
Abstract
Introduction
Innovation
Flavonoid-rich foods and beverages have been shown to exert a multiplicity of neuroprotective effects in the brain, whereas little is known regarding the actions of smaller phenolics/polyphenols. Our data provide the first evidence that phenolics such as gallic acid, protocatechuic acid, tyrosol, caftaric acid, and caffeic acid, delivered via moderate Champagne wine intake, also induce improvements in spatial memory in aged rodents and are dependent on the potential of these phenolics to modulate the expression of a number of proteins in the cortex and hippocampus related to the control of cell signaling, neuronal plasticity, cell cycle, and apoptosis.
Previously, Champagne wine consumption has been observed to improving acute vascular function (62), in a similar manner to that of red wine (7, 12). Further, tyrosol, caffeic acid, and gallic acid, the phenolic compounds found at relatively high concentrations in Champagne, have been shown to potently inhibit peroxynitrite-induced cellular injury at physiologically relevant concentrations (0.1 to 10 μM) (65), while nanomolar levels of tyrosol, caffeic acid, and p-coumaric acid protect cortical neurons against 5-S-cysteinyl-dopamine induced injury (61). Indeed, the levels of protection induced by these phenolics were equal to that, if not greater than, observed for a similar concentration of the flavonoids, (+)-catechin, (−)-epicatechin, and quercetin (61). The hydroxycinnamate, caffeic acid, has also been shown to be neuroprotective, counteracting inflammatory injury induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) by decreasing the production of a number of inflammatory cytokines, downregulating the expression of inducible nitric oxide synthase, cyclooxygenase-2, and glial fibrillary acidic protein and lowering the production of NO and PGE2 (56). In addition, caffeic acid–phenethyl ester may protect cerebellar granule neurons against glutamate-induced neuronal death via inhibition of p38 phosphorylation and caspase-3 activation (67) and significantly prevents hypoxic ischemic-induced neonatal rat brain damage in the cortex, hippocampus, and thalamus (68).
Despite these data, the impact of such phenolics on neuronal function/activity has received little attention relative to flavonoids, in particular in animal and human trials, and there is a lack of understanding with regard to their mechanisms of action in the brain in vivo. In particular, there are no data relating to the consumption of small phenolics and hydroxycinnamates, such as those present in Champagne wine on memory and neurocognitive performance. To address this, we have investigated the potential effects of moderate, medium-term Champagne wine intake on age-related deficits in spatial working memory and motor performance and the potential mechanisms of action in the hippocampus and cortex.
Results
Analysis of phenolic constituents in Champagne wine
The high performance liquid chromatography (HPLC) analysis of the phenolic extract obtained from Champagne wine allowed us to identify and quantify the major constituents present (Fig. 1). The chromatographic profile of the organic extract at 280 nm (Fig. 1A) and 320 nm (Fig. 1B) highlights the major phenolic components in the Champagne extract utilized: gallic acid (peak 1), protocatechuic acid (peak 2), tyrosol (peak 3), caftaric acid (peak 4), and caffeic acid (peak 5). Overlapping standard runs with the flavonoids catechin (Std 6), epicatechin (Std 7), and the stilbene resveratrol (Std 8) indicated that these polyphenols were undetectable in the Champagne used (Fig. 1). The major phenolics identified (Fig. 1C) were quantified to precisely characterize the intake on supplementation of the Champagne intervention (Fig. 1D).
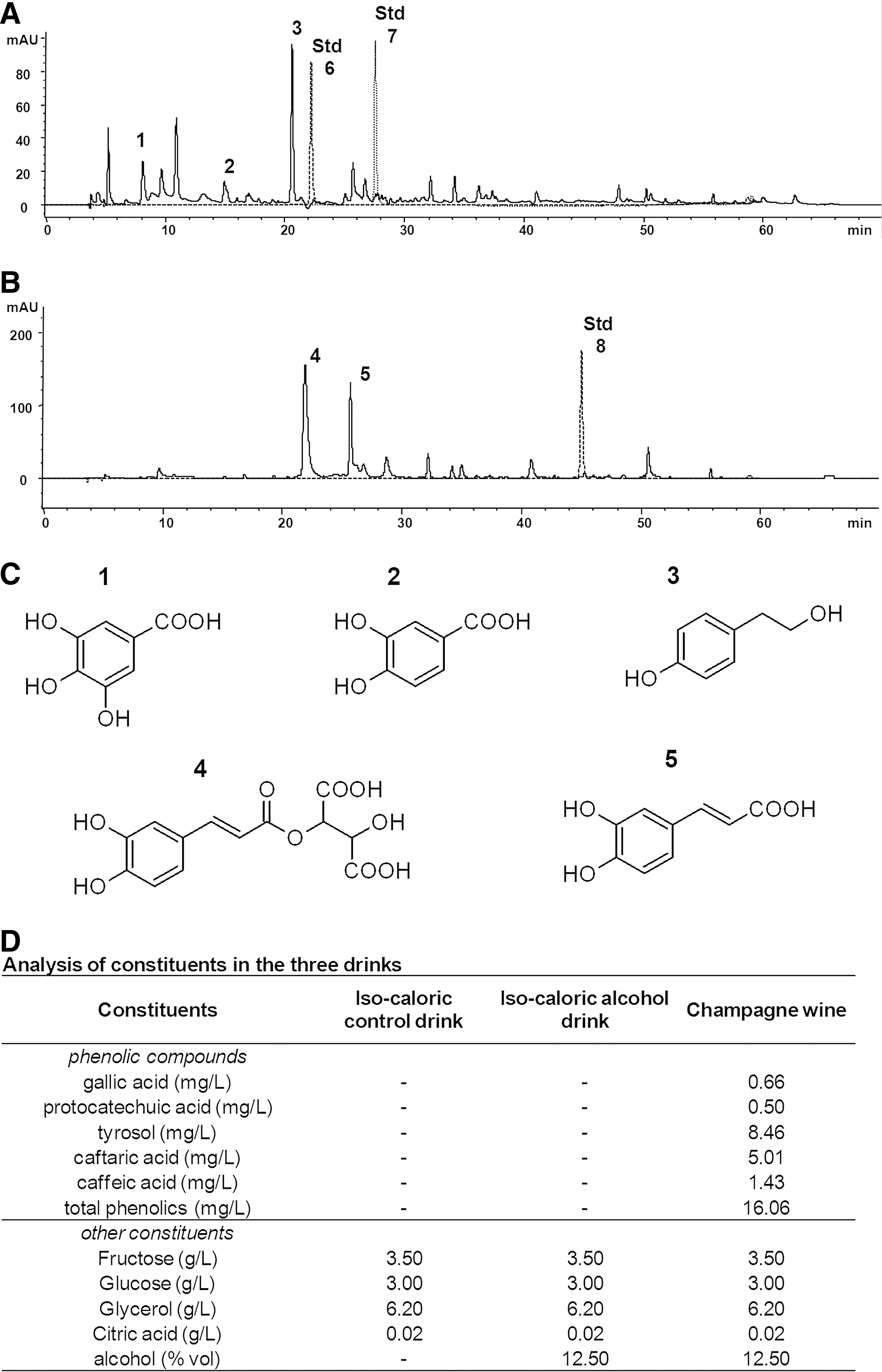
Food intake and animal weight
Animals were fed daily with Champagne, an isocaloric control or an isocaloric alcohol-containing control (Fig. 1D). There were no significant differences in food intake among the three groups throughout the study period (isocaloric control: 20.81 g/day; alcohol control: 20.42 g/day; Champagne group: 20.36 g; p>0.05). At baseline, the control, alcohol, and Champagne group animals weighed 478.11 (±18.23), 484.63 (±48.06), and 455.15 (±36.97) g, respectively, and there was no significant weight change throughout the intervention period in any of the groups (average weights at week 6: 480.70±15.75, 490.80±50.43, and 462.19±31.20 g, respectively; p>0.05).
