
Abstract
Introduction
R

In addition to being an intrinsic part of metabolism, redox status plays an active role in metabolic regulation. In this regard, the redox status operates as a major integrator of cellular metabolism and is simultaneously regulated itself by metabolic processes. This enables readjustment of global metabolic pathways and redox status homeostasis in response to changes in environmental conditions, involving reprogramming of gene expression and post-translational modification of target enzymes by thiol-disulfide modulations (19, 21, 44, 177). The underlying signaling pathways have only partially been discovered in the previous years. While much is known about redox status signals involved in light activation of photosynthetic processes, little is known about redox regulation of other metabolic pathways in the plastid and of extra-plastidial metabolism. While recent studies provide evidence for the existence of redox signals coordinating metabolism and gene expression between different organelles, such as plastid, mitochondrion, and nucleus, their nature has not yet been clarified.
In this review, we will describe the redox status control of metabolism and the metabolic control of redox status at both the cellular and subcellular levels, mainly focusing on post-translational mechanisms. Despite the vastness of literature concerned with redox status-regulated gene expression, we will only describe this in passing given that it is the subject of a couple of excellent recent reviews (114, 309). In the first part, redox status-related metabolic processes will be described within their subcellular context, with regard to redox status-regulatory properties and intra-organellar signals involved in their co-ordination. In this regard, our major focus is placed on organelles such as plastids, mitochondria, and peroxisomes with readers being referred to other comprehensive reviews for details on both cytosolic and apoplastic aspects of redox status (96, 265, 270) with only a broad overview of the most important features in the context of cellular metabolism and function being provided here. In the second part, we will discuss the integration at the cellular level while mainly focusing on inter-organellar signals co-ordinating redox status regulation of metabolism between different subcellular compartments.
Plastidial Redox Status Biology
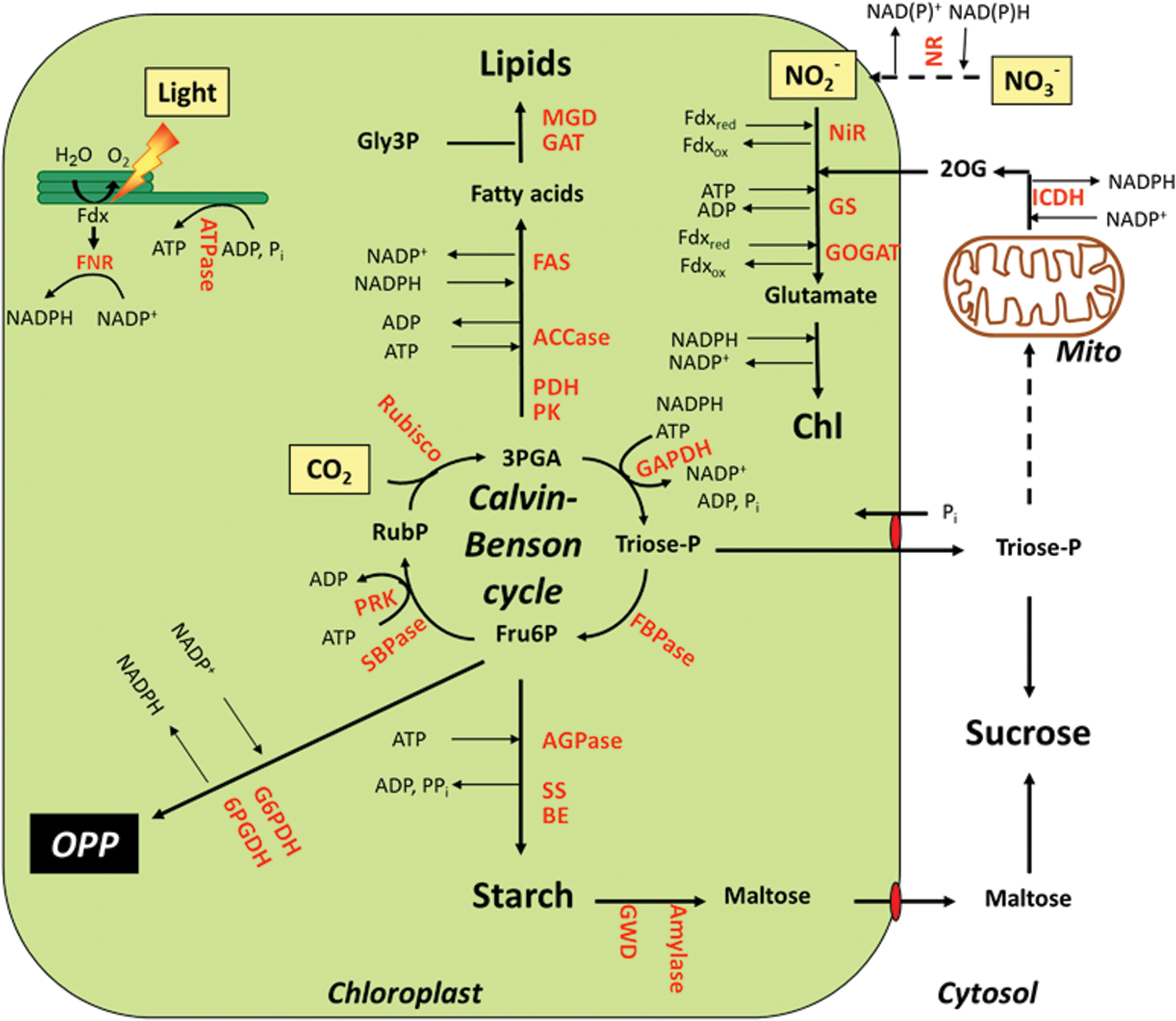
Chloroplasts are plant-specific organelles with important properties, the most prominent being their ability to carry out oxygenic photosynthesis (51). During this process, light energy is absorbed by photosystems I (PS I) and II (PS II) located in the thylakoid membrane and used to activate photosynthetic electron transport (Fig. 1). Linear electron flow requires both photosystems working in series, leading to electron transfer from water to NADP+ to generate NADPH as a reducing power and a trans-thylakoid proton gradient that drives ATP synthesis via CF0F1ATPase. This involves electron transfer from PS II to PS I via plastoquinone (PQ), the cytochrome b6 f complex, and plastocyanin as additional redox carriers (Fig. 1). At the stromal side of PS I, electrons are subsequently donated to Fdx, which functions as a mobile electron carrier distributing electrons to NADP+ via Fdx-NADP-reductase (FNR) to produce NADPH or directly to specific processes located in the stroma, such as S and N assimilation, the synthesis of chlorophyll and fatty acids, and reactions involved in chloroplast redox regulation (128).
In the latter, electrons are transferred from Fdx to thioredoxins (Trxs) via Fdx-Trx-reductase (FTR). Trxs are small regulatory proteins containing a redox-active disulfide group that controls the thiol-disulfide exchange of target proteins (145, 237, 280, 307). In plants, Trxs comprise a medium-sized gene family with 10 different isoforms (f1–2, m1–4, x, y1–2 and z) being located in the chloroplast of Arabidopsis (70), while other isoforms are located in the cytosol and mitochondria (219). In vitro studies using purified proteins indicate Trxs f and m to be involved in the regulation of stromal metabolism, while x-, y-, and z-types serve as reducing substrates for antioxidant enzymes (61) (Table 1). More recently, genetic studies have been used to further dissect the specific roles of different Trxs in vivo, providing evidence for different isoforms of Trxs f and m having different functions in plants (34, 346, 375).
For Abbreviations, see Abbreviations Used section.
In Arabidopsis, FNR has been found to be associated with the thylakoid and inner chloroplast envelope membranes, in addition to its location in the soluble stroma (128). Two FNR-binding proteins have been previously identified, Tic62 a component of the chloroplast import machinery (36) and the intrinsic thylakoid protein TROL (thylakoid rhodanese-like protein) (163), leading to binding of FNR to envelope and thylakoid membranes, respectively. Membrane recruiting of FNR has been found to be redox regulated in response to light signals (36). The role of membrane binding in the activity of FNR and the distribution of electrons to stromal reactions has, however, not yet been fully resolved. Recently, a combined knockout of Tic62 and TROL in an Arabidopsis double mutant led to restriction of FNR to the soluble stroma, resulting in decreased FNR level, decreased NADPH/NADP+ ratio, and altered starch metabolism in leaves (204).
In contrast to linear electron transfer, cyclic electron flow is driven by PS I alone to produce ATP without generating NADPH (Fig. 1). Here, electrons from Fdx are reintroduced into the electron transport chain rather than being distributed to stromal components. Two distinct pathways of cyclic electron flow have been identified as involving the proton-gradient-regulation-5/proton-gradient-regulation-like protein 1 (PGRL1) complex or the NADH dehydrogenase-like complex (158). The transmembrane thylakoid protein PGRL1 has recently been found to act as an Fdx-PQ reductase in Arabidopsis plants (141). The enzyme is most likely redox regulated, as its activity requires several redox-active cysteine residues and is controlled by Trx m4, providing a possible mechanism to link cyclic electron transport and stromal redox state (63, 141). Sensitive regulation of cyclic and linear electron flow will be required to rapidly adjust the ratio of NADPH and ATP production in response to fluctuating light conditions and to changes in the activities of stromal reactions with different requirements of ATP and/or NADPH as cofactors (160). In addition to this, excess reducing equivalents can be exported to the cytosol via the malate/oxaloacetate (OAA) shuttle, involving redox-dependent NADP-dependent malate dehydrogenase (MDH) in the chloroplast and NAD-dependent MDH in the cytosol for interconversion (302) (see also Table 1).
In the next section, we will not only describe the most important metabolic processes in the plastid stroma using NADPH, Fdx, and/or ATP provided by the photosynthetic light reactions, but we will also cover redox signals that are involved in their regulation. Light has been identified as the most important factor that is involved in the redox-status control of stromal metabolism via the Fdx/Trx system (Fig. 3a and Table 1). In addition to this, metabolic, developmental, and abiotic/biotic stress signals have to be integrated into this redox-regulatory network, possibly via alternative redox systems linked to NADPH-dependent thioredoxin reductase C (NTRC) or antioxidant metabolism (Fig. 3b and Table 1).

Carbon fixation cycle
The Calvin–Benson cycle utilizes the products of the light reactions of photosynthesis, ATP and NADPH, to reduce inorganic CO2 to organic sugars, serving as the primary pathway for carbon fixation in higher plants (106, 223). The sequence of reactions of this cycle is well known since its discovery in the years 1946–1954 (35, 50, 246). It can be divided into three stages (Fig. 2). In the initial stage, carbon enters the cycle via carboxylation of ribulose-1,5-bisphosphate (RuBP) by ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) to produce two molecules of glycerate-3-phosphate (3PGA). In the second stage, 3PGA is reduced to triose phosphates via two subsequent reactions catalyzed by phosphoglycerate kinase (PGK) and NADP-dependent glyceraldehyde-3-phosphate dehydrogenase (GAPDH) using most of the ATP and NADPH delivered by the light reactions. In the final stage, the acceptor RuBP is regenerated from triose phosphates via a sequence of different reactions catalyzed by aldolase, fructose-1,6-bisphosphatase (FBPase), transketolase (TK), sedoheptulose bisphosphatase (SBPase), and phosphoribulokinase (PRK), with the latter reaction also requiring ATP.
