
Abstract
Introduction
P
Several centuries later, we are looking specifically at the redox components and mechanisms that orchestrate plant growth and development in an effort to find answers and solutions, which are urgently needed for our voyage into the uncertain future in which population growth and climate change will inevitably threaten food security. Here, we review plant redox systems that have already served or might be exploited in the future as a basis to engineer crops with enhanced resistance to adverse environmental conditions, to develop efficient strategies for weed control, and to provide means to increase food security while combating nutritional deficiencies. We primarily focus on strategies that are already used in agriculture or have the potential to be adopted in the near future and discuss the science behind these technologies.
Generation of Reactive Oxygen Species in the Plant Cell
Today, the majority of living organisms use oxygen (O2) as a final electron acceptor to sustain their energy levels. This requirement is a direct consequence of the accumulation of oxygen in the primordial atmosphere mediated by photosynthetic organisms able to split water into O2, electrons, and protons (39). The reactivity of O2, however, entails the formation of toxic O2 derivatives as by-products of normal metabolism (Fig. 1). These by-products, that is, hydrogen peroxide (H2O2), superoxide radical (O2 •−), hydroxyl radical (HO•), and singlet oxygen (1O2), are commonly referred to as reactive oxygen species (ROS) and their levels have to be tightly controlled to avoid excessive oxidative damage of cellular constituents (Fig. 2).
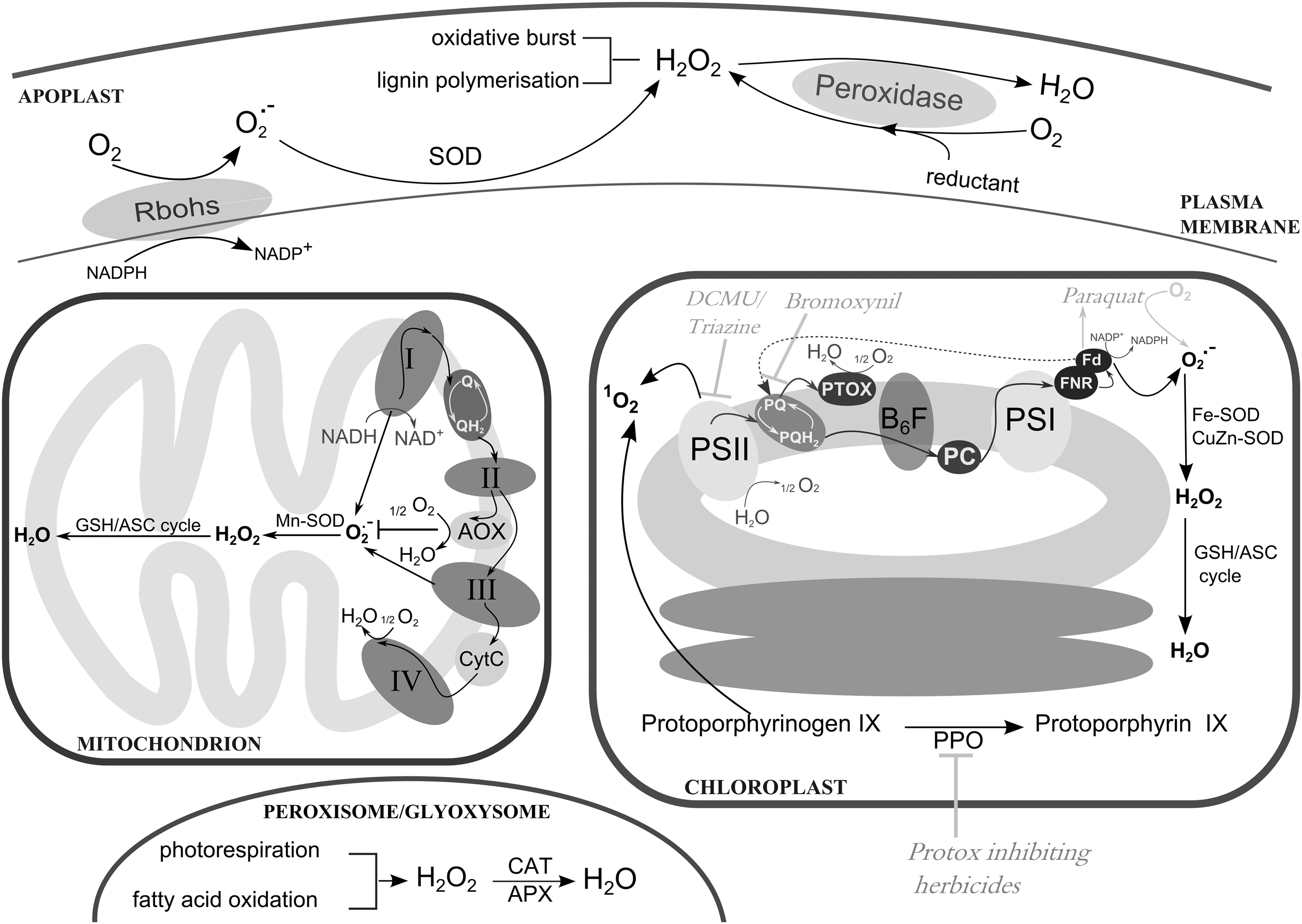
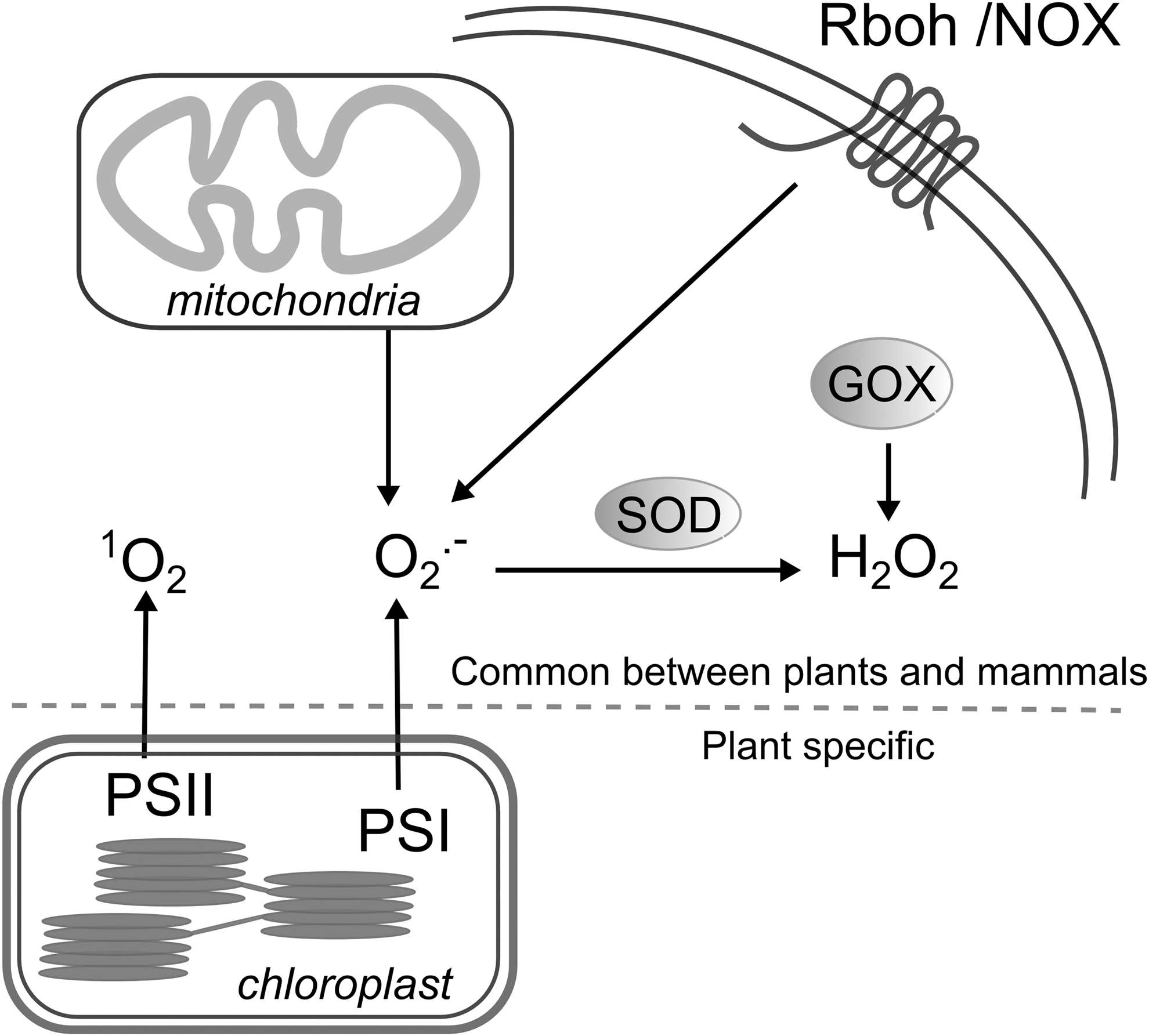
ROS production in plant cells reflects their functional specialization and is largely dependent on the fluctuating external environment. In contrast to animal cells, in which mitochondria are the main sites of ROS production, plant mitochondria are just one of the cellular compartments that contribute to the overall ROS abundance (Fig. 1) (123). In addition, chloroplasts, the plant organelles that harbor the photosynthetic machinery and combine O2 production with active electron transport chains, are the most important source of ROS generation in photosynthesizing tissues (4). Peroxisomal ROS production is also indirectly linked to photosynthesis via the photorespiratory pathway initiated by the enzymatic promiscuity of RuBisCO (16).