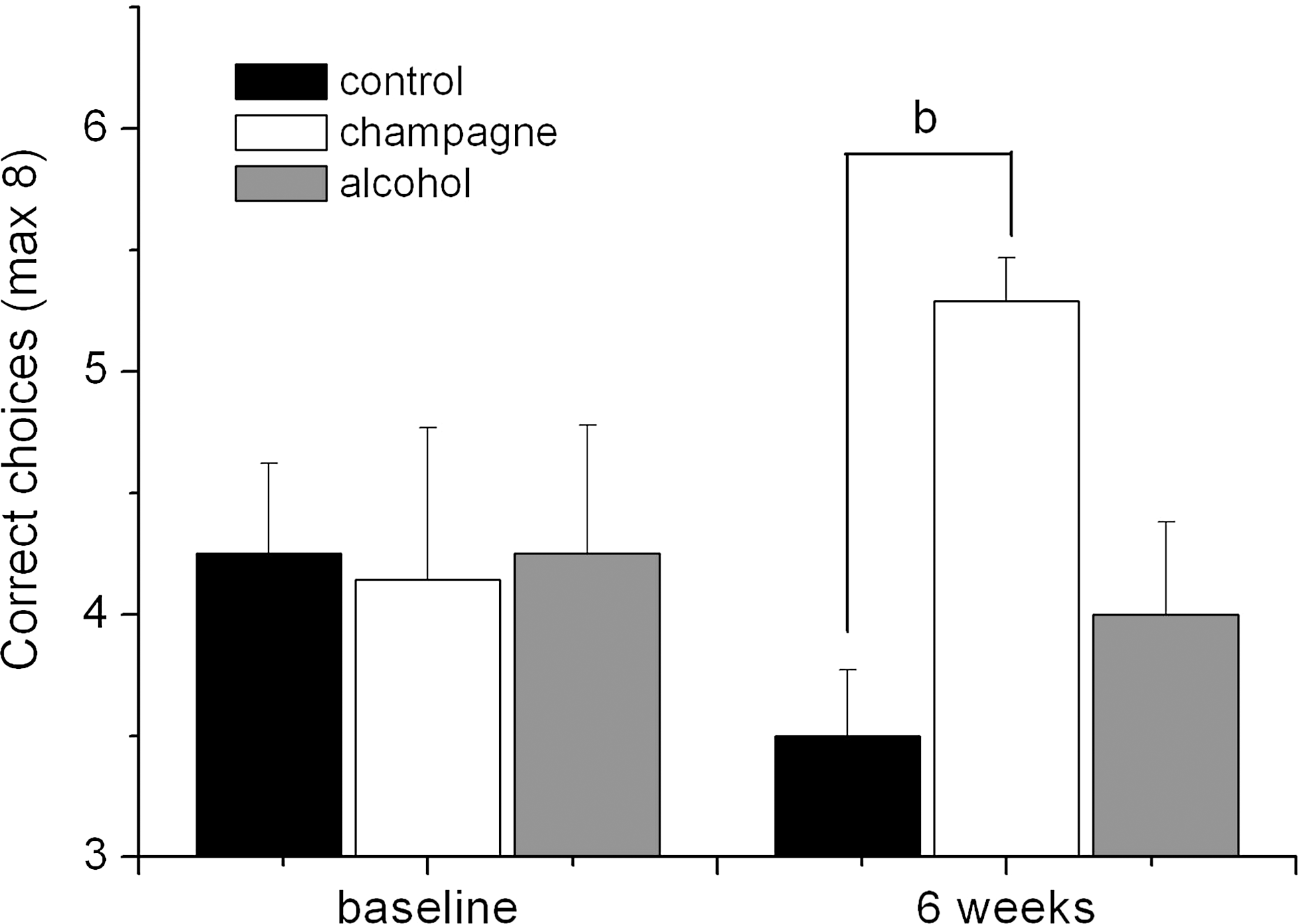
Spatial working memory
At baseline, the animals performed at an average choice accuracy of 4.25 (±0.27), and there was no significant difference in choice accuracy between the three experimental groups (Fig. 2). At 6 weeks, there was a reduction in the choice accuracy of the both isocaloric control group (3.50±0.27) and alcohol group (4.00±0.38) and an increase in the choice accuracy of the Champagne group (5.29±0.18). A two-way analysis of variance (ANOVA) with repeated measures indicated that there were no significant changes with respect to time (F(1,20)=0.022, MSE=0.026, p=0.884), or between treatments (F(2,20)=1.863, MSE=2.734, p=0.181), but there was a trend to significance with regard to the interaction between treatment and time (F(2,20)=2.955, MSE=3.536, p=0.075). Bonferroni post-hoc analysis indicated that this trend to significance was predominantly driven by a significant difference in choice accuracy between the isocaloric control group and the Champagne group at 6 weeks (p<0.01).