Studies in the 1960's on subcellular metabolite levels in leaves showed that activation of the Calvin–Benson cycle on illumination is accompanied by a transient decrease in the NADPH/NADP+ and ATP/ADP ratios in the chloroplast stroma (136). This indicated that light activation of the Calvin–Benson cycle is regulated by a mechanism which can override regulation by changes in the stromal NADPH and ATP levels. The mechanism was discovered by Buchanan and coworkers who found that FBPase and other enzymes of the photosynthetic carbon reduction cycle are activated by the Fdx/Trx system, which is directly linked to light-driven photosynthetic electron transport, rather than to stromal metabolism (48). Illumination promotes the reduction of Fdx at the acceptor side of PS I, which, in turn, leads to the sequential reduction of FTR and Trxs, which activate several target enzymes of the Calvin–Benson cycle (GAPDH, FBPase, SBPase, PRK, and Rubisco activase) by reducing intramolecular disulfide bonds (307). This enables the co-ordinated activation of the carbon fixation cycle in the light and its inhibition in the dark, when the Trx system is reoxidized (307). As shown by biochemical studies, the Trx-dependent enzymes of the Calvin–Benson cycle are preferentially or exclusively regulated by f-type Trxs, with m-type Trxs playing a much less prominent role (223, see Table 1). Thiol modulation is combined with additional regulatory mechanisms such as (i) protein complex formation of GAPDH and PRK via CP12 (146) and (ii) metabolic fine control acting on the mid-point redox potential of the regulatory cysteines (i.e., redox activation of FBPase requires fructose-1,6-bisphosphate) (281) or via allosteric effects (101). Light regulation is additionally supported by changes in pH and Mg2+ concentrations in the chloroplast stroma on illumination, which have direct regulatory effects on Rubisco, FBPase, SBPase, and PRK (48, 101). The combination of these mechanisms act to ensure that the Calvin–Benson cycle can be switched on in the light and off in the dark to avoid futile cycling of carbon metabolism. Moreover, they enable balancing of the different reactions of the cycle to avoid its blockage by accumulation or depletion of intermediates. While recent proteomic studies suggest that Calvin–Benson cycle enzymes also undergo other types of post-translational redox modifications, such as nitrosylation and glutathionylation, the importance and function of these additional redox modifications still have to be determined (223).
Rubisco also catalyzes a side reaction with O2, leading to the formation of one molecule of 3PGA and one molecule of 2-phosphoglycolate (2PG). The latter is detoxified and recycled back to 3PGA via photorespiration, a complex energy consuming pathway that liberates fixed carbon and nitrogen as CO2 and NH4 +, respectively (31). Moreover, the photorespiratory cycle is a highly compartmentalized process with enzymatic reactions occurring across four compartments. In the chloroplast stroma, 2PG is dephosphorylated to glycolate, which is subsequently exported from the chloroplasts—via the cytosol—to the peroxisomes where it is oxidized to glyoxylate and transaminated to glycine, which—via the cytosol—enters the mitochondria, where two molecules of glycine are deaminated and decarboxylated to form one molecule each of serine, ammonia, and carbon dioxide. Serine is exported from the mitochondria to the peroxisomes, where it is mainly converted to glycerate, which leaves the peroxisomes and is taken up—via the cytosol—into the chloroplast, where it is phosphorylated to yield 3PGA. While the enzymes involved in the photorespiratory cycle have been identified and characterized, little is known about the identity of the transporters catalyzing the exchange of photorespiratory metabolites between the different subcellular compartments. However, first steps in this direction are being taken with a chloroplastidic glycolate/glycerate transporter recently being identified in Arabidopsis that is necessary for the function of the photorespiratory cycle (269). Strategies to decrease photorespiration by CO2-concentrating mechanisms or to improve photorespiratory pathways have been recently reviewed extensively in the literature and will not be covered here (32, 267, 326). The role of the photorespiratory cycle in mitochondria and peroxisomes and its role in the exchange of redox equivalents between organelles will be discussed in greater detail in the respective chapters later.
Starch metabolism
The stoichiometry of the Calvin–Benson cycle requires 5/6th of the triose phosphates to be used to regenerate RuBP as CO2 acceptor, while the remainder can exit the cycle to be used to synthesize sucrose and starch as major end products (Fig. 2), which serve as the ultimate source of carbon for plant growth (326, 338). While sucrose synthesis follows the export of triose phosphates to the cytosol via the triose phosphate/inorganic phosphate (Pi) translocator at the inner membrane of the chloroplast envelope (293), starch is synthesized in the chloroplast stroma, using fructose-6-phosphate (Fru6P) delivered by FBPase in the Calvin–Benson cycle (103, 391). After conversion of Fru6P to glucose-1-phosphate (Glc1P) via the sequential action of phosphoglucose isomerase (PGI) and phosphoglucomutase (PGM), the first committed step of starch synthesis involves the conversion of Glc1P and ATP to ADP-glucose (ADPGlc) and inorganic pyrophosphate (PPi), catalyzed by ADPGlc pyrophosphorylase (AGPase). ADPGlc acts as the glucosyl donor for different classes of starch synthases (SS), which elongate the α-1,4-linked glucan chains of the starch polymers. Five distinct SS classes are known in plants: granule-bound SS, which is responsible for the synthesis of amylose, and soluble SS 1–4, which is responsible for amylopectin synthesis. Branch points are introduced by two classes of starch branching enzymes (SBE 1 and 2), which differ in terms of length of the glucan chains transferred and substrate specificities. Interestingly, starch synthesis also involves two types of debranching enzymes (ISA 1 and 2), which cleave branch points and are probably involved in tailoring the branched glucans into a form that is capable of crystallization within the starch granule.
In Arabidopsis leaves, the majority of control of starch biosynthesis has been found to reside in the reaction catalyzed by AGPase (79). AGPase is rapidly activated on illumination by reduction of an intermolecular disulfide bond between the Cys residues joining the two small subunits (APS1) of this heterotetrameric enzyme (139, 346). Using transgenic Arabidopsis plants expressing a mutated AGPase in which the regulatory Cys 81 of APS1 has been substituted by Ser, genetic evidence has been provided that redox regulation of AGPase contributes significantly to photosynthetic starch turnover during the light/dark cycle in leaves (125). AGPase from potato tubers and pea (Pisum sativum) leaf chloroplasts has been shown to be reduced by Trxs f and m in vitro (17, 104, 346). Using recombinant purified Arabidopsis proteins, it was shown that Trx f1 redox activates AGPase more efficiently than other types of plastidial Trxs, such as Trx m1, x, and y1 (346). Recent studies in Arabidopsis mutants with an insertion in the trx f1 gene provided in planta evidence for the role played by Trx f in the light activation of AGPase and photosynthetic carbon partitioning in plants (346). In these mutants, inactivation of Trx f1 led to decreased light activation of AGPase and decreased starch accumulation in leaves (346), while inactivation of isoforms of Trx m and Trx y in other Arabidopsis mutants had no such effects (I. Thormählen, M. Paul, E. Issakidis-Bourguet, and P. Geigenberger, unpublished results). This indicates a specific role of Trx f in regulating starch synthesis, similar to the established role of Trx f in regulating the Calvin–Benson cycle (see Table 1). The common regulation by Trx f of both metabolic processes enables photosynthesis and end-product synthesis to be co-ordinately regulated in response to light via the same signaling pathway.
Interestingly, Arabidopsis mutants lacking Trx f1 revealed no changes in photosynthetic parameters and growth, although Trx f protein levels were decreased by more than 95% relative to the wild type (346). This is surprising, given the exclusive regulation of individual steps of the Calvin–Benson cycle (i.e., FBPase) by f-type Trxs (see Table 1). While this may point to a more complex regulation of the Calvin–Benson cycle as initially expected, this may also be due to Trx f having much lower affinities for AGPase (17) compared with FBPase (239), indicating that changes in Trx f expression levels in response to transgenesis (346) circadian rhythms (26), thiol status, and sugars (25) or indeed any other stimulus will affect starch synthesis to a greater extent than the Calvin–Benson cycle. Consistently, in transplastomic tobacco plants, very severe over-expression of Trx f led to a strong increase in leaf starch accumulation, without leading to an increase in photosynthesis (290, 298).
In addition to redox, AGPase is also the subject of allosteric regulation, being activated by the first product of the Calvin–Benson cycle (3PGA) and inhibited by Pi (18). Since the concentrations of these effectors will change during light/dark transitions in a characteristic manner, this will contribute to light activation of AGPase. Specifically, allosteric regulation and redox regulation will act synergistically on AGPase to ensure starch synthesis is efficiently activated in the light and inactivated in the dark. While the allosteric activator 3PGA promotes light-dependent redox activation of AGPase by Trxs (139), the sensitivity of AGPase to its allosteric effectors is strongly affected by reversible oxidation of its regulatory cysteine (347).
In addition to light, redox activation of AGPase is also promoted by sugars, in illuminated as well as in darkened leaves and in non-photosynthetic tissues (139, 178, 347). In leaves, this enables starch synthesis to be regulated in response to changes in the balance between carbon supply and growth (111); while in non-photosynthetic storage organs such as potato tubers, this enables starch synthesis to be regulated in response to fluctuations in the supply of sucrose from the leaves (105, 347). Redox activation of AGPase was found to be closely correlated with the sugar content across a range of physiological and genetic manipulations, with light leading to an additional activation in leaves (139, 347). Reductive activation of AGPase in non-photosynthetic tissues or in nocturnal leaves requires alternative systems of electron transfer linked to NADPH generated from sugars, rather than to photoreduced Fdx (Fig. 4). This involves dark operative redox systems that are based on (i) Fdx/Trx systems involving a more oxidizing Fdx receiving electrons from NADPH in amyloplasts (20) or (ii) NTRC, localized in both chloroplasts and amyloplasts (176).
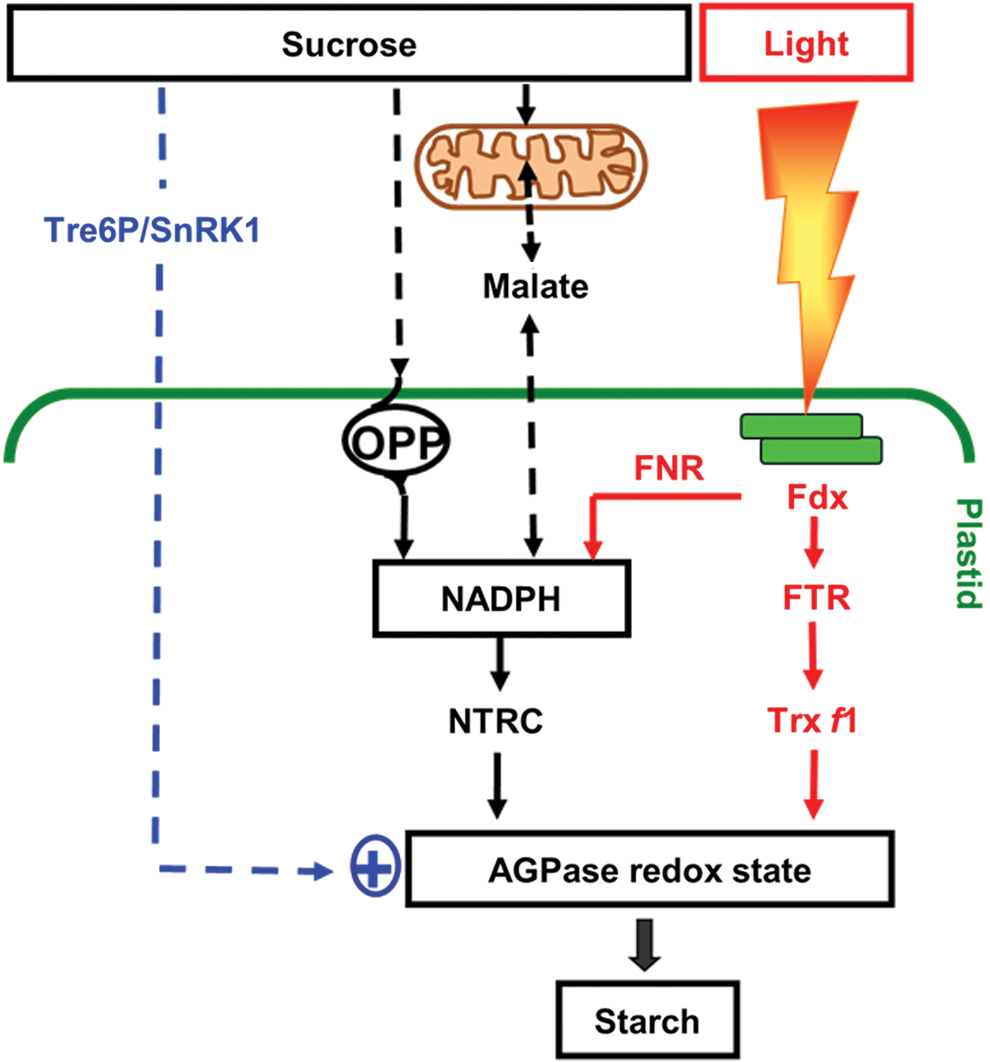
NTRC is a bimodular protein containing both an NADPH-dependent thioredoxin reductase (NTR) and a Trx domain on a single polypeptide (38). By using NADPH as a reducing power, NTRC conjugates both NTR and Trx activities to reductively activate AGPase and concomitant starch synthesis (222). Using an insertional knock-out mutant, NTRC has been found to play a role in the regulation of AGPase and starch synthesis in leaves and non-photosynthetic tissues in response to both light and sugars in the dark (196, 197, 222). In the light, NTRC can use photo-reduced NADPH provided by FNR; while in the dark, NADPH is most likely provided by the initial reactions of the OPP, glucose-6-phosphate dehydrogenase (G6PDH), and 6-phosphogluconate dehydrogenase (6PGDH). The role of the OPP in redox regulation of starch synthesis has been confirmed by recent studies showing chloroplast-localized 6PGDH to be required for starch accumulation in maize mutants (322). The chloroplast isoforms of G6PDH are subject to post-translational redox regulation via Trx f, leading to inactivation of the OPP in the light and activation in the dark (244). In heterotrophic amyloplasts, G6PDH isoforms are less sensitive to redox status control, enabling high levels of NADPH to be produced to drive biosynthetic processes in non-photosynthetic tissues (378). While glucose leads to redox activation of AGPase via an increase in the NADPH/NADP+ ratio, sucrose and trehalose most likely act via additional signaling pathways that have not yet been clarified (178). These disaccharide sugars are linked to trehalose-6-phosphate (Tre6P), a signal metabolite implemented in diverse developmental (371) and metabolic responses, including redox regulation of AGPase and starch metabolism (178, 209). In addition to Tre6P, redox activation of AGPase is also linked to other elements of the cytosolic sucrose signaling cascade, such as the conserved SNF1-related protein kinase (SnRK1) in Arabidopsis leaves (161) and potato tubers (216, 348), although the underlying mechanisms have not yet been resolved.
