
Abstract
Direct recordings of electron currents mediated by cytochromes b561 (CYB561) are not available yet, despite the importance of these proteins in a variety of physiological functions, including neurotransmitter synthesis and dietary iron uptake. Here, we used the two-electrode voltage-clamp technique applied to Xenopus oocytes to demonstrate, for the first time, the generation of electron currents by a Drosophila member of the CYB561 superfamily named stromal cell-derived receptor 2 (SDR2). This experimental method, along with the theoretical development of a three-state kinetic model, supports the hypothesis that electron donor/acceptor concentrations and transmembrane voltage mutually control SDR2-mediated electron transport activity in a complex but predictable manner. Antioxid. Redox Signal. 21, 384–391.
Introduction
C
Stromal cell-derived receptor 2 (SDR2) contains a canonical CYB561 core domain and two additional extracellular domains named DOMON and reeler (1). Despite its proposed role in iron homeostasis in the brain (6), any further understanding of SDR2 biological activity is missing. In this study, we show that expression of Drosophila SDR2 in Xenopus oocytes allows a thorough characterization of this elusive redox system by the two-electrode voltage-clamp technique. SDR2 is the first transmembrane electron transporter ever analyzed by this powerful method.
We used a classical electrophysiological approach to demonstrate, for the first time, the generation of electron currents by stromal cell-derived receptor 2 (SDR2), a Drosophila member of the cytochrome b561 superfamily. The description of our experimental data with an extension of the Michaelis–Menten equation, derived from a simple and powerful kinetic model, sheds light on the complex relationships among the electron acceptor, electron donor, and membrane voltage in controlling SDR2 activity. Future investigation will confirm if this work represents a breakthrough in the field of protein-mediated electron transport.
Ferricyanide elicits inward currents in SDR2-expressing oocytes
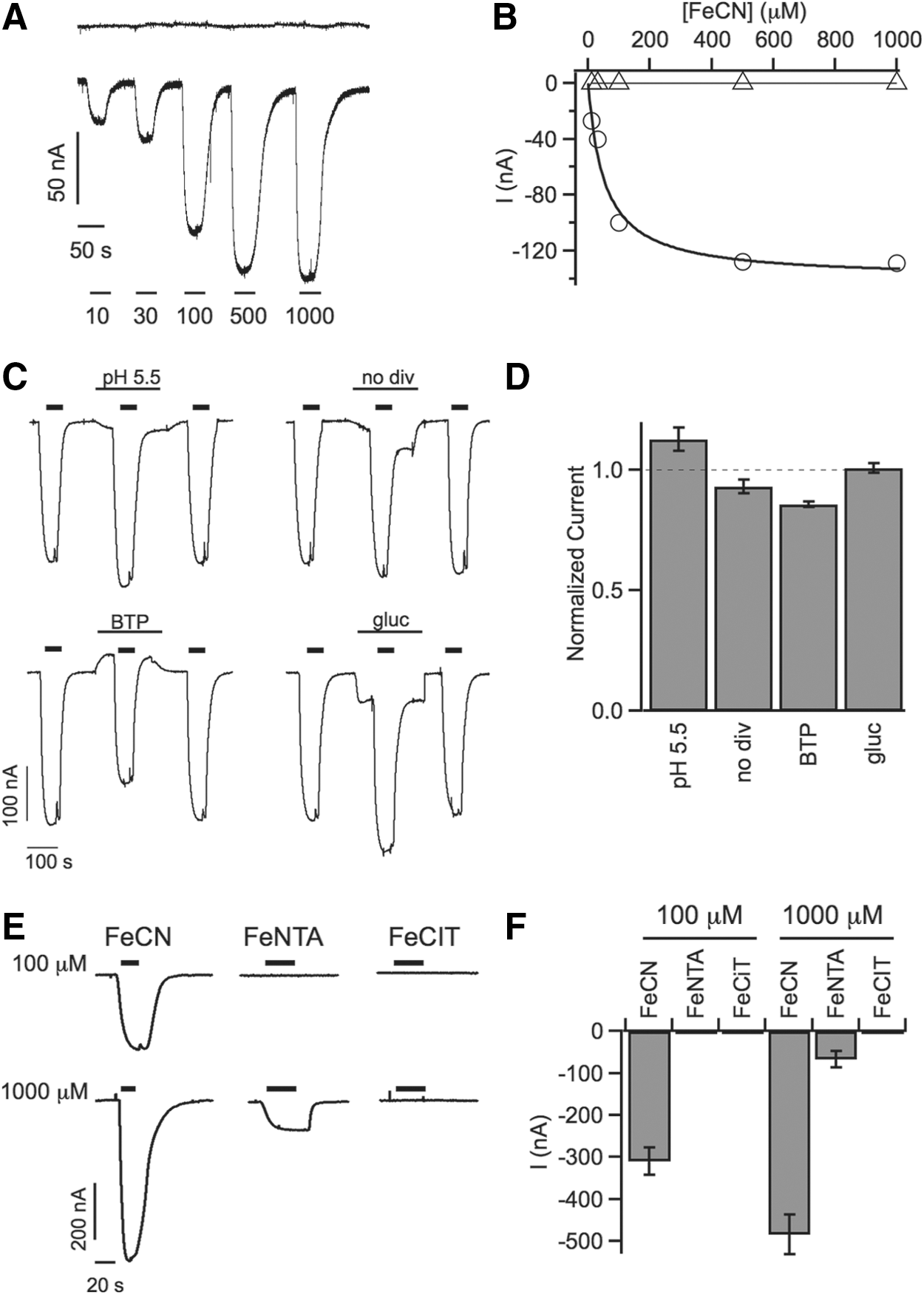
Preliminary evidence for transmembrane redox activity of SDR2 in the presence of external ferric chelate was obtained by colorimetric assays performed in heterologous expression systems like Xenopus oocytes (8). These results prompted us to verify the possibility to directly record currents in Xenopus oocytes applying the two-electrode voltage-clamp technique. In SDR2 cRNA-injected oocytes held at a membrane potential of −20 mV, brief exposure