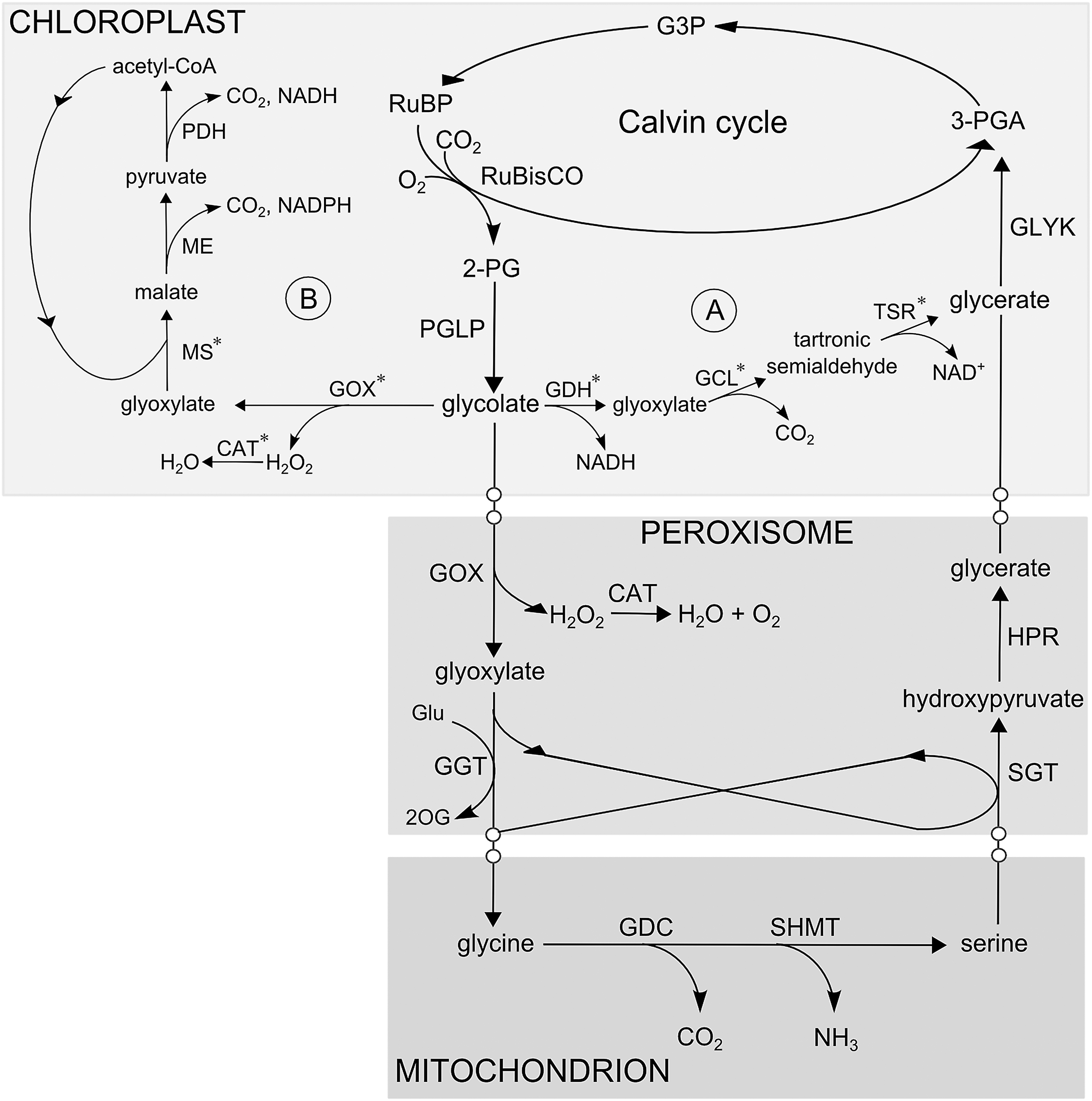
Photorespiratory reactions recycle toxic 2-phosphoglycolate resulting from the oxygenation reaction of RuBisCO via a series of enzymatic steps dispersed between chloroplasts, peroxisomes, and mitochondria (Fig. 3) (11). The peroxisomal glycolate oxidase (GOX) generates substantial amounts of H2O2 that under conditions favoring photorespiration pose a significant load on the cellular redox poise (129).
The enzymatic activities of several ROS-generating enzymes, such as plasma membrane-localized NADPH oxidases and apoplastic peroxidases and polyamine oxidases, are other important determinants of ROS homeostasis in plants (Fig. 1) (2). The presence of NADPH oxidase homologs both in animals (NOX) and plants (Rboh) indicates a common evolutionary origin for these superoxide-producing enzymes. NOX activity in human blood phagocytic cells is crucial for eliminating invading pathogens, whereas plant NADPH oxidases are similarly involved in ROS production in response to pathogens (Fig. 2). Apart from their involvement in defense, different NOX and Rboh family members have been implicated in diverse developmental and signal transduction events mediated through ROS production (185).
The Yin–Yang of ROS Homeostasis: Maintaining the Balance Between Signaling and Oxidative Damage
The accumulation of ROS has been demonstrated not only for multiple stress conditions, such as wounding (114), excess light (45), pathogen infection (79), and low CO2 availability (108), but also during developmental processes like root growth (43), gravitropism (60), and pollen tube growth (61). Furthermore, the production of ROS is an intrinsic feature of multiple cellular processes, such as mitochondrial respiration, photosynthesis, photorespiration, and β-fatty acid oxidation (123).
Under conditions favoring photorespiration, for example, the rate of peroxisomal H2O2 production in photosynthesizing tissues exceeds the contribution of chloroplasts and mitochondria to the overall ROS levels (44). The peroxisomes of germinating seeds, on the other hand, are a significant source of H2O2 generated during fatty acid β-oxidation of mobilized storage lipids (8). Turnover of fatty acids that can be used as an energy source during developmental and also stress-induced senescence has been suggested to enhance the rate of fatty acid β-oxidation, but given the complexity of the senescence process, the impact of fatty acid β-oxidation on ROS homeostasis remains to be deconvoluted (20, 192). Given the central role of lipid metabolism, however, it might constitute a potentially beneficial avenue for crop improvement.