In addition to AGPase, reductive activation by thiol/disulfide modulation has also been demonstrated for other enzymes involved in the pathway of starch synthesis downstream of AGPase, such as SS 1 and SS 3, SBE 2, and ISA 1 and ISA 2, by using comprehensive in vitro studies (115). This enables a co-ordinated activation of the supply and the use of ADPGlc for starch synthesis by Trx-dependent redox regulation, linking external factors such as light and sucrose to redox activation of the whole pathway. It also implies that redox status regulation may influence starch structure in addition to quantity, depending on the contribution of the different redox-regulated isoforms of SS and (de-)branching enzymes to synthesize the polymers. In addition to this, redox regulation of SS 1, SS 3, and SBE 2 might be important for the involvement of these isoforms in the formation of protein complexes, which have been suggested to play a role in the regulation of starch biosynthesis (140, 345). Interestingly, SS isoforms and NTRC have been implicated in the accumulation of starch in response to microbial volatiles (198).
In leaves, starch is remobilized at night to support chloroplast metabolism and sucrose export when photosynthesis is inactive; while in heterotrophic storage organs, starch is remobilized later in development to support phases of reproductive growth (391). The pathway of starch degradation has been recently established in Arabidopsis leaves (391). After reversible phosphorylation of glucans on the surface of the starch granule via glucan water dikinase (GWD) and glucan phosphatase (SEX4), starch granules are attacked most probably by alpha-amylase and the resulting branched glucans are subsequently converted to unbranched alpha-1,4-glucans via debranching enzymes (isoamylase and pullulanase). Linear glucans are metabolized by the concerted action of β-amylase and disproportionating enzyme to glucose and maltose, which leaves the chloroplast via a maltose transporter in the inner envelope membrane to support cytosolic metabolism (Fig. 2).
Despite our sound knowledge with regard to the pathway, relative little is known about its regulation. There is evidence that starch degradation is controlled by diurnal rhythms (117) and by redox status (115). It has been found that both of the enzymes involved in reversible starch phosphorylation, GWD (226) and SEX4 (313), as well as the more downstream acting enzymes alpha-amylase (115, 381), beta-amylase (321), and limit dextrinase (115, 306), are reductively activated by Trxs f and m. While this shows that redox status regulation also extends to enzymes of starch degradation, reductive activation of these enzymes is counterintuitive in the context of light/dark regulation in the chloroplast, as it would imply that starch degradation is inactivated in the dark, when the Trx system is oxidized. While the roles, in this context, of different Trx isoforms and NTRC have not been clarified, the use of NADPH as an electron donor via NTRC could enable redox status activation of starch degrading enzymes also in the dark. In addition to this, recent studies provide evidence for a role of the redox status in regulating starch degradation under certain stress conditions (165, 166, 356) or in the context of specific tissues, cell types (356), or subcellular compartments (376) having different requirements for regulation of starch degradation.
Lipid synthesis
Similar to the metabolism of starch in most species and tissues, the reactions of de novo fatty acid biosynthesis occur exclusively in the plastid (259). In leaf chloroplasts, carbon used for fatty acid synthesis mainly derives from the Calvin–Benson cycle, using a pathway that converts 3PGA to acetyl-CoA involving pyruvate kinase (PK) and the pyruvate dehydrogenase (PDH) enzyme complex (23). Acetyl-CoA carboxylase (ACCase) catalyzes the first committed step of de novo fatty acid synthesis in the plastid stroma, converting acetyl-CoA and CO2 to malonyl CoA by using ATP as an energy donor (Fig. 2). Malonyl CoA is used as a substrate for fatty acid synthase (FAS), a large multisubunit enzyme complex that performs the cyclic condensation of two carbon units with the extending acyl chains being covalently bound to acyl carrier protein (259). These reactions are a strong sink of ATP and NADPH, which are provided by the light-driven photosynthetic electron transport in leaves, or respiratory processes in non-photosynthetic tissues. Synthesis of one molecule of C16-palmitic acid from eight molecules of acetyl-CoA requires 14 molecules of NADPH and seven molecules of ATP. Conversion of saturated fatty acids to unsaturated forms is catalyzed by fatty acid desaturases, which require Fdx as an electron acceptor in the plastid (259). Fatty acids are used as substrates for the synthesis of glycerolipids, which form the lipid bilayer of all cellular membranes or serve as important carbon reserves in selected seeds (170).
ACCase is a key-regulatory step in the pathway of fatty acid biosynthesis in different organisms, including plants (149). The chloroplast enzyme is a multienzyme complex that consists of four different polypeptides, biotin carboxylase, biotin carboxyl carrier protein, and the carboxyltransferase alpha and beta subunits, encoded in the nucleus and the chloroplast, respectively. In vitro studies revealed that chloroplast ACCase is activated by Trxs f and m (299), catalyzing the reduction of an intermolecular disulfide bond between the carboxyltransferase subunits (183). Light leads to reductive activation of ACCase in isolated pea chloroplasts (183) and Arabidopsis leaves (A. Kolbe, M. Ehrlich, and P. Geigenberger, unpublished results), indicating that fatty acid biosynthesis is regulated by the Fdx/Trx system in a similar manner as the Calvin–Benson cycle (see Fig. 3A). Since fatty acid synthesis is a strong sink of NADPH and ATP (see 51), the use of these compounds has to be strictly coordinated with their supply by photosynthetic light reactions. In addition to light, reductive activation of ACCase has also been shown to be promoted by sucrose, in darkened leaves as well as in non-photosynthetic tissues (A. Kolbe, P. Waldeck, M. Ehrlich, H. Vigeolas, and P. Geigenberger, unpublished results). Similar to the sucrose-dependent redox regulation of AGPase (see Fig. 4), this could involve NTRC as a dark operative redox system in the plastid as well as specific sucrose signals from the cytosol, although direct evidence for their contribution in the regulation of ACCase is lacking at the moment. Recent studies show that in addition to the carboxyltransferases, the other subunits of ACCase are also potentially controlled by redox regulation. There is proteomic evidence for S-thiolation of the biotin carboxyl carrier subunit by glutathione in Chlamydomonas (224) and for glutathionylation of biotin carboxylase in Arabidopsis (71). Moreover, redox status regulation also extends to downstream metabolic reactions of galactolipid biosynthesis, as the envelope bound monogalactosyldiacylglycerol synthase that synthesizes the major lipid component of chloroplast thylakoid membranes has been found to be subject to Trx-dependent redox regulation (388).
Nitrate assimilation, chlorophyll synthesis, and antioxidant metabolism
Key reactions of many other important metabolic pathways, such as N-assimilation, chlorophyll synthesis, and antioxidant metabolism, are also located in the plastid (Fig. 2). The initial step of nitrate assimilation, the conversion of nitrate to nitrite, is catalyzed by nitrate reductase in the cytosol, using NAD(P)H as a reducing power. The highly reactive nitrite is immediately transported into the plastid, where it is converted to ammonium by nitrite reductase (NiR) using Fdx as an electron donor (144, 297). Eight molecules of reduced Fdx are required to reduce one molecule of nitrate to ammonium. The ammonium ions provided by NiR or photorespiration are rapidly assimilated into amino acids via the sequential action of glutamine synthetase (GS) and glutamine:oxoglutarate amino transferase (GOGAT), which catalyze the ATP-dependent conversion of ammonium and glutamate (Glu) to glutamine (Gln) and the conversion of Gln and 2-oxoglutarate (2OG) to two molecules of Glu, respectively (190). In the chloroplast, GOGAT accepts electrons directly from Fdx; while in non-photosynthetic plastids, electrons are provided by NADPH. Provision of 2OG is most likely a function of isocitrate dehydrogenase (ICDH) isoforms in the cytosol and mitochondria catalyzing the conversion of isocitrate and NAD(P)+ to 2OG and NAD(P)H, with 2OG being transported into the chloroplast via a 2OG/malate transporter (143).
Given that the reactions of nitrate assimilation are a strong sink for electrons, they have to be closely coordinated with the electron pressure of the photosynthetic light reactions. This is achieved by direct electron transfer from photoreduced Fdx to NiR and GOGAT in the chloroplast stroma (128). In addition to this, there is in-vitro evidence that NiR (212), GOGAT (201), and GS2 (19, 59, 241) are subject to redox status regulation by Trxs. Moreover, when DTT was fed to Arabidopsis leaves to increase the thiol state of the tissue, the levels of 2OG decreased while amino-acid levels increased, which indicates that redox status regulation of the GOGAT/GS cycle is most likely operational in vivo (177).
Glutamate (Glu), as the first product of ammonium assimilation, is used to synthesize many other amino acids and nitrogenous metabolites in plants. It serves as a precursor for the synthesis of 5-aminolevulinic acid (ALA), which is the starting point for the biosynthesis of chlorophyll (Chl) and heme in the chloroplast (51). Glutamate is converted to ALA by a sequence of reactions involving NADPH-dependent glutamyl-transfer RNA reductase (GluTR), Glu1-semialdehyde aminotransferase, porphobilinogen synthase, and uroporphyrinogen decarboxylase. ALA is subsequently converted in a series of enzymatic steps to protoporphyrin IX, the common branch point for the synthesis of heme and Chl. To make Chl, a polymeric magnesium (Mg) chelatase complex inserts Mg2+ into the protoporphyrin IX ring, which is further modified via the subsequent reactions of a methyltransferase (CHLM), an oxidative cyclase complex, an NADPH:vinyl reductase, and an NADPH:protochlorophyllide oxido-reductase (POR) to synthesize Chl a. The synthesis of Chl b occurs with Chl a as precursor most likely via Fd-dependent chlorophyll a oxygenase (283). After binding to the photosystems and light-harvesting complexes in the thylakoid membranes, both Chl a and Chl b serve as important components in photosynthetic light absorption.