to the electron acceptor ferricyanide (FeCN) elicited small downward deflections of the current trace, indicative of inward current activation, which were completely reversible and increased in amplitude in a [FeCN]-dependent manner (Fig. 1A, lower trace). The data points could be described by a Michaelis–Menten function with a Km of 53 μM (Fig. 1B). By contrast, the current levels of control oocytes did not respond to FeCN exposure at any concentration (Fig. 1A, B).
Which is the nature of SDR2-dependent currents?
Inwardly directed membrane currents could potentially be due to either cation influx or anion efflux. As such, they are fully compatible with electrons moving from an intracellular electron donor to the extracellular electron acceptor FeCN. To exclude the activation of alternative ionic fluxes across the oocyte plasma membrane, we compared FeCN-induced current amplitudes in standard bath solution to those recorded in modified bath solutions (Fig. 1C, D): FeCN (100 μM) elicited current responses of similar amplitude at increased extracellular [H+] (pH 5.5), in a solution without the divalent cations Ca2+ and Mg2+ (no div), in a solution in which the monovalent cations Na+ and K+ were substituted by bis-tris-propane (BTP), or in which chloride was substituted by equimolar gluconate (gluc). These results argue strongly against the possibility that FeCN-induced inward currents were carried by the major ionic constituents of the oocyte system, leaving the possibility that they correspond to electron currents from an internal electron donor to external FeCN.
Inward currents depend on the type of electron acceptor
We next investigated the response of SDR2-expressing oocytes to alternative electron acceptors. Oocytes were successively challenged with FeCN and two ferric chelates, ferric nitrilotriacetate (FeNTA) and ferric citrate (FeCIT) in standard bath solution (Fig. 1E, F). FeNTA and FeCIT differed strongly from FeCN in their ability to promote transmembrane electron transfer. Whereas FeCIT failed to elicit any measurable current, both at 100 and 1000 μM, FeNTA caused the appearance of small inward currents only at 1000 μM, which was about 1/10 of the amplitude produced by FeCN at the same concentration (Fig. 1F). Again, the current trace recorded in control oocytes did not respond to the presence of extracellular electron acceptors. In the absence of better information about the physiological electron acceptors of this system, further experiments were conducted with FeCN, which allowed to measure currents of higher amplitude.
