
Abstract
Introduction
C
Pluripotent stem cells (PSCs) from early embryos (called embryonic stem cells [ESCs]) (82) are capable of becoming all types of body cells under specific conditions both in vivo and in vitro. Thus, they hold great therapeutic potential for repairing nonregenerative organs, including the heart. However, establishing ESCs involves destroying early embryos (blastocysts), and this has continued to raise ethical problems. Furthermore, ESCs can form teratomas and are subject to immune rejection (99). The work of Yamanaka and colleagues in 2006 provided a partial solution to these problems by generating induced PSCs (iPSCs) from adult somatic cells (127). Since iPSCs can be generated from a patient's own somatic cells such as skin or blood cells, they circumvent ethical and patient-specificity problems associated with ESCs and, thus, constitute an attractive source of cells for therapeutic use. Research has also been directed toward the use of cardiac stem cells (CSCs) and cardiac progenitor cells (CPCs) and the development of a direct reprogramming method for the generation of CMs from somatic cells.
While the cell sources and methods described earlier have increased the hope for heart regeneration, there are critical hurdles to be overcome before putting them into practice. In this review, we will summarize currently available cell sources (Fig. 1) and methods for heart regeneration and discuss their potential and limitations.

Embryonic Cell Source
ESC-derived CMs
ESCs can be considered an ideal source for replacing dysfunctional CMs, as their pluripotency and ability to self-renew allow them unlimited expansion and to differentiate into specialized cell types, such as CMs, under appropriate conditions. ESCs were first isolated from mouse blastocysts (82) and, subsequently, from other species, including rat (46), rabbit (36), monkey (131), and human (130). The pluripotency of ESCs is mediated via common transcription factors, including Oct3/4, Sox2, and Nanog (5, 14, 18, 95), but the maintenance of rodent and human ESCs (hESCs) depends on different signaling cascades [i.e., leukemia inhibitory factor or fibroblast growth factor (FGF) signals for rodent or hESCs, respectively (1, 96)], suggesting different requirements for their self-renewal despite a common transcriptional network for pluripotency.
The cardiogenic potential of ESCs was well characterized by studies of mouse (26) and human (56) ESCs. Similar to adult CMs, hESC-derived CMs express the cardiac transcription factors GATA4 and Nkx2.5, as well as the cardiac-specific sarcomeric genes cardiac troponin I, cardiac troponin T (cTnT), atrial myosin light chain, ventricular myosin light chain (MLC-2v), and α-myosin heavy chain (αMHC) (56). Furthermore, the hESC-derived CMs express the CM gap junction proteins connexin (Cx)-43 and Cx45 and have action potentials similar to those of human ventricular myocytes (57). This suggests that ESC-derived CMs may be molecularly and functionally similar to CMs in vivo.
The transforming growth factor β, Wnt, FGF, and bone morphogenetic protein (BMP) signaling pathways play critical roles in establishing the cardiovascular system (30, 59, 66, 68, 83, 113, 114, 122, 139), and this knowledge has been applied for improving the efficiency of differentiating hESCs into CMs. The simplest way to obtain CMs from ESCs is to spontaneously differentiate ESCs by generating embryoid bodies (EBs), spherical cell aggregates formed via self-aggregation of ESCs (56). However, the efficiency of generating CMs is very low with the EB method. In 2005, Mummery and colleagues developed a coculture method to increase the differentiation rate of hESCs, based on the in vivo observation that the anterior endoderm is crucial for heart formation. They used a visceral endoderm-like cell line, END-2, as feeders for hESCs, and, to some extent, this enhanced cardiac differentiation of hESCs (102). Later, direct coaggregation of ESCs and END-2 in suspension was found to greatly promote CM formation (136). The effect was mediated by fibronectin secreted from END-2 cells, affecting Wnt signaling in ESCs (21). Subsequently, Keller and colleagues demonstrated that temporally mimicking an early in vivo environment with a defined set of growth factors, including activin A, BMP4, FGFs, vascular endothelial growth factor (VEGF), and Dickkopf-1, is sufficient to change the fate of ESCs to precardiac mesoderm and can be used to generate CMs from ESCs with a high efficiency (51, 53). However, this method requires substantial optimization due to ESC line variations. Recent studies showed that sequential promotion and inhibition of Wnt signaling result in a high yield of CMs from hESCs in a robust manner (77), and that PSC-derived CMs can be expanded by small molecules (138).