Chlorophyll synthesis has been identified as a light-dependent process in photosynthetic angiosperms (51). The CHL1 subunit of Mg-chelatase was found to be reductively activated by Trx f in vitro (153), while virus-induced gene silencing of Trx f and Trx m in transgenic pea plants showed that this mechanism is also operational in vivo (153). Proteomic approaches also identified other enzymes of Chl biosynthesis as potential Trx targets. These involve enzymes catalyzing the conversion of Glu to ALA, such as of Glu1-semialdehyde aminotransferase, porphobilinogen synthase, and uroporphyrinogen decarboxylase (49). Recent studies provide evidence that Chl synthesis is also regulated by NTRC as a second redox system (292). NTRC uses NADPH to reductively activate GluTR, involved in the synthesis of ALA, and CHLM, involved in the conversion of protoporphyrin IX to Chl a. A physical interaction of NTRC with GluTR and CHLM was confirmed by biomolecular fluorescence complementation assays, while knockout of NTRC led to a decrease in GluTR and CHLM activities and concomitant chlorophyll synthesis in Arabidopsis mutants in vivo (292). This shows that different stages of Chl synthesis are subject to post-translational redox regulation, involving Fdx/Trx and NADPH/NTRC as redox systems (Fig. 3 and Table 1). While the Fdx/Trx system enables Chl synthesis to be regulated in response to light signals, the role of the NADPH/NTRC system is less clear in this context. Similar to the role of NTRC in activation of starch synthesis (222), NTRC could complement the Fdx/Trx system in light activation of Chl synthesis, by acting at different target enzymes in this pathway (Table 1). Moreover, NTRC could Iink the rate of Chl synthesis to the availability of NADPH, which is required as an important reducing equivalent at different steps of the pathway (Fig. 2) (51).
Finally, metabolic reactions are required to scavange reactive oxygen species (ROS) that are produced during photosynthetic electron transport in the chloroplast. Under conditions of acceptor limitation of PS I, electrons from PS I will photoreduce oxygen to superoxide radicals, which subsequently disproportionate to hydrogen peroxide, either spontaneously or in a reaction catalyzed by superoxide dismutase (15). While hydrogen peroxide becomes toxic when it accumulates at a high level (4), it also serves as an important signaling molecule regulating photosynthesis and other processes in plants (268). A sensitive regulation of hydrogen peroxide levels is, therefore, important for its signaling function and detoxification. Hydrogen peroxide is reduced to water by enzymes such as ascorbate peroxidase, converting ascorbate to monodehydroascorbate, with the latter being recycled to ascorbate by using electrons from photoreduced Fdx (15). Hydrogen peroxide detoxification in the chloroplast also involves a 2Cys-peroxyredoxin (Prx). In the light, photoreduced Fdx serves as a source of reducing power to reduce 2Cys-Prx by Trx x via the FTR/Trx x pathway (180). In addition to this, 2Cys-Prx can be reduced by NTRC using electrons provided by NADPH, which is generated via FNR in the light or the OPP in the dark (266). NTRC knockout in Arabidopsis mutants led to increased levels of hydrogen peroxide and lipid peroxidation under conditions of prolonged darkness followed by illumination, while no such effects were observed in a normal diurnal cycle (266). While NTRC is complementing the Fdx/Trx x system in diverting electrons to the reduction of 2Cys-Prx in the light, it may serve as a key detoxification system in the dark, using metabolically produced NADPH as a reducing power. The role of the OPP to provide NADPH for ROS detoxification under these conditions has been recently confirmed (67). The NADPH-dependent hydrogen peroxide-scavenging system via 2Cys-Prx and NTRC has been shown to be required for the protection of the highly ROS-sensitive Mg-protoporphyrin monomethylester cyclase involved in chlorophyll biosynthesis (323, 324). Redox status-sensitive regulation of the cyclase complex will affect accumulation of Mg-protoporphyrin IX, which has been suggested to act as a plastid retrograde signal controlling the expression of photosynthetic genes in the nucleus (24) (see section “Integration at the Cellular Level”).
Interaction of plastid metabolism and gene expression
Due to the endosymbiotic origin of chloroplasts, they contain their own genome and gene expression machinery. This machinery is known to be highly regulated, especially in response to different environmental stimuli (27). Light plays a crucial role, especially for chloroplast protein translation based on findings that synthesis rates of chloroplast encoded photosynthetic proteins increase on illumination despite the fact that their respective mRNA levels stay constant (78). In addition to this, metabolic and developmental signals have to be integrated in this context. Recent studies in Chlamydomonas provide evidence for a cross-talk between chloroplast protein translation and carbon metabolism. In the study of (41), it was shown that a subunit of the plastidial PDH complex (DLA2), which synthesizes acetyl-CoA as a precursor for lipid synthesis, forms ribonucleoprotein particles and influences chloroplast mRNA translation. Conversely, RNA binding affects plastidial PDH metabolic activity. This reciprocal regulation may function in co-ordinating the synthesis of lipids and proteins for the biogenesis of photosynthetic membranes. In a second example, the same group showed that redox status regulation of chloroplast mRNA translation of the psbD gene, encoding the D2 protein of PS II, involves the reduction of a single intermolecular disulfide bridge between two translation-activation proteins in the dark, most likely via NADPH-dependent NTRC, acting as a dark operative chloroplast redox system (308). This provides evidence for a connection between plastid carbon metabolism and protein synthesis via NTRC-mediated redox signaling.
Mitochondrial Redox Biology
Mitochondrial redox biology and, indeed, mitochondrial metabolism is dominated by its major flux-bearing pathways, namely respiration (228) and the pathway of photorespiration that it shares with the peroxisome, mitochondria, chloroplast, and cytosol (31, 351). Mitochondrial respiration provides ATP, reductant, and carbon skeletons in both the light and the dark (Fig. 5). It is, however, considerably downregulated in the light; however, to what extent remains somewhat controversial (256, 343). The mitochondrial proteome is responsive not just to light conditions but also to tissue type and to a range of biotic and abiotic stresses (156, 193, 336), as such it follows that the mitochondria need to harbor a considerable degree of dynamic flexibility in order to maintain both its own function and that of the cell in general (309). In this section, we not only will focus largely on the redox chemistry of the mitochondrial respiratory machinery but will also integrate the mitochondrial steps of photorespiration into this discussion as well as covering relevant signal pathways emanating from the mitochondria. Oxidative phosphorylation in the mitochondria of plants is most commonly fueled by the breakdown of sucrose (Fig. 5). Sucrolysis is the pathway by which sucrose is degraded into pyruvate in the cytosol [although some reactions are additionally localized in the plastid (3) and a small proportion of the enzyme activities of the entire glycolytic pathway is localized to the outer mitochondrial membrane (112, 119)], the reactions of the TCA cycle completely oxidize pyruvate to CO2 (85) with electrons being transferred to NAD+ and FAD yielding NADH and FADH2 while phosphorylating some ADP directly. Complete respiration of a molecule of sucrose will result in the release of 20 NADH and 4 FADH2—these reduced co-enzymes are subsequently oxidized by the mitochondrial electron transport chain (mETC) (Fig. 5). The free energy released by mitochondrial electron transport is partially coupled to the translocation of protons across the inner mitochondrial membrane, creating an electrochemical gradient across the inner membrane, although in plants flexibility exists here due to the presence of non-proton pumping enzymes. The free energy released by the movement of the protons back across the inner membrane through the F0 proton channel of the ATP synthase complex is used by the F1 component to convert ADP and Pi to ATP (85). However, it is important to note that many of the intermediates of this pathway are important precursor molecules in their own right for a plethora of biosynthetic pathways and, as such, these numbers are likely to highly overestimate the true respiratory yield of sucrose breakdown.

Mitochondrial contribution to energy metabolism—respiration
As stated earlier, the mitochondria is dominated by respiration and in photosynthetic cells, the oxidative reactions of photorespiration that reside in this organelle, namely the concerted action of glycine decarboxylase and serine hydroxymethyltransferase (32). Mitochondrial respiratory activity, in turn, is closely connected with NADH production with four of the eight core enzymes of the cycle as well as the intimately related mitochondrial PDH producing this co-factor. To re-capitulate, the TCA cycle is composed by a set of eight enzymes primarily linking the product of the oxidation of pyruvate and malate (generated in the cytosol) to CO2 with the generation of NADH for the oxidation by the mitochondrial respiratory chain (85, 228). Mitochondria are the only organelles in which a full cycle is present, although this does not in itself necessitate that it always functions in a fully cyclic mode (333). The presence of organic acids is known to support numerous and diverse functions within and beyond cellular metabolism; the level of accumulation of the various organic acids is extremely variable between species, developmental stages, and tissue types (7, 86). For this reason, it is likely that the enzymes involved in the inter-conversion of these metabolic intermediates are subject to tight regulatory control. Hints to the regulation of the TCA cycle have been provided by a recent metabolic control analysis which shows that much of the control through this pathway is resident in fumarase, MDH, and 2OG dehydrogenase (8). An important comment needs to be made here. Although the summed control co-efficient values reported exceed 1 and, as such, appear to violate the summation theory, it is critical to note that the summation theory is only valid in instances in which there is no control (which can, of course, be a negative as well as a positive resident outside of the pathway under consideration). This fact, apart from this study, is somewhat in conflict with the finding of modeling studies of Hagedorn et al., which suggested that the rate of oxygen consumption was insensitive to changes in MDH activity (126), suggesting a further study is required to clarify the actual contribution of this enzyme.
Unfortunately, the lack of subcellular information with regard to the levels of intermediates of the cycle (335) currently precludes us from being able to assess the potential of the constituent enzymes to play regulatory roles merely on the basis of disequilibrium ratios. That said, a massive amount of data has, nevertheless, been compiled from both reductionist in vitro studies and more recent holistic, systems-based approaches.
It has long been demonstrated that pyruvate decarboxylase (PDC) as well as TCA cycle dehydrogenases displays product inhibition in vitro by NADH [for a review, see Ref. (238)]. Accordingly, given that the in vivo activities of PDC and other TCA cycle enzymes are responsive to the NADH/NAD+ ratio, this provides a very sensitive mechanism by which it is possible to balance the rate of pyruvate oxidation by PDC and the TCA cycle activity with the rate of oxidative phosphorylation (272). Given that NAD+ is a common co-factor of three enzymes of the TCA cycle proper as well as by the PDC, it is reasonable to assume that mitochondrial NADH/NAD+ ratio has a major impact on the flux through the TCA cycle. However, it is equally important to note, with regard to these enzymes, that elegant studies from the Møller laboratory have revealed that the free NADH concentration is kept constant in plant mitochondria under different metabolic conditions, rendering it crucial to interpret implications of in vitro kinetics with caution (168). In addition to this regulation, the mitochondrial PDC is regulated by product inhibition by acetyl CoA (353) and activated by thiamine pyrophosphate (40).
The subsequent and first true reaction of the TCA cycle that was catalyzed by citrate synthase (CS) is also regulated by the cellular NADH/NAD+ ratio as well as activated by high ATP/ADP ratios and succinyl CoA levels, suggesting that its activity is tightly regulated at the metabolite level (253). The next step of the TCA cycle is catalyzed by aconitase, which catalyzes the reversible hydration of cis aconitate to either citrate or isocitrate. Irrespective of how many genes encode this enzyme, it appears that activities are localized in the mitochondria and cytosol (13, 53) and it is also well documented as being highly sensitive to oxidative stress (194, 258, 312). It has, furthermore, been demonstrated that the lack of manganese superoxide dismutase results in the inhibition of aconitase and the subsequent enzyme of the cycle, NAD-dependent ICDH (240). NAD-dependent ICDH additionally demonstrates product inhibition by NADH, and it is also inhibited by NADPH (192). The next reaction of the cycle that was catalyzed by the 2OG dehydrogenase complex (OGDHC) represents a metabolic branch point connecting 2OG (and the TCA cycle) with nitrogen assimilation with 2OG either being irreversibly degraded by the OGDHC or providing carbon skeletons for nitrogen assimilation. OGDHC is allosterically regulated in response to second messengers and metabolic indicators, such as Ca2+, ATP/ADP, NADH/NAD+, and thiamine pyrophosphate (11). The subsequent enzyme of the cycle, succinyl CoA ligase (ScoAL), is feedback inhibited by intermediates of the pathway of porphyrin biosynthesis as well as competitively inhibited by malonate in the reverse direction but is activated by 2OG and inhibited by both citrate and isocitrate and all downstream intermediates of the TCA cycle when assayed in the forward direction (331).