Modulation of electron currents by the cytosolic ascorbate concentration
Previous studies have established that electron transfer mediated by members of the CYB561-family is dependent on ascorbate as the electron donor. Consequently, we expected the FeCN-elicited inward currents of SDR2-expressing oocytes to respond to variations of the cytosolic ascorbate concentration. Membrane currents in response to [FeCN] between 10 and 1000 μM were recorded three times on the same oocyte (Fig. 2A): in baseline conditions using the endogenous cytosolic ascorbate pool (Asccyt; green traces), after the first ascorbate injection corresponding to Asccyt+10 mM (red traces) and after a further ascorbate injection raising the total [Asc] to Asccyt+40 mM (blue traces). Clearly, current amplitudes increased strongly after the first ascorbate injection, and even further after the second injection, lending additional support to the conclusion that FeCN-induced inward currents represent SDR2-mediated electron transfer from cytosolic ascorbate to extracellular FeCN.
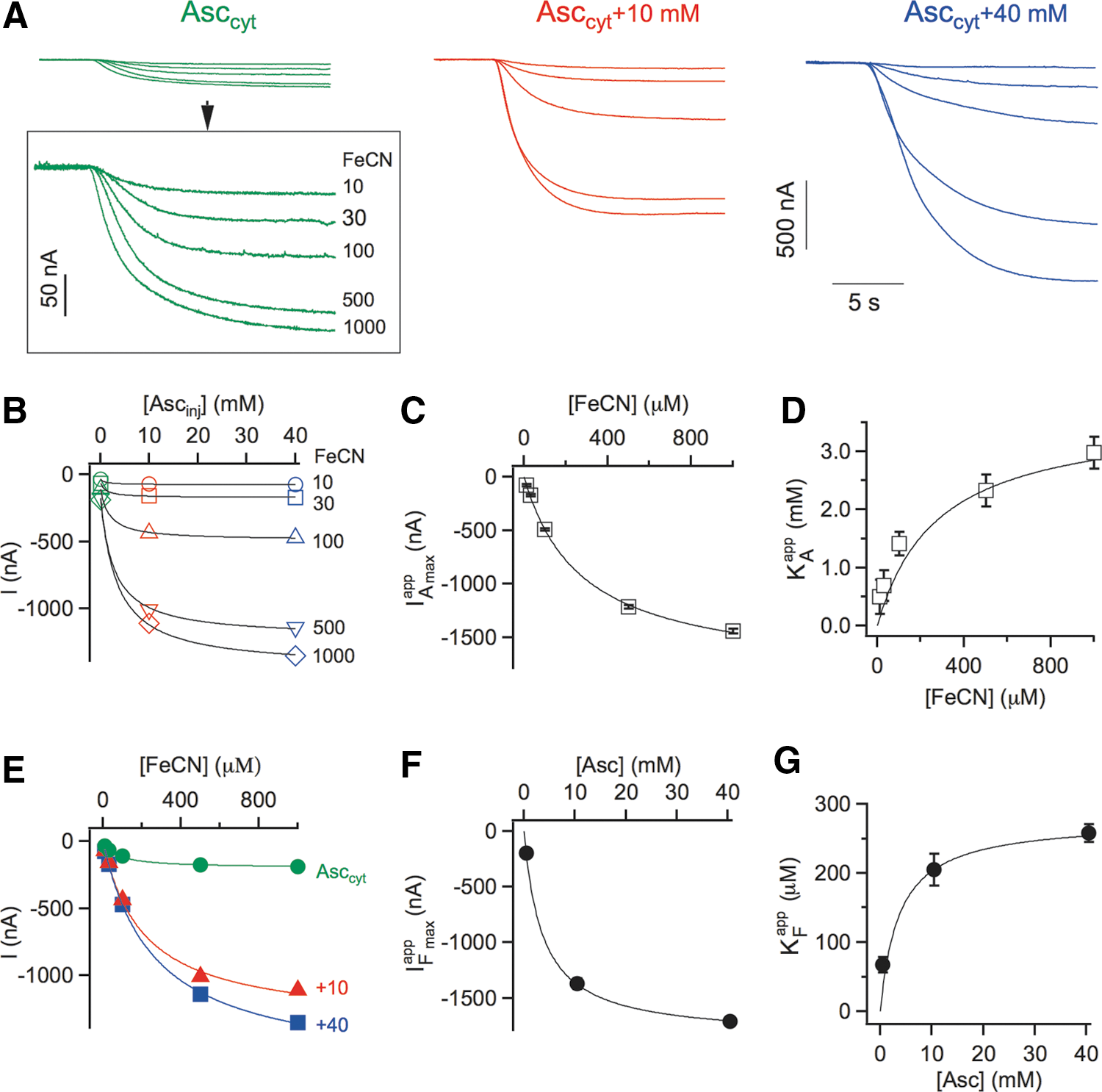
Current amplitudes obtained at five different [FeCN] were plotted as a function of injected ascorbate ([Asc]inj of 0, 10, and 40 mM; Fig. 2B). Data points were subjected to a global fit with a modified Michaelis–Menten function (see Notes section), which allowed the determination of the apparent maximum current at saturating [Asc] (IAmax app ; Fig. 2C), the apparent affinity constant for Asc (KA app ; Fig. 2D) and the concentration of the endogenous cytosolic ascorbate pool for each oocyte, Asccyt. This latter value of 0.60±0.11 mM was very consistent between individual oocytes (n=10) and is in line with the ascorbate concentrations typically found in animal cells, ranging from 0.5 mM in neutrophils to 10 mM in neurons. Current amplitudes were also plotted against FeCN concentrations (Fig. 2E) and fitted with a conventional Michaelis–Menten function yielding the apparent maximum currents (IFmax app ; Fig. 2F) and apparent affinity constants for FeCN (KF app ; Fig. 2G).