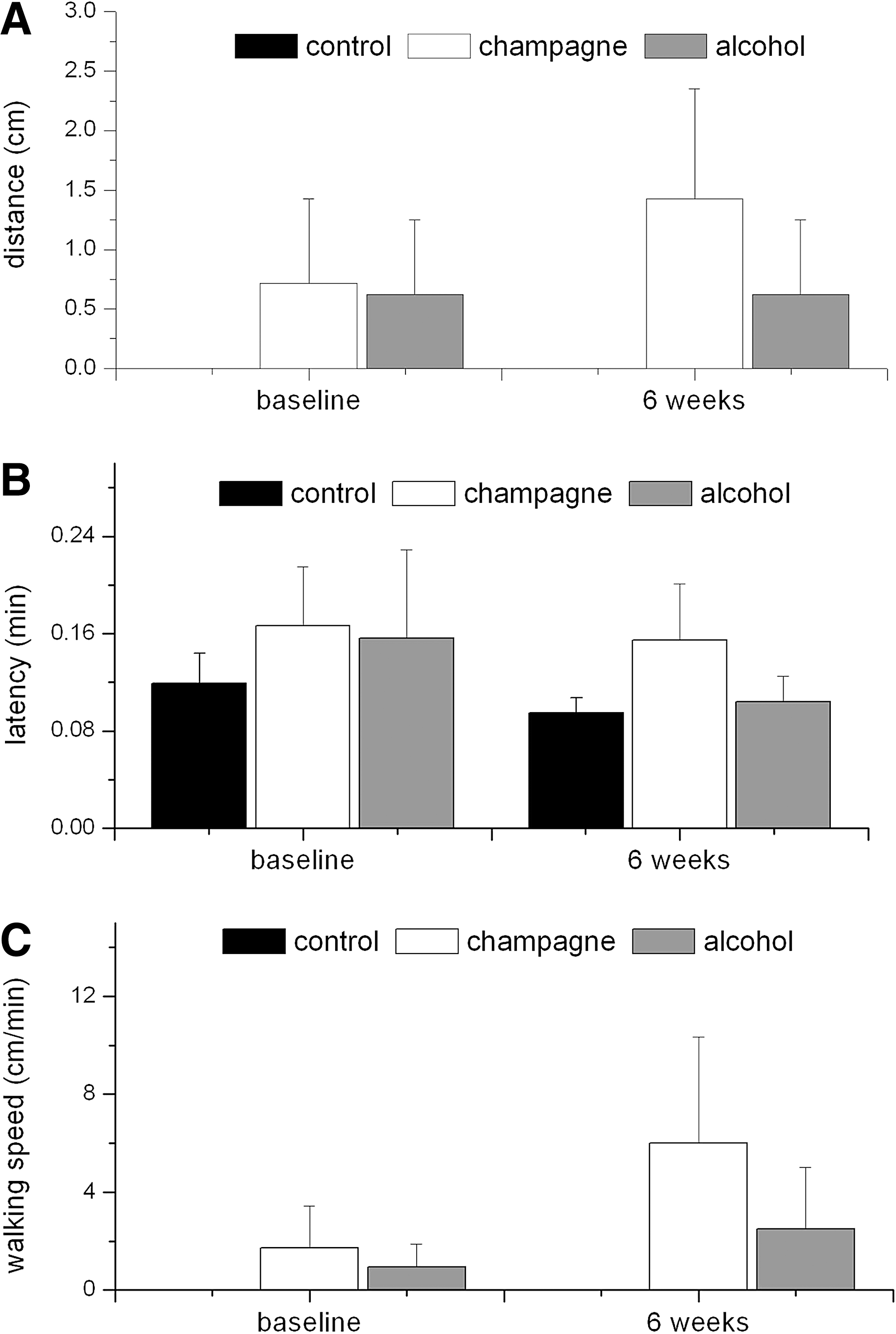
Motor performance
A two-way ANOVA with repeated measures indicated that there was no significant effect of the interventions on the distance covered by the animals on beam-walking tests (time: F(1,19)=1.101, MSE=0.621, p=0.307; treatment: F(2,19)=0.855, MSE=4.058, p=0.441; interaction: F(2,19)=1.080, MSE=0.609, p=0.360) (Fig. 3A). Similarly, the latency time (Fig. 3B) was not significantly affected by the control or Champagne intervention for 6 weeks (time: F(1,19)=1.703, MSE=33.783, p=0.208; treatment: F(2,19)=0.441, MSE=36.414, p=0.650; interaction: F(2,19)=0.293, MSE=5.814, p=0.749). In agreement with this, there were no significant effects of any of the treatments on calculated walking speed (Fig. 3C; time: F(1,19)=1.742, MSE=41.637, p=0.203; treatments (F(2,19)=1.155, MSE=52.296, p=0.336; interaction: F(2,19)=0.690, MSE=16.500, p=0.514). These data suggest that neither alcohol nor Champagne intervention had an influence on motor performance.
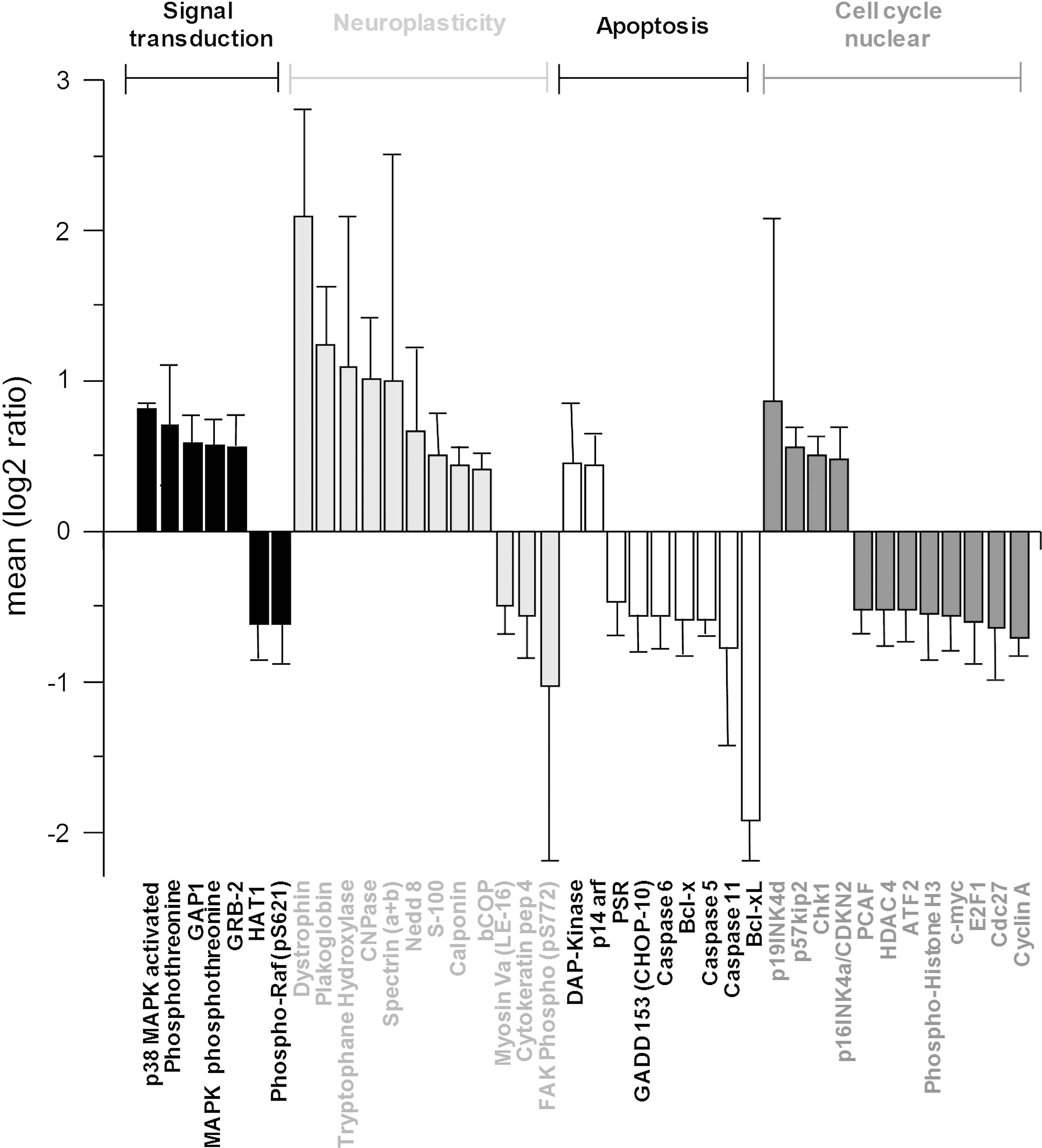
Protein expression array
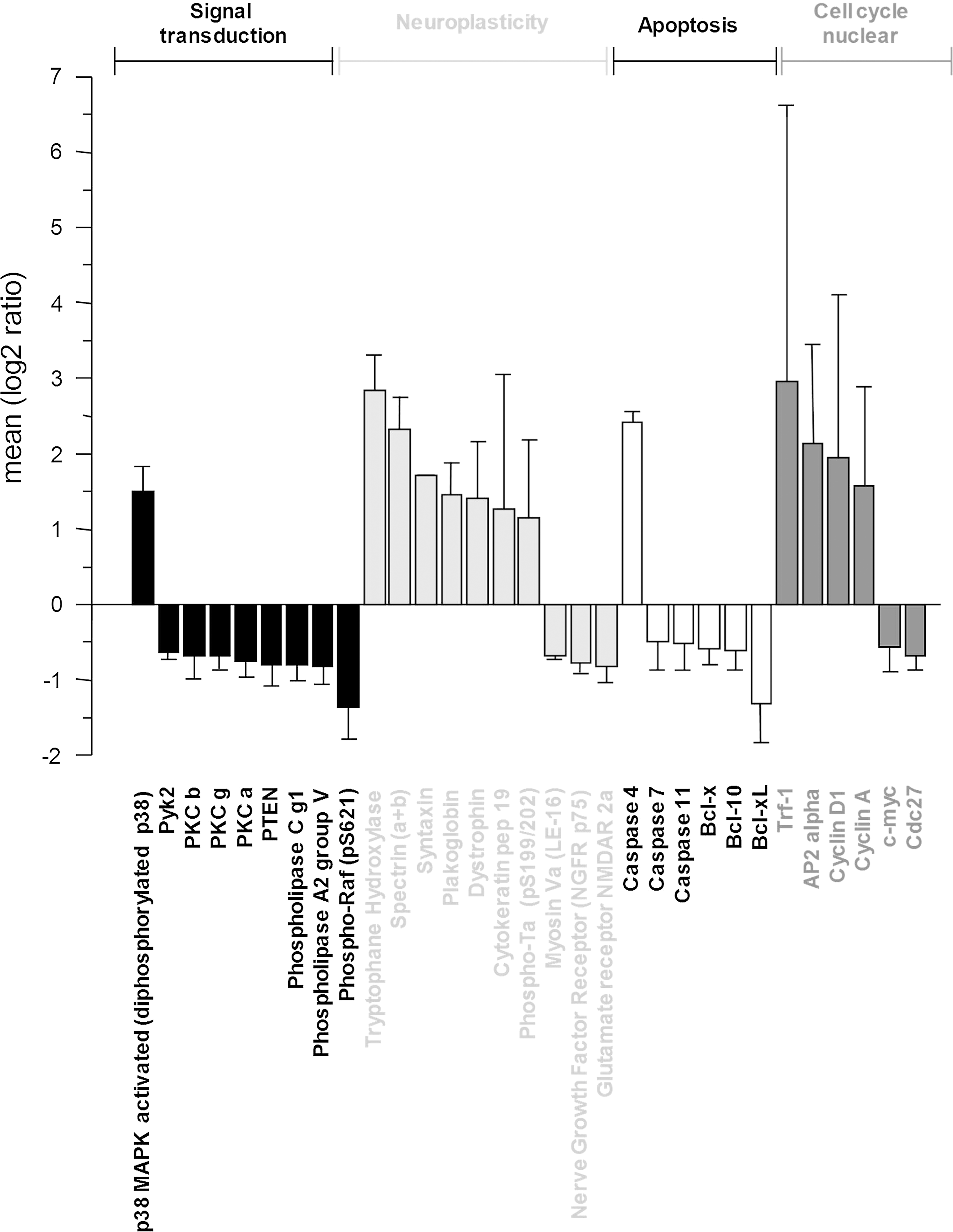
Forty hippocampal (Fig. 4) and 31 cortical (Fig. 5) proteins related to signal transduction, neuroplasticity, apoptosis, and cell cycle regulation were significantly modulated (p<0.05) after 6 weeks of Champagne intake relative to the isocaloric control. In both the hippocampus and cortex, Champagne intervention modulated a range of proteins involved in the mitogen-activated protein kinase (MAPK) signaling and the epidermal growth factor receptor signaling cascade, such as p38 and phospho-Raf. In addition, in the cortex, Champagne downregulated the expression of some calcium-activated protein kinases (PKC a, PKC b, and PKC g) and phospholipases C and A2. Champagne intervention increased the expression of the myelin-associated enzyme 2′,3′-cyclic-nucleotide 