Succinate dehydrogenase (SDH), also commonly referred to as complex II, plays a dual role in mitochondrial metabolism both as a member of the electron transport chain and TCA cycle (9, 148). The regulation of the enzyme was investigated in coupled mitochondria by simultaneously measuring oxygen uptake rates and ubiquinone reduction levels (2). This study revealed that the activation state level of the enzyme is unambiguously reflected in the kinetic dependence of the succinate oxidation rate on the ubiquinone redox poise. Kinetic results indicated that it is additionally activated by both ATP and ADP (2).
Allosteric properties of fumarase from pea (Pisum sativum L.) revealed inhibition of this enzyme by physiological concentrations of pyruvate, 2-OG and the adenine nucleotides ATP, ADP, and AMP (33). Accordingly, downregulation of this enzyme in tomato resulted in a relatively large reduction in the rate of respiration in comparison to the majority of other enzymes of the cycle (254). The cycle is completed by NAD-dependent MDH, which catalyzes the reversible oxidation of malate to produce OAA (255, 352). While the equilibrium position favors malate and NAD+ production, the in vivo removal of OAA by CS, coupled with the removal of NADH by the respiratory chain, causes the reaction to function in the direction of malate oxidation in most tissues (253). Therefore, it is again likely that accumulation of NADH would lead to an inhibition of the mitochondrial MDH activity, although as shown by the modeling study of (126) mentioned earlier, the additional presence of malic enzyme in plant mitochondria can generate the pyruvate required to operate in the direction of malate production.
In addition to these allosteric changes, some of the enzymes of the TCA cycle are regulated at the level of protein abundance—for example, in response to oxidative or flooding stress or during the diel cycle (179, 193, 341, 342). Similarly, they are potentially regulated by a wide number of post-translational modifications such as phosphorylation, Trx-mediated redox status regulation, and lysine acetylation (21, 89, 134). Indeed, interrogation of compiled metabolomics responses to abiotic stress reveals that the cellular levels of TCA cycle intermediates respond dramatically to multiple cues, including pharmacological manipulation of redox status, tissue oxygenation, and circadian clock-mediated processes (40, 99, 177, 257, 294). Returning to the sum effect of the TCA cycle dehydrogenases, it is clear that the redox balance of the NADH/NAD+ pool is set by the balance of electron influx and efflux (see Fig. 5)—an equation complicated considerably in plants by the action of alternative NADH dehydrogenases which are upregulated under conditions of stress (279, 309) and, to a lesser extent, by the operation of the NAD+ transporter, NDT2 (263). Furthermore, as stated earlier, the finding that the free NADH concentration is kept constant in plant mitochondria needs to be taken into account when analyzing the regulation of the TCA cycle. Indeed, the identification that many of the enzymes of the cycle bind to Trxs (21, 389) suggests that redox status regulation is more likely mediated by the NTR system than by direct allosteric considerations.
Mitochondrial contribution to energy metabolism—(photo)respiration, amino acid, lipid, and vitamin synthesis
In addition to the TCA cycle, other major redox-related pathways in mitochondria include the photorespiratory amino-acid metabolism mentioned earlier as well as cysteine, proline, and branched chain amino-acid and lipid and vitamin metabolism (334). In photosynthetically active tissues, the photorespiratory enzymes, glycine decarboxylase and serine hydroxymethyltransferase, are among the most prominent proteins of the mitochondrial matrix (31). As mentioned earlier, it is well established that glycine produced by photorespiration is taken up by mitochondria and oxidized by these two enzymes (85). Oxidation of NADH produced on glycine oxidation occurs preferentially over that from other substrates such as malate or succinate (37, 73). These findings have led to suggestions that protein complexes located in the vicinity of the respiratory chain may give preferential access of specific reducing equivalents to the respiratory chain (185); however, direct evidence for this is currently lacking. Irrespective of how glycine oxidation is prioritized, it is clear that the NADH requirement of peroxisomal hydroxypyruvate reductase is stoichiometrically equivalent to the NADH production by glycine oxidation in the mitochondrial matrix, leading to the proposal that the NADH produced in the mitochondria is utilized in the peroxisome (162). However, given that only 25% to 50% of the redox equivalents produced in the mitochondrial matrix are exported, another source of NADH is clearly required to cover this shortfall (304). Experimental evidence suggests that the activity of the malate-OAA shuttle in the chloroplastic envelope is sufficient to meet this demand (132). However, it remains likely that both chloroplasts and mitochondria simultaneously allocate some NADH to the peroxisomes although the exact contribution of the two sources remains to be quantified (186). Recently, a mechanism for facilitating the oxidation of photorespiratory NADH in the mitochondrion has been revealed. Biochemical and physiological analyses of a T-DNA insertional mutant of Arabidopsis deficient in the expression of the uncoupling protein AtUCP1 revealed a specific inhibition of photorespiration (337). Uncoupling proteins (UCPs) are integral to the inner mitochondrial membrane, where they catalyze proton conductance across this membrane, dissipating the mitochondrial proton gradient as heat (184, 366). It has been postulated that this is important when the demand for oxidation of NADH is high and may facilitate high TCA cycle flux (317). Consistent with this suggestion, the ucp1 mutants displayed a reduced photosynthetic carbon assimilation rate that was linked to a reduced rate of oxidation of photorespiratory glycine (337). The role of UCPs in other aspects of redox biology is discussed in detail next. As for the TCA cycle, operation of the mitochondrial steps of this pathway also requires the recycling of NAD+ in the mitochondrial matrix, a function that could be performed by the internal NADH dehydrogenase. Circumstantial support for this comes from the observation that the expression of a gene encoding an internal NADH dehydrogenase is strictly light dependent (332). Intriguingly, the abundance of this protein highly correlates with that of the alternative oxidase (AOX), raising the possibility of the operation of a truncated and entirely non-phosphorylating electron transport chain (85, 221).
Also partially localized in the mitochondria are the enzymes of cysteine synthesis, although a full complement of these enzymes is also duplicated in the plastid and the cytosol (137). That said, the mitochondria is clearly highly important for the production of O-acetyl serine and, as such, is likely of high importance for the overall cellular redox balance (123, 380). Similarly, redox reactions play an important role in proline degradation with mutants of the delta1-pyrroline-5-carboxylate dehydrogenase, catalyzing the second step in this pathway, being hypersensitive to proline and leading to programmed cell death, callose deposition, ROS production, and DNA laddering via a salicylic acid-mediated signal transduction pathway (69). Catabolism of proline has been demonstrated to be highly important in plants, not only in programmed cell death (138), but also in protection of catalase, peroxidase, and complex II activities (127, 262), as well as potentially having an important role in stimulating seed germination (130). Beyond these metabolites and the alternative substrates of the mETC (discussed in the section below), the mitochondria are also the site of metabolism for, among others, fatty acids, vitamins, and haem (334) as well as have their own ascorbate-glutathione cycle (AGC) (57, 157). Due to space constraints, we cannot detail redox status constraints of the constituent reactions in the former processes nor the subsequent roles in redox regulation of their products. However, readers are referred to earlier articles for details thereof (157, 334) as well as to the next section on the cytosol in the case of the AGC.
The mETC
As already implied in the earlier sections, the mETC is intimately linked to the redox status of the mitochondria. As with other eukaryotes, the mETC of plants comprises four large protein complexes (Fig. 5). In contrast, to other eukaryotes, however, the plant mETC is highly branched (6, 277) and features several alternative pathways catalyzed by NAD(P)H dehydrogenases and by the AOX with the former bypassing complex I and the latter complexes III and IV (230, 233, 363). Both of these proteins decrease respiratory ATP output by 30%–60%, while the UCP discussed earlier enables proton flow that bypasses ATP synthesis, conferring additional flexibility to plant metabolism (277). That said, although the role for UCP is at least partially elucidated in plants, that for AOX remains elusive despite many hypotheses, including roles in minimizing ROS production and as a defence against metabolic oscillations (122, 215, 276, 291).
Recently, several additional electron inputs into the mETC have been characterized in plants; for example, the electron transfer flavoprotein: quinone oxidoreductase (ETFQO), L-galactono-1, 4-lactone dehydrogenase, and glycerol 3phosphate dehydrogenase (28, 155, 255, 310) and, indeed, many more redox centers are involved. Depending on their respective contributions, these alternative inputs could potentially have a major effect on the redox status of the mitochondria and beyond. However, to get a better understanding, this requires that we initially consider the conventional cytochrome pathway. In all mitochondria, the principal respiratory transfer is through four membrane-bound proton complexes that catalyze electron transfer from NADH and FADH2 to oxygen. The two-electron reduction of ½ O2 by NADH involves a reduction potential difference of 1.14 V, which translates into 219.2 kJ of free energy released for every mole of NADH oxidized. The mETC, thus, facilitates stepwise modestly exergonic redox reactions rather than a single explosive one. As mentioned earlier, the plant mETC is augmented by a wide variety of novel or newly uncovered pathways for the oxidation of NAD(P)H and the reduction of oxygen. In brief, the cytochrome pathway links four multiple subunit protein complexes. Complex I is an NADH dehydrogenase that oxidizes the NADH generated in the mitochondrial matrix by the operation of the TCA cycle. Complex II, which includes one of the constituent enzymes of the TCA cycle, namely SDH, oxidizes succinate to fumarate. Similar to Complex I, Complex II transfers electrons to ubiquinone. Ubiquinone can carry one or two electrons, and both fully oxidized and fully reduced molecules are freely diffusible within the inner membrane, enabling it to shuttle electrons from Complexes I and II to Complex III. Complex III, subsequently, transfers electrons from ubiquinone to cytochrome c, a small peripheral membrane protein that carries one electron at a time from Complex III to Complex IV (also known as cytochrome c oxidase), the terminal electron carrier in the chain. For every four molecules of cytochrome c oxidized, one molecule of oxygen is reduced to two molecules of water.
While of critical importance to plants, the cytochrome pathway is augmented by multiple other elements in plants; arguably, the best researched are the alternative dehydrogenases and the AOX. The type II NAD(P)H dehydrogenases are located on the internal and external sides of the inner mitochondrial membrane and oxidize NADH or NADPH from the mitochondrial matrix or the cytosol (206, 276, 277). These reactions bypass the first energy conservation step maintained by the proton pumping complex I, which oxidizes matrix NADH. Among the type II NAD(P)H dehydrogenases, NDB proteins of potato (Solanum tuberosum) and Arabidopsis (Arabidopsis thaliana) are present on the external surface of the inner mitochondrial membrane and can be subject to calcium-mediated regulation (80, 107, 276). NDA and NDC proteins are, by contrast, matrix directed (276).
The function of these internal dehydrogenase is by no means fully elucidated; however, the fact that the K m for NADH of the internal alternative NADH dehydrogenase(s) is 10-fold higher than that of complex I (235, 278) renders this enzyme likely to oxidize NADH only under high matrix NADH concentrations, for example those attained during photorespiration (74, 150, 151). Circumstantial support for such a role is provided by the characterized light- and photoreceptor-dependent upregulation of nda1 gene expression in A. thaliana (81, 275). However, further study is required to ascertain how the interplay between complex I and these dehydrogenases is regulated.