Secondary plots of apparent kinetic parameters versus substrate concentrations showed again hyperbolic dependencies, with apparent maximum currents that increased with increasing concentrations of the varied substrate (Fig. 2C, F), and apparent affinity constants that followed a similar trend (Fig. 2D, G). Thus, the lower the concentration of one substrate (either the donor Asc or the acceptor FeCN), the higher was the affinity for the other substrate. Overall, these data demonstrated that the electron transport activity of SDR2 strongly depends on the concentrations of both the electron donor (cytosolic ascorbate) and electron acceptor (extracellular FeCN) in a tightly connected manner.
Can SDR2-mediated currents be interpreted by a simple Michaelis–Menten equation?
A minimal three-state model was developed to describe SDR2-mediated electron transport (Fig. 3).
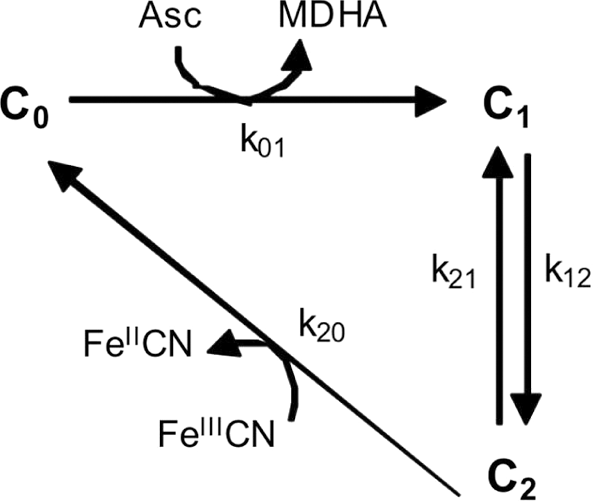
Upon binding of cytosolic ascorbate, free cytochrome in its oxidized form (C 0) is reduced by electron transfer to the internal heme (C 1) and semioxidized ascorbate (monodehydroascorbate [MDHA]) is concomitantly released (rate constant k 01). The rate constant k 12 describes the electronic transition from the internal to the external heme (C 2), the reverse reaction depending on rate constant k 21. The return from C 2 to C 0 (rate constant k 20) implies the reduction of extracellular ferricyanide (FeIIICN) and release of reduced ferrocyanide (FeIICN). Reverse reactions from C 1 to C 0 and from C 2 to C 0 are not included in the scheme because they depend on [MDHA] or [FeIICN], respectively, which were negligible in our experimental conditions. In fact, although SDR2 generated FeIICN and MDHA in the reaction cycle, FeIICN was continuously removed by perfusion, and intracellular MDHA was kept low by spontaneous dismutation and/or enzymatic reduction.