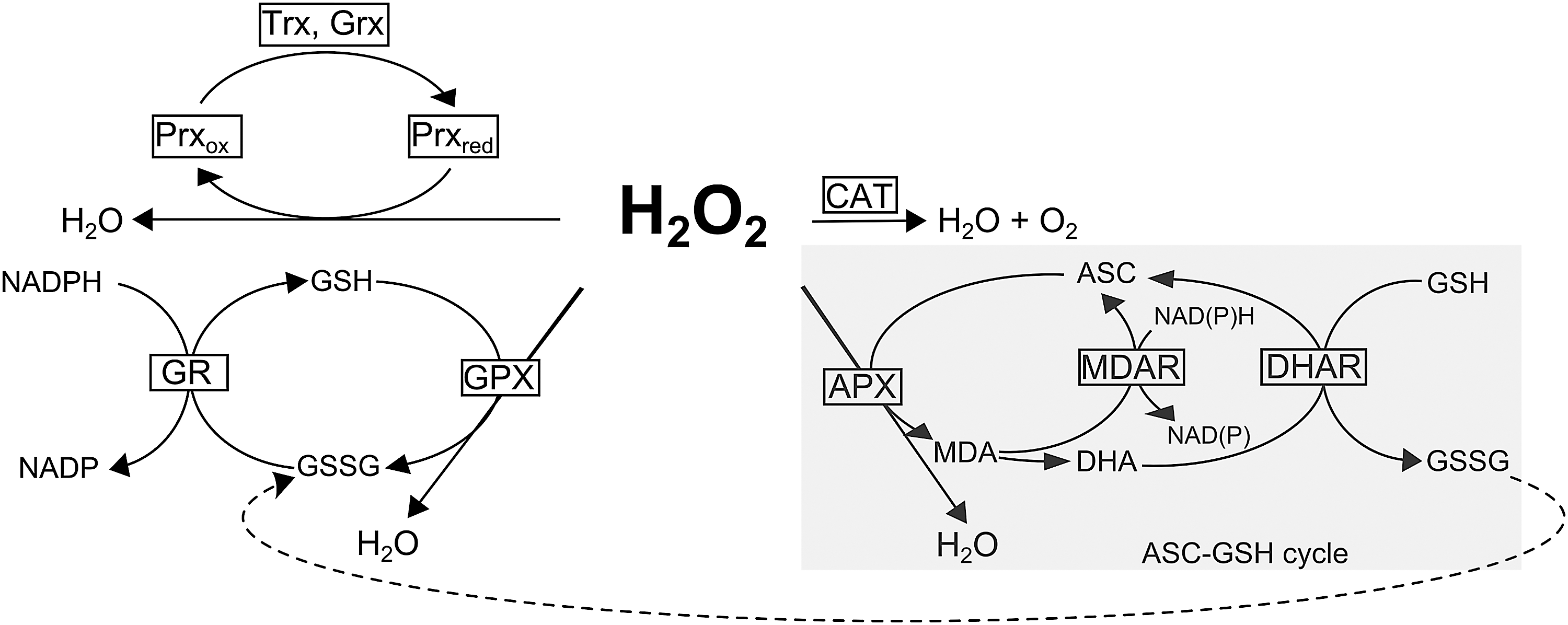
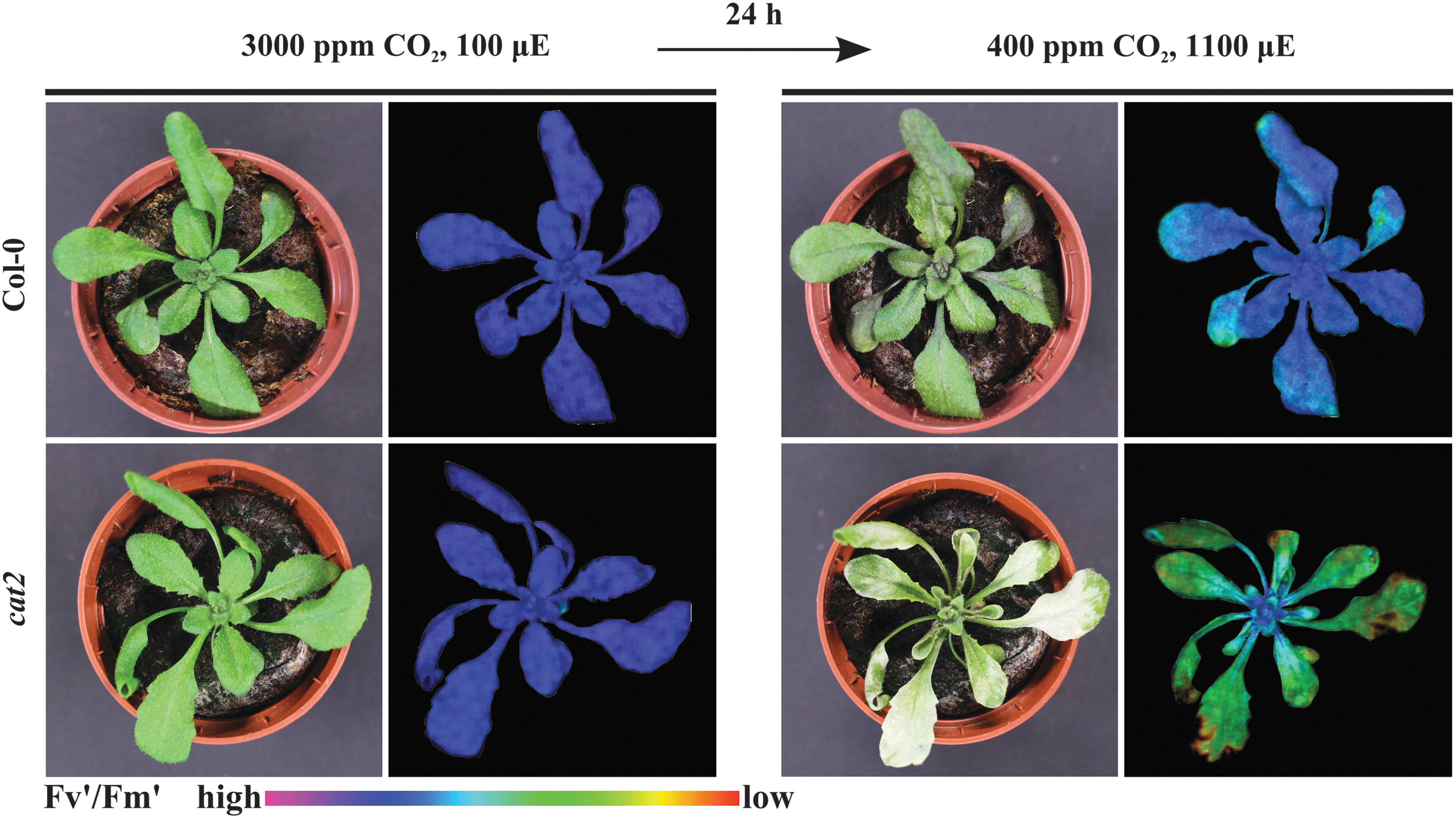
Although frequently reported, obtaining an accurate estimation of ROS levels is inherently limited and such results should be regarded with some caution (128). Often the low methodological resolution obscures the contribution of ROS within individual cellular compartments. It is hard to distinguish between different ROS species and we cannot adequately capture the transient nature of their accumulation (128). To circumvent these limitations, biochemical (144) and molecular markers (46) have been developed with the use of established genetic models exhibiting elevated ROS levels. However, the use of single marker genes does not seem to provide a good proxy for the site and nature of ROS production, therefore the use of ROS signatures has recently been proposed (138). Plants lacking peroxisomal catalase (cat2) (Fig. 4, 5), for example, have been used to deconvolute the transcriptional response triggered by peroxisomal H2O2 (170). In a similar approach, the Arabidopsis fluorescent (flu) mutant, which accumulates the photosensitizer protochlorophyllide, was used to extract an 1O2-related transcriptional (113) and metabolic signature (127). Obviously, cautionary interpretation is necessary as these responses can be influenced by timing, interconversion of ROS, and site of production (68, 143, 155). Additionally, the ability of ROS to differentially oxidize lipids to specific hydroxy fatty acids was exploited to discriminate between peroxidation reactions mediated by individual ROS forms (162). The attack of 1O2 on carotenoids results in the formation of oxidation products, such as β-cyclocitral, which might not only serve as indicators of ROS accumulation but also act as potent signaling components (130).
Oxidation of amino acid residues by ROS serves as a crucial factor governing the stability and function of both metabolic enzymes and signaling proteins (57, 142). Thus far, the best documented evidences of such post-translational modifications are those involving oxidation of cysteine and methionine residues (Fig. 6). In both cases, the targets of oxidation are sulfur atoms found in sulfhydryl (R-SH) and thioether (R-S-R′) groups of cysteines and methionines, respectively (30). Methionine sulfoxide (Met-SO), the oxidation product of methionine, is relatively stable and rarely undergoes further oxidation to methionine sulfone (Fig. 6) (30).

In contrast, the initial oxidation product of cysteine, sulfenic acid (Cys-SOH), is highly unstable and, unless stabilized within the protein environment, can be further oxidized (Fig. 6) either reversibly to sulfinic acid (Cys-SO2H) or irreversibly to sulfonic acid (Cys-SO3H) (31, 137). The reduction of sulfinic acid back to cysteine has been debated, but the existence of an enzyme, sulfiredoxin (Srx), catalyzing this reaction in plants supports this notion (135). The only two known AtSrx substrates (chloroplast 2-Cys peroxiredoxins and mitochondrial PrxIIF), however, question the general occurrence of this process (56, 135).