The role of the NDB, however, is much better understood in plants. Nicotiana sylvestris overexpressing StNDB1 displayed a decrease in the NADPH/NADP+ ratio independent of changes in the NADH/NAD+ ratio in illuminated leaves. This, thus, demonstrated that the NDB1 enzyme was active and able to specifically modify the cellular NADP(H) pools, although given that no visual phenotype was observed it was difficult to conclude the physiological importance of mitochondrial NADPH oxidation (205). However, in a subsequent study, it was demonstrated that plants overexpressing StNDB1 exhibit an earlier transition from rosette stage to bolting, whereas a line suppressing the expression of both StNDB1 and NsNDB1 is delayed in this parameter. The phenotype was strongly correlated with stem-specific changes in NADPH reduction levels (206), supporting an important role of this enzyme in mediation of stem physiology. It remains highly likely, however, that the NDBs as well as NDAs and NDC have further functions which remain to be elucidated.
Another protein proving rather difficult to pin down a physiological role on is the AOX. As mentioned earlier, this protein, similar to the UCP, provides a means to relax the coupling between respiration and ATP production and it appears to be particularly important under stress (359). A small number of thermogenic plants use AOX in order to generate heat from respiration (228, 370, 377); however, given the limited range of plants possessing this capacity, the function of AOX should differ from this in most species. Specific AOX gene family members are strongly induced at the transcript and protein level by complex III or complex IV dysfunction (167, 361), suggesting that AOX expression is highly responsive to insufficient cytochrome pathway capacity downstream of the ubiquinone pool. However, AOX is also commonly induced by complex I dysfunction and by other disruptions in respiratory metabolism such as the inhibition of ATP synthase, uncoupling of the mETC, and inhibition of the TCA cycle (164, 362). However, since some other studies reported no change in AOX levels in response to dramatic changes in the ETC (319), it would seem reasonable to conclude that the expression level of AOX is governed by multiple and complex signals from the mETC (359). Isotope discrimination studies have revealed that AOX activity does not correlate directly with protein abundance (369) and that it is subject to a wide range of post-translational modulations. First, the plant AOX is embedded within the inner mitochondrial membrane as a homodimer that is either non-covalently linked (the reduced, active form) or covalently linked by a regulatory disulfide bond between the two monomers (the oxidized, inactive form) (354). The responsible residue, a conserved cysteine toward the N-terminus and exposed within the matrix, is responsible for the formation of this disulfide bond (364). Reduction of the disulfide bond is facilitated by the oxidation of specific TCA cycle substrates and, based on the substrate specificity, it is hypothesized that specifically NADPH provides the reducing power for this regulatory reduction (360). Furthermore, a mitochondrially localized Trx has been demonstrated to be able to reduce this disulfide bond (108). Once reduced, AOX is sensitive to activation by specific organic acids, most notably pyruvate (229, 260, 360). Recent studies indicate that this effect is due to the enzyme's apparent Vmax resulting from the ability of pyruvate to stabilize the active AOX (54).
L-galactono-1, 4-lactone dehydrogenase, the terminal enzyme of ascorbate biosynthesis is associated to the cytochrome pathway although its precise localization was unclear (28, 227). A recent study showed that it is a component of Complex I (305), while reverse genetics and inhibitor studies have indicated the responsiveness of ascorbate biosynthesis to inhibition of respiration (227, 255). This link is rather intriguing, as it may represent a powerful manner by which to integrate organellar energy production; however, recent observations summarized earlier suggest that it is far from fully understood.
In addition, an input from a mitochondrial ubiquinone-reducing glycerol-3-phosphate dehydrogenase has been characterized in plants (310, 311). Thus, as in animals, it seems likely that a glycerol-3-phosphate shuttle exists in which an NADH-dependent glycerol-3-phosphate dehydrogenase produces glycerol-3-phosphate which permeates the outer mitochondrial membrane and gets oxidized by the ubiquinone reducing glycerol-3-phosphate dehydrogenase is present on the outer surface of the inner mitochondrial membrane with the dihydroacetone phosphate formed being recycled back to the cytosol (46, 129). However, despite the presence of the required enzymatic complement in plants, as yet no transporter of glycerol 3 phosphate has been identified. Furthermore, transcript levels of components of the putative glycerol 3 phosphate shuttle neither correlate with each other nor correlate with the NADH dehydrogenases (277); however, as the authors of this analysis suggest, it is conceivable that their co-regulation occurs at the enzyme level.
A third recently uncovered route of electron donation is that afforded by the ETF complex which was first discovered in plants after transcript profiling of senescent plants (47) and subsequently directly demonstrated to be functionally linked to branched chain amino-acid and lysine degradation by the isovaleryl and hydroxyglutarate dehydrogenases, respectively (6, 155). In utilizing this system, the plant cell considerably increases the energy efficiency of protein degradation by both providing substrates for the TCA cycle and directly donating electrons to the mETC (10).
Since genotypes of Arabidopsis deficient in the expression of at least some of the proteins involved in these alternate electron donor systems exist (6, 113, 155, 205, 310, 311, 330), future studies should be able to assess the hierarchy of their contribution to respiration across a range of environmental conditions.
Redox status signaling and other mitochondrially emitted signals
Given the extensive discussion of the metabolic control of the major redox systems and the fact that mitochondrial redox signaling has been recently comprehensively and expertly reviewed (309), we will only highlight a few brief aspects of this vast topic here and refer the reviewer to this and other previous reviews (93, 334) for details. Similar to the plastid, the mitochondrion originated from a bacterial symbiosis and still harbors its own genome. As such, redox status signaling from the mitochondria alongside other has wide implications. Although not traditionally regarded as major sources of ROS in leaves, it is well known that reactions associated with Complex I and Complex III produce superoxide (233) and as mentioned earlier, many mitochondrial enzymes are highly susceptible to oxidative damage. Even though on a cellular basis mitochondrial ROS production in the illuminated leaf is relatively modest, this does not preclude a role in setting the cellular redox status, particularly given that the ROS detoxification capacity is relatively small in comparison to that housed by the plastid or peroxisome (93). However, similar to the other organelles, mitochondria house both enzymic and non-enzymic antioxidants (157, 278), including a Trx system. While retrograde signaling from mitochondria is far less well characterized than its plastidial counterpart with no single pathway being established thus far (309), several candidate redox-related signals have been proposed, including superoxide, NO, H2O2, ascorbate, glutathione, lipid peroxide, peroxynitrite, lipoic acid, cysteine, NAD+/NADH, and oxidized proteins [for a detailed discussion, see Ref. (309)] as well as TCA cycle organic acids (88). The superoxide radical anion is not membrane permeable and has a very short half life in the mitochondrion. Hence, a potential superoxide sensor would need to reside in close proximity to the location of its generation, particularly given the high activity of Mn-superoxide dismutase. However, several mitochondrial proteins, including aconitase, are highly susceptible to oxidation by this radical (240); therefore, a possible signaling role for it should not be dismissed too quickly. The possibility that mitochondrial NO signaling plays an important role in cellular co-ordination is reasonably supported by experimental evidence, despite the fact that a classical NO synthase has as yet not been identified in plants (122). NO inhibits aconitase and via nitrosylation glycine decarboxylase and constituents of the mETC (122, 263). It furthermore reacts with superoxide to form peroxynitrite, which can influence tyrosine-dependent kinase signaling pathways (357). Little is currently known with regard to mitochondrially initiated H2O2 signaling. As mentioned earlier, ascorbate is produced in the mitochondria and has strong effects on the transcription of a wide range of nuclear and photosynthetic genes (174, 316, 355), and recent studies imply important roles of ABI4 and the cullin protein degradation network in setting or mediating this signaling system (172, 374). While synthesized in the plastid rather than in the mitochondria, the mitochondrial pool of glutathione is particularly large, stable, and independent from variations in cellular levels (390). This pool is, however, sensitive (in terms of oxidation) to changes envoked by mutations in either the mETC or other elements of the mitochondrial antioxidant system (75, 87, 240, 396), suggesting that it may work as a good signaling molecule most likely exerting its effects by oxidation of nearby protein thiols. Lipoic acid and free cysteine are proposed to signal in an analogous manner (309). Protein oxidation could be mediated by a broad number of different systems, including peroxidases, Trxs, and glutaredoxins, with the evidence for the importance of each of these being critically discussed by (309). Finally, returning to molecules intimately related to the TCA cycle, both NAD+/NADH and the TCA themselves need to be considered in the context of mitochondrial signaling. We have discussed the regulation of mitochondrial NAD+ levels in detail earlier. Further to this, it is intriguing that the tobacco cytoplasmic male-sterile (CMS) II mutant revealed a massively altered cellular redox status homeostasis as a consequence of an impaired function of Complex I (75). In mammals, sirtuin type protein deacetylases act as metabolite sensors for NAD+ (367). The study of sirtuins in plants, alongside that of mutants of the recently identified mitochondrial NAD+ transporter (264) will be instrumental in furthering our understanding as to the extent of NAD+ signaling in plants. Similarly, poorly characterized as yet in plants are carboxylic acid signaling pathways, although first works explicitly addressing this are now beginning to be published (88). Earlier studies focused on using the AOX transcript as a marker for such retrograde signaling due to the large increases in the expression of this gene after incubation with exogenous carboxylic acids (120, 260). More recent studies demonstrated that a similar response can be achieved by a non-metabolizable analog of citrate, suggesting specificity and that this response was not abolished in known hormone signaling mutants (88). However, as for all of the candidate signals, far more work will be needed to firmly establish the underlying reception and relay pathways. To summarize, signaling from the mitochondria is likely to be a highly complex network of overlapping responses. Once one of these pathways is fully elucidated, it would seem likely that we will be able to get a better handle on the others and finally an understanding of the hierarchy of their control and influence.
