experiments carried out in our team with M. truncatula–S. meliloti mature nodules resulted in the identification of about 80 S-nitrosylated proteins, from both the plant and the bacterial partner, many of them being related to carbon and N2-fixing metabolism (Brouquisse et al., unpublished observations). Interestingly, 27 proteins were found to be both sulfenylated and nitrosylated (Table 2). It may be noted that 21 of them are related to carbon, nitrogen, and energy metabolism, which underlines the key role of the redox state in the control of primary metabolism. Many of the proteins identified as being S-sulfenylated or S-nitrosylated in the symbiotic interaction were also reported to be S-glutathionylated in Arabidopsis thaliana (34), suggesting that the same protein may be precisely and differently regulated depending on redox state and signaling (135).
Proteins commonly identified as sulfenylated (102) and nitrosylated (Brouquisse et al., unpublished observations).
Accession number and gene number according to the Mt3.5 database (
Data collected from
References are limited to plant and bacteria proteomics analyses focused on S-NO or S-OH modifications.
TCA, tricarboxylic acid cycle.
Overall, it appears that H2O2 and NO are playing crucial roles in the legume–Rhizobium symbiosis, in being involved at various steps of the interaction (Fig. 1). They are important players in the cross-talk between plant and bacterial partners during the establishment of symbiosis, and in the nodule functioning, where they modulate gene expression and protein activity in both partners (Fig. 7).
Reactive Species in Mycorrhizal Symbiosis
Arbuscular mycorrhizal fungi (AMF) represent a monophyletic fungal lineage (Glomeromycota) that benefits terrestrial ecosystems worldwide by establishing an intimate association with the roots of most land plants: the mycorrhizal symbiosis. This relationship results in an improved acquisition of nutrients (phosphate, nitrate) from the soil by the plant partners and, in exchange, allows the AMF to obtain the photosynthetically fixed carbon sources (sugars) which are necessary for their survival and propagation (25). Fungal lipochitooligosaccharide signals (Myc factors) stimulate the formation of arbuscular mycorrhiza in plant species of diverse families (82).
H2O2 was detected in infection structures during AM symbiosis (AMS) (41, 74, 126). Intracellular accumulation of H2O2 was observed in the plant cytoplasm close to intact and collapsing fungal structures, whereas intercellular H2O2 was located at the surface of fungal hyphae. These characteristics of ROS accumulation in AM roots suggest similarities to ROS accumulation during the senescence of legume root nodules (41). On the other hand, accumulation of H2O2 was especially pronounced in cells containing arbuscules and around hyphal tips attempting to penetrate a host cell. In contrast, no H2O2 accumulation was observed in hyphal tips growing along the middle lamella, or in appressoria or vesicles, suggesting that a locally restricted oxidative burst is involved in the temporal and spatial control of the intracellular colonization of M. truncatula cells by the AMF Glomus intraradices (126).
The expression of a CuZn SOD (GmarCuZnSOD) in the Gigaspora margarita has been shown to be associated with fungal structures inside the root where H2O2 has been also highlighted (74). It was suggested that this gene may offer protection as an ROS-inactivating system against localized host defense responses raised in arbuscule-containing cells (74). However, it may be also suggested that GmarCuZnSOD contributes to produce an H2O2 signal to optimize the interaction. The identification of a G. intraradices gene (GintPDX1) encoding a protein involved in vitamin B6 biosynthesis supports this hypothesis. Vitamin B6 has recently been implicated in defence against cellular oxidative stress (10). On oxidative stress, GintPDX1 is up-regulated in the intraradical fungal structures, suggesting a role as an antioxidant and a modulator of ROS in G. intraradices (10). This fits with the idea that a minimal H2O2 production is required for an optimal symbiotic interaction, as already shown in the rhizobial interaction (67).
H2O2 production during AMS raised the question of its origin. A partial answer can be suggested by the study of the mutualistic interaction between Epichloë festucae and Lolium perenne (140, 141). Regulated synthesis of ROS by specific fungal Noxs was shown to play a key role in fungal cellular differentiation and development (140, 141). Fungi have up to three different Nox isoforms, and ROS production by NoxA is critical for maintaining a fungal–plant symbiosis (142). Thus, even if the identification of AM Nox gene remains to be performed, a similar regulation may be suggested. Plant Rbohs genes may also be good candidates. Indeed, MtROP9i transgenic lines revealed neither ROS generation nor MtRBOHE/3 gene expression after AM infection (71). However, early hyphal root colonization is promoted, suggesting a putative role of MtRBOHE/3 in ROS-mediated early infection signaling (71).