The scheme in Figure 3 is described by a simple steady-state Michaelis–Menten-like equation in which the current amplitude (I) is expressed as a function of substrate concentrations and four rate constants (k
01 to k
20) or, more conveniently, three limiting kinetic parameters with the usual operational meaning (Imax
, KA
, and KF
) in which rate constants can be merged (see Notes section).
This equation provided an adequate description of the data (continuous lines in Fig. 2C, D, F, G; see Notes section, for details): in spite of its minimal complexity, the scheme in Figure 3 and its related Equation 1 were thus powerful enough to provide a satisfactory interpretation of the SDR2 transport activity at a fixed membrane potential.
How does membrane voltage affect SDR2-mediated currents?
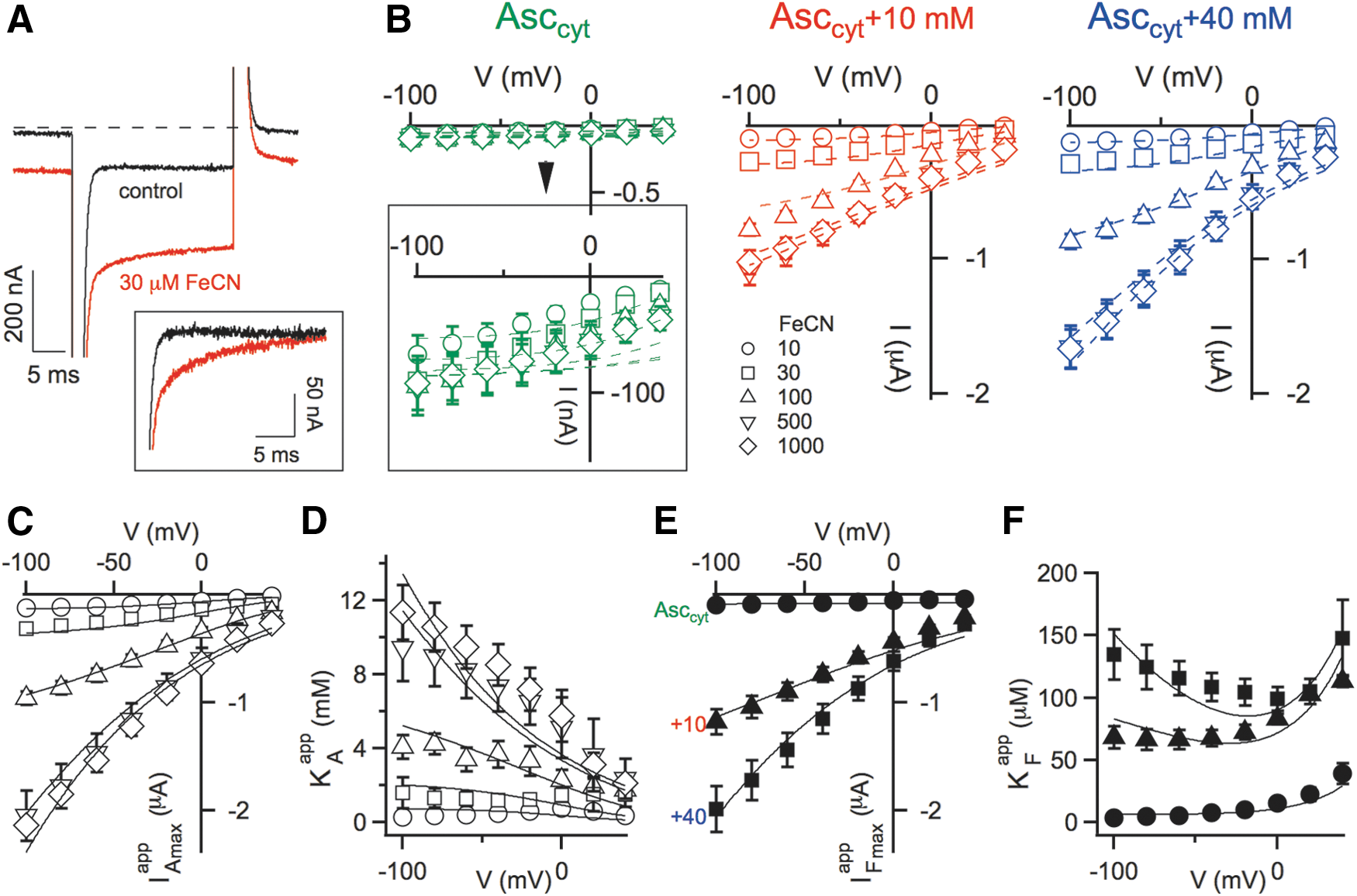
Cytochrome-mediated electron transport in the oocyte plasma membrane is supposed to be influenced by changes of the electrical potential difference across the membrane. We thus investigated the voltage dependence of FeCN-evoked membrane currents. In current recordings in response to a voltage step performed on SDR2-expressing oocytes, we observed a transient phase that was absent in control conditions, but appeared after addition of FeCN to the bath solution (Fig. 4A). In this study, we concentrated on the steady-state component of SDR2-mediated currents recorded in the voltage range between+40 and −100 mV (Fig. 4B). As expected, negative membrane potentials stimulated inward currents, but the activating effect was strongly dependent on the substrate concentrations (Fig. 4B). Using a similar strategy as in Figure 2, we obtained four apparent kinetic parameters (IAmax app , KA app , IFmax app , KF app ) and plotted them as a function of the applied membrane potential (Fig. 4C–F). At low concentrations of either Asc or FeCN, SDR2-mediated currents were almost voltage independent (Fig. 4B), and consequently, all apparent kinetic parameters (Fig. 4C–F). At high substrate concentrations, on the other hand, all parameters strongly increased with hyperpolarization and, except for KF app , all decreased at positive membrane potentials (Fig. 4B–E). The apparent affinity constant for FeCN (KF app ) behaved differently in that, it showed a minimum around 0 mV (Fig. 4F). Importantly, the complex voltage dependence of SDR2 was nicely described by scheme in Figure 3 extended by the assumption that the rate constants k 12 and k 21 were voltage dependent, that is, the transmembrane voltage (V) would have mainly influenced the electron transfer between the two hemes of the cytochrome. According to the theory of rate processes, we assigned to k 12 and k 21 exponential voltage dependence: k 12=k 12(0) exp(−δVF/RT); k 21=k 21(0) exp((1−δ)VF/RT)