To use ESC-derived CMs to improve heart function, it is necessary for them to interact properly with endogenous CMs and function normally after transplantation. Initial in vivo trials demonstrated that hESC-derived CMs are capable of forming new myocardium when transplanted into the hearts of rats (70) and pigs (57). The engrafted CMs expressed cardiac markers, including αMHC, MLC-2v, and atrial natriuretic factor (ANF), and the size of the CM graft significantly increased over time, implying their proliferation in vivo. Moreover, electrophysiological mapping and histopathological examination revealed that hESC-derived CMs could form pacemakers and couple electrically with surrounding host CMs in pig (70) and guinea pig (149) hearts. When these cells were transplanted into the infarcted heart in rats, they appeared to improve the contractile function of the heart (69). The engrafted CMs were also electrically coupled to the host myocardium and improved the function of the postinfarcted heart (61, 89, 118). These studies suggest that hESC-derived CMs have characteristics similar to adult CMs and may be a suitable cell source to repair the damaged heart. However, despite the numerous advantages, there are several impediments to be overcome for therapeutic use. For instance, ESC-derived CMs show structural and functional properties of early-stage CMs instead of those of mature adult CMs (55) and remain immature after engraftment. In addition, there are safety concerns, such as tumor formation and immune rejection (99), and ethical issues on the isolation of ESCs from human blastocysts.
iPSC-derived CMs
In 2006, Yamanaka and colleagues developed a novel method to induce ESC-like cells—referred to as iPSC—from mouse fibroblasts by introducing a defined set of transcription factors (i.e., Oct3/4, Sox2, and c-Myc) using viral vectors (127). Similar to ESCs, the iPSCs were capable of differentiating into derivatives of all three germ layers both in vivo and in vitro and of forming teratomas when transplanted into nude mice. The next year, human iPSCs (hiPSCs) were generated with the same combination of transcription factors (126) or a different set of factors (i.e., Oct3/4, Sox2, Nanog, and Lin-28) (152). These factors were originally delivered via viral methods with retroviruses or lentiviruses, and, therefore, their random integration creates insertional mutations. Furthermore, incomplete silencing of c-Myc, a proto-oncogene, can lead to tumor formation (42). To resolve these issues, alternative methods have been developed with the LoxP/Cre system (119), Piggy Bac system (146), episomal plasmids (151), nonintegrating viruses (115), and siRNA (90). While the LoxP/Cre and Piggy Bac methods still leave small insertions, the others offer ways to generate integration-free iPSCs.
The differentiation potential of iPSCs into CMs has been extensively studied with iPSCs derived from mice (87), pigs (91), and humans (155). In comparison to hESCs, hiPSCs showed a lower efficiency of differentiation. However, both hiPSC- and hESC-derived CMs displayed similar patterns of cardiac gene expression, including Nkx2.5, cTnT (troponin T type 2 [TNNT2]), αMHC, α-actinin, myosin light chain 2 atrial isoform, myosin light chain 2 ventricular isoform, ANF, and phospholamban (155), with spontaneous rhythmic intracellular Ca2+ fluctuations (87). Moreover, iPSC-derived CMs contained atrial- and ventricular-like cells and responded to β-adrenergic signaling, a canonical CM signaling pathway (155). This suggests that iPSC-derived CMs are molecularly and functionally similar to ESC-derived CMs.
The clinical potential of iPSCs in cardiac repair was demonstrated in animal models. Okano and colleagues generated cardiac cell sheets with hiPSC-derived CMs in two-dimensional culture (86). These sheets showed spontaneous and synchronous beating, even after being detached from the culture dishes, and extracellular action potential propagation when two sheets were partially overlaid. hiPSC-derived cardiac cell patches can also be generated using a three-dimensional (3D) culture system (135). In this system, hiPSC-derived CMs were cultured in collagen type I to generate contractile cardiac tissue patches, and their proliferation, hypertrophy, and alignment were controlled by mechanical stress. When these patches were transplanted into the adult rat heart, they formed grafts with contractile function (135). The hiPSC-derived CM sheets also improved cardiac function and left ventricular remodeling in porcine myocardial infarction (MI) models (54).