3′-phosphodiesterase (CNPase) in the hippocampus, and a range of neuroplasticity-related proteins in both the hippocampus and cortex (dystrophin, plakoglobin, tryptophane hydroxylase, and spectrin), whereas it downregulated NGFR p75 and NMDAR-2a receptors in the cortex only. Finally, Champagne supplementation also led to the modulation of apoptotic proteins (i.e., caspases and Bcl family proteins) and cell cycle/nuclear proteins, including those of the antiapoptotic proteins Bcl-xL and Bcl-x, and cyclin A and Cdc27 in both the hippocampus and cortex. Probe protein array analysis provided several new candidate proteins that were selected based on the expression levels and role in neuronal plasticity (p38, CNPase, dystrophin, and Bcl-xL), along with the previously identified proteins brain-derived neurotrophic factor (BDNF), pro-BDNF, cAMP response-element-binding protein (CREB), mammalian target of rapamycin (mTOR), Erk, JNK, Akt, Arc, and PKA C-α absent in the array, for further evaluation and/or confirmation by western immunoblotting. A complete list of the proteins analyzed with the protein array is presented as a supplementary material (Supplementary Table S1: hippocampus, and Supplementary Table S2: cortex; Supplementary Data are available online at
Immunoblotting
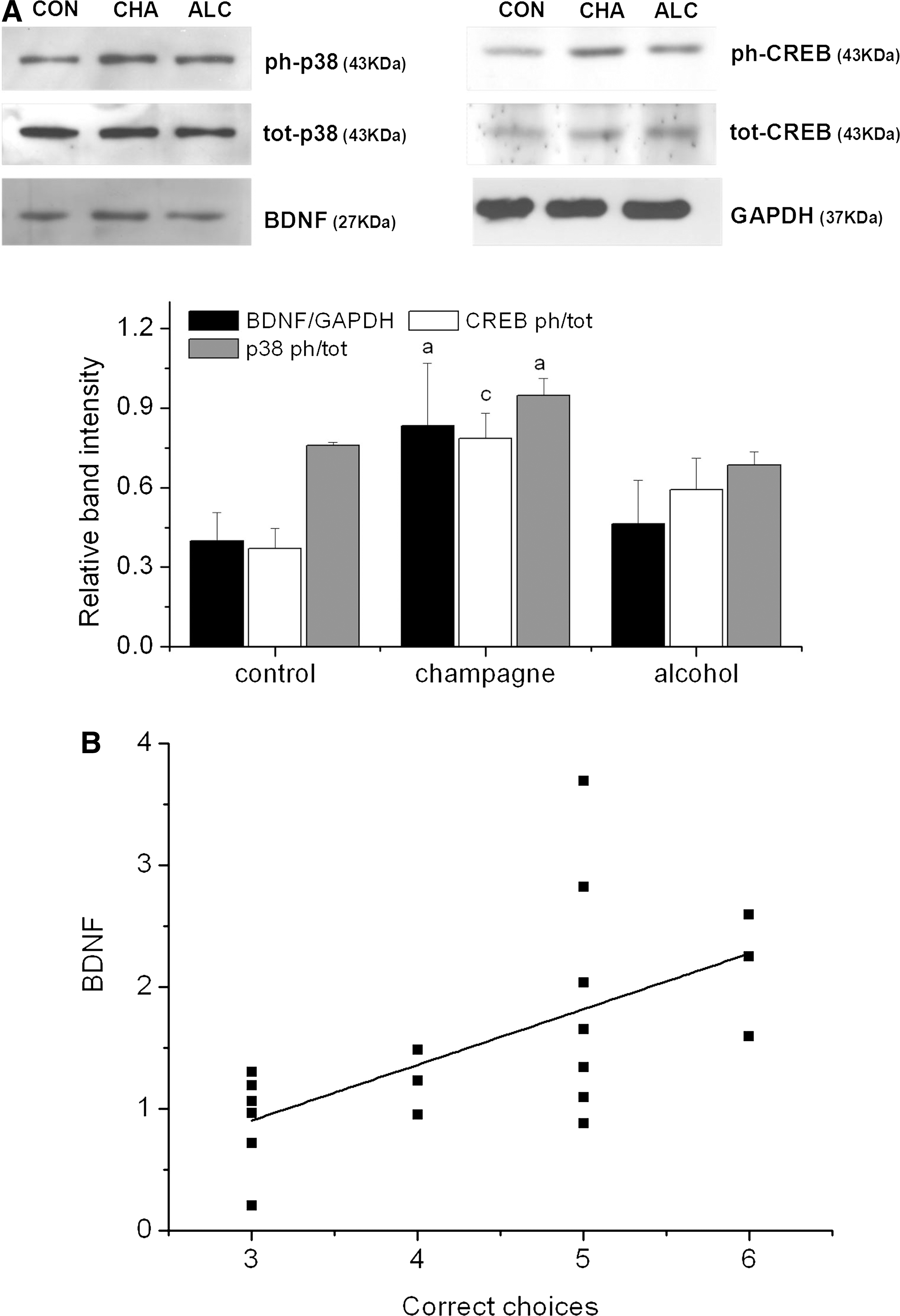
In comparison to the isocaloric and alcohol control interventions, Champagne intervention led to significant increases in the hippocampal levels of phospho-p38, pCREB (Ser133) and BDNF (p<0.001; p<0.05; and p<0.001, respectively) (Fig. 6A). In addition, there was a significant positive correlation between the hippocampal BDNF levels in individual animal performance on spatial memory tasks (R=0.613, p<0.01) (Fig. 6B) and between the hippocampal phospho-p38 levels and spatial performance (R=0.681, p<0.01). Similarly, Champagne intervention led to the increased expression of dystrophin and CNPase (p<0.05 and p<0.01, respectively) (Fig. 7), with a significant positive correlation existing between spatial memory task performance and hippocampal levels of dystrophin (R=0.74, p<0.05) and CNPase (R=0.529, p<0.05).