Peroxisomal Redox Biology
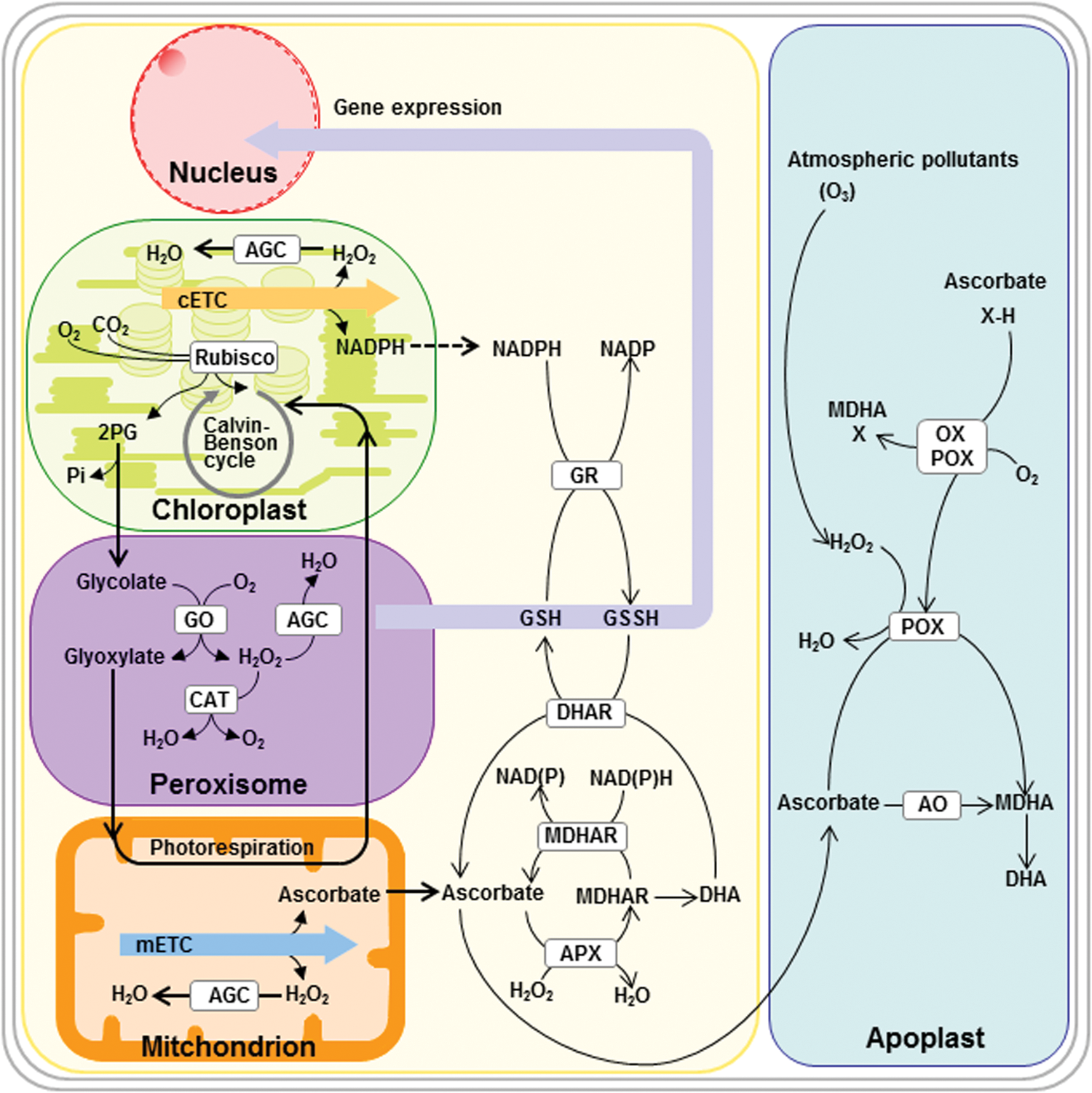
Although emerging roles for peroxisomes include participation in the biosynthesis of the plant hormone auxin and the signaling molecule jasmonic acid (JA) and role in sulfur and nitrogen metabolism (147, 182, 251, 284, 329, 394), the defining role of this organelle, at least in photosynthetically active tissues, is participation in photorespiration and concomitant production of H2O2 (Fig. 6). Indeed, a large proportion of proteins identified in peroxisomal proteomic studies are identified as reductases or dehydrogenases [see Ref. (147) and references therein], reflecting the importance of this organelle in cellular redox balance. The β-oxidation of fatty acids, as well as the role of peroxisomes in JA and auxin metabolism, is dependent on the redox status aspect of the peroxisome; while its role within photorespiration is also intimately related to the redox balance (31, 95, 147). For this reason, we will briefly describe the operation of the steps of each of these metabolic pathways, which is confined within the peroxisome before detailing their metabolic interaction with other compartments and signaling aspects of peroxisomally generated redox species.
β-oxidation of fatty acids
The β-oxidation of fatty acids is an essential process in the mobilization of the oil reserves that seeds lay down during development, predominantly as triacylglycerol, in order to support post-germinative growth during seedling establishment (76, 118, 171). Fatty acids are transported into peroxisomes by a peroxisomal ABC transporter protein variously known as CTS, PXA1, or PED3 (91, 133, 208, 394). Subsequent β-oxidation is particularly important for initial stages of germination (16) as well as in reserve mobilization during developmental or dark induced senescence (72, 188, 315). The first step after import into the peroxisome of straight chain saturated fatty acids is oxidation catalyzed by a family of acyl-CoA oxidases (173) that are linked to flavin adenine dinucleotide and pass electrons to molecular oxygen to produce H2O2. Next, multifunctional proteins containing both hydratase and dehydrogenase domains act on the resultant 2-trans-enoyl CoA (12). Finally, the product of these reactions, 3-ketoacyl Co A is cleaved by thiolase to produce acetyl-CoA and acyl-CoA (109, 296). The β-oxidation of unsaturated fatty acids is more complicated and is reliant on the activity of accessory enzymes to convert these metabolites into suitable substrates for the pathway described earlier (116). However, essentially it operates in a very similar manner. The acyl-CoA oxidase reactions described earlier generate considerable amounts of H2O2, the majority of which is metabolized by catalase; however, when production is excessively high, for example during the mobilization of TAG that occurs during seedling establishment, membrane-bound ascorbate peroxidase and monodehydroascorbate reductase also operate to prevent leakage of H2O2 into the cytosol (77). In addition, an active ascorbate/glutathione system has been reported to help maintain redox balance in the peroxisome of pea leaves (157, 220). The acetyl-CoA produced during β-oxidation is respired by either mitochondria (189) or feeds into the glyoxylate cycle in which it is converted to succinate and malate and used for gluconeogenesis (274). During β-oxidation, the operation of the hydroxyacyl-CoA dehydrogenase activity produces NADH. The reoxidation of the co-factor, and by implication continued activity of the process, therefore, depends on a malate-OAA shuttle involving peroxisomal and cytosolic isoforms of MDH and operating in a similar manner to that defined earlier in the mitochondria. Intriguingly, mutants deficient in both peroxisomal isoforms of MDH germinate but are dependent on sucrose for establishment and mobilize TAGs at a considerably slower rate (273).
Hormone biosynthesis
Recently, redox status links to hormone biosynthesis and modification have been uncovered largely due to the clarification of the structure and localization of these pathways. The JA hormone family comprises JA itself, derivatives of JA, and its precursor 12-oxo-phytodienoic acid. The latter is produced in the chloroplast from which it is released by an as yet undefined mechanism (1). After the uptake of 12-oxo-phytodienoic acid into the peroxisome, it is reduced to 3-oxo-2-(29-[Z]-penenyl)cytopentane-1-octanoic acid (62, 300, 325). This product subsequently undergoes three rounds of β-oxidation to form JA (65, 175, 182). Importantly, 12-oxo-phytodienoic acid regulates seed germination with severe mutants in core β-oxidation function only being able to germinate if the testa is manually ruptured (90, 271). Evaluation of a range of peroxisomal uptake and activity mutants has recently indicated that the peroxisomal import and metabolism of 12-oxo-phytodienoic acid are important for germination (68). Similarly, the conversion of IBA to IAA occurs in the peroxisome. IBA is structurally similar to IAA, the principal form of auxin, and is known for efficacy in root induction (387). However, it is actually a protoauxin that is transferred or stored without auxin activity (314, 327). The predicted pathway for the metabolism of IBA also parallels β-oxidation of fatty acids, but its exact constitution has not yet been clarified [see Refs. (147, 328, 395) for details].
Photorespiration
Whilst the pathways described earlier clearly play highly important roles in seedling establishment as mentioned earlier, the four compartment-spanning pathway of photorespiration is the major role of the peroxisomes in photosynthetically active tissues (32, 147). Peroxisomally localized enzymes include glycolate oxidase (124), the catalase (220) serine-glycolate and glutamate-glycolate aminotransferases mentioned earlier (152, 202), the major isoform of hydroxypyruvate reductase (351), and a considerable MDH activity (64), rendering the peroxisome a major site of photorespiration (32). Glycolate is formed in the plastid via the dephosphorylation of 2PG (32), the product of the oxygenase reaction of Rubisco. It is exported to the cytosol by the glycolate/glycerate transporter (269) and is considered, by many, to simply diffuse into the matrix of peroxisomes (147). There, glycolate is oxidized, yielding H2O2 and glyoxylate as products (124, 295). As detailed earlier, the mitochondrial glycine decarboxylase decomposes glycine to CO2, NH3, and NADH transferring a C group to 5, 10-methyl tetrahydrofolate in the process (60); while SHMT attaches this methylene unit to a second glycine molecule to produce Ser, which diffuses back to the peroxisome and is transaminated by the serine-glycolate transferase to yield hydroxypyruvate. This intermediate is reduced by hydroxypyruvate reductase and NADH, which is provided by the operation of the peroxisomal MDH to form glycerate (64, 351). Given the importance of the hydroxypyruvate reductase reaction, a minor isoform of this enzyme is also localized to the cytosol (351). The exact reason for this dual localization is, at present, unknown; however, it has been speculated that it enables greater flexibility under a wide range of subcellular redox conditions (335). Whichever the route of hydroxypyruvate reduction, the glycerate produced is converted via glycerate kinase to the Calvin–Benson cycle intermediate 3PGA (43, 214). Intriguingly, by contrast to its C3 counterparts, the maize glycerate kinase enzyme is redox regulated via an additional C-terminal autoinhibitory domain, which forms a disulfide bridge in the dark, rendering the oxidized enzyme inactive (29). Attempts to optimize the potential of C3 plants by manipulating the photorespiratory pathway have been undertaken. Although promising, these have exposed unanticipated problems in that they exhibit many additional consequences likely due to the fact that the photorespiratory pathway is strongly embedded in both plant primary metabolism and subcellular redox balances (83, 169, 219, 267).
Peroxisomal transporters
Having described the major metabolic routes operating in the peroxisome, an obvious question is how they are sustained with regard to energy, reducing equivalents, and other co-factors. Great strides have been taken toward addressing these questions by the use of proteomics and reverse genetic studies in recent years. Arabidopsis PNC1 and PNC2 proteins are members of the mitochondrial carrier family (263), but they function as adenine nucleotide carriers. Both proteins import cytosolic ATP to support energy-consuming processes such as those regulated by the many kinases identified in the peroxisomal proteome (284, 289), as well as, perhaps more significantly, the pathway of β-oxidation described earlier. Interestingly, simultaneous repression of both genes after an RNAi strategy severely restricted this pathway, indicating the absence of a major, peroxisome-autonomous, ATP-generating system such as substrate level phosphorylation (5, 203). Similarly, the peroxisomal NAD+ transporter, PXN, is a highly abundant peroxisomal membrane protein (82, 98) that was additionally isolated in a screen for mutants of abnormal peroxisomal morphology (211). Despite having very high similarity to the PNCs, transport studies (39) revealed that it is functionally more closely related to the recently characterized mitochondrial and plastidic NAD+ transporters (264). Given that NAD+ is known to be synthesized exclusively in the cytosol (131, 249), it seems reasonable to assume, on the basis of the kinetic characterization of the recombinant protein (39), that PXN mediates an NAD+ import/AMPexport antiport (147). By contrast to the specific transport of adenylates and NAD+, diffusion of carboxylic acids appears to be facilitated by a peroxisomal pore-forming channel (147, 286). Those from spinach leaves and castor beans are anion selective and enable diffusion of small carboxylic acids such as intermediates of photorespiration, β-oxidation, and the glyoxylate cycle (285 –288). It is likely that further transport functions will be uncovered given that our understanding of peroxisomal metabolism has had to be expanded to include enzymes of the OPP (218), purine catabolism (93, 191), and polyamine metabolism (320).
The transporters described earlier are likely to be vital in co-ordinating not only cellular redox metabolism, but also transmission of redox-based signals. Peroxisomes are known to release signals that regulate nuclear gene expression, photomorphogenesis, plant development, light signaling, and stress responses as well as peroxisomal biogenesis itself (93, 100, 358, 379). The nature of the signals responsible are not fully defined; however, three candidates that are regularly discussed are H2O2, NO, and redox hormone interactions (93). In the case of H2O2, insufficient activity of catalase in C3 plants has been documented, during high rates of photorespiration, and results in a marked accumulation of oxidized glutathione (379). Furthermore, even with a complete complement of catalase, photorespiration-linked alteration of the redox states of ascorbate and glutathione can occur transiently (250). It is possible that the glycollate oxidase reaction acts as an important mediator of H2O2-derived signals (295) as could the superoxide produced on the conversion of xanthine to urate (220) and even the acyl-CoA oxidase (220). It is still unclear how the signal is relayed within the cell with both direct signaling (232) and signaling by oxidized peptide intermediaries (236) being debated. The strongest arguments for the latter theory would be their specificity as well as the fact that a combination of cytoplasmic streaming and degradation of H2O2 would likely render direct signaling unlikely. Indeed, elegant mathematical modeling provides strong support for this theory in that while it demonstrated that signaling over the distance equivalent to that typical between the plasma membrane and the nucleus would theoretically be sustainable tellingly, this requires a much faster enzymatic degradation and a much lower background concentration of H2O2 than has been observed experimentally (234, 368). Another possible signaling candidate is the well characterized, yet somewhat controversial NO (93, 121). This intermediate is produced in peroxisomes by the reduction of nitrite as catalyzed by the peroxisomal enzyme xanthine oxidoreductase—the redox regulation of which can reversibly convert the enzyme function from a xanthine dehydrogenase into a xanthine oxidase. A role for this enzyme in signaling has been postulated in pea (318) as well as on phosphate deficiency in cluster roots of white lupin (Lupinus albus) (373), and there is a large body of evidence that NO has a signaling function in plants (121). The third possibility is that the redox status of the peroxisome interacts with hormone signaling particulary with regard to those phytohormones whose synthesis are closely linked to β-oxidation-like processes in the mitochondria. This possibility has recently been expertly critically discussed in the context of the overlap between peroxisomal function and plant defence (320). In addition to these direct signals, the impact of peroxisomal activity on cytosolic metabolite pool sizes and redox status should also be considered as potential indirect signaling mechanisms by which the peroxisome can influence both nuclear gene expression and, either subsequently or in parallel, cellular phenotypes. As recently discussed (52), the application of integrated functional genomics approaches and modeling will likely enable greater resolution of these aspects.