Given that iPSCs share numerous features in common with ESCs but circumvent the ethical and allogeneic transplantation problems associated with ESCs (126, 152), they are considered a suitable alternative to ESCs. However, genome-wide analysis of iPSCs and ESCs has revealed significant differences in gene expression, methylation, and microRNA (miRNAs) expression patterns (23, 145). The gene expression patterns of iPSCs also depended on their origins (33, 81). Moreover, Xu and colleagues showed that ESCs derived from a C57BL/6 mouse could induce teratoma formation without immune rejection, whereas some of the iPSCs derived from the same mouse resulted in T-cell-dependent immune rejection and failed to form teratomas. It turned out that the immunological difference between ESCs and iPSCs was caused by the abnormal expression of antigens in iPSCs (158). This indicates that iPSCs could have immunogenic problems, which may be linked to mutations that occur before, during, or after reprogramming (35). Thus, it will be necessary to evaluate genetic and epigenetic modifications and immunogenicity of iPSCs before clinical applications.
Cardiac progenitor cells
While the potential of CSCs remain to be further clarified, the biology of CPCs has been extensively studied both in vivo and in vitro. CPCs are identified in developing embryos and used as building blocks to generate the heart during embryogenesis (Fig. 2). The basic helix-loop-helix transcription factor mesoderm posterior 1 (Mesp1) is transiently expressed in the primitive streak during gastrulation and marks early CPCs (110, 111). Mesp1+ CPCs migrate anteriorly and form the first heart field (FHF) and second heart field (SHF) (17, 52, 92, 140). FHF cells give rise to the atria and left ventricle, whereas the outflow tract, right ventricle, and some of the atria are derived from the SHF. The heart fields express late CPC markers, such as Islet1 (Isl1), fetal liver kinase 1 (Flk1), and Nkx2.5, which are often used to identify and isolate CPCs from ESCs/iPSCs (44, 52, 92, 147). Similar to CPCs found in vivo, ESC/iPSC-derived CPCs have the capability of expanding and differentiating into various types of cardiac cells, including CMs, smooth muscle cells, and endothelial cells in vitro (52, 64, 67, 92, 147). With this capability, CPCs are thought to have tremendous regenerative potential to repair damaged hearts. However, although they can be expanded by Wnt, Notch, and Fgf signals (25, 64, 65), CPCs are heterogeneous cells that spontaneously differentiate, and no method is available to clonally expand them in a renewal state. This poses a concern for potential PSC carryovers that can lead to tumor formation. A recent study showed that a subset of CPCs can undergo self-renewal as a homogeneous population in a dedicated cellular environment (117). Determining their environmental factors may enable long-term maintenance and mass production of CPCs. When engrafted into infarcted heart, ESC-derived CPCs differentiated into adult-sized CMs with Cx43 expression, reconstitute ∼20% of the scar tissue in a nonhuman primate model (12). Consistently, unlike PSC-derived CMs, PSC-derived CPCs appear to differentiate into mature CMs and vascular cells in the adult mouse heart without causing teratomas or arrhythmias (24, 98). While these studies revealed the unique potential of CPCs to form adult-like CMs for heart regeneration, further investigation will be required to determine their morphological and functional maturation, coupling, and functional integration with host myocardium, and long-term survival and effects on heart function.

Adult Cell Source
Cardiac stem cells
The adult heart is considered a terminally differentiated organ that lacks regenerative capacity. In fact, nearly all CMs withdraw from the cell cycle shortly after birth in mammals (79, 109). However, the human heart contains cycling myocytes under both normal and pathological conditions (9, 94, 106), and by measuring the concentration of 14C in the DNA, Frisen and colleagues proposed that human CMs renew at the rate of 1% per year and 45% of the CMs are exchanged during an individual's life span (11). These studies suggested the existence of endogenous cells contributing to new myocytes in the adult heart. In fact, several groups have identified CSC populations in the adult heart, although their origins still remain unclear.