In contrast, mTOR phosphorylation and Bcl-xL were altered by both the Champagne and alcohol treatments (Fig. 8). Hippocampal levels of Bcl-xL were found to be significantly reduced in animals supplemented with both Champagne (p<0.001) and alcohol (p<0.01) compared to the control, although to a greater extent in the Champagne group (p<0.001). mTOR activation on the other hand was significantly higher in both the Champagne- (p<0.001) and alcohol-supplemented animals (p<0.05) compared to the isocaloric control. Champagne-induced hippocampal changes in the p38, CNPase, dystrophin, and Bcl-xL levels in the hippocampus were in agreement with those observed using antibody arrays (Fig. 4). There were no significant alterations in the activation state, or expression of, Akt, Arc, Erk, JNK, PKA C-α, and pro-BDNF in the hippocampus in response to either alcohol or Champagne intervention (Table 1).

Statistical analysis (one-way ANOVA) was performed. Differences were all nonsignificant (p>0.05).
With regard to protein expression in the cortex, the effect on mTOR was opposite to that observed in the hippocampus, with both alcohol (p<0.001) and Champagne (p<0.001) intervention leading to significant reductions in mTOR levels relative to the isocaloric control (Fig. 9). However, in a similar manner to that observed in the hippocampus, both alcohol and Champagne evoked significant increases in cortical phospho-CREB (p<0.05). There were no significant alterations in the activation state, or expression of, Akt, Arc, Bcl-xL, BDNF, CNPase, Erk, JNK, p38, and pro-BDNF in the cortex (Table 1).