Alternatively, sulfenic acid might react with free protein thiols to form intra- or intermolecular disulfide bonds or with low-molecular-weight thiols such as glutathione (GSH) leading to cysteine-S-glutathionylation (Fig. 6) (137, 169). S-glutathionylation has long been believed to serve as a mechanism protecting active cysteine residues from overoxidation and permanent protein damage. Only recently, S-glutathionylation gained recognition for its role in redox signaling (195). S-glutathionylation, formation of intra-/intermolecular disulfides, and methionine sulfoxide are readily reversible and their redox status is controlled by glutaredoxins (Grx), thioredoxins (Trx), and methionine sulfoxide reductases (Msr), respectively (Fig. 6) (30, 137).
Apart from methionine and cysteine residues, side chains of other amino acids, such as proline, histidine, arginine, lysine, and threonine, might also be subjected to oxidative modifications leading to the formation of carbonyl derivatives (Fig. 6). Protein carbonylation is an irreversible process caused by hydroxyl radicals (13) generated during metal-catalyzed cleavage of H2O2 (186). Formation of carbonyl groups is widely recognized as a marker for protein oxidative damage and is mainly associated with their proteolytic degradation (58, 110, 179).
Redox Regulation of Plant Immunity
Upon recognition of pathogens, plants trigger a broad spectrum of defense processes, which have been reviewed extensively, and include the hypersensitive response (HR) caused by ROS accumulation. The initial oxidative burst following pathogen infection has been largely seen as mediated by class III cell wall peroxidases and membrane-bound NADPH oxidases (Respiratory burst oxidase homologs, Rbohs) generating H2O2 and superoxide radicals, respectively (29, 161). The precise mechanism of the peroxidase-dependent oxidative burst is far from elucidated and is likely to require reductants such as phenolic compounds and NADH. Upon infection, the ROS profile is governed by the lifestyle of the pathogen. Avirulent species induce a rapid transient ROS formation, followed by sustained higher amplitude of oxidative burst associated with HR and defense (91, 107). In contrast, during virulent and symbiotic interactions, only the initial ROS formation is detected (91).
Plants with compromised Rbohs and apoplastic peroxidase activities display altered resistance to pathogens, exemplifying the important role of ROS accumulation during plant–pathogen interaction (6, 194). One member of the Rboh protein family, RbohD, is further required for triggering and propagating a systemic signal initiated by various biotic and abiotic stimuli (98). The effective long-distance communication depends on apoplastic ROS accumulation along the signal path since the application of the NADPH oxidase inhibitor, diphenylene iodonium (DPI), is sufficient to block further spread of the signal. Interestingly, rbohd mutants are susceptible to green peach aphid (Myzus persicae) attack, implying a functional role for RbohD and systemic ROS signaling during plant–aphid interactions (98). Other invasive species, such as the parasitic nematode Heterodera schachtii, similarly trigger Rboh-dependent ROS production (145). Infected plants lacking RbohD and RbohF display increased cell death lesions in comparison with the wild type.
Despite the primary role of Rbohs in ROS generation during plant defense, emerging evidence points toward alternative ROS sources that might be activated under such scenarios and function independently from Rbohs. Peroxisomal GOXs, for example, modulate the outcome of the nonhost resistance in tobacco and Arabidopsis, with plants lacking GOX activity being more susceptible to pathogens (136). Moreover, silencing of RbohD within the gox mutant background indicated independent roles in plant defense because lack of RbohD had largely no additive effect on bacterial growth in the respective double mutants (136).
The highly reactive nature of ROS is capable of imposing a direct cytotoxic effect on invading pathogens (121). For example, the virulence of Ustillago maydis (the causing agent of corn snut) depends on its ability to detoxify plant-generated ROS (99). A U. maydis strain lacking an ortholog of the Saccharomyces cerevisiae YAP1 (Yeast AP-1-like) transcription factor, which controls the response toward oxidative stress conditions (33, 34), is more susceptible to H2O2 produced by NADPH oxidases during oxidative burst (99). In contrast to biotrophic pathogens, necrotrophs utilize dead plant cells as source of nutrients and often induce ROS production during the infection process to stimulate the HR-related cell death (49).
Salicylic acid (SA) has a prominent role in plant–pathogen responses through a coordinated action of the NONEXPRESSER OF PR GENES 1 (NPR1) transcriptional coactivator and its interacting transcription factors from the TGA family (Fig. 7). The molecular properties of both NPR1 and its TGA family interactors depend on changes in the cellular redox status that occur upon pathogen infection (104, 153). Under nonstressed conditions, NPR1 oligomers are stabilized by intramolecular disulfide bonds involving two cysteine residues (Cys82 and Cys216) and localized to the cytoplasm (Fig. 7). SA triggers fluctuations in the cellular redox status, which lead to a Trxh3/h5-dependent reduction of these disulfides, NPR1 monomerization, and subsequent nuclear import (104, 153). This process is in direct competition with the oligomerization and disulfide formation induced by the S-nitrosylation of Cys156 (Fig. 7).
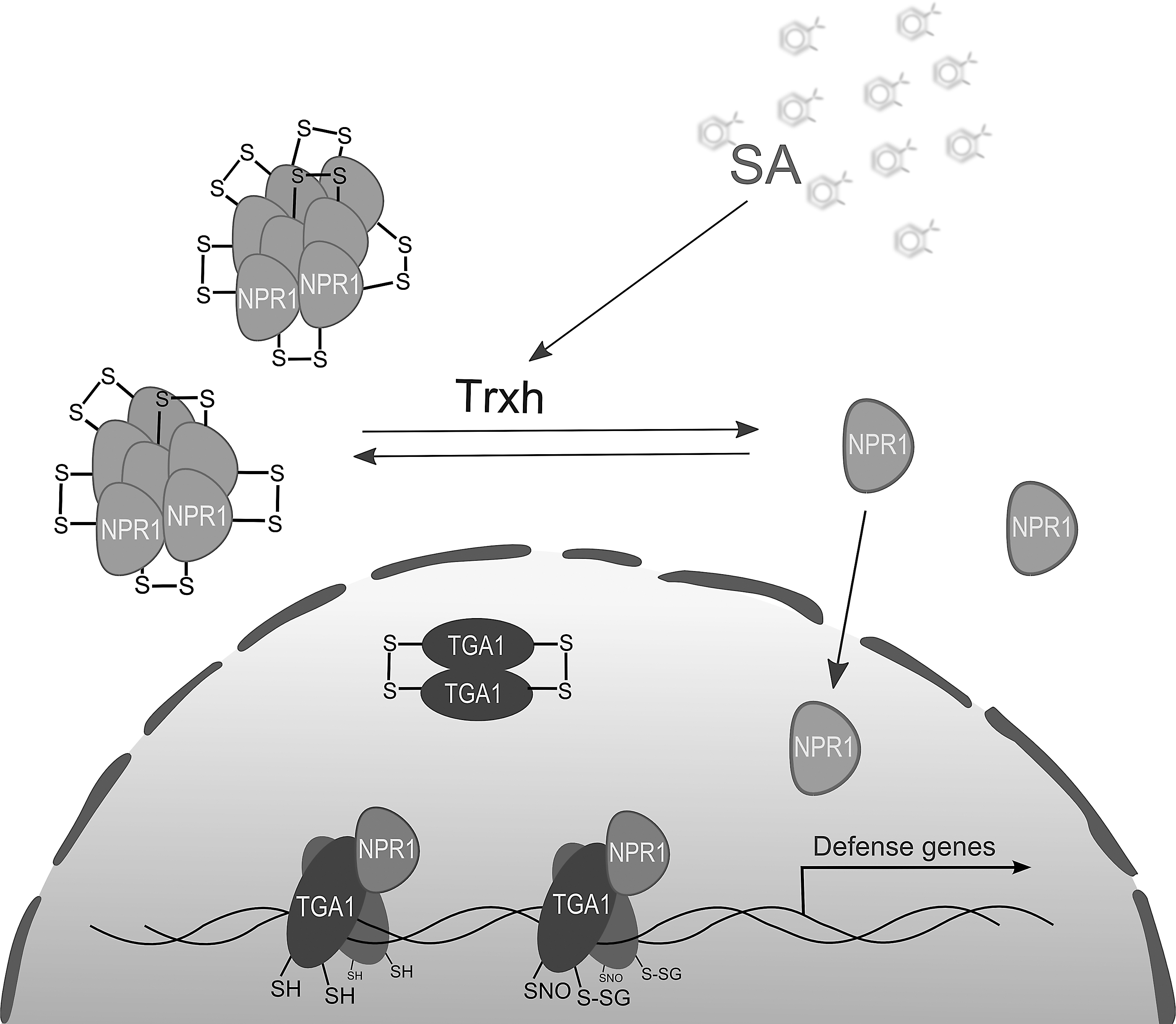
The interplay between these two modifications tightly controls NPR1-dependent signaling (153). Upon nuclear import, NPR1 interacts with TGA basic leucine zipper transcription factors to promote the transcription of pathogenesis-related (PR) genes. Clade 2 TGA transcription factors (TGA2/5/6) act as constitutive repressors of PR transcription. Binding of NPR1 to TGA2 results in coactivation of transcription and depends on NPR1 Cys521 and Cys529 required for binding of SA via the transition metal, copper (190). The interaction of NPR1 with clade 1 TGA transcription factors (TGA1/4) relies on their redox status. When oxidized, TGA1 and/or TGA4 form an intramolecular disulfide bridge (Cys260-Cys266) that hinders binding. Reduction of this disulfide stimulates the interaction with NPR1 and subsequent binding to the as-1 element for the transcription of PR genes (35). Recent data indicate that the two remaining TGA1 cysteine residues, Cys172 and Cys287, are also involved in regulatory disulfide formation (83). Furthermore, all four Cys residues were found to undergo S-nitrosylation/S-glutathionylation after treatment with GSNO (Fig. 7). These modifications serve to protect the Cys residues from disulfide formation and increase the DNA-binding activity of TGA1 (83).