The tyrosine kinase receptor c-kit was originally identified as a marker of hematopoietic stem cells (48) and later shown to mark CSCs (3). Cardiac c-kit+ cells were isolated from adult heart tissue and expressed cardiac transcription factors, such as Gata4, Gata5, MEF2C, and Nkx2.5. These cells were named CSCs, as they could self-renew and give rise to CMs, smooth muscle cells, and endothelial cells (8) after differentiation (7, 8). When injected into an ischemic heart, c-kit+ CSCs differentiated into myocardium, vessels, and myocytes and reduced the infarct size with functional improvement (8), and c-kit+ CSCs are necessary and sufficient for cardiac regeneration and repair (29). Stem cell antigen-1 (Sca-1), a cell-surface marker for somatic and hematopoietic stem cells, was also shown to mark resident heart cells with stem cell properties (43, 85). Sca-1+/CD31− cells were identified in the adult myocardium as small interstitial cells and expressed cardiac transcription factors, such as Gata4 and MEF2C. Similar to c-kit+ CSCs, transplantation of Sca-1+/CD31− cells after MI improved heart function by promoting angiogenesis and CM regeneration (144). Sca-1-deficient mice exacerbated cardiac hypertrophy with disorganized CMs and increased cell apoptosis under pressure overload, suggesting that Sca-1 may also have an essential role in cardiac remodeling and integrity (108). In addition to c-kit+ or Sca-1+ CSCs, cells expressing ATP-binding cassette transporter ABCG2/breast cancer resistance protein 1 (BCRP) were identified as a CSC/CPC marker and shown to play a pivotal role in cardiac repair post-MI (40, 159). However, the existence and potential of CSCs remain controversial due to reproducibility issues (2, 153), and recent studies indicated that the benefits associated with adult stem cell injection might come from paracrine effects rather than from their myocardial differentiation (32). While cytokines and growth factors affecting angiogenesis and cell survival and proliferation, including VEGFs, BMPs, FGFs, and Wnt modulators, were proposed as the potential paracrine mediators responsible for cardiac protection and functional improvement (34, 75), further studies will be necessary to confirm their beneficial effects and mechanisms.
Cardiac fibroblasts
Although PSC-derived CMs/CPCs have demonstrated their therapeutic potential for cell-mediated heart regeneration, there are still concerns about carry-over undifferentiated PSCs that can form tumors. In addition, a large numbers of CMs/CPCs are required to replenish cells lost due to heart damage. Direct in vivo reprogramming may resolve these concerns, as the method enables generating CMs in situ, directly from endogenous non-CM cells. Ieda and colleagues developed the protocol for direct reprogramming of cardiac fibroblasts by introducing the transcription factors Gata4, Mef2c, and Tbx5( GMT cocktail) in mice (47). This method greatly reduced the steps required to produce CMs while limiting the formation of teratomas. Introducing these factors reprogrammed cardiac fibroblasts into CM-like cells (induced CMs [iCMs]) in vitro but they were morphologically and molecularly similar to neonatal CMs (47). However, intramyocardial delivery of the factors into infarcted hearts, whose fibroblasts were genetically labeled, resulted in conversion of cardiac fibroblasts into mature iCMs (105), suggesting that environmental factors, such as extracellular matrix (ECM), secreted factors, and contracting neighboring cells, may play an important role in their maturation. The GMT-treated mice showed significant improvement in heart function, particularly in cardiac output and stroke volume for approximately 3 months postintervention (105). Several other groups reported that the reprogramming efficiency can be enhanced by adding one more factor to the original GMT cocktail (120), fusing the MyoD transactivation domain to the transcription factors (41), or using miRNAs (miRNAs 1, 133, 208 and 499) instead of the cocktail (49).
With the abundance of cardiac fibroblasts and their in situ reprogramming, this method has great therapeutic potential to repair the damaged heart. Further investigation will be required to improve the reprogramming efficiency and to minimize the formation of incompletely reprogrammed or immature iCMs, as abnormal CMs can result in deleterious effects in patients, such as arrhythmias. In addition, it will be necessary to establish a robust protocol to generate iCMs in a highly reproducible manner (20). It is worth noting that given the low reprogramming efficiency, newly generated iCMs appear to be not sufficient to explain the high levels of functional improvement achieved by introducing the factors into the infarcted heart (120). Thus, it will be also of importance to investigate additional roles and mechanisms of the reprogramming factors leading to the functional improvement.