While there is now strong evidence for NO production in NFS, less information exists for AMS. NSF and AMS share several common cellular and molecular features, referred to as the “common symbiotic pathway” under the control of 3 DMI (Doesn't make Infection) genes (105), and it was asked whether NO could be involved in AMS establishment and functioning as well. A recent report provides evidence of NO production in the roots of M. truncatula plant within the first minutes after treatment with G. margarita exudate (19). Using dmi mutants, the authors showed that NO production is involved in the signaling pathway mediating the early stage of AM interaction. Based on the activation of one NR gene before the establishment of the symbiosis between M. truncatula and Glomus mosseae, Weidmann et al. (150) suggested a possible role of NO in mycorrhiza. Recently, Moche et al. (97) showed that NO-producing capacity of tobacco root plasma membrane was strongly correlated with G. mosseae colonisation, and suggested that NO could be produced by the coordinated activity of plasma membrane NR and nitrite:NO reductase. This hypothesis was recently supported by the observation that, during M. truncatula-G. margarita interaction, a slight up-regulation of MtNR gene occurs within the first hours of the interaction, and the production of NO is reduced when the enzyme is inhibited by Tg (19). Thus, similar to what happens in NFS, NO is probably involved in both the establishment and functioning of AMS, but additional experiments are necessary to clearly decipher its specific functions in this interaction.
Future Directions
One of the most challenging questions is to define more precisely when and where ROS and RNS are generated during symbioses. As mentioned earlier, it appears that both H2O2 and NO are formed at various times during the symbiotic interaction, where they most probably play different roles. For instance, based on the observation that ROS are produced during the IT progression and the endocytosis process, the question is raised of their role in the release of bacteria from the IT and in their further differentiation into bacteroids.
Adapted tools to detect in vivo ROS production are needed. Thus, using a H2O2-specific fluorescent protein probe (9), we have been able to visualize different H2O2 levels in the infection zone of M. truncatula nodules (2). In vivo NO production was evidenced at various steps of the symbiotic interaction using DAF-2DA and DAF-FM fluorescent probes (6, 20, 31, 100, 132). However, DAF was shown to react with ascorbic and dehydroascorbic acids to generate fluorescent compounds interfering with DAF-2T (158), and its use in plant tissues was called into question (116). Recently, cell permeant fluorescent probes reacting directly with NO have been developed, such as copper-base fluorescent probe (78) and the fluorescent NO cheletropic traps (38). With sensitivity in the nM range and a good selectivity for NO, these probes appear ideally suited to detect NO in vivo during symbiosis. A technical point should also be underlined. Since N2-fixing nodules are microoxic organs, the protocols used to analyze either H2O2, or NO, production, metabolism, and function in symbiotic nodules should take this parameter into account and respect microoxic conditions to avoid biased results. Thus, the measurement of NO production in nodule crude extracts, or in sliced nodules, should be called into question if the oxygen pressure in the samples is not controlled during the experiment.
The systems responsible for ROS production also need further attention. With regard to H2O2, it appears that Noxs are good candidates, even if the involvement of other systems cannot be excluded. Indeed, the presence of a redox system identified in M. truncatula lipid rafts from plasmalemma raised the question of its function during the symbiotic interaction (43, 77). Thus, the use of Rboh mutants appears to be necessary to define more precisely the importance of these enzymes in the symbiotic interaction. In this context, it is worth noting that phylogenetic analysis of RBOH encoding genes in M. truncatula (83) highlighted the existence of genes pairs resulting from recent duplication (134). The persistence of these pairs, in spite of the selective pressure, points to their importance and to possible new functions in the nodulation process. In this framework, the study of double mutants under the control of inducible promoters will be of great interest. NO synthesis still remains an open issue. Data presented here highlight the involvement of NR and possible NOS-like enzymes. Three NR genes have been identified in the M. truncatula genome, with MtNR3 being specifically expressed during the two first weeks of symbiosis (Boscari et al., unpublished observations). Plant double and triple mutants will be useful to underpin their respective role in NO production during symbiosis. NOS-like activities have been evidenced in symbiotic nodules, but neither the gene nor the enzyme was evidenced in plants (14, 99). The discovery of a specific NOS-like enzyme is a promising challenge and should help in elucidating NO signaling and metabolism.
Another important question is “how the ROS message is perceived.” It is indeed unlikely that ROS regulate gene expression directly, and it is already established that post-translational modifications may modify the activity of proteins involved in the development and functioning of the nodule (102). In this framework, the search for redox-dependent transcription factors will be fruitful. Based on the data presented earlier, it is most likely that ROS interfere with NF signaling (Fig. 6). The recent finding that a gene encoding a symbiotic kinase is regulated by both H2O2 and NF strongly supports this hypothesis (2). Thus, it will be important to define where H2O2 and NO are involved in the NF signalization cascade; the same question should be addressed with regard to the Myc factor cascade.