Discussion
Human clinical trials and animal studies have identified flavonoid-rich foods and beverages as being capable of delaying the onset of age-related cognitive impairment (20, 21, 41, 48, 49, 63, 69). Flavonoids present in such foods have been postulated to evoke protection against cognitive aging through the actions of absorbed flavonoids and their metabolites on neurons and glia, enhancing the existing neuronal function and/or to stimulating neuronal regeneration (20, 26, 49, 63). Although antioxidant mechanisms cannot be excluded (70), recent datasets suggest that such effects are mediated by their ability to modulate neuronal signaling (48, 50, 52), stimulate neurotransmitter release (22), increase cerebrovascular blood flow (14), and even stimulate hippocampal neurogenesis (5). Although a large body of evidence has accumulated for flavonoid-rich foods (26), there is less known regarding the influence of nonflavonoid phenolics on similar behavioral and molecular parameters. In the current study, we show that Champagne wine, which is rich in phenolics, such as gallic acid, protocatechuic acid, tyrosol, caftaric acid and caffeic acid, but essentially lacking in flavonoids, is also capable of enhancing spatial working memory (without altering motor performance) in aged animals. In contrast, moderate alcohol intake failed to induce spatial memory changes. These observations are in agreement with those observed following long-term red wine intake in a similar model of hippocampal-dependent spatial memory (2).
The effects of Champagne on spatial memory were paralleled by a number of changes in the hippocampal and cortical protein expression, which may underpin the performance on spatial memory tasks. Perhaps, most notably, dystrophin and CNPase were modulated by Champagne supplementation, but not alcohol. CNPase (2′,3′-cyclic nucleotide 3′-phos phosphodiesterase) is a myelin-associated enzyme that constitutes around 4% of total central nervous system myelin protein, and is thought to undergo significant age-associated changes (17), and is reduced in Alzheimer's disease and Down's syndrome patients (66). Further, Champagne-induced hippocampal increases in the cytoskeletal associated protein, dystrophin, may be beneficial, as a lack of this protein in the hippocampus has been associated with impaired cognitive function (35), spatial memory (59), and long-term potentiation (58). Indeed, patients lacking dystrophin in the hippocampus and neocortex (due to mutation in the dystrophin gene) display a range of cognitive deficits (1). Intervention with the phenolic-rich Champagne also led to the increased expression of a range of other cytoskeletal proteins, including plakoglobin (γ-catenin), spectrin, calponin, cytokeratin pep4 and pep19, myosin Va, and focal adhesion kinase (FAK). Such proteins facilitate complex neuronal network formation in the brain and operate with neuronal membrane proteins (e.g., ion channels, scaffolding proteins, and adaptor proteins) at the sites of synaptic contacts to regulate synaptogenesis and coordinate synaptic strength (16, 38).
BDNF, a major regulator of synaptic transmission and plasticity at adult synapses (31), has previously been shown to partly mediate flavonoid-induced improvement in spatial memory through the ERK/Akt-CREB-BDNF pathway (42, 69). We show that phenolics present in Champagne also increase the BDNF and CREB levels in the hippocampus and a number of other neuroplastic proteins, notably dystrophin, plakoglobin, tryptophane hydroxylase, and CNPase. CREB is well known to be involved in memory and learning processes (53), via its selective activation of numerous downstream genes, including BDNF (3), and the CREB/BDNF pathway plays a critical role in synaptic plasticity and memory consolidation (34). In contrast to previous findings with flavonoid-rich interventions, we observed no significant modulation of ERK1/2 or Akt by Champagne intervention, suggesting that BDNF is not regulated in this manner. Rather, phenolic-induced BDNF expression may be regulated through p38 signaling in the hippocampus, leading to a decrease in the ERK/p38 activity ratio and the direct inhibition of raf-1 by p38 (18). Indeed, raf-1 was significantly downregulated by Champagne intervention, together with a number of other proteins connected to raf-1 in the hippocampus (p38, ATF2, GADD 153, p57kip2, Cdc27, Cyclin A, E2F1, PCAF, c-myc, GRB2, FAK, and spectrin) and in the cortex (p38, Cyclin A, Cyclin D1, c-myc, caspase 7, NGRF p75, PTEN, phospholipase C g1 and A2 group V, PKC a, b, and g, NMDAR 2a, Pyk2, and Tau), as indicated by STRING functional protein association networks (19). Further, the observed increases in p38 activation are in agreement with previous findings, showing that p38 activity may mediate BDNF synthesis in the rat hippocampus (23).
Further, unlike flavonoids, Champagne phenolics did not specifically regulate Akt-mTOR signaling, although alcohol treatment did appear to increase in the activation status of mTOR, a protein that regulates survival, differentiation, and development of neurons and is crucial for synaptic plasticity, learning, and memory formation in the adult brain (54), something which agrees with previous datasets (29). Similarly, reductions in the hippocampal levels of Bcl-xL levels were reduced by both Champagne and alcohol, suggesting that alcohol intake itself may play a role in apoptotic regulation in the brain (13). A number of other apoptotic proteins were found to be significantly downregulated by Champagne in the hippocampus (Bcl-xL, Bcl-x, caspases 5, 6, and 11, PSR, and GADD 153) and in the cortex (Bcl-xL, Bcl-x, Bcl-10, and caspases 7 and 11), although it is unclear whether alcohol alone would also mediate similar reductions. Antiapoptotic proteins such as Bcl-xL and other members of the Bcl-2 family have been implicated in the selective neuronal loss observed in AD (47) and have been found to be upregulated in the Alzheimer brains (24).