Existing CMs
While CMs in primitive animals, such as zebrafish, are capable of dedifferentiating and proliferating to repair damaged heart muscle (50), CMs in mammals are considered terminally differentiated and unable to re-enter the cell cycle. However, there is growing evidence that mammals retain the regenerative capacity at their neonatal stage (104) and the Hippo signaling pathway—known to control organ growth in Drosophila (101)—plays a key role in CM proliferation in the neonatal heart (141). Hippo inactivation, which leads to activation of its effector molecule Yes-associated protein 1 (Yap1), resulted in heart enlargement with increased CM proliferation during development (39). Similarly, genetic activation or deletion of Yap1 causes myocardial hyperplasia or hypoplasia, respectively, in the postnatal heart as well as in the prenatal heart, but CM size appeared to be unaffected (141). More recently, cardiac-specific deletion of Yap1 was shown to impair neonatal heart regeneration and increased levels of an active form of Yap1 in the adult heart promoted CM proliferation and dramatically enhanced heart regeneration after MI (148). These studies suggest that Hippo/Yap1 signaling is a crucial regulator of CM proliferation during embryonic and neonatal heart development and can push adult CMs to re-enter the cell cycle. However, it is unknown whether CM proliferation in the adult heart requires dedifferentiation of CMs. Interestingly, increased levels of oncostatin M (OSM), an interleukin-6 (IL-6) class of cytokine, were shown to induce dedifferentiation of adult CMs, evidenced by loss of sarcomeric structures and upregulation of fetal genes and stem cell markers, and to enhance their cell-cycle progression (62). Thus, it appears that adult CMs are capable to undergo dedifferentiation and proliferation, and elucidating the precise mechanisms of the events may facilitate the development of an alternative way to regenerate the heart.
Cell Purification
For embryonic or adult cell-based heart regeneration, it is crucial to isolate specific precardiac or cardiac cell types without genetic manipulation and contamination, and efforts have been expended to identify cell-surface proteins (Fig. 2), which enables cell purification via antibody-mediated fluorescence-activated cell sorting (FACS).
PSC-derived CPCs can be isolated using cell-surface markers, such as KDR and/or PDGFRA (53, 150). Purified KDR/PDGFR+ cells express high levels of Mesp1 (21) and can differentiate into all types of cardiac cells, including CMs, endothelial cells, and smooth muscle cells (51, 53). FHF progenitors, which represent committed CPCs, can be isolated with the cell-surface protein hyperpolarization-activated, cyclic nucleotide-gated K 4 (HCN4) and FACS-purified HCN4+ cells preferentially differentiate into myocardial lineages (121). However, despite extensive in vivo characterization of the SHF, cell-surface molecules specifically expressed in SHF progenitors remain unclear. This may reflect the heterogeneity of SHF CPCs.
Signal-regulatory protein alpha (SIRPA) and vascular cell adhesion molecule 1 (VCAM-1) are cell-surface proteins highly expressed in PSC-derived CMs, and antibodies interacting with their extracellular domains can be used to isolate the CMs. FACS-purified SIRPA+ or VCAM-1+ cells were shown to contain 98% or 95%, respectively, of cTnT+ CMs derived from PSCs (28, 137). Different approaches were also proposed to purify PSC-derived CMs. Tetramethylrhodamine methyl ester perchlorate (TMRM) is a fluorescent dye that specifically labels mitochondria, and CMs can be distinguished from non-CMs by the intensity of TMRM (38). This method was shown to enrich PSC-derived CMs of approximately 99% purity (38). PSC-derived CMs can also be purified based on their metabolic properties to survive in a glucose-depleted and lactate-enriched condition (133). This method does not involve FACS, and therefore can be suitable for isolating a large quantity of CMs. While the described methods may enable us to efficiently purify CMs from PSC culture, CMs are heterogeneous, and it is unclear which chamber CMs are generated and isolated from PSCs. Determining the molecular and functional properties of CMs in vivo may enable us to identify and purify homogeneous or specific CMs from PSCs.