The recognized role of ROS accumulation for an effective immune response was harnessed in a technology that provides resistance to the economically important pathogen, Sclerotinia sclerotiorum (WO199904012; 15). Transgenic sunflower, canola, and soybean plants overexpressing the H2O2-producing enzyme, oxalate oxidase, displayed an improved performance after infection under field conditions (WO199904012; 15). Moreover, introducing the transgenic trait into conventional pathogen-tolerant cultivars had a synergistic effect. Boosting H2O2 levels by introducing glucose oxidase (GO) from Aspergillus sp. also positively affected plant immunity and led to broad-spectrum resistance against various fungal and bacterial pathogens (WO1995014784; 76, 189). For example, potato tubers with enhanced GO levels displayed tolerance against Erwinia carotovora, an economically important postharvest pathogen causing potato soft rot (WO1995014784; 76, 189).
Redox Homeostasis in Plant Growth and Development
Until recently, developmental processes have been largely seen as orchestrated by hormonal cross talk and transcriptional regulators (73, 174). The redox control of plant growth and development, however, is now being increasingly recognized in the light of ample evidence for an extensive network of interactions between ROS and hormonal (e.g., auxin, gibberellin, and abscisic acid) signaling pathways (1, 183). For example, the complex machinery involved in auxin transport, perception, and biosynthesis is influenced by the cellular redox homeostasis (158). One of the most strongly induced transcripts after H2O2 treatment of Arabidopsis plants encodes a UDP-glucosyltransferase 74E2 (UGT74E2; 171), acting on indole-3-butyric acid (WO2008155139; 160, 167). Endogenous overexpression of UGTE74E2 in Arabidopsis altered the endogenous auxin pools and led to distinct morphological changes accompanied by enhanced drought and salt tolerance (WO2008155139; 160, 167).
The organ-forming capacity of the shoot and root meristem depends on continuous cell proliferation that is affected by fluctuations of the redox potential (43, 86, 164). The root quiescent center (QC) is maintained in a highly oxidized state that might be linked to the cell cycle arrest observed in the stem cells (59, 84). Moreover, high levels of the lipid peroxidation product, malondialdehyde, were detected in the meristematic tissues, implying increased rates of ROS formation (141). Active rearrangements of the intracellular GSH pools, major water-soluble antioxidants, have been observed during cell cycle progression, with GSH migrating to the nucleus at the G1 stage and oxidizing the cytosol (177, 178).
The pivotal importance of GSH in plant development was further demonstrated by the inability of the GSH biosynthetic mutant, root meristemless (rml), to maintain cell division (175). The low levels of GSH in the rml mutant (less than 5% of the wild-type content) could be mimicked by inhibiting GSH biosynthesis with buthionine sulfoximine (BSO), which arrests root growth as well. The crucial role of redox homeostasis maintenance for proper development is also evident in Arabidopsis mutants lacking NADPH-thioredoxin reductases (NTRs), which display stunted growth phenotypes similar to those observed upon perturbation of GSH biosynthesis (132). These effects are likely to be partially mediated through the perturbation of polar auxin transport since GSH depletion negatively impacts various auxin transporters (70). Supporting this view is the fact that the triple mutant rml ntra ntrb, impaired in GSH biosynthesis and thioredoxin reductase activity, phenotypically mimics auxin transport mutants (10).
The agriculturally important processes of bud dormancy and bud release are also predominantly driven by redox mechanisms [excellently reviewed by Considine and Foyer (25)]. Synchronization of bud burst is of particular importance for economically relevant perennial plants, such as grapevine and kiwi, because it ensures uniform flowering and predictable harvesting times. Current agricultural practices applied to simultaneously break bud dormancy include physical and chemical treatments, such as cyanamide (NH2CN) (53). Transcriptional responses triggered by NH2CN suggest that it might exert its action via the perturbation of the redox homeostasis since numerous NH2CN-induced genes, such as glutathione S-transferases (GSTs), catalase, superoxide dismutase (SOD), ascorbate peroxidase (APX), and glutathione peroxidase, are also responsive to H2O2 (53).
Various transcription factors, important for plant development, are regulated via redox mechanisms (38). PERIANTHIA (PAN/TGA8), a member of the TGA family of transcription factors, controls floral development via an interaction with ROXY1, a floral monothiol glutaredoxin required for petal development (80). Class II, III, and IV members of the plant-specific homeodomain-leucine zipper (HD-ZIP) transcription factor family modulate apical embryo development, meristem function, and epidermal cell fate through the formation of intermolecular disulfides impairing their DNA-binding activity (24, 163). The TCP transcription factors implemented in the control of various developmental processes contain a highly conserved cysteine residue within their DNA-binding/dimerization domain that mediates the oxidative inhibition of DNA-binding via the formation of intramolecular disulfide bonds. Treatments with H2O2 and oxidized glutathione (GSSG) inhibit binding of TCP15 to DNA (176).
Given the tightly regulated redox homeostasis required for plant development, it is not surprising that external treatments resulting in redox perturbations might trigger distinct morphological changes. Both GSH and its biosynthetic inhibitor, BSO, modulate the flowering time of Arabidopsis (WO2001080638; 111, 112). Similar effects were achieved by using H2O2 and the redox regulators, KCN and NaN3 (WO2001080638; 111). Moreover, ROS has been shown to regulate cell elongation (43, 105). In cotton (Gossypium hirsutum), fiber length is an important quality parameter for fiber production that can be modified by altering ROS homeostasis. Exogenous H2O2 and Ca2+ starvation are crucial determinants of early cotton fiber elongation, which is also fine-tuned by the ROS-dependent calcium sensor GhCaM7 (156). The redox requirement of maize anther development has been exploited as a means to control archesporial cell abundance. By lowering the partial pressure of O2 or exposing developing anthers to reducing agents, the inventors achieved a higher pollen production. In contrast, oxidizing conditions led to a lower pollen production or to male sterile plants. Moreover, this approach might be used for the generation of hybrid monocot plants (WO2013123051; 64, 65).
A beneficial impact of ROS signaling on plant growth and development is exemplified by the effect of a class of pesticides called strobilurins. First introduced in the market as fungicides, they quickly attracted attention due to the significantly higher yield observed in strobilurin-sprayed crops that could not be attributed to their fungicidal properties related to the inhibition of fungal mitochondrial electron transport at the level of complex III (9). Even though the exact molecular mechanisms triggered by strobilurins are not entirely elucidated, a transient inhibition of plant mitochondria is believed to alter the cellular redox homeostasis, which causes a long-lasting positive impact on growth (14, 36). Examples of strobilurin compounds in commercially available products that combine both fungicidal and yield-enhancing properties are pyraclostrobin and kresoxim-methyl.