where F, R, and T have conventional meanings, k 12(0) and k 21(0) are the voltage-independent components of the rate constants and δ represents the electrical distance between the cytosolic heme (i.e., the cytosolic electron binding site) and the energy maximum that the electron has to overcome to reach the external heme.
Although only the C 1 to C 2 interconversion was considered to be voltage dependent, the steady-state model of SDR2 predicts that all kinetic parameters must be affected by voltage, because all of them contain the rate constants k 12 and k 21 (see Notes section). Specifically, both Imax and KA were predicted to increase exponentially (in absolute values) with negative-going membrane potentials, since both parameters are linearly related to k 12. On the other hand, the voltage dependence of KF is predicted to be more complicated because it includes both k 12 and k 21, which are influenced by the membrane potential in opposite ways. As shown in Figure 4, the modified model satisfactorily accounted for the experimentally determined voltage dependence of SDR2, including the complex voltage responses of the apparent affinity constant for FeCN (Fig. 4F).
What about other electron transporters?
Before SDR2, the only example of a transmembrane electron transport protein studied by electrophysiological techniques was NADPH oxidase in human phagocytes, which is structurally unrelated to SDR2 (3, 5, 7). However, a comprehensive kinetic model for NADPH oxidase activity could not be drawn because information on its oxygen dependence was missing. The experimental setup used here allowed to perform a complete steady-state kinetic analysis of SDR2 activity and to include the effects of the membrane potential into a single integrated model. Future efforts will be directed at the application of this novel approach to further members of the CYB561 superfamily and other electronic transporters.
Notes
Materials and methods
Oocyte expression
Full-length Drosophila melanogaster SDR2 cDNA in the pOT2 vector was obtained from the Berkeley Drosophila Genome Project. The clone identity was confirmed by sequencing. In vitro transcription of XhoI-linearized SDR2 DNA template was done using the mMESSAGE mMACHINE T7 kit (Life Technologies, Monza, Italy) and the quality of the transcribed cRNA was confirmed by agarose gel analysis.