Most of the adult CSC/CPC markers, such as c-kit, Sca-1, and ABCG2/BCRP, are cell-surface proteins, and their antibodies are directly used for isolation (8, 85, 159). However, these proteins are also expressed in many other cell types, and the self-renewal and differentiation potential of the isolated cells remain to be confirmed.
Unlike adult cell sources, CPCs/CMs isolated from PSC culture have concerns of tumorigenicity due to potential contamination of residual undifferentiated PSCs. A recent study suggested a way to selectively remove undifferentiated PSCs: They inhibited the activity of anti-apoptotic proteins (Bcl10 and survivin) enriched in PSCs with small-molecule inhibitors (e.g., quercetin or YM155) (73). The inhibitors did not seem to affect the survival and function of differentiated cell types, such as dopamine neurons and smooth muscle cells (73). However, it will be necessary to determine whether the inhibitors do not affect cardiac differentiation and CM viability and function, because survivin deficiency causes premature cardiac death and CM proliferation defects (74).
Cell Delivery and Survival
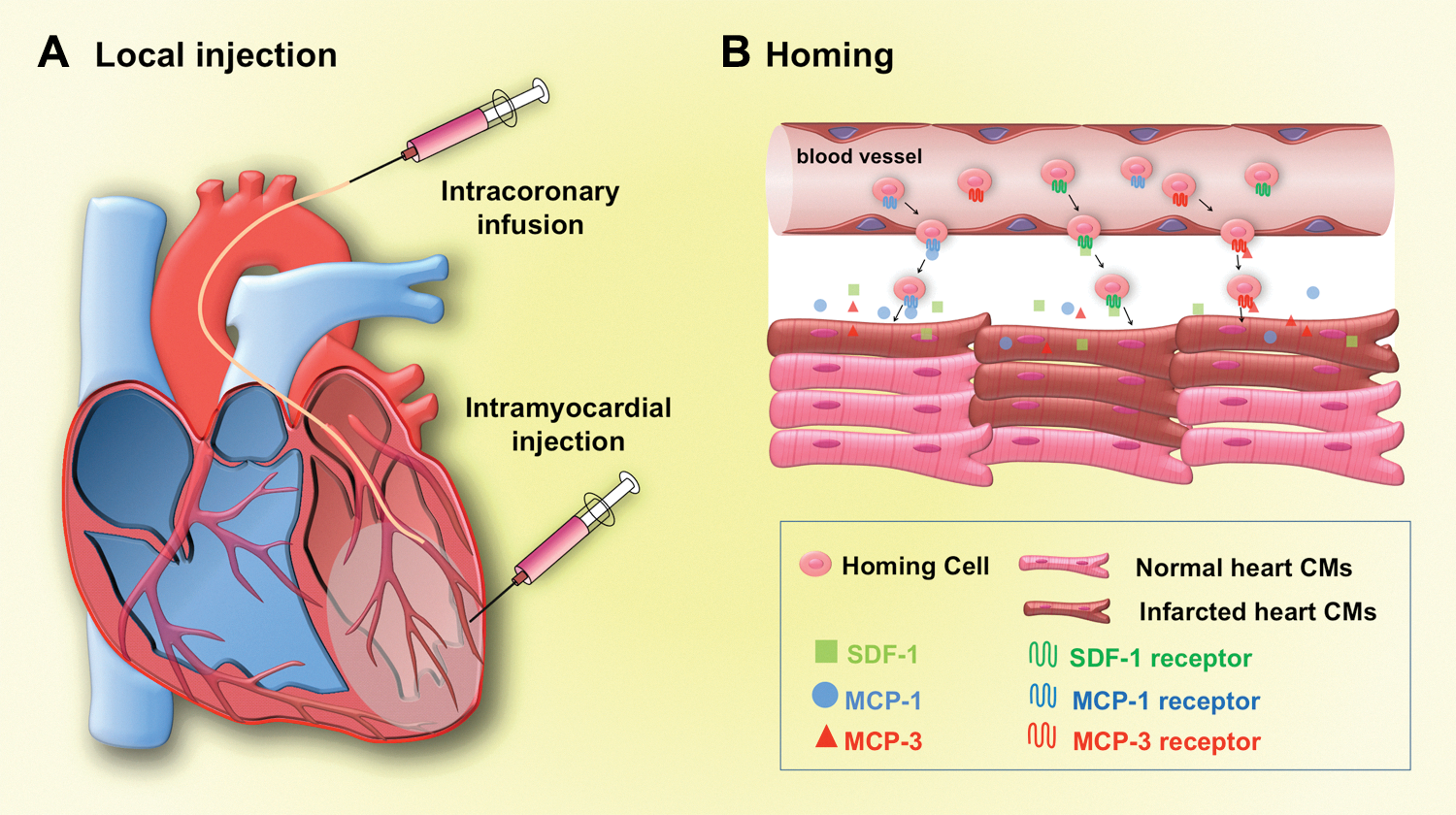
To treat the damaged heart via cell transplantation, it is important to develop cell delivery strategies and protocols that enable precise and efficient introduction of cells into the site of injury. Direct and indirect methods are currently used for cell delivery (Fig. 3). Direct intramyocardial injection is a common way to deliver cells into the injured region of the heart (103). In this method, cells can be injected directly into infarcted or scar regions of ventricular wall. Although this method has the advantage of direct visualization and delivery with proven safety (103), needle punctures can cause mechanical leakage and lead to massive loss of cells injected (129). Intracoronary infusion is another option to deliver cells into the myocardial region of injury. While this approach can minimize cell leakage and may limit their systemic diffusion, injected cells may not be retained well (16) and can cause acute myocardial ischemia and myocardial microinfarction (143). Cells may be delivered via a homing system, which enables endogenous or exogenous cells to migrate to a specific region. Homing systems generally depend on chemo-attractants expressed in injured tissues, and cell delivery is often done systemically. On MI, expression levels of chemo-attractants, such as stromal-cell-derived factor 1 (SDF-1) (4), monocyte chemotactic protein (MCP)-1 (128), and MCP-3 (112), were shown to increase in the heart and this could lead to cell homing