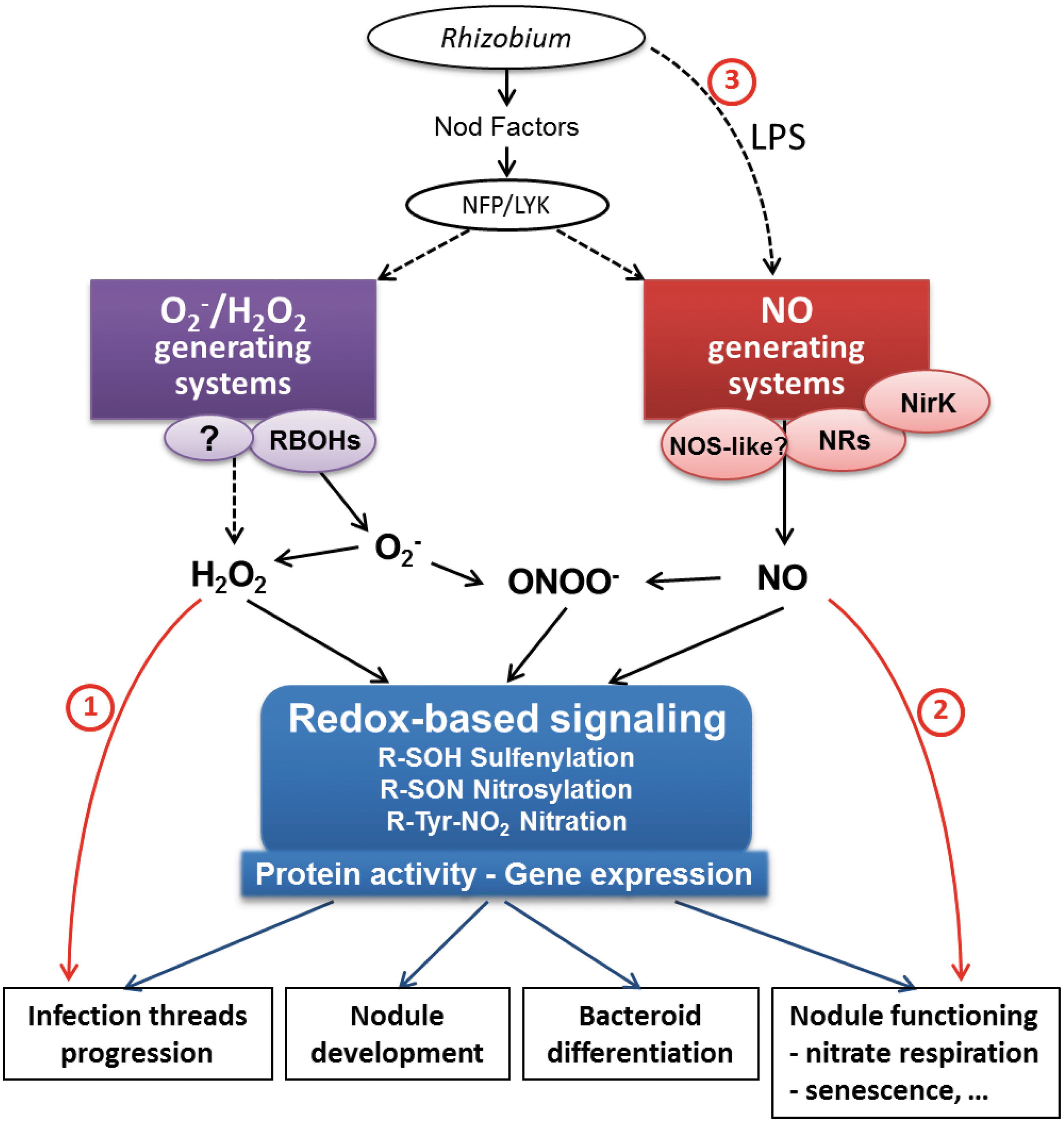
Crosstalk between ROS and NO appears to be another metabolic and signaling key to decipher symbiosis regulation. Crossed ROS/NO regulations have been already evidenced during plant-pathogen (5, 8) and pollen-stigma (60, 87) interactions. Possible crossed regulations in the legume symbioses are shown in Fig. 8. In terms of reactive species, peroxynitrite (ONOO − ) is an RNS that is formed when NO reacts with O2 − . To date, peroxynitrite is emerging as a potential signaling molecule in the induction of defense response against pathogens, and this could be mediated by the selective nitration of Tyr residues in a small number of proteins (147). Since both NO and O2 − are produced in symbiotic nodules, it is conceivable that peroxynitrite is formed in these organs. Lb was shown to scavenge peroxynitrite, thus precluding any damaging effect of this species in the nodules (58). Interestingly, Melo et al. (91) recently reported that glutamine synthetase GS1a, a key enzyme in N2 fixation (Fig. 2), is nitrated in nodules; whereas GS2a is subjected to S-nitrosylation. Such NO/ONOO− mediated post-translational inactivation of GS provides a direct link between NO/O2 − signaling and nitrogen metabolism in root nodules (91). Peroxynitrite signalization appears to be a promising field of investigation to understand redox regulation of NFS.