Engineering Stress Tolerance in Plants by Reducing Oxidative Stress
Prolonged drought, heat waves, suboptimal temperatures, and saline soils are among the common environmental scenarios that negatively affect crop yield. Such suboptimal conditions often lead to over-reduction of the chloroplast electron transport chain, resulting in excessive ROS formation (4). Singlet oxygen is typically produced at photosystem II (PSII) following energy transfer from excited chlorophyll molecules to the ground state triplet oxygen (3O2) (72), whereas PSI contributes to the generation of O2 •− via the Mehler reaction (5). Excessive superoxide levels in the chloroplasts are controlled by iron (Fe) and copper/zinc (Cu/Zn) SOD isoforms. Cu/Zn SOD also resides in the cytosol and peroxisomes, whereas manganese (Mn) SOD can be found in both mitochondria and peroxisomes (3). The crucial antioxidant role of SOD in dismutating highly reactive O2 •− into O2 and H2O2 has attracted significant attention as a tool to restrict oxidative damage during stress conditions (WO199002804; 17, 148, 168, 181). Targeting MnSOD, otherwise localized in the mitochondria, to the chloroplasts in rice has resulted in an overall increase of SOD activity and an improved performance under cold and paraquat-induced stress (103).
Stacking of multiple antioxidant enzymes has also been used to engineer transgenic plants with superior abilities to withstand stress conditions. Simultaneous overexpression of APX, dehydroascorbate reductase (DHAR), and Cu/Zn SOD led to an increased survival under multiple abiotic stresses (77). Moreover, driving APX and SOD under the control of the oxidative stress-inducible promoter of the sweet potato peroxidase (pSWPA2) resulted in the conditional activation of the transgenes under ROS-promoting scenarios, such as cold, heat, and salt stress (WO2006054815; 74). Similarly, overexpression of Cu/Zn SOD, cytosolic APX, or their combination conferred higher tolerance to in vitro salt stress in plums (37) and to drought conditions in tobacco (41).
During the evolution of multicellular organisms, the O2 environment has served as a selective force, predominantly retaining O2-resistant cellular components (39, 150). Nevertheless, certain O2-sensitive elements such as iron–sulfur (Fe-S) clusters are still present as building blocks in plant proteins (168). One such example is ferredoxin (Fdx), the major electron carrier protein in chloroplasts, which is reduced at the level of PSI by the electron flow via the chloroplast electron transport chain (184). In a reaction catalyzed by ferredoxin-NADP+ reductase (FNR), the reducing equivalent carried by Fdx is transferred to NADPH, which is required for numerous biosynthetic processes, including CO2 assimilation, among others. Other Fdx-dependent enzymes involved in sulfur and nitrogen assimilation and amino acid and fatty acid synthesis also rely on reducing equivalents carried by Fdx (54). Importantly, light activation of some key enzymes of the Calvin cycle is mediated via the reduction of regulatory disulfide bonds by thioredoxin/ferredoxin-thioredoxin reductase and requires a flow of reducing equivalents through Fdx (142).
Despite its crucial role, Fdx integrity is highly vulnerable and its activity quickly declines under adverse environmental conditions in all photosynthetic species, including microorganisms (159). In contrast, certain eukaryotic algae and all prokaryotes can counteract the negative effects of Fdx depletion by inducing the expression of alternative electron carriers containing flavin mononucleotide (42). These carriers named flavodoxins, however, are not present in the genome of higher plants (197). Despite their absence, many Fdx-dependent plant enzymes can accept electrons from flavodoxin in vitro, reflecting their common ancestral origin. Based on these evolutionary conserved properties, Palatnik et al. (116) designed transgenic plants with enhanced tolerance to oxidative stress-promoting conditions. Expression of chloroplast-targeted cyanobacterial flavodoxin in tobacco was sufficient to compensate for the Fdx decline and to provide protection against adverse environmental conditions, whereas cytosolic expression did not have any beneficial effects (WO2003035881; 116, 159). Plants overexpressing flavodoxin in the chloroplasts sustained predominantly reduced ascorbate and GSH pools under stress and displayed improved performance under heat, cold, drought, and UV radiation. Interestingly, the transgenic plants also accumulated more biomass in the field in comparison with control plants (WO2003035881; 116). Overexpression of flavodoxin has direct implications for disease resistance and was shown to alleviate symptoms of fungal, bacterial, and viral attacks (WO2003035881; 116). Furthermore, bacterial flavodoxin prevented the decline of nitrogen fixation in nodules of Medicago truncatula (32). Since nodules are highly susceptible to drought and high salinity, flavodoxin overexpression might serve as a way to sustain legume symbiotic performance under unfavorable conditions. The broader agricultural application of flavodoxin was also shown in the important crop species, such as rice, corn, alfalfa, sugarcane, cotton, canola, and wheat (WO2013150402; 134). The transgenic plants displayed improved traits, such as increased biomass, higher seed yield, higher sugar content, and enhanced tolerance against various abiotic stresses.
Due to the losses in carbon fixation associated with photorespiratory metabolism, multiple efforts have been focused on reducing the rate of photorespiration in plants. Next to yet unsuccessful efforts toward the modification of RuBisCO oxygenase activity (119), independent strategies targeting downstream photorespiratory reactions have been described. This seminal approach (WO2003100066; 52, WO2011095528; 63, 71) is based on heterologous overexpression of E. coli enzymes constituting the glycolate catabolic pathway (Fig. 7A). In this strategy, glycolate is converted to glycerate directly in the chloroplast, which significantly reduces the photorespiratory flux through peroxisomes and mitochondria. Plants that utilize this photorespiratory bypass exhibited improved biomass production and increased sugar content (WO2003100066; 52, WO2010012796; 62, WO2011095528; 63, 71, 109). Similarly to the first approach, a recent strategy (89) introduced a complete glycolate catabolic cycle in chloroplasts (Fig. 7B). This cycle comprises GOX, malate synthase, and catalase and was designed to metabolize glycolate into malate that is catalyzed by endogenous enzymes into pyruvate with the release of CO2 (WO2009103782; 89, 93). This approach remains controversial since no 3-PGA is recycled and therefore the reaction depletes the intermediates of the Calvin cycle (122). Nevertheless, a growth improvement was reported for transgenic plants generated in this study (89).
Initially associated with responses to high temperature, heat shock transcription factors (HSFs) are currently implicated in various other ROS-promoting conditions (152, 166). Importantly, HSFs play vital roles in the adaptation to oxidative stress (31, 117), which can be exploited in crop engineering. That said, the ectopic expression of HSF3 increases seed yield under control and drought stress conditions and enhances tolerance toward biotrophic pathogens (106). Similarly, the overexpression of sunflower (Helianthus annuus) heat shock transcription factor (HaHSFA9) in Arabidopsis increases tolerance toward oxidative stress and dehydration (126).
In response to drought, plants accumulate abscisic acid (ABA) required for activation of survival programs, stomatal closure, and reduction of water loss (7, 22). Changes of the ABA level are perceived by the PYR/PYL/RCAR (PYRABACTIN RESISTANCE/PYR1-LIKE/REGULATORY COMPONENT OF ABA RECEPTOR) receptor proteins (87, 118). Conformational changes triggered by ABA lead to interaction between the receptors and the PP2C (PROTEIN PHOSPHATASE 2C) protein phosphatase family members, ABI1 (ABA-INSENSITIVE 1) and ABI2, resulting in their inactivation. Both ABI1 and ABI2 have been shown to be negatively regulated by oxidative modifications, which amplify the ABA signaling cascade (Fig. 8) (95, 96). In the case of ABI2, GLUTATHIONE PEROXIDASE 3 (GPX3) relays oxidizing equivalents via thiol–disulfide exchange and inhibits the phosphatase activity of ABI2 (97). The inactivation of PP2C releases SnRK type 2 family of kinases (SnRK2), which phosphorylate transcription factors mediating ABA-related gene expression and ion channels orchestrating stomatal movements (78, 139).