In summary, the potential of Champagne to beneficially influence spatial memory appears to be mediated by its ability to induce a number of changes in hippocampal and cortical protein expression, which occur in parallel to behavioral effects. Such effects on protein expression may result via the direct actions of individual phenolics such as tyrosol, caffeic acid, and their metabolites on hippocampal and cortical neurons/glia after their uptake into the brain, or via the synergistic effects of these phenolics along with other Champagne wine components. Although we did not observe this directly, previous data support the potential for such phenolics to undergo transfer across the blood–brain barrier (BBB) (11, 44). While the ability of flavonoids to cross the BBB is believed to be dependent on lipophilicity, small phenolics may transverse the BBB via amino acid transporters, such as have been reported for 4-ethylcatechol (33). Further, caffeic acid shares structural similarities with
Materials and Methods
Materials
Gallic acid, protocatechuic acid, caffeic acid, Bradford reagent, panorama antibody array Cell Signaling Kit, CelLytic™ MT Cell Lysis Reagent, and HRP-conjugated goat anti-rabbit and anti-mouse secondary antibodies were obtained from Sigma. Caftaric acid was obtained from Apin Chemicals. Tyrosol was purchased from Extrasynthese. PhosSTOP phosphatase inhibitors and Complete Miniprotease inhibitors were obtained from Roche. Precision Plus Protein Western C Standards and Precision Protein StrepTactin–HRP conjugate were purchased from Biorad. Primary antibodies used were anti-BDNF (1:200) and anti-pro-BDNF (1:200) from Millipore; anti-phospho-p38 Thr180/Tyr182 (1:1000), anti-total-p38 (1:1000), anti-phospho-CREB Ser133 (1:1000), anti-total-CREB (1:1000), anti-phospho-Erk1/2 Thr202/Tyr204 (1:2000), anti-total-Erk1/2 (1:2000), anti-phospho-Akt Ser473 (1:1000), anti-total-Akt (1:1000), anti-phospho-JNK Thr183/Tyr185 (1:1000), anti-total-JNK (1:1000), anti-Arc/Arg 3.1 C-7 (1:1000), anti-phospho-mTOR Ser2448 (1:1000), and anti-mTOR (1:1000) from New England Biolabs; anti-phospho-PKA C-α (1:1000), anti-total-PKA C-α (1:1000), anti CNPase (1:1000), anti-dystrophin (1:500), and anti-Bcl-xL (1:500) from Sigma. ECL reagent and hyperfilm-ECL were purchased from Amersham Biosciences. All other reagents were obtained from Sigma or Merck.
Extraction and quantification of polyphenols from Champagne wine
Champagne extracts were prepared using Champagne wine derived from both Chardonnay and Pinot Noir and Pinot Meunier grapes as reported previously (65). Briefly, 375 ml of Champagne was degassed, filtered, concentrated under vacuum, and extracted with ethyl acetate. The organic phase was separated from the aqueous phase and concentrated under reduced pressure to yield a brown aromatic residue (yield=1g/375 ml). The obtained fraction was dissolved in methanol, filtered, and analyzed by HPLC. HPLC analysis was carried out with an Agilent 1100 series liquid chromatograph equipped with a diode array detector linked to the HP ChemStation Software system. Samples were analyzed by reverse-phase HPLC using a NovaPak C18 column (4.6×250 mm, 4-μm particle size) fitted with a guard column C18 NovaPak from Waters. The temperature of the column was maintained at 30°C. The mobile phases consisted of a mixture of aqueous methanol 5% in 0.1% hydrochloric acid 5 M (A) and a mixture of aqueous acetonitrile 50% in 0.1% hydrochloric acid 5 M (B) and were pumped through the column at 0.7 ml/min. The following gradient system was used (min/% B): 0/5, 5/5, 40/50, 55/100, 59,9/100, and 60/5, with 10-min postrun. The eluent was monitored by photodiode array detection at 280 and 320 nm, and spectra of products were obtained over the 200–600-nm range. Calibration curves of the phenolic compounds were constructed using authentic standards (0.1–100 mg/ml), and in each case were found to be linear with correlation coefficients of .0.995. All data were analyzed using ChemStation software.
Animals and supplementation
All procedures were followed according to the specifications of the United Kingdom Animals (Scientific Procedures) Act, 1986, and to the national guidelines for the care and use of animals. Three groups of adult, male Wistar rats (n=8 per group; housed in cages each containing 2 rats) were kept under controlled temperature (22°C±1°C) and humidity (50%±10%), and on a 12:12 light–dark cycle (lights off 10:00 h). All rats were 15 months old at the beginning of the trial. Animals were pseudorandomly assigned to 3 groups based on their baseline spatial working memory correct-choice scores. Once allocated to a group, animals were fed daily with Champagne, an isocaloric control drink, or an isocaloric drink containing alcohol (Fig. 1D). The isocaloric control was matched for macro- and micronutrient content, notably glycerol, fructose, glucose, and citric acid content, and was dissolved in carbonated water. The alcohol control was similarly matched, but also contained alcohol (12.5% vol). The Champagne wine used in the study also contained phenolic compounds (gallic acid, protocatechuic acid, tyrosol, caftaric acid, and caffeic acid). Champagne and alcohol supplementation (1.78 ml/kg bw) was approximately equivalent to one 125-ml intake for a 70-Kg human and was adjusted according to each animal's body weight (approximate range 450–480 g). Alternatively, if we calculate the dose according to the Human Equivalent Dose (HED) formula: HED (mg/kg)=animal dose (mg/kg) * ([animal weight (kg)/human weight (kg)]0.33 (40), the Champagne dose utilized in our experiments is equivalent to 0.34 mg kg−1 day−1 in humans (23.8 mg Champagne in a 70-kg person per day). This equates to a lower intake equivalent to 1.3 glasses of Champagne (125 ml) per week in humans. To control daily intake of the intervention diets, the three drinks were prepared freshly each day and administered in the form of a mash, by mixing the test solutions with a small amount (8 g/10 ml) of powdered standard diet (RM3; Special Diets Services), and the correct amount given to each rat based on the body weight. At the end of the spatial memory trials, animals were sacrificed and the brains removed, the hippocampal and cortical regions dissected, snap-frozen in liquid nitrogen, and stored at −80°C until analysis.
Spatial working memory
Spatial working memory performance was assessed using a cross-maze apparatus (36), which is a commonly used paradigm to evaluate spatial learning, working memory, and reference memory performance in rodents (46, 69). All rats were fasted overnight before the procedure to provide sufficient motivation to ensure a high level of response during testing. Extra maze cues (laboratory furniture, lights, and several prominent visual features on the walls) were held constant throughout the experiment. Rats were given 2-week pretraining (habituation) that they ran reliably from the start arm to locate the reward pellets in both goal arms of the maze. This was followed by 6 weeks of shaping sessions (each consisting of six trials) before initiation of the supplementation regimes. During each shaping trial, rats were trained to enter an open goal arm and collect a reward pellet (entry to the alternate other goal arm was restricted). Across each shaping session, the open goal arm varied between trials according to a pseudorandom design. After completion of the shaping sessions, dietary supplementation and testing sessions were started. During the testing phase, each animal received eight trials per test session, with each consisting of a sample phase and a choice phase. During the sample phase, rats were placed in the start arm and allowed access only one goal arm to recover a food reward. Once the reward pellet had been collected, animals were removed and returned to the start arm for 10 s. During the choice phase, both goal arms were accessible, and animals were allowed a free choice between the two, with correct choice responses recorded for animals entering arm not visited during the sample phase. An animal was deemed to have selected an arm when it had placed a hind foot within the confines of the arm correct arm (retracing was measured as a failure on the task). Correct choices were rewarded with a food reward, whereas incorrect arm choices meant the animal was returned to its home cage. For 50% of the choice phases, the rat was placed in the alternate start arm opposite to the normal start arm. For each trial, accuracy and time taken to make a choice were measured. The maze was cleaned with 50% ethanol solution between trials to remove any olfactory clues. Each test session contained a pseudorandom sequence of correct choices between the two arms, as well as a pseudorandom sequence for the start arm during the choice phase. Rats were given 2 test sessions on the cross-maze, one session administered immediately before supplementation (baseline) and one administered at 6 weeks. Each group of aged animals was pseudorandomly assigned before the intervention based on the baseline correct-choice scores.