to the damaged tissue. In fact, increased levels of SDF-1 and the SDF-1 receptor CXCR4 in mesenchymal stem cells (MSCs) enhanced the homing efficiency and improved cardiac function (4, 22, 154, 156). Similarly local overexpression of MCP-3 was shown to improve cardiac function by recruiting MSCs to the injured cardiac tissue (112). Homing systems can also be activated by growth factors, such as stem cell factor, VEGF, neural growth factor, granulocyte-colony stimulating factor, insulin-like growth factor, hepatocyte growth factor, and FGF [for review, see Taghavi and George (125)]. However, it is unknown whether a homing system can be used to deliver PSC-derived CPCs/CMs, and its clinical efficacy remains to be determined.
After cell delivery, the majority of injected cells are often washed out, and the engrafted cells do not survive long enough in the host heart. Thus, current cardiac tissue engineering focuses on developing methods to improve cell retention and survival of grafted cells (Fig. 4). Endogenous CMs are tightly attached to ECM molecules, such as laminin, collagen, fibronectin, and entactin, that typically provide structural support and play crucial roles in cell growth, wound healing, and fibrosis (71). For this reason, much effort has been put into the development of matrices that mimic native ECM. For instance, synthetic hydrogels share physical and biomechanical properties to native ECM and have widely been used as a 3D scaffold in tissue engineering (31). Intra-myocardial injection of hydrogel-encapsulated CSCs supplemented with ECM binding proteins significantly improved viability, proliferation, and long-term retention of the CPCs after MI (88). However, progressive loss of engrafted cells over time still needs to be resolved (88). Collagen is a natural ECM component and has been frequently used for tissue grafts, because it offers many advantages for cell transplantation such as low immunogenicity and biodegradability (13, 72). Coinjection with collagen matrices was shown to significantly enhance the survival rate of engrafted ESC-derived CMs and contribute to functional improvement of ischemic rat hearts (63). Similarly, the survival rate of hESC-derived CMs was significantly increased when they were delivered with matrigel (gelatinous ECM mixture from mouse sarcoma cells) into the infarcted heart of athymic rats (69). Addition of AVAD-fmk (antiapototic agent), cyclosporine A (immunosuppressant), pinacidil (ATP-dependent K+ channel opener), and insulin-like growth factor 1 (described as prosurvival cocktail together with matrigel) further enhanced their survival (69).