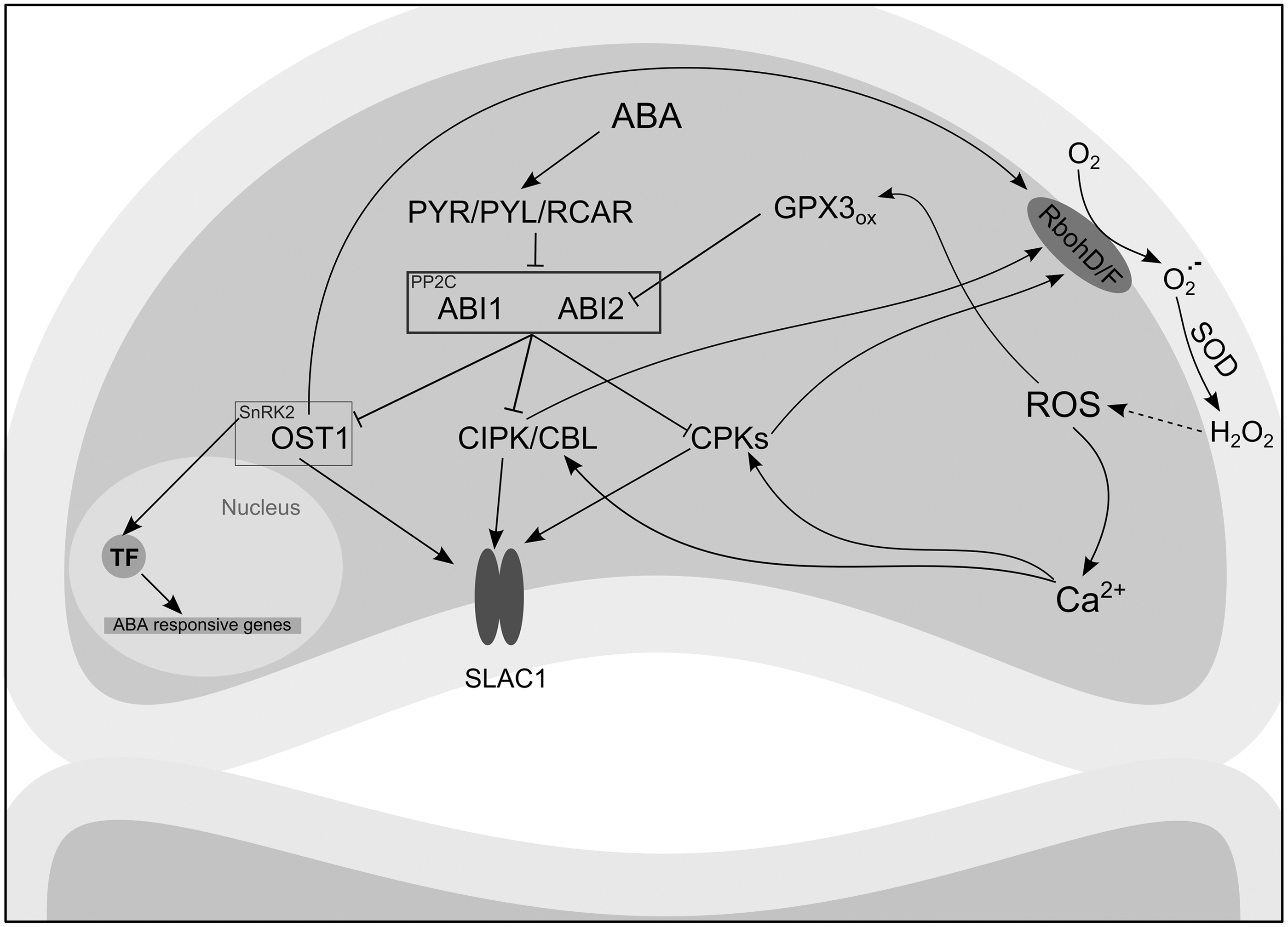
Among the targets of OST1, a member of the SnRK2 family, are the ROS-producing NADPH oxidases RbohD/F (147). Furthermore, OST1 phosphorylates SLAC1 (SLOW ANION CHANNEL-ASSOCIATED 1), which controls the polarity of the guard cell membrane and hence the opening of K+ efflux channels governing stomatal movement (78). SLAC1 is further regulated by phosphorylation mediated by calcium (Ca2+)-dependent protein kinase (CPK) family members upon increase in cytosolic Ca2+ levels (47), which has been shown to be dependent on ABA-induced H2O2 accumulation (120). Several functionally redundant CPKs, such as CPK23, CPK21, and CPK6, are activated by elevated Ca2+ levels (18, 47). Moreover, calcineurin B-like (CBL) calcium sensors, CBL1 and CBL9, in complex with CIPK 23, a member of the CBL-interacting protein kinases family, could also activate SLAC1 (90). Similarly, CPKs and CIPK26 can induce oxidative burst by the phosphorylation of Rbohs (40, 69). Among the upstream SLAC1 regulators, the activity of CPK21 is inhibited by H2O2 following formation of an intramolecular disulfide bond, which can be reduced by Trx h1 (165).
Overexpression of constitutively active ABA PYL4 receptors leads to improved water use, drought tolerance, and enhanced ABA sensitivity (124). Conserved PYL4 homologs exist in many crop species, including maize, rice, barley, soybean, alfalfa, and potato, implying that PYL4 overexpression might be an interesting avenue to engineer drought-resistant crops.
Putative membrane-associated phospholipid hydroperoxide glutathione peroxidase (PHGPx) has been proposed to act as a scavenger of lipid peroxidation products in analogy with glutathione peroxidase that removes excess of H2O2 with the formation of GSSG and water (WO2010108836; 94). Transgenic Arabidopsis plants expressing chloroplast-targeted glutathione peroxidase from S. cerevisiae (YIR037W) displayed an increased yield under water-limiting conditions (WO2010108836; 94). When the protein was targeted to the mitochondria, however, a protective effect was not observed.
In animals, GSH can be degraded by the action of γ-glutamyl transpeptidase (GGT) (82). The Arabidopsis genome contains four putative GGT genes (GGT1, GGT2, GGT3, and GGT4) (151). Knockouts of GGT3 show a reduced number of siliques and a reduced seed yield (92). Expressing γ-glutamyl transpeptidase (slr1269) from Synechocystis sp. in Arabidopsis with chloroplast and mitochondrial signals, negatively affected the photosynthetic performance and plant biomass. However, when the protein was not targeted to any subcellular compartment, the transgenic plants displayed higher biomass in comparison with controls under cycling drought conditions (WO2010108836; 94). An alternative strategy based on boosting cellular GSH levels through the overexpression of the GSH-synthesizing enzymes, γ-glutamylcysteine synthetase and GSH synthetase, enhances the oxidative stress tolerance of lettuce and tomatoes (WO2002033105; 26). Although modifications of the GSH pool might be beneficial in certain cases and are relatively straightforward to generate, it remains to be proven if such approaches can be employed in crop improving strategies. Crucial for the viability of such technologies will be to test their effect in the field under fluctuating environmental conditions.
The pentose phosphate pathway is induced under abiotic stress conditions and generates reducing equivalents (NADPH) and pentose sugars. One of the steps in the nonoxidative phase of this metabolic pathway is a reversible reaction between sedoheptulose-7-phosphate and glyceraldehyde-3-phosphate catalyzed by transaldolase. The reaction products are erythrose-4-phosphate and fructose-6-phosphate, the latter being a glycolytic intermediate linking both pathways. Arabidopsis plants overexpressing bacterial transaldolase A (b2464) performed better under drought conditions. Interestingly, under optimal watering conditions, these plants accumulated higher biomass (WO2010108836; 94).
Glutaredoxins are ubiquitous oxidoreductases that play a major role in the regulation of S-glutathionylation, a reversible post-translational modification affecting protein stability and redox signaling (Fig. 6) (195). Arabidopsis plants expressing PvGRX5, encoding a glutaredoxin from fern Pterris vittata, displayed enhanced tolerance to heat during seed germination and at the mature stage (WO2010021798; 131). Additionally, these transgenic plants were more tolerant to drought, which was associated with reduced oxidative damage of proteins, such as HSP70, OEC33, RubisCO's large subunit, RubisCO activase, and chlorophyll a/b-binding protein. These proteins have previously been shown to be extremely sensitive to oxidation (WO2010021798; 131).
Oxidative Stress-Based Strategies for Crop Protection
Modern agriculture relies heavily on herbicides to control invasive weed species that might severely impair crop yield. Given the high importance of ROS metabolism and their immediate toxicity, not surprisingly, nearly half of the herbicides exert their effect either by directly boosting ROS levels or indirectly triggering ROS overproduction, eventually leading to cell death.
Perhaps, the most notorious example of an ROS-generating herbicide is paraquat. Being among the most toxic and most widely used herbicides for decades, paraquat application is gradually declining because of environmental concerns and tougher regulations (67). It acts quickly and nonselectively by transferring electrons from PSI to O2, which leads to the generation of O2 •− (Fig. 1) (75). The precise mechanism involves the reduction of paraquat by ferredoxin and a subsequent electron transfer to O2 with paraquat being recycled back to its initial redox state, ready to accept a new electron (75). Thus, the potency of paraquat is a direct consequence of the architecture and the reactions executed at PSI, which functions as a plastocyanin–ferredoxin oxidoreductase.