Stationary-beam test
The motor skills of the animals during Champagne supplementation were tested on a stationary beam (55) at baseline and at the end of the intervention (6 weeks). The dynamic equilibrium was tested on a beam consisting of a scale (150 cm, 3-cm diameter) located 40 cm above a soft carpet. At the beginning of the experiment, the rat was placed on the middle of the beam, its body axis perpendicular to the beam's longitudinal axis. The distance covered and latency times before falling were recorded, and the walking speed was calculated. The trial was stopped when the animal fell or 3 min had elapsed.
Antibody array
The Panorama antibody array Cell Signaling Kit (Sigma-Aldrich) was used, which contains 224 different antibodies spotted in duplicate on nitrocellulose glass slides, and represents proteins involved in biological pathways such as apoptosis, cell cycle, nuclear proteins, neurobiology, cytoskeleton, and signal transduction. Three biological replicates were used for the hippocampus region and two biological replicates for the cortex region. Statistical analysis was conducted on biological replicates, and significantly modulated protein expression in the hippocampus and in the cortex is reported in Supplementary Tables S1 and S2, respectively. Proteins from both the brain regions of the Champagne-treated and control-treated animals were extracted, purified, labeled with Cy-fluorescent dyes, hybridized to microarrays (at concentration of 5 μg/ml of each labeled target), and washed according to the protocols provided by the manufacturer. Briefly, the protein extracts were prepared using the extraction buffer provided in the kit, and the protein concentration was determined in the supernatant by the Bradford method. One mg of protein extract from each sample was labeled with Cy5 or Cy3, according to the manufacturer's instructions. Free, nonincorporated Cy3 and Cy5 dyes were separated by applying the labeled extracts on the SigmaSpin Post-Reaction Clean-Up Columns. The dye-to-protein molar ratio (D/P) was calculated, and only samples having D/P>2 were used. An equal amount of labeled protein of both extracts (5 μg/ml) was incubated on the microarray slide for 30 min; all washes were done in PBS-Tween 0.05%. The microarray slides were scanned on a GenePix 4200 Professional scanner (Molecular Devices), with laser power reduced to 25% and PMT settings manually adjusted to avoid saturated pixels in the experimental spots, and the resulted images were analyzed with GenePixPro 6 software (Molecular Devices). Spot intensities were local background-adjusted, and spots with negative values were removed from further analysis. For the remaining spots, resulted intensities were averaged for replicates, treatment-to-control ratios calculated, log2-transformed, and normalized to median.
Western immunoblotting
The dissected brain regions were homogenized on ice with a glass homogenizer using CelLytic™ MT Cell Lysis Reagent (Sigma) containing cOmplete Mini protease inhibitors and PhosSTOP phosphatase inhibitors. The homogenates were left on ice for 45 min before centrifugation at 4000 g for 5 min at 4°C to remove unbroken cell debris and nuclei. The protein concentration in the supernatants was determined by the Bradford protein assay. Samples were incubated for 2 min at 95°C in boiling buffer (final concentration 62.5 mM Tris, pH 6.8, 2% SDS, 5% 2-mercaptoethanol, 10% glycerol, and 0.0025% bromophenol blue). Boiled samples (20–60 μg/lane) and protein standards were run on 9%–12% SDS–polyacrylamide gels, and proteins were transferred to nitrocellulose membranes (Hybond-ECL; Amersham) by semidry electroblotting (1.5 mA/cm2). The nitrocellulose membrane was then incubated in a blocking buffer (20 mM Tris, pH 7.5, 150 mM NaCl; TBS) containing 4% (w/v) skimmed milk powder for 45 min at room temperature followed by 2×5-min washes in TBS supplemented with 0.05% (v/v) Tween 20 (TTBS). Blots were then incubated with the primary antibodies of interest (1:200–1:2000 dilution) in TTBS containing 1% (w/v) skimmed milk powder (antibody buffer) overnight at 4°C on a three-dimensional rocking table. The blots were washed 2×10 min in TTBS and incubated with secondary anti-rabbit or anti-mouse IgG conjugated to HRP (1:1000 dilution) and Precision Protein StrepTactin–HRP conjugate (1:10.000 dilution) for 45 min. Finally, the blots were washed 2×10 min in TTBS rinsed in TBS and exposed to ECL reagent for 1–2 min and developed. The bands were analyzed using Bio-Rad Quantity One 1-D Analysis software. Molecular weights of the bands were calculated from comparison with prestained molecular-weight markers that were run in parallel with the samples. Relative band intensities were calculated as a ratio of the phosphorylated protein to total protein in the case of CREB, p38, and mTOR. For BDNF, dystrophin, CNPase, and Bcl-xL, the relative band intensity was calculated in comparison to the GAPDH levels.
Statistics
All behavioral data (means±SEM, n=8) were subjected to two-way ANOVA for repeated measures with the diet group (control, alcohol, and Champagne) and time (0 and 6 weeks) as main factors, followed by a Bonferroni post hoc test to examine the differences between individual treatments (GraphPad Prism 5). For the microarray data (means±SEM, n=4 for hippocampus, n=4 for cortex), to evaluate the difference in protein expression between the Champagne-treated and isocaloric control-treated samples, we used Rank Product Test (4), as implemented in MeV 4.6 microarray data analysis software (43), with a cutoff of log2-ratio more than 0.4 or less than −0.4 (more than 1.3-fold change) and p<0.05. For the western immunoblotting data (means±SEM, n≥3), the results were statistically evaluated by one-way ANOVA, followed by a Student–Newman-Keuls post hoc t-test (GraphPad Prism 5). Significant changes are indicated as follows: a=p<0.001; b=p<0.01; c=p<0.05. Correlations between animal performance on spatial memory tasks and protein levels in the brain were calculated using the Pearson product moment correlation coefficient and the associated p value.
Footnotes
Acknowledgment
The authors wish to thank Dr. Viacheslav Bolshakov for his help with the protein array assays.
Author Disclosure Statement
No competing financial interests exist.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.