Other prosurvival cocktail have been developed. miRs significantly improved CPC engraftment without ECM in a murine infarct model (45). The cocktail included miR-21, miR-24, and miR-221 that bind to the 3′ UTR region of Bim, an apoptotic activator, and suppress its expression (45). These prosurvival cocktail studies suggest that cell death caused by apoptosis is an important factor affecting the cell engraftment. Apoptotic signaling in transplanted cells can also be inhibited by heat shock protein 70, whose expression is induced by heat shock (93) or by activation of the serine-threonine kinase Akt that targets the apoptotic molecules Bcl-2 and caspases (58). Heatshock treatment improved graft size of cardiac-enriched hESC progeny about threefold in rat hearts (70), and Akt overexpression reduced the percentage of engrafted neonatal CM undergoing apoptosis by 40% (157). While these studies point to the importance of blocking apoptotic pathways for cell survival in host myocardium, it remains to be determined whether the proposed methods have long-term effects.
While the approaches described earlier sought to develop cell-surviving methods through cell-autonomous mechanisms, it is also important to understand the cellular environment of damaged myocardium as it directly influences cell engraftment. For example, oxidative stress—caused by the imbalance between reactive oxygen species (ROS) levels and antioxidant defense—is increased in the failing heart (134) and can cause heart failure (97).
ROS are chemically reactive molecules, such as oxygen ions and peroxides, and are naturally generated by normal oxygen metabolism. Since high ROS levels increase oxidative stress, their levels are usually kept low by antioxidant molecules, such as vitamins C and E, superoxide dismutase (SOD), and glutathione peroxidase (132). Injured heart tissues are exposed to high ROS levels, and this condition adversely affects the survival and function of endogenous CMs (19, 142). Thus, high levels of ROS could negatively affect the viability of injected cells in the damaged heart. Indeed, SOD treatment increased the survival rate of engrafted skeletal myoblasts in the heart (124), and increasing antioxidant levels in the ischemic heart enhanced survival and proliferation of CSCs engrafted to the myocardium (37). The beneficial effect was also observed when injected cells were pretreated with N-acetylcysteine that increases antioxidant levels (27). Since mitochondria are the major source of ROS (15), ROS levels might be more efficiently controlled by manipulating thioredoxin reductase-2, a key regulator of the overall emission of peroxides from cardiac mitochondria (123).
Although these studies demonstrated the importance of altering oxidative environment and the potential of antioxidants for survival of grafted cells, high-dose antioxidants were shown to negatively affect genetic stability, which may predispose to cancer (76). Similarly, increased levels of the angiogenic factors VEGF (107) and hypoxia inducible factor-1α (6) could improve survival of grafted cells by enhancing blood vessel formation, and their overexpression is closely associated with tumor development (10, 116). Thus, it will be critical to determine safe dose ranges of antioxidants or angiogenic factors when they are used for cell transplantation therapies.
Outlook
Stem cells have enormous potential for treating heart failure. Efforts are now underway around the world to improve the current strategies and methodologies for heart regeneration, which may be applied for clinical interventions in the near future. As discussed, methods, including stem cell differentiation, purification, maturation, delivery, survival, and retention, should be thoroughly tested and optimized for stem cells to be used for clinical applications. New approaches may be needed to overcome obstacles, such as those that have been encountered in clinical trials.
In addition to cell-based therapies, the potential of stem cells is being actively investigated with multidisciplinary approaches. There are ongoing trials to generate bioartifical hearts in vitro by recellularizing a decellularized heart with CMs (100) or CPCs derived from PSCs (78). A novel in vivo method is under development for generating an entire organ by introducing donor iPSCs into host blastocysts of different species that are incapable of forming the organ (60). A therapeutic method could be sought from another direction such as 3D printers that have now been applied to generate organs using a special nozzle which can provide cells and a matrix of a specific shape (80). As such, integration of new technologies with stem cells is expected to facilitate and accelerate the development of innovative stem-cell therapies for heart regeneration, which may lead to cures for heart failure in the future.
Footnotes
Acknowledgments
The authors thank Kwon laboratory members for helpful discussions. This work was supported by grants from NHLBI/NIH (R01HL111198, R00HL092234), MSCRF, and the Magic That Matters Fund.