The triazine herbicides are widely applied blockers of PSII with atrazine being possibly the most well-known representative of this class (182). Their physicochemical properties allow them to occupy the plastoquinone-binding site at the D1 protein, a crucial component of PSII (23). Since triazines are nonreducible, the electron flow from pheophytin is blocked, thus preventing dissipation of the excited energy state. The resulting high-energy (triplet) chlorophyll molecules readily form 1O2 and trigger oxidative stress (Fig. 1); 3-(3,4-dichlorophenyl)-1,1-dimethylurea, commonly known as DCMU, is another chemically distinct herbicide that exerts its effect via the same mechanism (157).
The protoporphyrinogen oxidase (protox)-inhibiting herbicides kill plants by generating 1O2 in the chloroplasts (51). Blocking the metabolic flux at this chlorophyll biosynthetic enzyme results in the accumulation of protoporphyrinogen-IX, which is a powerful photosensitizer generating 1O2 in illuminated leaves (Fig. 1) (51). Strategies to develop Protox herbicide-resistant plants have been reviewed previously and include identification and engineering of insensitive plant protoporphyrinogen oxidases (PPOs) or introduction of the Bacillus subtilis PPO coding sequence (US6084155; 81, 180).
Bromoxynil and other structurally related 3,5-disubstituted 4-hydroxy benzonitriles act as PSII blockers at the level of plastoquinone reduction, thus provoking oxidative stress in high light conditions (Fig. 1) (28). The possibility to detoxify bromoxynil via nitrilase-mediated hydrolysis served as the basis to develop bromoxynil-tolerant transgenic canola (Brassica napus). This was achieved by introducing a nitrilase-encoding gene from Klebsiella pneumonia (WO1987004181; 27, 149). The combined application of bromoxynil and bromoxynil-tolerant canola allows an effective control against broadleaf weed species threatening this crop.
Discovered in an effort to generate novel chemical weapons during World War II, 2,4-dichlorophenoxyacetic acid (2,4-D) was quickly recognized as a powerful selective herbicide and is currently one of the most widely used herbicides worldwide (188). 2,4-D belongs to the group of the synthetic phenoxyacetic auxins that largely mimic the effects of the naturally occurring auxin, indole-3-acetic acid. Auxin is often regarded as a master regulator of plant physiology due to its involvement in virtually every aspect of plant growth and development (173). Binding of 2,4-D and other synthetic auxins to the receptor transport inhibitor response, TIR1, and its homologs activates the auxin signaling machinery (66).
At low concentrations, these chemicals stimulate growth, which makes them powerful growth regulators used in agriculture and tissue culture technologies (50). Applied at herbicidal rates, the synthetic auxins cause profound alterations of cellular morphology that eventually result in the disintegration of membrane structures and cell death. These effects are accompanied with increased rates of ROS formation. Thus, ROS-triggered processes, resulting in progressive chlorosis, wilting, and necrosis, mediate cell collapse upon auxin herbicide treatment (50). Recently, metabolic resistance to 2,4-D and structurally related herbicides was engineered by transforming plants with bacterial aryloxyalkanoate dioxygenase (188). Given the low cost and high selectivity of 2,4-D, this technology might play a major role in the production of herbicide-resistant crops in the future.
Biofortification and Food Security
Food shortage and unequal distribution of food supplies pose imminent threats to the well being of billions of people worldwide. Especially vulnerable are poor communities in developing countries relying predominantly on plant-based food. Their diet often cannot deliver the recommended daily vitamin allowances, leading to health problems and even death.
Severe deficiencies of vitamin A take thousands of children's lives each year worldwide (187). In an effort to overcome million personal tragedies originating from a diet poor in vitamin A, researchers have developed a transgenic rice variety, accumulating high amounts of β-carotene (provitamin A) in the rice endosperm (115, 193). This gives the rice seeds a characteristic golden color and hence the name Golden Rice. The provitamin A levels in the optimized Golden Rice 2 allow only 100 g (50 g for children) rice per day to cover the daily requirements of vitamin A (155). The introduction of the carotenoid biosynthetic machinery in the rice endosperm was achieved by combining maize phytoene synthase (psy) with the carotene desaturase (crtI) from Erwinia uredovora (115). Current regulatory measures pause the adoption of the Golden Rice technology in countries that need solutions to overcome nutrient deficiencies.
β-Carotene and other plant carotenoids fulfill important functions in plants (21). By serving as accessory pigments of the photosynthetic machinery, they absorb light and transfer its energy to the photosynthetic reaction centers (146). Moreover, carotenoids scavenge high-energy chlorophyll molecules and 1O2 formed during the photosynthetic activity (125). Oxidation products of carotenoids such as β-cyclocitral and β-ionene are potent signaling molecules that govern plant acclimation to adverse environmental conditions (130).
There is an increasing interest to enrich edible plants with metabolites known to have beneficial health effects. Introduction of only two genes, Delila (Del) and Rosea1 (Ros1), from Antirrhinum majus in tomato boosted anthocyanin production and gave the fruits a deep purple color (19). Del and Ros1 encode basic helix-loop-helix and MYB-related transcription factors, respectively. Anthocyanin functions in plants include among others pigmentation important to attract pollinators, protection against UV radiation, and ROS scavenging (154).
Increasing food production, one of the main goals of the agro-biotech industry, is only relevant if it is adequately used and not wasted. Currently, a significant amount of food is lost due to short shelf life, which restricts its logistical distribution and long-term storage. Tight control of ROS levels has been observed during the early ripening stages of tomato and guava, whereas oxidative stress progressively increased during maturation, implying a major role of ROS in fruit development (100, 101). Moreover, tomato and guava cultivars with a long shelf life displayed higher antioxidant capacity in comparison with cultivars with a short shelf life. Transgenic broccoli expressing isopentenyltransferase (ipt) accumulate antioxidant proteins such as SOD and APX, which is accompanied with delayed postharvest yellowing (85). Taken together, these observations suggest that redox control during fruit maturation and storage could be exploited as a strategy to extend the shelf life. This is supported by the fact that the purple tomatoes described above displayed a prolonged shelf life possibly due to the antioxidant properties of the accumulated anthocyanins (196).
Postharvest physiological deterioration (PPD) observed in cassava roots, a staple food for millions in the tropics, significantly affects their quality within the first 3 days after harvest, making them unpalatable and impossible to sell (133). PPD has been associated with ROS accumulation and increased abundance of various proteins related to GSH homeostasis (133, 172). Delay of PPD and shelf life increase have been observed in cassava cultivars containing higher levels of β-carotene (102). By overexpressing alternative oxidase from Arabidopsis, Liu et al. (85) were able to extend the cassava shelf life to 10–21 days both under field and greenhouse conditions. Control of ROS levels through the overexpression of Cu/Zn SOD and catalase similarly delayed the onset of PPD by at least 10 days (191). Cytosolic glutathione peroxidase overexpressors also displayed postponed PPD symptoms and lower levels of H2O2 (172).
Concluding Remarks
Over the years, scientists have attempted to elucidate the role of ROS in plant development and stress responses. The gathered knowledge has repeatedly been used to improve plant traits, such as biomass, stress tolerance, and nutritional value. We expect that the growing knowledge about ROS homeostasis, perception, and signaling will further stimulate efforts toward improving plant traits and future application of this knowledge into crop species.
Footnotes
Acknowledgments
This work was supported by Ghent University (Multidisciplinary Research Partnership Biotechnology for a Sustainable Economy, Grant 01MRB510W), VIB and Marie Curie (OMICS@VIB PCOFUND-GA-2010-267139), FWO (Fonds Wetenschappelijk Onderzoek–Vlaanderen; Sulfenomics: oxidatieve schakelaars in planten), European Cooperation in Science and Technology (COST Action BM1203 EU-ROS), and the Interuniversity Attraction Poles Program (IUAP P7/29 MARS) initiated by the Belgian Science Policy Office. This work was also supported by the European Cooperation in Science and Research (COST Action BM1203/EU-ROS) by FWO through a Pegasus postdoctoral grant funded to P.K, by IWT through a predoctoral fellowship to B.D.S, and by VIB through a predoctoral scholarship (International PhD Program in Life Sciences) granted to C.W.