Oocytes were prepared from adult female Xenopus laevis frogs and injected with SDR2 cRNA at 35 ng per oocyte using a Drummond “Nanoject” microinjector. Oocytes were incubated in a solution containing 96 mM NaCl, 2 mM KCl, 1 mM CaCl2, 1 mM MgCl2, and 5 mM HEPES, pH 7.5 (solution A), supplemented with 0.1 mg/ml gentamycin. Oocytes were incubated at 18°C for at least 24 h before voltage-clamp experiments.
Electrophysiology
Whole-cell membrane currents were measured at room temperature using a home-made two-electrode voltage-clamp amplifier and 0.2–0.4 MΩ glass electrodes filled with 3 M KCl. The standard bath solution was solution A. A gravity-driven perfusion system was used for continuous superfusion of oocytes during voltage-clamp recordings and for switching between different bath solutions. Ferric compounds were prepared as stock solutions and diluted appropriately in a bath solution, potassium FeCN at 100 mM, FeNTA and FeCIT at 30 mM.
For ascorbate injection experiments, the ascorbate solution (100 mM stock) was made freshly and adjusted to pH 7.0 with KOH. After the initial recording series, the oocyte was removed from the voltage-clamp setup and injected with a defined volume of ascorbate stock solution using the Drummond “Nanoject” microinjector. The first injection of 50 nl stock solution and the second injection of 150 nl resulted in a final [Asc]inj of 10 and 40 mM, respectively, considering a volume of ∼500 nl for a mean oocyte diameter of 1 mm. After each injection, the oocyte volume transiently increased, but recovered to its original size within minutes. Injected oocytes were allowed to recover for 30 min at RT before voltage-clamp measurements were resumed.
Each experimental condition was tested in at least five different oocytes. The presence of extracellular electron acceptors elicited membrane currents in all SDR2 cRNA-injected oocytes tested (n=71). None of the control oocytes (n=41) subjected to the different experimental paradigms, including double ascorbate injection, showed detectable currents.
Kinetic model
In Figure 3, C
0 represents the probability of the cytochrome to be in its oxidized form. C
1 and C
2 are the probabilities of the cytochrome to be in a reduced form with an electron on the internal or on the external heme, respectively. The expression of the electronic current derived from Figure 3 is
where N is the total number of cytochromes on the oocyte membrane, and e is the electronic charge (−1.60×10−19 C). Solving the system of differential equations associated with the scheme in Figure 3 in steady-state conditions, we obtain the following:
where A and F are the internal ascorbate and the external FeCN concentrations, respectively. We can write Equation M2 in the form of Equation 1 (see main text) introducing the following kinetic parameters:
We assume k
12 and k
21 to be voltage dependent; thus, all three kinetic parameters have voltage dependence. At fixed FeCN concentrations and voltage, we can write the current as follows:
where
The two apparent kinetic parameters
where
Also, in this case,
Data analysis
In panel B of Figure 2, the following Michaelis–Menten function was used to perform global data fitting:
These parameters are linked to the rate constants as follows:
Imax (0), KA 1, KF 1, and KF 2 are, together with the electrical distance δ, the five free parameters of the model in this experimental condition. Imax (0) is the maximum current at V=0 mV when F >> KF and A >> KA ; KA 1 is the affinity constant for ascorbate at V=0 mV and F >> KF ; KF 1 is the affinity constant for FeCN at V<< 0 and A >> KA and KF 2 is the affinity constant for FeCN at V >> 0 and A >> KA . Inserting the relationships M10 in M5, M6, M8, and M9 and using these four equations to globally fit data in panel C, D, E, and F, we obtained Imax (0)=718±15 nA, KA 1=4.0±2.0 mM, KF 1=52±2 μM, KF 2=48±3 μM, and δ=0.35±0.01.
In the following, we focus on the voltage dependence of the current in limiting conditions. If A >> KA
and F >> KF
, then the theoretical current becomes
Footnotes
Acknowledgments
We thank Franco Gambale, Michael Pusch (IBF-CNR, Italy), and Paolo Pupillo (University of Bologna, Italy) for critical comments on the article. The technical assistance of Francesca Quartino, Alessandro Barbin, Damiano Magliozzi (IBF-CNR, Italy), and Nunzio Tango (University of Bologna) was highly appreciated. A.C. and F.S. were supported by the Italian “Progetti di Ricerca di Interesse Nazionale” (