Cytosolic Redox Biology
ROS have received great attention due to both their inherent reactivity and their potential as signaling molecules (4, 66, 231, 242). That said, despite intense research activity, the exact mechanisms underlying this signaling remain controversial and it is still uncertain whether superoxide, hydrogen peroxide, or singlet oxygen are themselves signaling molecules or whether they rather merely provide an environment that is a suitable conduit for signaling by other pathways (242). The primary targets of ROS signals appear to be amino acids such as Cys and small molecule thiols such as glutathione (66), although their abundance is additionally closely linked to that of the signal molecule nitric oxide (245), rendering the unraveling of the underlying mechanisms even more problematic. The halftimes of ROS turnover are determined by the antioxidant environment, which includes two factors. First, antioxidant enzymes such as catalase in the peroxisome, ascorbate peroxidase in the apoplast, and enzymes of the ascorbate/glutathione cycle in the cytosol (Fig. 6), and second, small-molecule antioxidants such as ascorbate and glutathione as well as the plastidial carotenoids and tocopherols (242). Alongside peroxiredoxins localized in the plastid, cytosol, and mitochondria, these small-molecule antioxidants play a major role in the detoxification of ROS (97). Returning specifically to the cytosol while both ascorbate and glutathione pools have been clearly demonstrated to act as redox buffers, our current understanding suggests that these pools, nevertheless, appear to have quite distinctive functions within ROS signaling. The principal effect of ascorbate has been suggested to be in setting the thresholds for cytoplasmic and apoplastic signaling (242). However, several recent studies provide evidence which would appear to suggest that its role transcends such limitations, suggesting roles in plant defence and maintenance of optimal photosynthesis (172, 255, 374). In contrast, glutathione appears to exert a greater influence via the hydrogen peroxide signaling pathways and is widely regarded as a common arbiter of the intracellular redox potential (94, 242). However, the roles of both cytosolic ascorbate and glutathione are reviewed in detail in several excellent recent articles (94, 96, 242), so we will not dwell on them here.
Relatively little is known about the functional role of cytosolic Trxs, comprising mainly Trx h isoforms (h1-h6 in Arabidopsis), which are reduced by NTRA and NTRB using NADPH as a redox donor (282). The redox status of the NADP(H) system in the cytosol is most likely determined by (i) the activities of G6PDH and 6PGDH regulating NADPH provision by the OPP and (ii) NADP-ICDH catalyzing NADPH metabolism as a part of the amino-acid synthesis pathway. In Arabidopsis, there are six G6PDH isoforms with two (G6PD5 and G6PD6) located in the cytosol, with G6PD5 being insensitive to redox changes, while G6PD6 is subject to inactivation by oxidation (372). The latter has been found to be regulated by protein phosphorylation with cytosolic glycogen synthase kinase 3 leading to its activation (67). Glycogen synthase kinase 3 might play a crucial role in counterbalancing oxidative inhibition of G6PD6 by phosphorylating and thereby enhancing cytosolic G6PD6 activity. Intriguingly, Arabidopsis plants overexpressing glycogen synthase kinase 3 have increased G6PDH activity and lower levels of ROS in response to stress and are more tolerant to salt stress (67). In confirmation to this, changes in G6PDH expression in the cytosol of transgenic tobacco plants provide evidence for a role of cytosolic G6PDH in biotic and abiotic stress tolerance (301).
While the redox status of the cytosol seems to be well buffered under normal conditions, large changes in the cytosolic redox balance can occur in response to environmental stress conditions, such as hypoxia. Oxygen deficiency inhibits respiration and leads to a subsequent increase in the NADH/NAD+ ratio in the cytosol. The resulting decrease in NAD+ recycling is critical for the operation of glycolysis, as it will limit NAD-GAPDH activity. Plants, therefore, respond to low oxygen concentrations by inducing hypoxic genes involved in fermentative pathways in the cytosol, which convert pyruvate to lactate, ethanol, or other products to recycle NAD+ from NADH to enable glycolysis and its attendant ATP production to proceed (102). There has been rapid progress in our understanding of the underlying low oxygen sensing and signaling pathways in the last years. Recent reports indicate group VII ethylene response factor (ERF VII) transcription factors such as HRE1, HRE2 (200), RAP2.2, and RAP2.12 (142) as important regulators of hypoxic gene expression and survival in Arabidopsis plants. Intriguingly, it was shown that group-VII ERFs are stabilized under hypoxia and degraded on re-oxygenation via the N-end rule pathway of targeted proteolysis, functioning as an oxygen-sensing mechanism in Arabidopsis plants (110, 199). In animals and plants, N-end rule-regulated proteolysis involves spontaneous or enzymatic oxidation of an exposed Cys to Cys-sulfinate or further to Cys-sulfonate in an oxygen-, ROS-, or NO-dependent manner (365). The role of the redox status of the tissue in modulating oxidation of the penultimate Cys of group-VII ERFs in response to changes in oxygen concentrations has not yet been investigated. Moreover, it will be interesting to study the impact of this oxygen-sensing pathway on adaptive responses in metabolism to balance the redox and energy status of the tissue in response to changes in oxygen concentrations.
Integration at the Cellular Level
Plastid signals play important roles in various cellular processes that are vital to the plant by influencing nuclear gene expression during different stress conditions (24). This includes different signal components related to photosynthetic electron transport, changes of the chloroplast redox state, accumulation of ROS, and metabolic intermediates (i.e., protoporphyrin IX or sugars). These aspects of plastid retrograde signaling to the nucleus have been presented by many excellent recent reviews and will not be covered here (24, 58, 195, 268, 340, 344, 386). However, there are also signals transferred between plastids and mitochondria to coordinate their metabolic activities. In the next section, we will discuss the role of redox status signals involved in the communication between these two organelles.
Turning attention to cellular processes that are regulated by metabolic redox signals emanating from the mitochondria, recent reverse genetic and functional genomic studies have consolidated the role of the mitochondria in optimizing photosynthesis (252) and elucidated roles for the mitochondria in the regulation of normal fruit ripening (55). Considerable cumulative evidence has accrued suggesting a vital role for mitochondrial function during the photosynthetic process (256). Depending on a plant`s developmental stage and/or environmental considerations, reducing equivalents generated by the photochemical reactions accumulate in the chloroplast stroma, causing over-reduction of the photosynthetic electron transport chain and the generation of ROS, leading to photoinhibition (92). It is generally accepted that this excess of reducing equivalents can be dissipated by their export from the chloroplast to the mitochondria via the malate valve (186, 302); once within the mitochondria, these reducing equivalents are oxidized by the mitochondrial respiratory chain (248), thus enabling continued high rates of photosynthesis. However, while the reduced rates of photosynthesis in UCP mutants provided some weak circumstantial evidence for the importance of the operation of the malate valve (337), mutants in the plastidial NADP-dependent MDH essential for its operation revealed that the plant harbors additional mechanisms to protect against photoinhibition (135). A second link between these organelles, one working in the opposite direction, was recently uncovered after the downregulation of the mitochondrial isoform of MDH in tomato (255). These studies demonstrated that plants deficient in this TCA cycle enzyme were able to effectively utilize L-galactono-1,4—lactone as an alternative electron donor to the mETC, resulting in a considerable increase in ascorbate content. Intriguingly clear and direct evidence for such an interaction between respiration and ascorbate biosynthesis was also provided by Bartoli et al. (28). Another consequence of the inhibition of the mitochondrial MDH is that the transgenic plants have enhanced rates of photosynthesis and growth. However, the exact mechanism by which this is achieved is not known. It is speculated that this is a direct result of the upregulated ascorbate biosynthesis (255) likely via one of the myriad of processes by which ascorbate can exert an effect on photosynthesis (316). Perhaps unsurprisingly, this effect seems to be highly environmentally dependent, neither being observed in short day conditions in tomato (256) nor in corresponding mutants in Arabidopsis (352). The role of photorespiration in linking chloroplastic and mitochondrial metabolism and function has also been the subject of multiple studies (32, 84, 349). While we have mainly dealt with it in the preceding sections, one recent study which is particularly pertinent to this discussion is the recent finding that overexpression of the H-protein of glycine decarboxylase considerably enhanced net photosynthesis and growth of A. thaliana (350). At the molecular level, lower glycine levels confirmed elevated glycine decarboxylase activity in vivo, and lower levels of the CO2 acceptor RuBP indicated higher drain from CO2 fixation. Thus, the photorespiratory enzyme glycine decarboxylase in mitochondria appears to constitute an important feedback mechanism that contributes to the control of the Calvin–Benson cycle in the plastid and, hence, carbon flow through both photosynthesis and photorespiration (350).
The second example is likely also to be mediated by the malate valve described earlier but this time in reverse direction that is, the redox status of the mitochondria is transmitted to the plastid and influences plastidial metabolism and function with subsequent whole cell and, subsequently, whole organ consequences (Fig. 7). It was again, at least partially, uncovered by the suppression of the expression of mitochondrial MDH alongside independent suppression of fumarase in tomato, but this time in a fruit-specific manner (55). Detailed characterization suggested that, although the rate of ripening was essentially unaltered in these lines, there were minor changes in the accumulation of pigments which are modulated in a redox-mediated manner (154, 243, 382). Furthermore, lines containing higher levels of malate were characterized by lower levels of transitory starch and lower soluble sugar content at harvest, whereas those with lower malate contained higher levels of these carbohydrates. Analysis of the redox-activation state of AGPase revealed that it correlated with the accumulation of transitory starch (55). Intriguingly, most likely as a consequence of the altered sugar content, these lines were characterized as having elevated or reduced shelf life and inversely reduced or elevated susceptibility to post-harvest bacterial infection. More recent studies have revealed that the operation of this reverse malate shunt is context dependent and does not work in fully heterotrophic tissues such as the potato tuber (339), but that similar effects can be brought about by altering the malate-related redox status balance in other compartments of the tomato fruit (261).
In this review, we have discussed recent developments with regard to the redox status control of metabolism and the metabolic control of redox status, at both the organellar and cellular levels. It is becoming clear that redox status plays a central role in sensing physiological and environmental alterations within chloroplasts and mitochondria. These organelles respond actively to these inputs by regulating their own central metabolic processes via internal redox systems and by emitting interorganellar redox signals to maintain the homeostasis at a cellular level. Further work is required to resolve the network of intra- and interorganellar redox signals. This will require genetic and metabolomics approaches (52, 346) as well as the application of techniques to analyze redox states at the subcellular level (187, 309). Such methodological advances will be imperative to get a comprehensive understanding of how compartmental issues affect both the metabolic control of redox and its corollary, the redox control of metabolism.
Footnotes
Acknowledgments
This work on redox has been supported by the Deutsche Forschungsgemeinschaft (grants Ge 878/5-1 & 8-1 to P.G. and the Research Unit Promics FOR 1186 to A.R.F.).